Embed Size (px)
Citation preview

INFECTION AND IMMUNITY,0019-9567/01/$04.00�0 DOI: 10.1128/IAI.69.11.6931–6941.2001
Nov. 2001, p. 6931–6941 Vol. 69, No. 11
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Cloning of the Streptococcus mutans Gene Encoding GlucanBinding Protein B and Analysis of Genetic Diversity and
Protein Production in Clinical IsolatesRENATA O. MATTOS-GRANER,1 SONG JIN,1 WILLIAM F. KING,1 TSUTE CHEN,2
DANIEL J. SMITH,1 AND MARGARET J. DUNCAN2*
Department of Immunology1 and Department of Molecular Genetics,2
The Forsyth Institute, Boston, Massachusetts 02115
Received 6 June 2001/Returned for modification 23 July 2001/Accepted 22 August 2001
Streptococcus mutans, the primary etiological agent of dental caries, produces several activities that promoteits accumulation within the dental biofilm. These include glucosyltransferases, their glucan products, andproteins that bind glucan. At least three glucan binding proteins have been identified, and GbpB, the proteincharacterized in this study, appears to be novel. The gbpB gene was cloned and the predicted protein sequencecontained several unusual features and shared extensive homology with a putative peptidoglycan hydrolasefrom group B streptococcus. Examination of gbpB genes from clinical isolates of S. mutans revealed that DNApolymorphisms, and hence amino acid changes, were limited to the central region of the gene, suggestingfunctional conservation within the amino and carboxy termini of the protein. The GbpB produced by clinicalisolates and laboratory strains showed various distributions between cells and culture medium, and amountsof protein produced by individual strains correlated positively with their ability to grow as biofilms in an invitro assay.
Dental caries is a transmissible infectious disease in whichmutans streptococci (MS) play the major role. Streptococcusmutans, the primary etiological agent, possesses several viru-lence factors that allow it to accumulate within the dentalbiofilm and to produce and tolerate the acids that cause cari-ous lesions. Biofilm development occurs in two distinct phases;during the first, bacterial surface proteins interact with host orbacterial products adsorbed on the tooth surface. In the secondphase, a biofilm forms as bacteria accumulate by aggregationwith the same or other species and produce an extracellularpolysaccharide matrix (13). Bacterial components associatedwith the accumulation phase of MS include glucosyltrans-ferases, their glucan products, and proteins that bind glucan.At least three S. mutans glucan binding proteins (Gbp) havebeen identified: GbpA (23), GbpB (26), and GbpC (24). Aftercloning and sequencing, the gbpA gene product was found toshare homology with the putative glucan binding domain ofglucosyltransferase (2) and the gbpA gene was found to encodea constitutively expressed secreted protein (1, 2). Cell surface-associated GbpC was related to the Spa family of streptococcalproteins and was expressed only during conditions of stress(24). GbpB was immunologically distinct from other Gbpsexpressed by S. mutans and Streptococcus sobrinus and alsodiffered in size and purification properties (26).
For successful colonization of the oral cavity, MS need non-shedding tooth surfaces; thus, the period of highest coloniza-tion occurs from approximately 18 to 30 months of age, coin-ciding with the eruption of primary molars (3, 12). At this age,
the mucosal immune system is mature enough to respond toimmune interventions against infecting MS (27) and MS col-onization of young children results in a measurable immuno-globulin A response to GbpB (27). Furthermore, in experi-mental infection of rats, systemic or mucosal immunizationwith GbpB induced protective immunity to dental caries, indi-cating that GbpB may be an important target for the develop-ment of caries vaccines (29). Preliminary studies have shownthat GbpB is expressed in all laboratory and clinical S. mutansstrains tested so far (D. J. Smith, W. F. King, and M. A.Taubman, J. Dent. Res. 74:123, 1995). However, the biologicalfunction of GbpB and its role in the virulence of S. mutans arestill unclear. The cloning and sequencing of gbpB were brieflyreported previously (S. Jin, M. J. Duncan, M. A. Taubman, andD. J. Smith, J. Dent. Res. 79:224, 2000). The protein showshomology to a putative peptidoglycan hydrolase from group Bstreptococcus, suggesting that GbpB plays a role in peptidogly-can biosynthesis. In this study, we examined the genotypic andprotein diversity of gbpB in clinical isolates of S. mutans. Wealso determined the production and localization of GbpB pro-tein in clinical isolates and laboratory strains and amounts ofprotein produced correlated positively with biofilm growth inan in vitro assay.
MATERIALS AND METHODS
Bacterial strains and growth conditions. S. mutans strain SJ32 was describedpreviously (26, 27). Additional S. mutans strains were obtained from a subset ofchildren derived from a previously described larger population (16, 17). Clinicalstrains were genotyped by arbitrarily primed PCR (15) and included 44 distinctS. mutans amplitypes and an S. sobrinus strain. Laboratory strains of S. mutansstrains included in this study were UA130, UA159, T8 (kindly provided by P. W.Caufield, University of Alabama), and GS5 (kindly provided by H. K. Kuramitsu,State University of New York at Buffalo). MS strains were grown in either ToddHewitt broth (THB) or brain heart infusion broth or plates and in chemicallydefined medium (COM) (31) under anaerobic conditions, 10% H2–10% CO2–
* Corresponding author. Mailing address: Department of MolecularGenetics, The Forsyth Institute, 140 Fenway, Boston, MA 02115.Phone: (617) 262-5200, ext. 344. Fax: (617) 262-4021. E-mail: [email protected].
6931
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

80% N2. Escherichia coli strains DH5� and BL21 (Novagen, Madison, Wis.) weregrown in Luria Bertani (LB) broth or plates, and ampicillin (100 �g/ml) was usedto select and maintain recombinant plasmids. Unless stated otherwise, all chem-icals were obtained from Sigma Chemical Co., St. Louis, Mo.
DNA isolation. S. mutans chromosomal DNA was isolated using a MasterPureDNA purification kit from Epicentre Technologies (Madison, Wis.). Recombi-nant plasmid DNA was isolated using a Perfectprep kit from Eppendorf Scien-tific Inc. (Westbury, N.Y.).
S. mutans library construction. Chromosomal DNA was partially digested withSau3AI, and 2.5- to 5.0-kb fragments were gel purified using a QIAquick gelextraction kit (QIAGEN Inc., Valencia, Calif.). Fragments were ligated toBamHI-digested, calf intestinal phosphatase-treated pUC19 and used to trans-form competent E. coli DH5� cells. Ampicillin-resistant recombinant cloneswere selected on plates of LB and ampicillin and containing X-Gal (5-bromo-4-chloro-3-indolyl-�-D-galactopyranoside) and IPTG (isopropyl-�-D-thiogalacto-pyranoside) to screen for insert-containing clones.
Southern hybridization. Hybridization conditions and signal developmentwere as recommended in the Enhanced Chemiluminescence gene detectionsystem (Amersham-Pharmacia, Piscataway, N.J.).
Cloning of gbpB. GbpB was purified by anion exchange chromatography in thepresence of urea as previously described (28). Peptide sequences were obtainedat the Molecular Biology Core Facilities, Dana-Farber Cancer Institute, Boston,Mass., and the Harvard Microchemistry Facility, Harvard University, Cambridge,Mass., by trypsin digestion, high-performance liquid chromatography, matrix-assisted laser desorption ionization–time of flight (MALDI-TOF) (mass spec-trometry), and Edman degradation. PCR primers derived from these peptidesand used to isolate gbpB are described in Table 1. Reaction mixture volumeswere 50 to 100 �l and contained 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 2.5 mMMgCl2, 50 �M deoxynucleoside triphosphates, approximately 0.5 �M primers,0.1 to 0.5 �g of template, and 2.5 U of AmpliTaq Gold DNA polymerase(Perkin-Elmer, Branchburg, N.J.). Thermal conditions were as follows: 10 min at95°C; 6 cycles of 30 s at 95°C, 30 s at 40°C, and 1.5 min at 72°C with 5-sincrements per cycle; 30 cycles of 30 s at 95°C, 30 s at 46°C, and 2 min at 72°Cwith 5-s increments per cycle; and 5 min at 72°C. PCR and specific sequencingprimers were obtained from The Midland Certified Reagent Company (Midland,Tex). A Peltier Thermal Cycler model PTC-200 (MJ Research, Cambridge,Mass.) was used for PCR. Sequencing was carried out with either dRhodamineor Big Dye Terminator cycle sequencing kits (Perkin-Elmer, Foster City, Calif.)
using a PE 9700 Thermocycler. Reactions were run on Perkin-Elmer ABI 377Sequencer.
Expression of gbpB in E. coli. The gbpB gene from S. mutans SJ32 genomicDNA was amplified using a 5� PCR primer with an NcoI restriction enzymerecognition site for insertion of the amplicon into an expression vector and forsequence coding for the first seven amino acids of the mature GbpB protein. The3� primer contained an XhoI restriction site and sequence coding for the lasteight amino acids of GbpB, but excluding the stop codon. The 1,212-kb PCRproduct was ligated to the NcoI-XhoI-digested vector pET22B and then trans-formed into E. coli BL21(DE3). GbpB expression in E. coli was induced withIPTG, protein extraction, and purification of the His-tag protein on nickelcolumns as described by the supplier (Novagen).
RFLP analysis of gbpB. gbpB genotypes were detected by restriction fragmentlength polymorphism (RFLP) analysis of PCR-amplified gbpB genes from 44amplitypes of S. mutans clinical isolates. Laboratory strains SJ32, T8, UA130,UA159, and GS5 were also included in the analysis. The gbpB DNA sequencefrom SJ32 was aligned with that of strain UA159 (S. mutans genome database[http://www.genome.ou.edu/smutans.html]), and from the latter, primers weredesigned to amplify gbpB plus 162 bp of the upstream and 195 bp of thedownstream sequences flanking the open reading frame (ORF) of GbpB (Table1).
Measurement of GbpB in culture supernatants and cell extracts. After over-night growth in CDM, bacterial suspensions were adjusted to the same opticaldensity (A550) so that the same size inoculum was added to fresh CDM (4 ml)containing 10 �M hydrochloride 4-(2-aminoethyl)-benzolsulfonylfluoride(AEBSF) from Roche Diagnostics (Indianapolis, Ind.) to inhibit protease activ-ity. After growth for 18 h, cells were pelleted from 2 ml of culture, and thesupernatant was filtered through 0.22-�m-pore-size Spin-X filter membranes(Costar, New York, N.Y.) and immediately frozen at �70°C. Cell-associatedGbpB was extracted with urea as previously described (8). For direct compari-sons, culture supernatants and urea extracts were dialyzed overnight at 4°Cagainst 0.02 M sodium phosphate buffer (PB) (pH 6.5) with 5 mM �-mercapto-ethanol. All samples were stored at �70°C.
GbpB in culture supernatants and cell extracts was measured by immuno dotblotting. Nitrocellulose membranes (Bio-Rad, Foster City, Calif.) were washedwith PB and applied to a dot blot apparatus (Bio-Rad). Two hundred microlitersof PB was applied to each well and dried under vacuum, followed by 50 �l ofsample diluted 1:2 in PB, and was allowed to drain. An additional 200 �l of PB
TABLE 1. PCR primers used in this study
Procedure 5� Primer 3� Primer a Comment b Ampliconsize (bp)
PCRStep 1 (Fig. 1A) GAT/C GAT/C TTT/C
TGT/C GCI CAA/GATT/CA GC
TGT/C TGI CCI AA/GIGTI GCI GAT/C TGIGC
Derived from N terminus and internalpeptide of mature protein
180
Step 2 (Fig. 1B) TAA ATA AAA ATGTAA TAA TG
AGT TGT GCT GCT TCTAAT
Derived from GenBank acc.no. 4098503
691
Step 3 (Fig. 1D) TGC AGC TAA TCAGGA GAC AA
GA TAA ATA TAA CTGACA CT
5� primer, derived from GenBank acc.no. 4098503; 3� primer, from S.mutans database
765
Step 4 (1st round)(Fig. 1E)
CAA GCA GCA GCCGCA CAA
TAA TAC GAC TCA CTATAG GG N NNN NNNNNN GAT ATC
5� primer, derived from 765-bpamplicon; 3� primer, arbitraryprimer (4)
Step 4 (2ndround)(Fig. 1E)
GCC GCA GCT GATAAC AC
ATT ATG CTG AGT GATATC CC
5� primer, derived from 765-bpamplicon; 3� primer, arbitraryprimer (4)
1,054
Step 5 (Fig. 1F) ATA TAC TGG CTT TTCTCT TCT TTC
ACT TTC TCT GCT AATTCT TTG TTT
5� primer, derived from 100 bpupstream of gbpB promoter; 3�primer, derived from 100 bpdownsteam of gbpB terminationcodon
1,644
RFLP analysis(Fig. 5A)
ATA TAC TGG GCT TTTCTC TTC TTT C
GTC TTA GCC ACT TTCTCT GC
Derived from UA159 sequenceflanking ORF of GbpB (S. mutansgenome database [http://www.genome.ou.edu/smutans.html])
1,653
a I is inosine; N is A, T, G, or C.b acc, accession.
6932 MATTOS-GRANER ET AL. INFECT. IMMUN.
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

was applied per well and dried under vacuum. Samples of serially diluted purifiedGbpB were included as standards. Membranes were blocked for 1.5 h in amixture of 100 mM Tris-HCl, 2.5 mM NaCl (pH 7.5), 10% skim milk, and 0.2%NaN3, followed by incubation with rat polyclonal antiserum (26) to GbpB (1:700dilution) for 2 h at room temperature. As negative controls, samples wereincubated with preimmune antibody. Following incubation with primary or pre-immune antibody, membranes were washed three times for 5 min with 100 mMTris buffer containing 0.25% Tween (TBST), pH 7.5. Incubation with the sec-ondary antibody, fluorescein-conjugated rabbit anti-rat immunoglobulin G(Zymed, San Francisco, Calif.) was at room temperature for 2 h in the dark.Membranes were washed twice with TBST, then three times with 100 mM Trisbuffer, pH 7.5. Signals were detected with a fluorescence reader (Storm 840;Molecular Dynamics, Sunnyvale, Calif.) and converted to digital images, and dotblot intensities were quantified with ImageQuant software (Molecular Dynam-ics). Samples of all strains were blotted to the same membrane for comparison,and the immunoassay was repeated at least twice. The amounts of GbpB inculture fluids and cell extracts were quantified by comparison with serial dilutionsof purified GbpB of known concentration within a linear range. The samesamples were also tested as described above for amounts of GbpA and totalglucosyltransferases using polyclonal antibodies against these proteins at serumprotein concentrations of 1:200 and 1:700, respectively.
Biofilm formation in microtiter plates. Biofilm formation in microtiter plateswas assayed by the method described by O’Toole and Kolter (21). An aliquotfrom a 5-ml overnight THB culture was diluted 1:100 in fresh THB, and 200 mlwas transferred to sterile polystyrene U-bottom microtiter plates (Dynatech Lab,Chantilly, Va.). Plates were incubated anaerobically for 18 h, and biofilm growthwas revealed and quantified by staining with crystal violet (21). Crystal violetabsorbance was determined with a plate reader at 575 nm (Dynatech, Winooski,Vt.). The absorbance (A550) of planktonic cultures grown under the same con-ditions was measured to monitor growth. Biofilm formation for all strains wasmeasured in triplicate plates. The laboratory strains S. mutans UA130 and SJ32were also included in the same experiments.
Statistical analysis. Pearson correlation analyses were used to evaluate asso-ciations between production of GbpB, biofilm growth, and planktonic growth ofcultures. To avoid bias because of GbpB production by different S. mutansamplitypes, only one of each amplitype identified per child was used after ran-dom selection.
Nucleotide sequence accession numbers. The nucleotide sequence of the gbpBgene from S. mutans strain SJ32 was assigned GenBank accession numberAY046410; from strain 3VF4, AY046411; from 15JP2, AY046412; from 3SN1,AY046413; and from 5SM3, AY046414.
RESULTS
Cloning of gbpB. Cloning was carried out during the begin-ning stages of the S. mutans genome project, and before gbpB-related sequences were available. Therefore, mature GbpBwas purified from culture supernatants of S. mutans SJ32 andthe N-terminal and two internal peptide sequences were de-termined after tryptic digestion, MALDI-TOF (mass spec-trometry), and cycle sequencing. The N-terminal sequences ofpeptide 1 (DDF?AQIASCD[A]KI[V][N][T]) and internal pep-tides 2 (GWFNPGSVSYIYP[L]) and 3 (LEAQSATLGQQIQFIG. 1. Strategy for cloning gbpB from S. mutans SJ32. (A) From S.
mutans SJ32 template DNA, PCR primers 1 and 3 yielded a 180-bpfragment with 100% DNA and amino acid identity to GenBank acces-sion (Acc.) no. 4098503, a partially cloned but unknown S. mutansgene. Peptides 1 and 3 were found in the partial putative ORF; thus,peptide order within GbpB was 1-3-2. N-term, N terminus. (B)Specific 5� and 3� primers designed from accession no. 4098503 gen-erated a 691-bp amplicon. (C) The purified amplicon was used toprobe an S. mutans SJ32 genomic library for clones containing gbpB-related sequences. A single hybridizing clone contained a 3.5-kb insertwith the 5� promoter region, a 300-amino-acid (aa) ORF comprising asignal sequence, the N terminus of mature GbpB, peptide 3, and 89amino acids more than the gene fragment reported in GenBank ac-cession no. 4098503. The ORF did not contain peptide 2 or a stopcodon. (D) To obtain the 3� region, the PCR primer specific to the 5�sequence of GenBank accession no. 4098503 was used with a 3� specificprimer designed after BLAST searching the S. mutans genome data-base with peptide 2. A 765-bp amplicon was aligned with alreadyobtained sequences, but still without the stop codon. (E) A randomly
primed PCR strategy (4) was used to generate the C terminus of theprotein and the 3� noncoding region. First-round PCR products weregenerated with specific primer 1 derived from the 765-bp amplicon andrandom primer 2 that contained a 3� EcoRV sequence. The secondround 5� specific primer 3 was again derived from sequences within the765-bp amplicon, and the 3� random primer 4 retained the EcoRVsequence tag. A 1,054-bp fragment was generated that contained the Cterminus, the stop codon, and 3� downstream untranslated sequence.(F) All sequences were aligned, and PCR primers were designed forthe sequence 100 bp up- and downstream of the promoter and stopcodons to generate the complete gene from SJ32. (G) GbpB expressedin E. coli reacted with antibody to the native protein. Shown is aWestern blot of GbpB expressed in E. coli. Lane 1, induced GbpB; lane2, induced vector alone; lane 3, purified GbpB.
VOL. 69, 2001 GENETIC AND PROTEIN ANALYSIS OF GbpB FROM S. MUTANS 6933
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

TLSSSK) were obtained. However, the positions of 2 and 3relative to each other were unknown, since at that time therewere no homologues in GenBank, nor did they have identitywith sequences released from the genome sequencing project.Degenerate oligonucleotide primers 1 and 3 (Table 1) weredesigned from peptides 1 and 3, respectively (Fig. 1A), sincepeptide 2 was unsuitable for primer design. Southern blotanalysis showed that primer 1, designed from N-terminal pep-tide 1, hybridized to single fragments of restriction enzyme-digested S. mutans SJ32 chromosomal DNA (data not shown),indicating that at least the 5� terminus of the gene existed insingle copy in the genome. The steps involved in cloning gbpBare depicted in Fig. 1. The gbpB gene was subcloned into thepET22B expression vector and transformed into E. coli BL23.Following induction with IPTG, cell-associated protein ex-tracts and culture supernatants contained proteins that reactedwith polyclonal antibody to purified GbpB (Fig. 1G), providingevidence that we had cloned the correct protein.
DNA and protein analysis of GbpB. The DNA sequence ofgbpB together with the predicted coding sequence is shown inFig. 2. A putative Pribnow box, T97AATATA103, was foundnext to the predicted transcription start site at A103 (PromoterPrediction by Neural Network [http://www.hgc.lbl.gov.gov
/projects/promoter.html]) (10; M. G. Reese, N. L. Harris, andF. H. Eeckman, Proc. 1996 Pacific Symp. Biocomputing, 1996),and a ribosome binding site (147AGGA150) was identified 12 bp5� to the ATG start codon. The coding sequence was 1,295 bpin length, and following the TAA termination codon, a hairpinloop was detected (1468A-1499T) with �40.3 kcal of free energyper ml. Therefore, it appears that gbpB is an independentlyregulated gene.
The open reading frame encoded a protein of 431 aminoacids with a molecular mass of 44.58 kDa. A 27-amino-acidsignal peptide was predicted by SignalP (20), and the cleavagesite confirmed the N-terminal sequence of the purified maturesecreted protein. Thus, the mature form of the polypeptide isca. 41.3 kDa, approximating the size estimated by mass spec-trometry. The protein was glutamine and alanine rich, 13.9 and21.4%, respectively, by composition. There was a leucine zip-per motif, LQTQQAELQAENQRLEAQSATLGQQIQTL,within the N-terminal half of the protein. GbpB did not possessthe C-terminal cell wall anchor motif LPXTG (18), consistentwith it being found in culture fluids as well as cell associated;however, the C-terminal region contained hydrophobic se-quences (11) also found in cell wall targeted or secreted pro-teins in gram-positive bacteria (6).
FIG. 2. DNA and encoded protein sequence of gbpB from S. mutans SJ32. The putative Pribnow box in the 5� untranslated sequence isunderlined, and the predicted transcription start is shown in bold. The ribosome binding site is shown in bold and underlined. Within the proteinsequence the leucine zipper is underlined.
6934 MATTOS-GRANER ET AL. INFECT. IMMUN.
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

Apart from the leucine zipper sequence, there were noamino acid motifs within GbpB that suggested putative biolog-ical function. However, nine stress-responsive genes were re-cently identified in S. mutans by differential display reversetranscription PCR (5). One of these, GSP-781, was activated byacid stress, high osmolarity, and temperature and shared 100%amino acid identity with GbpB. Further insight into a potentialfunction for GbpB was provided by the high homologythroughout its length (60% complete and 69% positive iden-tities) to PcsB, a recently identified secreted protein in group B
streptococcus (22) (Fig. 3). Both PcsB and GbpB show somehomology to P45, a protein from Listeria monocytogenes thatwas identified as a peptidoglycan hydrolase (25). These en-zymes play important roles in peptidoglycan cycling, cell wallsynthesis, and daughter cell separation. Although hydrolyticactivity could not be demonstrated with purified PcsB, electronmicrographs showed that pcsB knockout mutant cells werelarger than wild-type cells, with abnormal cell shape and divi-sion septa, suggesting that PcsB played a role in cell wallsynthesis or division (22).
FIG. 3. Alignment of deduced amino acid sequences of GbpB and PcsB precursor proteins. The ClustalW multiple alignment program(http://npsa-pbil.ibcp.fr/cgi-bin/align_clustalw.pl) was used to align amino acids. Identical and similar amino acids are marked at the bottom asasterisks (strong similarity) and dots (weak similarity), respectively. A conserved cysteine residue at the C-terminal region is observed at position350.
VOL. 69, 2001 GENETIC AND PROTEIN ANALYSIS OF GbpB FROM S. MUTANS 6935
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

Comparative genomic analysis of gbpB region. Evidencefrom comparative genomics also suggests that GbpB is in-volved in cell wall biology. Analysis of the S. mutans genomedatabase (http://www.genome.ou.edu) revealed that two genesupstream from and in the same orientation as gbpB, encodedhomologues of cell shape-determining proteins from othergram-positive bacteria. These ORFs contained approximately50% amino acid similarity to cell shape-determining proteinsMreC and MreD from Streptococcus pneumoniae, Lactococcuslactis, Enterococcus faecalis, and Enterococcus faecium (Fig. 4).Downstream from gbpB, other genes were identified that en-coded homologues of proteins from gram-positive organisms(L. lactis, Bacillus subtilis, L. monocytogenes, Streptococus gor-donii, and Streptococcus sanguis) that were involved in aminoacid and lipid synthesis, ATP-dependent transport system,DNA repair, and competence. The gene immediately down-stream from pcsB in group B streptococci encoded a polypep-tide with high similarity to phosphoribosyl pyrophosphate syn-thetase (23), and the same locus was found immediatelydownstream from gbpB. Thus, in several gram-positive bacte-ria, including S. mutans, there appears to be conservation ofgenomic structure suggesting a functional relationship betweengenes involved in cell shape and cell wall maintenance.
RFLP analysis of the gbpB gene. The gbpB gene was ob-tained by PCR from 44 amplitypes of S. mutans strains (15),and a fragment of the predicted size, 1,653 kb, was obtainedfrom all strains (data not shown). Figure 5A shows the pre-dicted restriction map of gbpB, and endonucleases PvuII andSau3AI were selected as the most informative for RFLP anal-ysis because of the higher number of cutting sites and yield offragments that could be well separated by electrophoresis. ByPvuII digestion, the gbpB genes could be divided into threeclasses, and the most prevalent class, represented by strain
5SM3 in Fig. 5B, was found in 63.7% of the strains, includingSJ32, from which gbpB was originally sequenced, as well aslaboratory strains T8 and UA130. Also, 34% of the amplitypesshowed a second pattern, represented by strain 3VF4 (Fig. 5B)and strains UA159 and GS5. Sau3AI-generated polymor-phisms were less informative since only four amplitypes (9.1%)
FIG. 4. Comparative genomics of the S. mutans gbpB locus. Genes flanking gbpB were examined for homologies in gram-positive bacteria.ORFs 1 and 2 showed homology to MreC and MreD, described as cell shape-determining proteins. GbpB shares homology with Usp45 from S.pneumoniae and L. lactis and to SagA from E. faecalis and E. faecium, which are secreted proteins related to peptidoglycan hydrolases fromgram-positive bacteria. ORFs 4 and 5 were variable with regard to protein function. Accession numbers were obtained from the GenBank and TheInstitute for Genomic Research (�) databases.
FIG. 5. RFLP analysis of gbpB from clinical isolates of S. mutans.(A) Restriction map of 1,653-bp amplicon that included 162 bp up-stream and 195 bp downstream flanking ORF of GbpB from UA159.(B) RFLP patterns with restriction enzyme PvuII. (C) Restrictionpatterns obtained with Sau3AI. Lanes M, molecular size markers.
6936 MATTOS-GRANER ET AL. INFECT. IMMUN.
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

showed a different pattern, as shown with strain 3SN1 in Fig.5C.
A representative from each of the PvuII and Sau3AI classeswas sequenced to identify base changes. A total of 18 changeswere found, of which 17 were point mutations. One class,represented by 15JP2 (Fig. 6), contained a point mutation inthe DNA sequence encoding the signal peptide that resulted ina conservative residue change (valine to isoleucine) three res-idues upstream from the signal peptidase cleavage site. Thestrain still secreted GbpB (see below), so it appears unlikelythat the mutation affects processing. Another class, repre-sented by strains 3SN1 and 15JP2, had a 3-bp insertion thatcoded for an extra asparagine residue at position 264. This andthe remaining point mutations occurred within a relativelydefined region (456 to 972 bp) corresponding to the centralregion of the protein (Fig. 5). Eight mutations were silent,another four resulted in conservative amino acid changes, andfive represented changes to amino acids of different classes.
Production of GbpB by clinical isolates of S. mutans. Theamounts of GbpB measured in culture supernatants from 76clinical isolates were found to be highly variable. Values werenormalized to microgram of protein/A550 of culture, and themean (� standard deviation [SD]) production level was 5.58 �0.167 �g/ml of culture supernatant. The A550 of cultures didnot vary significantly (mean, 0.147; SD, 0.116); thus, there wasno correlation between growth and GbpB production (Pear-
son, r � 0.05, P � 0.667). Strain 20A3 produced the largestamounts of GbpB in the culture supernatant (8.52 �g/ml), andstrain 8VS3 produced the smallest amounts (1.16 �g/ml).Twenty isolates with the widest spectrum of GbpB proteinlevels, including S. mutans laboratory strains UA130 and SJ32and S. sobrinus as a negative control, were selected for furtheranalysis of the distribution of GbpB between cells and super-natants (Fig. 7). It then became apparent that levels of GbpBin culture supernatants alone did not reflect total levels since insome strains a large amount of protein remained cell associ-ated, e.g., strain 8VS3. On the other hand, 20A3 containedlittle cell-associated protein and other strains produced smallamounts of GbpB in both culture supernatants and cell ex-tracts, e.g., 4JP2. Out of 19 S. mutans clinical strains analyzed,eight (42.1%) retained most of the GbpB they produced in acell-associated form, e.g., 3A1 and 5SM3, while in 11 strains(57.9%), most of the protein was secreted into the culturesupernatants. Among the laboratory strains, most of the GbpBproduced by SJ32 was found in culture supernatants, while forUA130, most of the protein was in a cell-associated form. Asexpected, S. sobrinus strain 3SSA1 did not produce GbpB. Thepatterns of GbpB distribution between cell surface and culturesupernatants were reproducible, and in similar control exper-iments, GbpA was extracted from 20 strains and the majorityof this protein was in supernatants and was not cell associated(data not shown). Thus, the protein localization patterns ap-
FIG. 6. Polymorphisms of gbpB in different S. mutans strains lead to changes in protein sequence. Out of 18 mutations identified, 9 causedchanges in the protein level (highlighted bars) and 5 represented changes to residues of different classes.
VOL. 69, 2001 GENETIC AND PROTEIN ANALYSIS OF GbpB FROM S. MUTANS 6937
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
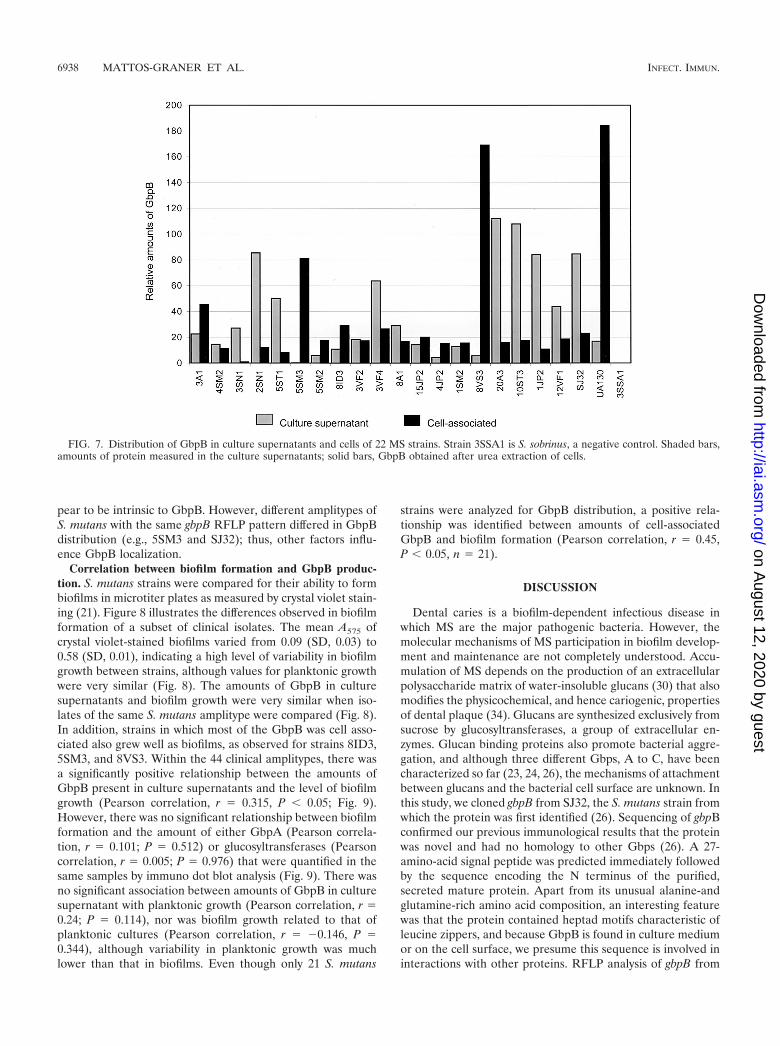
pear to be intrinsic to GbpB. However, different amplitypes ofS. mutans with the same gbpB RFLP pattern differed in GbpBdistribution (e.g., 5SM3 and SJ32); thus, other factors influ-ence GbpB localization.
Correlation between biofilm formation and GbpB produc-tion. S. mutans strains were compared for their ability to formbiofilms in microtiter plates as measured by crystal violet stain-ing (21). Figure 8 illustrates the differences observed in biofilmformation of a subset of clinical isolates. The mean A575 ofcrystal violet-stained biofilms varied from 0.09 (SD, 0.03) to0.58 (SD, 0.01), indicating a high level of variability in biofilmgrowth between strains, although values for planktonic growthwere very similar (Fig. 8). The amounts of GbpB in culturesupernatants and biofilm growth were very similar when iso-lates of the same S. mutans amplitype were compared (Fig. 8).In addition, strains in which most of the GbpB was cell asso-ciated also grew well as biofilms, as observed for strains 8ID3,5SM3, and 8VS3. Within the 44 clinical amplitypes, there wasa significantly positive relationship between the amounts ofGbpB present in culture supernatants and the level of biofilmgrowth (Pearson correlation, r � 0.315, P 0.05; Fig. 9).However, there was no significant relationship between biofilmformation and the amount of either GbpA (Pearson correla-tion, r � 0.101; P � 0.512) or glucosyltransferases (Pearsoncorrelation, r � 0.005; P � 0.976) that were quantified in thesame samples by immuno dot blot analysis (Fig. 9). There wasno significant association between amounts of GbpB in culturesupernatant with planktonic growth (Pearson correlation, r �0.24; P � 0.114), nor was biofilm growth related to that ofplanktonic cultures (Pearson correlation, r � �0.146, P �0.344), although variability in planktonic growth was muchlower than that in biofilms. Even though only 21 S. mutans
strains were analyzed for GbpB distribution, a positive rela-tionship was identified between amounts of cell-associatedGbpB and biofilm formation (Pearson correlation, r � 0.45,P 0.05, n � 21).
DISCUSSION
Dental caries is a biofilm-dependent infectious disease inwhich MS are the major pathogenic bacteria. However, themolecular mechanisms of MS participation in biofilm develop-ment and maintenance are not completely understood. Accu-mulation of MS depends on the production of an extracellularpolysaccharide matrix of water-insoluble glucans (30) that alsomodifies the physicochemical, and hence cariogenic, propertiesof dental plaque (34). Glucans are synthesized exclusively fromsucrose by glucosyltransferases, a group of extracellular en-zymes. Glucan binding proteins also promote bacterial aggre-gation, and although three different Gbps, A to C, have beencharacterized so far (23, 24, 26), the mechanisms of attachmentbetween glucans and the bacterial cell surface are unknown. Inthis study, we cloned gbpB from SJ32, the S. mutans strain fromwhich the protein was first identified (26). Sequencing of gbpBconfirmed our previous immunological results that the proteinwas novel and had no homology to other Gbps (26). A 27-amino-acid signal peptide was predicted immediately followedby the sequence encoding the N terminus of the purified,secreted mature protein. Apart from its unusual alanine-andglutamine-rich amino acid composition, an interesting featurewas that the protein contained heptad motifs characteristic ofleucine zippers, and because GbpB is found in culture mediumor on the cell surface, we presume this sequence is involved ininteractions with other proteins. RFLP analysis of gbpB from
FIG. 7. Distribution of GbpB in culture supernatants and cells of 22 MS strains. Strain 3SSA1 is S. sobrinus, a negative control. Shaded bars,amounts of protein measured in the culture supernatants; solid bars, GbpB obtained after urea extraction of cells.
6938 MATTOS-GRANER ET AL. INFECT. IMMUN.
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

44 S. mutans amplitypes revealed that most sequence changeswere clustered in the central region of the protein, suggestingfunctional conservation of sequences within the N- and C-terminal domains. Recent results indicate that the N terminusis immunologically active (unpublished data) and may possessthe protective epitopes associated with the native molecule(29).
Although sequences with partial homology to GbpB werefound in the GenBank database, there were no suggestions asto function, apart from the ability to bind glucan. Recently,a differential display reverse transcription-PCR screen for S.mutans stress-responsive genes identified a gene with completeidentity to gbpB (5). Expression increased under conditions ofhigh osmolarity and temperature, and it was speculated thatthe encoded protein might be a structural component of thecell wall or membrane. More significantly with regard to GbpBfunction, a protein designated PcsB was recently identified ingroup B streptococcus (22) and was shown to have good ho-
mology to GbpB (Fig. 3). Generating a pcsB mutant proveddifficult, and by using osmotically protective conditions forboth transformation and the subsequent growth of candidates,one mutant was obtained. In addition to osmotic sensitivity, themutant grew slowly, was hypersensitive to several antibiotics,and showed abnormal septum formation during growth. Fromthese characteristics, together with some homology to a pep-tidoglycan hydrolase from L. monocytogenes, it was concludedthat PcsB was involved in wall separation during cell division.Database analyses indicated that PcsB-like proteins werepresent in several gram-positive bacteria; these proteins in-cluded P45 of L. monocytogenes, SagA from E. faecium and E.faecalis, and Usp45 from Streptococcus pneumoniae and L.lactis (7, 32, 33). GbpB homologues are preceded by two ORFsthat code for cell shape-determining proteins (Fig. 4). As withGbpB and PcsB, these proteins were both secreted and cellassociated and contained a conserved cysteine residue in theC-terminal domain that was necessary for peptidoglycan hy-
FIG. 8. Biofilm formation, planktonic growth, and GbpB production in clinical strains of S. mutans. Strains within vertical lines were isolatedfrom the same child. One child (10ST) was colonized by two different amplitypes: one represented by isolates 10ST1 and 10ST4 and the other byisolates 10ST2 and 10ST3. In all other samples, one amplitype per child was identified. The lower panels show the amounts of GbpB present inculture supernatants that were quantified by immuno dot blot analysis.
VOL. 69, 2001 GENETIC AND PROTEIN ANALYSIS OF GbpB FROM S. MUTANS 6939
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

drolytic activity (35). We observed polymorphic forms of GbpBin clinical isolates of S. mutans; however, most of the changeswere clustered in the central region of the protein (Fig. 6)while the N- and C-terminal regions were conserved, suggest-ing functional roles. Because of the close homology of GbpB toPcsB, and in turn to the other proteins with sequence similar-ities that include the conserved cysteine active site residue, wehypothesize that GbpB has a similar function in peptidoglycanbiosynthesis. To test this hypothesis, we attempted to isolategbpB mutants in several strains using different integrating con-structs. Inactivation of gbpB by simple Campbell-like integra-tion of a suicide vector yielded viable mutants containing mul-tiple and deleted versions of gbpB, as indicated by PCR andsequencing analysis (R. O. Mattos-Graner et al., unpublisheddata). Scanning electron micrographs of one class of slow-growing mutants showed clumps of enlarged cells instead ofthe chains of cells observed with the parent strain, a growthphenotype similar to that of pcsB mutants (22). Thus, althoughglucan-binding properties were originally ascribed to GbpB,our results to date indicate that the protein plays other roles inthe biology of S. mutans.
GbpB was previously described as a secreted protein inseveral laboratory strains of S. mutans (D. J. Smith et al., J.Dent. Res), however, we observed large variations in theamounts of GbpB secreted in the culture supernatants of 74clinical isolates of S. mutans. In the majority of clinical isolates,most of the GbpB was secreted into culture medium, but insome amplitypes the majority of the protein remained cellassociated (Fig. 7). Unlike GbpC, GbpB does not possess theC-terminal cell wall anchor motif, LPXTG, found in severalsurface proteins of gram-positive bacteria (18). However, thiscell-sorting signal is not contained in all surface proteins ofgram-positive organisms (19) and was not present in P45 fromL. monocytogenes (25).
The amounts of GbpB secreted by strains of the same am-plitype were very similar, suggesting that distribution of theprotein between cells and culture medium is intrinsic to spe-cific clones of S. mutans. Different genotypes, even when iso-lated from the same host, showed different amounts of se-creted GbpB, as exemplified by strains 10ST1 and 10ST4 (Fig.8), which have the same amplitype and secrete large amountsof GbpB, and strains 10ST2 and 10ST3, which were isolatedfrom the same child but have a different DNA fingerprint andsecrete less GbpB. Since GbpB may be involved in cell wallformation, it would be interesting to determine the response ofeach amplitype to high osmolarity, temperature, pH, and an-tibiotic exposure.
In this study, a large number of S. mutans clinical isolateswere examined for the ability to form biofilms in low-sucrosemedium (THB) (9) using an in vitro assay that was previouslyused to screen for biofilm-defective mutants in both gram-negative and gram-positive bacteria (14, 21). Despite high vari-ability between isolates, a significant finding was a positivecorrelation between the ability of strains to grow as biofilmsand GbpB production. Even strains in which a large proportionof the protein remained cell associated showed good growth inbiofilms. That no correlations were observed between produc-tion of GbpA or glucosyltransferases and biofilm formationalso underscores the importance of GbpB in this mode ofgrowth (Fig. 9). However, for several strains, the level of se-
FIG. 9. (A) A positive relationship was detected between amountsof GbpB present in culture supernatants and biofilm formation amongthe 44 amplitypes of S. mutans. No significant associations were de-tected between amounts of GpbA (B) and glucosyl transferases (GTF)(C) and biofilm growth in the same strains.
6940 MATTOS-GRANER ET AL. INFECT. IMMUN.
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

creted GbpB did not correlate with biofilm growth, a predict-able finding since many unknown factors, beside GbpB, areinvolved in the complex process of biofilm formation. Since thevirulence of S. mutans is directly related to its ability to colo-nize tooth surfaces and accumulate in the dental biofilm, it isimportant to define a function for GbpB and its role in biofilmformation. Given the extent of homology between GbpB andPscB from group B streptococcus, one hypothesis is that GbpBis involved in cell wall formation. Several biofilm-defectivemutants of S. gordonii Challis had disruptions in genes involvedin peptidoglycan biosynthesis, and a putative osmoregulatoryrole in biofilm formation was suggested (14). This providescircumstantial supporting evidence for our hypothesis thatGbpB is also involved in peptidoglycan biosynthesis.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grant R37 DE-06153 (NIDCR) to D.J.S. and grant 99/08278-9 (FAPESP) to R.O.M.-G.
REFERENCES
1. Banas, J. A., H. C. Potvin, and R. N. Singh. 1997. The regulation of Strep-tococcus mutans glucan-binding protein A expression. FEMS Microbiol.Lett. 154:289–292.
2. Banas, J. A., R. R. Russell, and J. J. Ferretti. 1990. Sequence analysis of thegene for the glucan-binding protein of Streptococcus mutans Ingbritt. Infect.Immun. 58:667–673.
3. Caufield, P. W., G. R. Cutter, and A. P. Dasanayake. 1993. Initial acquisitionof mutans streptococci by infants: evidence for a discrete window of infec-tivity. J. Dent. Res. 72:37–45.
4. Chen, T. R., R. Yong, H. Dong, and M. J. Duncan. 1999. A general methodfor direct sequencing of transposon mutants by randomly primed PCR,T01834. Technical Tips Online T01834. [Online.] http://research.bmn.com/tto.
5. Chia, J. S., Y. Y. Lee, P. T. Huang, and J. Y. Chen. 2001. Identification ofstress-responsive genes in Streptococcus mutans by differential display re-verse transcription-PCR. Infect. Immun. 69:2493–2501.
6. Davis, N. G., and P. Model. 1985. An artificial anchor domain: hydropho-bicity suffices to stop transfer. Cell 41:607–614.
7. Furst, P., H. U. Mosch, and M. Solioz. 1989. A protein of unusual compo-sition from Enterococcus faecium. Nucleic Acids Res. 17:6724.
8. Hamada, S., T. Horikoshi, T. Minami, N. Okahashi, and T. Koga. 1989.Purification and characterization of cell-associated glucosyltransferase syn-thesizing water-insoluble glucan from serotype c Streptococcus mutans.J. Gen. Microbiol. 135(Part 2):335–344.
9. Hamada, S., and M. Torii. 1978. Effect of sucrose in culture media on thelocation of glucosyltransferase of Streptococcus mutans and cell adherence toglass surfaces. Infect. Immun. 20:592–599.
10. Harley, C. B., and R. P. Reynolds. 1987. Analysis of E. coli promoter se-quences. Nucleic Acids Res. 15:2343–2361.
11. Hopp, T. P., and K. R. Woods. 1981. Prediction of protein antigenic deter-minants from amino acid sequences. Proc. Natl. Acad. Sci. USA 78:3824–3828.
12. Kohler, B., D. Bratthall, and B. Krasse. 1983. Preventive measures in moth-ers influence the establishment of the bacterium Streptococcus mutans intheir infants. Arch. Oral Biol. 28:225–231.
13. Kolenbrander, P. E. 2000. Oral microbial communities: biofilms, interac-tions, and genetic systems. Annu. Rev. Microbiol. 54:413–437.
14. Loo, C. Y., D. A. Corliss, and N. Ganeshkumar. 2000. Streptococcus gordoniibiofilm formation: identification of genes that code for biofilm phenotypes.J. Bacteriol. 182:1374–1382.
15. Mattos-Graner, R. O., Y. Li, P. W. Caufield, M. Duncan, and D. J. Smith.2001. Genotypic diversity of mutans streptococci in Brazilian nursery chil-
dern suggests horizontal transmission. J. Clin. Microbiol. 39:2313–2316.16. Mattos-Graner, R. O., D. J. Smith, W. F. King, and M. P. Mayer. 2000.
Water-insoluble glucan synthesis by mutans streptococcal strains correlateswith caries incidence in 12- to 30-month-old children. J. Dent. Res. 79:1371–1377.
17. Mattos-Graner, R. O., F. Zelante, R. C. Line, and M. P. Mayer. 1998.Association between caries prevalence and clinical, microbiological and di-etary variables in 1.0 to 2.5-year-old Brazilian children. Caries Res. 32:319–323.
18. Navarre, W. W., and O. Schneewind. 1994. Proteolytic cleavage and cell wallanchoring at the LPXTG motif of surface proteins in gram-positive bacteria.Mol. Microbiol. 14:115–121.
19. Navarre, W. W., and O. Schneewind. 1999. Surface proteins of gram-positivebacteria and mechanisms of their targeting to the cell wall envelope. Micro-biol. Mol. Biol. Rev. 63:174–229.
20. Nielsen, H., J. Engelbrecht, S. Brunak, and G. von Heijne. 1997. Identifica-tion of prokaryotic and eukaryotic signal peptides and prediction of theircleavage sites. Protein Eng. 10:1–6.
21. O’Toole, G. A., and R. Kolter. 1998. Initiation of biofilm formation inPseudomonas fluorescens WCS365 proceeds via multiple, convergent signal-ling pathways: a genetic analysis. Mol. Microbiol. 28:449–461.
22. Reinscheid, D. J., B. Gottschalk, A. Schubert, B. J. Eikmanns, and G. S.Chhatwal. 2001. Identification and molecular analysis of PcsB, a proteinrequired for cell wall separation of group B streptococcus. J. Bacteriol.183:1175–1183.
23. Russell, R. R. 1979. Glucan-binding proteins of Streptococcus mutans sero-type c. J. Gen. Microbiol. 112:197–201.
24. Sato, Y., Y. Yamamoto, and H. Kizaki. 1997. Cloning and sequence analysisof the gbpC gene encoding a novel glucan-binding protein of Streptococcusmutans. Infect. Immun. 65:668–675.
25. Schubert, K., A. M. Bichlmaier, E. Mager, K. Wolff, G. Ruhland, and F.Fiedler. 2000. P45, an extracellular 45 kDa protein of Listeria monocytogeneswith similarity to protein p60 and exhibiting peptidoglycan lytic activity.Arch. Microbiol. 173:21–28.
26. Smith, D. J., H. Akita, W. F. King, and M. A. Taubman. 1994. Purificationand antigenicity of a novel glucan-binding protein of Streptococcus mutans.Infect. Immun. 62:2545–2552.
27. Smith, D. J., W. F. King, H. Akita, and M. A. Taubman. 1998. Association ofsalivary immunoglobulin A antibody and initial mutans streptococcal infec-tion. Oral Microbiol. Immunol. 13:278–285.
28. Smith, D. J., W. F. King, and R. Godiska. 2001. Passive transfer of immu-noglobulin Y antibody to Streptococcus mutans glucan binding protein B canconfer protection against experimental dental caries. Infect. Immun. 69:3135–3142.
29. Smith, D. J., and M. A. Taubman. 1996. Experimental immunization of ratswith a Streptococcus mutans 59-kilodalton glucan-binding protein protectsagainst dental caries. Infect. Immun. 64:3069–3073.
30. Smith, D. J., and M. A. Taubman. 1997. Vaccines against dental cariesinfection, p. 913–930. In M. Levine, G. C. Woodrow, J. B. Kraper, and G. S.Cobon (ed.), New generation of vaccines. Marcel Dekker, New York, N.Y.
31. Socransky, S. S., J. L. Dzink, and C. M. Smith. 1985. Chemically definedmedium for oral microorganisms. J. Clin. Microbiol. 22:303–305.
32. Tettelin, H., K. E. Nelson, I. T. Paulsen, J. A. Eisen, T. D. Read, S. Peterson,J. Heidelberg, R. T. DeBoy, D. H. Haft, R. J. Dodson, A. S. Durkin, M.Gwinn, J. F. Kolonay, W. C. Nelson, J. D. Peterson, L. A. Umayam, O. White,S. L. Salzberg, M. R. Lewis, D. Radune, E. Holtzapple, H. Khouri, A. M.Wolf, T. R. Utterback, C. L. Hansen, L. A. McDonald, T. V. Feldblyum, S.Angiuoli, T. Dickinson, E. K. Hickey, I. E. Holt, B. J. Loftus, F. Yang, H. O.Smith, J. C. Venter, B. A. Dougherty, D. A. Morrison, S. K. Hollingshead,and C. M. Fraser. 2001. Complete genome sequence of a virulent isolate ofStreptococcus pneumoniae. Science 293:498–506.
33. van Asseldonk, M., G. Rutten, M. Oteman, R. J. Siezen, W. M. de Vos, andG. Simons. 1990. Cloning of usp45, a gene encoding a secreted protein fromLactococcus lactis subsp. lactis MG1363. Gene 95:155–160.
34. Van Houte, J., J. Russo, and K. S. Prostak. 1989. Increased pH-loweringability of Streptococcus mutans cell masses associated with extracellular glu-can-rich matrix material and the mechanisms involved. J. Dent. Res. 68:451–459.
35. Wuenscher, M. D., S. Kohler, A. Bubert, U. Gerike, and W. Goebel. 1993.The iap gene of Listeria monocytogenes is essential for cell viability, and itsgene product, p60, has bacteriolytic activity. J. Bacteriol. 175:3491–3501.
Editor: E. I. Tuomanen
VOL. 69, 2001 GENETIC AND PROTEIN ANALYSIS OF GbpB FROM S. MUTANS 6941
on August 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from





























