Embed Size (px)
Citation preview

Cloning of the human claudin-2 5’-flanking region revealed a TATA-less
promoter with conserved binding sites in mouse and human for Caudal-
Related Homeodomain Proteins and Hepatocyte Nuclear Factor-1
Takanori Sakaguchi*, Xiubin Gu*, Heidi M. Golden*, EunRan Suh$, David B.
Rhoads#, and Hans-Christian Reinecker*
* Gastrointestinal Unit, Department of Medicine, Center for the Study of
Inflammatory Bowel Disease; # Pediatric Endocrine Unit, Department of
Pediatrics; Massachusetts General Hospital & Harvard Medical School, 32 Fruit
Street, Boston, Massachusetts 02114; and $ Department of Internal Medicine,
University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania
19104
Running Title: Organ specific regulation of claudin-2 gene expression.
Abbreviations: HNF, hepatocyte nuclear factor; NF-κB, nuclear factor-kappa B;
SI, sucrase isomaltase; and LPH, lactase-phlorizin hydrolase
Address correspondence to: Hans-Christian Reinecker
Gastrointestinal Unit
Massachusetts General Hospital
32 Fruit Street, Boston, MA 02114
Phone (617) 724 2172, Fax (617) 726 3673
E-mail: [email protected]
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on April 4, 2002 as Manuscript M110261200 by guest on June 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Sakaguchi et al. 2
Summary
Claudin-2 is a structural component of tight junctions in the kidneys, liver
and intestine, but the mechanisms regulating its expression have not been
defined. The 5’-flanking region of the claudin-2 gene contains binding sites for
intestinal specific Cdx homeodomain proteins and hepatocyte nuclear factor
(HNF)-1, which are conserved in human and mouse. Both, Cdx1 and Cdx2,
activated the claudin-2 promoter in the human intestinal epithelial cell line Caco-
2. HNF-1α augmented the Cdx2- but not Cdx1-induced transcriptional activation
of the human claudin-2 promoter. In mice, HNF-1α was required for claudin-2
expression in the villus epithelium of the ileum and within the liver, but not in the
kidneys, indicating an organ specific function of HNF-1α in the regulation of
claudin-2 gene expression. Tight junction structural components, which
determine epithelial polarization and intestinal barrier function, can be regulated
by homeodomain proteins that control the differentiation of the intestinal
epithelium.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 3
Introduction
Claudin-2 is a regulatory component of tight junctions in the liver, the
kidneys, and the epithelium of the small and large intestines (1,2). Claudin-2
expression has been demonstrated to be involved in the regulation of the
intestinal barrier function by immune modulators (3). Claudins form a family of
proteins composed of at least 24 members, which are expressed in an organ
specific manner and regulate the tissue-specific physiological properties of tight
junctions (4,5). Tight junctions not only create a primary barrier to prevent
paracellular passage of solutes and pathogens but they also restrict the lateral
diffusion of membrane lipids and proteins to maintain cellular polarity (5-8).
Evidence is mounting that claudins are actively involved in the regulation of
paracellular transport of ions through tight junctions (9,10). The modulation of
selective transport through tight junction may require the coordinated expression
of distinct claudins in a particular cell type (1,11). Therefore the regulation of
claudin expression may determine the fundamental ability of the intestinal
epithelium to modulate water- or ion-transport and barrier function. However, the
transcriptional events involved in the organ specific expression of claudins have
not been determined.
Cdx1 and Cdx2 are members of the caudal-related homeobox gene family
based on their sequence homology to the caudal gene of Drosophila
melanogaster (12-14). In vitro and in vivo studies of Cdx1 and Cdx2 suggest that
these transcription factors are important in the early differentiation and
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 4
maintenance of intestinal epithelial cells. In vitro experiments show significant
functional effects of Cdx genes on intestinal differentiation (15,16), proliferation
(15,17), and intestine-specific gene transcription (18-22). Overexpression of
Cdx2 in undifferentiated IEC-6 intestinal epithelial cells leads to the development
of a differentiated phenotype (15). Cdx1 and Cdx2 have been shown to regulate
intestine-specific gene transcription by binding to several intestine-specific
promoters (18-20,23,24). In intestinal epithelial Caco-2 cells, Cdx2 expression
induces the expression of sucrase-isomaltase (SI) and lactase-phlorizin
hydrolase (LPH), two markers of intestinal differentiation (25).
In the regulation of LPH expression Cdx2 directly interacts with
Hepatocyte Nuclear Factor (HNF)-1α (21). HNF-1α and HNF-1β are related
transcription factors which bind to DNA as homo- or hetero-dimers (26). HNF-1α
and HNF-1β are known to be important for liver-specific gene transcription, but
are also expressed in other organs, such as pancreas, kidney, stomach and
intestine (27-29).
In this report, we examined the Cdx and HNF-1α mediated regulation of
the claudin-2 promoter in the human intestinal epithelial cell line Caco-2 and
determined the claudin-2 mRNA and protein expression in HNF-1α deficient
mice. These experiments identify claudin-2 as a target of Cdx homeoproteins and
HNF-1α function in human intestinal epithelial cells. HNF-1α regulated the
complex pattern of claudin-2 expression along the crypt-villus axis of the mouse
ileum and was required for claudin-2 expression in the liver.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 5
Experimental procedures
Antibodies and expression vectors
Polyclonal antibodies recognizing claudin-1 and claudin-2 were obtained
from Zymed Laboratories, Inc. (San Francisco, CA). Anti-Cdx1 and Cdx2
polyclonal antibodies were previously described (30,31). Antibodies for human
HNF-1α and HNF-1β were from Santa Cruz Biotechnology, Inc. (Santa Cruz,
CA). Horseradish peroxidase conjugated anti-rabbit and anti-goat antibodies
were obtained from Amersham (Arlington Heights, IL) and Santa Cruz,
respectively. Mouse Cdx1- or Cdx2-expression vectors (pRc/CMV-Cdx1 or
pRc/CMV-Cdx2) were previously described (15,18). Human HNF-1α- or HNF-
1β-expression vectors (32) and mouse HNF-1α-expression vector (pBJ5mHNF-
1α) (27) were kindly provided by Dr. Marco Pontoglio (Institut Pasteur, Paris,
France) and Dr. Gerald R. Crabtree (Stanford University School of Medicine,
CA), respectively.
Cell culture
The human colon cancer derived cell line Caco-2, human hepatocellular
carcinoma derived cell line HepG2, and mouse mesenchymal cell line NIH3T3
cells were obtained from American type Culture Collection (Rockwell, MD).
These cells were grown in Dulbeco’s modified Eagle’s medium (DMEM) (Cellgro,
Mediatech Inc., Herndon, VA), supplemented with 100 IU/ml penicillin, 100 µg/ml
streptomycin and 10 % (for HepG2 and NIH3T3) or 20% (for Caco-2) heat-
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 6
inactivated fetal calf serum (Sigma, St. Louise, MO), in a humidified 5% CO2
atmosphere at 37 °C. The human colon cancer derived cell line T-84 cells were
grown in DMEM/F-12 (1:1) with antibiotics described above and 10 % heat-
inactivated fetal calf serum.
Isolation of the 5’ flanking region of the human claudin-2 gene and cloning
of human claudin-2.
A degenerate primer approach with primers: 5’-TGG ATG GA(AG) TGT
GC(ATGC) AC(ATGC) CA(CT) -3’; 5’-GA GCA (GA)GA (AG)AA GCA (ATGC)AG
(AGTC)AT(GTA)AT (AGTC)CC-3’, corresponding to the mouse claudin-2
sequence, was used to amplify 407bp of the open reading frame of human
claudin-2. Database searches with the putative human claudin-2 sequence
identified several human claudin-2 EST clones, which were used to complement
the 5' and 3’ sequence. Additional polymerase chain reaction (PCR) with the
primers: 5’-GCT TCT ACT GAG AGG TCT G -3’; 5’-TTC TTC ACA CAT ACC
CTG-3’, and DNA sequencing was utilized to confirm the expression of the full-
length human claudin-2 sequence in T-84 cells. DNA and amino acid sequence
of human claudin-2 has been submitted to Genbank, and is available under the
accession number AF250558. The GenomeWalker Kit (Clontech, Palo Alto, CA)
was used to isolate the 5’-flanking region of the human claudin-2 gene. In brief,
the first PCR was performed with the gene specific primer 1 (5’-CAA AAG CCC
CAG AAG GCC TAG GAT GTA G -3’; +30 to +57 relative to the adenosine of
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 7
the methionine start codon of the human claudin-2 cDNA; accession number
AF250558 or AF177340) and the Adaptor Primer 1. The second PCR was done
with the gene specific primer 2 (5’-GGC AGA CCT CTC AGT AGA AGC GTC
TTC -3’; -27 to -1; corresponding to 493 – 519 sequence of AF177340) and the
Adaptor Primer 2. The longest PCR fragment was purified and subcloned into
pCR2.1 vector (Invitrogen, Carlsbad, CA). The resulting plasmid was designated
as pCR-hCL2p and sequenced.
Deletion constructs, mutagenesis, and reporter gene assay
The KpnI / XhoI fragment of pCR-hCL2p was subcloned into the KpnI /
XhoI site of the pGL3B vector (Promega, Madison, WI). Various length
fragments of the 5’-flanking region of the human claudin-2 gene were amplified
by PCR and subcloned into pGL3B. To obtain –62 construct, HindIII / XbaI
fragment from –84 construct was ligated to EcoRI / HindIII-digested –84
construct with complementary 38-base oligonucleotides (designated as -62wt,
from –62 to –31: sense, 5’-AAT TCA TAT TTA ATC TGG TTT ATG GAT TTT
TTT TAG GT-3’; antisense, 5’-CTA GAC CTA AAA AAA ATC CAT AAA CCA
GAT TAA ATA TG) with 5’-EcoRI and 3’-XbaI overhangs (underlined). To make
mutant claudin-2 promoter constructs, mutated 38-base oligonucleotides were
substituted for wildtype sequence. For Mut 1, Mut 2, and Mut 1+2, 5’-AAT TCA
TAT TTA ATC TGG TGG CTG GAT TTT TTT TAG GT-3’, 5’-AAT TCA TAT TTA
ATC TGG TTT ATG GAT TTT TTG GCG GT-3’, and 5’-AAT TCA TAT TTA ATC
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 8
TGG TGG CTG GAT TTT TTG GCG GT-3’ were used, respectively. (Only sense
strands are shown, nucleotide substitutions are indicated in bold letters.)
For reporter assays, a DNA transfection mixture was prepared consisting
of 1 µg of the reporter construct and 20 ng of pRL-CMV (Promega) as an internal
control. Cells were split onto 6-well plates 18 hr before transfection. Cell
confluency at transfection was 40-60 %. The individual DNA mixtures were
transfected with LipofectAMINE Plus (Life Technologies, Gaithersburg, MD)
according to the manufacturer’s protocol. For co-transfection experiments, 0.5
µg of the expression vector was transfected along with reporter vectors.
pcDNA3.1 vector (Invitrogen) was used to equalize the amount of transfected
DNA. Cells were harvested 48 hrs after transfection, and the luciferase activity
was measured using Dual-Luciferase Reporter Assay System (Promega) and a
luminometer. Transfection efficiencies were normalized to renilla luciferase
activity of the pRL-CMV vector and results are expressed as mean relative
luciferase activity ± SD, of at least three independent experiments.
Electrophoretic Mobility Shift Assay (EMSA)
Nuclear proteins were prepared as previously described (33). Cytosolic
fractions obtained during this procedure were separated for Western blot
analysis. The double-stranded oligonucleotides, -62wt, Mut1, Mut2 and Mut1+2,
were used as probes or cold competitors to analyze the interaction between Cdx
protein and DNA. The HNF-1 wild type probe from –67 to –51 of human claudin-
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 9
2 gene sequence consisted of complementary 29-nucleotide oligonucleotides
(sense; 5’-AAT TCC TGG TCA ATA TTT AAT CTG T -3’, antisense; 5’-CTA GAC
AGA TTA AAT ATT GAC CAG G) with 5’-EcoRI and 3’-XbaI overhangs
(underlined). Mutant HNF complementary oligonucleotides were following:
sense; 5’-AAT TCC TAA TTC AGG TTT AAT CTG T-3’, antisense; 5’-CTA GAC
AGA TTA AAC CTG AAT TAG G-3’ (nucleotide substitutions are indicated by
bold).
The probes were labeled with Klenow enzyme by fill-in incorporation with
nucleotide triphosphates, including [α-32P] dATP. The binding reaction was
performed as previously described (34). For a competition assay, a 100-fold
excess of unlabeled oligonucleotide was added to the reaction. To perform
supershift assay, the binding mixtures were incubated for 10 minutes at room
temperature in the presence of 1 µL of antibodies. Samples were fractionated on
4% non-denaturing polyacrylamide gel in 0.5X TBE buffer. The resultant DNA-
protein complexes were detected by autoradiography.
Western blot analysis
The protein concentration of each sample was quantified by the Bradford
method. Samples were electrophoresed through a 4-20 % gradient SDS
polyacrylamide gel and transferred onto polyvinylidene difluoride (PVDF)
membranes (Millipore, Bedford, MA). The blots were blocked overnight at 4 °C
with 10 % dry milk in PBS containing 0.1 % Tween 20 (PBS-T), followed by the
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 10
incubation for 3 hours at room temperature (RT) with primary antibodies diluted
in blocking buffer at 1:1,000. After washing in PBS-T for 30 minutes, the blots
were incubated with secondary antibodies diluted in blocking buffer for 45
minutes at RT. The hybridized bands were detected by ECL kit (Amersham),
according to the manufacturer’s instruction.
RNA extraction and Northern Blot analysis
Total RNA was isolated from tissues using Trizol reagent (Life
Technologies). Total RNA (30 µg) was electrophoresed in a 1 % agarose
formaldehyde gel and transferred to a nylon membrane (Magna NT,
MicroSeparations Inc., Westbrough, MA) by capillary blotting. Probes were
labeled with [α-32P] dCTP using Rediprime Random Primer Labeling Kit
(Amersham). Membranes were hybridized with radio-labeled probes in Quickhyb
solution (Stratagene, La Jolla, CA) at 65 °C for 1 hour. The membranes were
washed with 0.1% SDS, 2X sodium chloride sodium citrate buffer at RT for 15
minutes and at 65 °C for 10 minutes. The blots were analyzed by
autoradiography. The probes used to detect claudin-1, claudin-2 and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were described
previously (3). Other probes were: Cdx1, a 0.9 kb HindIII / XbaI fragment of
pRc/CMV-Cdx1; Cdx2, a 0.9 kb HindIII fragment of pRc/CMV-Cdx2; and HNF-
1α, a 0.4 kb SmaI fragment of pBJ5mHNF1α. The HNF-1α probe derives from a
unique sequence in HNF-1α cDNA and does not cross-hybridize with HNF-1β.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 11
Northern blots were densitometrically analyzed and gene specific mRNA
expression levels were normalized to GAPDH mRNA expression levels in the
same samples, and expressed as mean density/area calculated from 3
independent experiments.
Tissue preparation and immunohistochemistry
Mice carrying the HNF-1α null allele were obtained from Dr. Frank J.
Gonzalez (NIH, MD) (35). Homozygous HNF-1α null and wildtype littermates
were obtained by mating heterozygous carriers. All animal experiments were
performed in accordance with National Institutes of Health guidelines and
protocols approved by the Subcommittee on Research Animal Care at our
institute. The liver and kidney were removed and washed with ice-cold PBS.
Segments of 2 cm from the most proximal jejunum and most distal ileum were
collected. For immunostaining, small tissue blocks were mounted in OCT
compound and frozen in dry ice-ethanol. For RNA extraction, small pieces of
tissues were snap-frozen at –80 °C.
Cryosections of frozen tissues with 4-µm thickness were prepared. The
sections were air-dried and fixed in methanol at -20 °C for 10 minutes followed by
rehydration in PBS at 4 °C for 30 minutes as previously described (2). The
sections were blocked with 0.5 % normal donkey serum in PBS (blocking
solution) for 1 hour at 20 °C, and incubated with primary antibodies or normal
rabbit serum diluted at 1:100 with blocking solution for 3 hours at RT. After three
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 12
washes with PBS, the slides were incubated at RT with FITC-labeled anti-rabbit
antibody (Vector Laboratories, Burligame, CA) diluted at 1:500 with blocking
solution for 1 hour in the dark and analyzed with an AX-70 Olympus fluorescent
microscope.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 13
Results
Isolation of the 5’-flanking region of the human claudin-2 gene
The 5’-flanking region of the claudin-2 gene isolated from the human
genomic library and T-84 cell-derived claudin-2 cDNA sequence were confirmed
by sequence comparison with the human genomic clone AL158821. BLAST
search revealed that the gene encoding human claudin-2 is located on
chromosome X, mapping to q22.3-23. The claudin-2 mRNA expressed in T-84
cells contains an open reading frame of 693bp. Human and mouse claudin-2
have a high sequence identity of 87% on mRNA level and 93% identity on amino
acid level.
Comparison with the mouse claudin-2 promoter in genomic databases
revealed that the promoters of the human and mouse claudin-2 genes possess a
remarkable homology of 84% for the region of -1 to –400 (Fig. 1). The mouse
claudin-2 cDNA (Genbank AK004990) recovered by cap-trapping revealed the
putative transcriptional start site at 152 basepair (bp) upstream of the
translational start codon. The transcriptional initiation site is located within a
consensus initiator (Inr) element (NCANNNNN) (36,37).
The promoters of the human and mouse claudin-2 genes have no TATA
box near the putative transcriptional initiation site (Fig. 1). However, a CAAT box
is located at –60 to -63 bp and two E-boxes (CANNTG) at –198 to -195 bp and
–67 to –62 bp (Fig. 1), suggesting that regulatory elements to initiate gene
transcription are present.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 14
The human claudin-2 promoter contains two sites for the intestine-specific
homeodomain protein family Cdx (18), designated CdxA and CdxB (Fig. 1). The
promoter has also binding sites for HNF-1 and HNF-3β (38), as well as putative
AP-1 (39), NF-κB (40) and GATA (41) binding sites. Particularly, the first Cdx
binding site CdxA, the HNF-1, HNF-3β and GATA binding sites are conserved in
human and mouse claudin-2 promoters (Fig. 1).
To identify the regions involved in regulating claudin-2 gene transcription,
sequentially deleted 5’-flanking regions (–1041, -393, -84, -62 and –31 to +148)
were cloned into the reporter plasmid pGL3B. Reporter constructs were
transfected into intestinal epithelial cell line Caco-2, hepatic cell line HepG2 or
fibroblast cell line NIH3T3. In Caco-2 cells the claudin-2 promoter fragments
containing –1040 to –62 bp of the 5’-flanking region induced a 18 - 29-fold
increase in relative luciferase activity above that observed after transfection with
the control null reporter construct (Fig. 2). In contrast, the same promoter regions
achieved only a 7-11-fold increase when transfected into HepG2 and NIH3T3
cells (Fig. 2).
Removal of the putative AP-1 and the NF-κB sites decreased the
promoter activity slightly. Disruption of the HNF-1 binding site in the -62 bp
construct did not alter the promoter activity significantly in Caco-2 cells. However,
removal of the Cdx binding sites resulted in a loss of promoter activity in Caco-2,
HepG2 and NIH3T3 cells (Fig. 2).
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 15
Claudin-2 promoter activity is regulated by Cdx homeodomain protein
overexpression in Caco-2 cells.
To examine the function of the two Cdx sites, mutations were introduced
into CdxA (Mut 1), CdxB (Mut 2), or both (Mut 1+2) (Fig. 3A). As shown in Figure
3B, mutation in CdxA (Mut 1) or CdxB (Mut 2) sites decreased the promoter
activity to 30% and 61% of that observed with -62 wildtype construct,
respectively. When both sites were mutated (Mut 1+2), promoter activity was
decreased to 15% of the wildtype construct, comparable to the –31 construct
lacking both Cdx sites.
Next we determined the ability of Cdx1 and Cdx2 to activate the claudin-2
promoter. As shown in Figure 4A, Cdx2 but not Cdx1 protein was detectable in
nuclear proteins from Caco-2 cells. Transient expression with either Cdx1 or
Cdx2 alone or in combination resulted in the strong expression of these proteins
in the nuclei of Caco-2 cells 48 hours after transfection (Fig. 4A). Ectopic
expression of Cdx1 did not alter the expression level of Cdx2, nor did Cdx2-
overexpression induce Cdx1 protein expression in the nuclei (Fig. 4A).
As shown in Figure 4B, Cdx1-overexpression resulted in 3.5-fold increase
of the promoter activity driven by -62 construct containing both intact Cdx sites in
Caco-2 cells (92-fold relative to the activity of null pGL3B vector). In contrast,
Cdx2-overexpression increased the activity of the same construct up to 6.7-fold
(177-fold of null pGL3B vector activity).
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 16
Overexpression of Cdx1 and Cdx2 together did not significantly increase
the activity above the values achieved by Cdx2 alone (Fig. 4B). Although
mutation of either CdxA (Mut 1) or CdxB (Mut 2) site retained the ability to
respond to Cdx2-overexpression, promoter activities induced by Cdx2-
overexpression were less than 25 % and 43 % of that observed in -62 construct,
respectively. Similarly, Mut 1 and Mut 2 constructs were less sensitive to Cdx1-
overexpression. In the absence of both Cdx sites neither Cdx1- nor Cdx2-
overexpression induced a significant induction of reporter gene transcription in
Caco-2 cells (Fig. 4B). The ability of Cdx2 to induce a stronger induction of
claudin-2 promoter activity in comparison to Cdx1 was specific for Caco-2 cells.
As demonstrated in Figure 4C, Cdx1 and Cdx2 enhanced claudin-2 promoter
activity in fibroblasts 2.7-fold and 2.8-fold, respectively, whereas Cdx1 induced a
2.9 fold and Cdx2 a 6.7-fold higher promoter activity in Caco-2 cells.
Cdx-2 binds to the Cdx responsive elements of the human claudin-2 promoter
To further define the interaction between Cdx2 and the two Cdx sites,
EMSA and supershifts were performed with nuclear proteins from Caco-2 cells.
These experiments were carried out in post-confluent Caco-2 cells since it was
shown that specific Cdx-DNA complexes can be obscured by unspecific binding
of unknown peptides in nuclear proteins from pre-confluent Caco-2 cells (22).
As shown in Figure 5A, three DNA-protein complexes (A, B and C) were
observed when binding reactions were carried out with radio-labeled wildtype
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 17
oligonucleotide containing both intact Cdx sites (Fig.5A, lane 1). Unlabeled Mut 2
oligonucleotide with an intact CdxA but a mutated CdxB competed with the
formation of all three complexes whereas unlabeled Mut 1 oligoncleotide with a
mutated CdxA but an intact CdxB prevented only the formation of complex B
(Fig. 5A, lane 3 and 4). In supershift assays, anti-Cdx2 antibody shifted only
complex A to reveal two distinct Cdx2-containing protein-DNA complexes (Fig.
5A, lane 7). In contrast, anti-Cdx1 antibody did not affect the mobility of the
complexes (Fig. 5A, lane 6). The Cdx2-containing complex A was also formed
with radio-labeled Mut 2 oligonucleotide used as a probe (Fig. 5A lane 10),
suggesting that this complex is preferentially formed with CdxA. Complex B did
not form on the Cdx sites since this complex was detected and consequently
competed by all three mutated oligonucleotides (Fig. 5A lane 3-5, 9, 11 and 13).
Mutation in CdxA site greatly reduced the formation of complex C, suggesting the
formation of complex C is dependent on this site (Fig. 5A, lane 8 and 12).
Within the SI gene promoter two adjacent Cdx consensus sites may be
able to direct the formation of Cdx2 homodimers (18). We therefore further
characterized the potential coordination of Cdx2 binding by the two Cdx sites. In
these experiments increasing amounts of nuclear proteins of Cdx2-transfected
Caco-2 cells were used. Complex A, which was supershifted by anti-Cdx2
antibody, was observed even in the absence of CdxA site when more nuclear
protein was used (Fig. 5B, lane 7 and 8). However, most of the Cdx2 containing
complexes formed in the presence of CdxA site and did not require the CdxB site
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 18
(Fig. 5, lane 2 - 4, and 10 -12). In addition, when 12 µg of nuclear proteins from
Cdx2-transfected Caco-2 cells were used, an additional complex D was
observed, which was shifted by anti-Cdx2 antibody (Fig. 5B, lane 3, 4, 11 and
12). Although we could not visualize an additional supershifted band derived from
complex D, it may correspond to Cdx2 homodimers, which could not be
distinguished in supershifts from monomeric complexes (18).
HNF-1 enhances Cdx2 mediated activation of human claudin-2 promoter in
Caco-2 cells.
Cdx2 has been shown to regulate intestinal specific LPH gene expression
in synergy with HNF-1α (21). We therefore determined if the HNF-1 site in the
human claudin-2 promoter could contribute to transcriptional regulation. We
compared the effect of HNF-1α- and HNF-1β-overexpression, since both proteins
share highly homologous DNA binding domains but have distinct activation
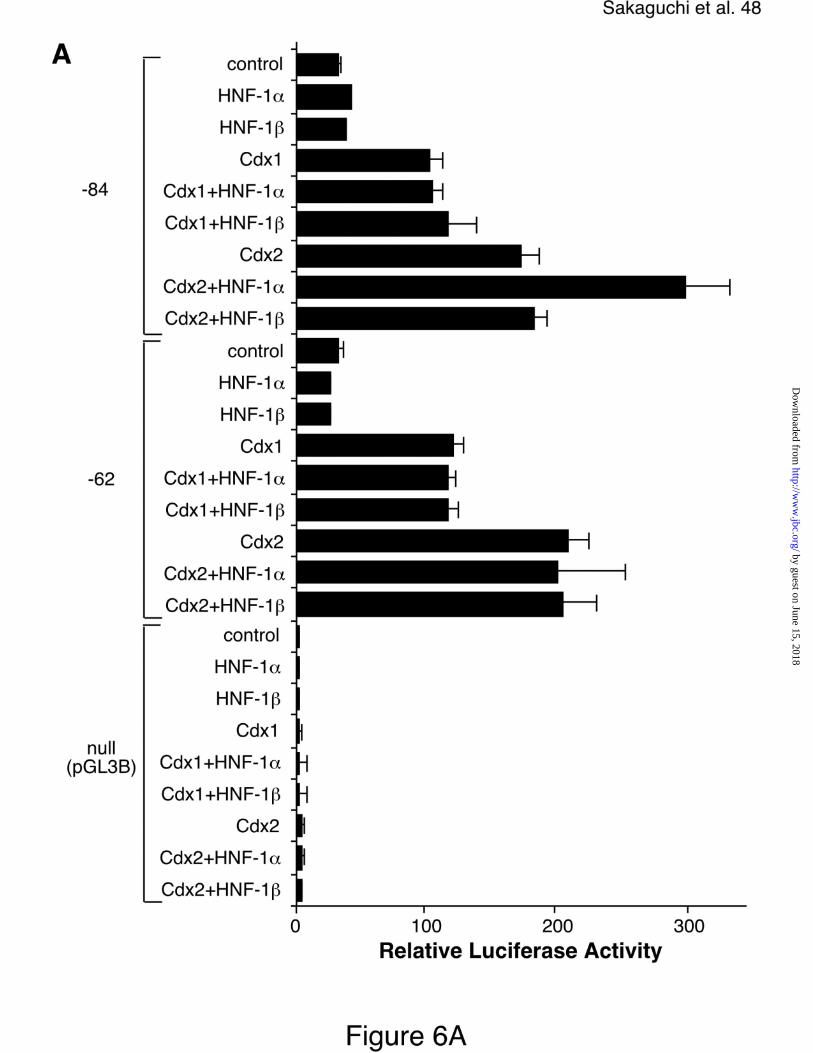
domains (42). As shown in Figure 6A, Cdx1- and Cdx2-overexpression resulted
in a 3-fold and 5-fold increase of the promoter activity driven by –84 construct
(100-fold and 170-fold relative to that of null pGL3B vector), respectively.
However, transfection of either HNF-1α or HNF-1β alone was not able to
increase promoter activity. In contrast, co-transfection of HNF-1α together with
Cdx2 but not Cdx1 resulted in a 9-fold increase of the promoter activity (293-fold
of the null pGL3B activity) (Fig. 6A). Disruption of the HNF-1 site in –84 reporter
construct prevented a synergistic co-operation of HNF-1α and Cdx2 (Fig. 6A, -62
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 19
construct). As shown in Figure 6B, transfection of Caco-2 cells with HNF-1α
expression constructs resulted in an increase of HNF-1α expression in both the
cytosolic and nuclear protein fractions. In contrast, Cdx2 was exclusively
expressed in nuclear protein fractions of Caco-2 cells even after ectopic
expression (Fig. 6B).
HNF-1 binds its recognition sequence within the human claudin-2 promoter.
To further determine the interaction between HNF-1 proteins and HNF-1
binding site in the human claudin-2 promoter, EMSA and supershifts were
performed with nuclear proteins from Caco-2 cells. As shown in Figure 7, a single
DNA-protein complex was observed when nuclear proteins from mock-
transfected Caco-2 cells was used (lane 1). Addition of 100-fold excess of
unlabeled wildtype but not mutant oligonucleotide prevented the formation of this
complex (Fig. 7, lane 2 and 3). The HNF-1 consensus sequence-protein
complex was supershifted efficiently by anti-HNF-1α antibody but only to a small
extent by anti-HNF-1β antibody (Fig. 7, lane 4 and 5). Transfection with either
HNF-1α- or Cdx2 alone did not alter the formation of this complex (Fig. 7, lane 6
and 7). In contrast, co-transfection with Cdx2 and HNF-1α together resulted in
the increased formation of the complex, which was supershifted by anti-HNF-1α
antibody (Fig. 7, lane 8 and 9).
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 20
HNF-1 is an organ specific regulator of claudin-2 expression.
The in vitro experiments identified HNF-1α as a potential regulator of
claudin-2 expression. Cdx1 and Cdx2 expression is restricted to the intestine,
whereas HNF-1α is also a regulator of gene expression in the liver and kidneys,
organs in which claudin-2 is expressed (1,5,42). In contrast to Cdx2 deficient
animals (43), HNF-1α deficient mice are viable and survive to adulthood (35,44).
We utilized these mice to determine the potential contribution of HNF-1α in the
expression of claudin-2 in different organs. Analysis of the claudin-2 mRNA and
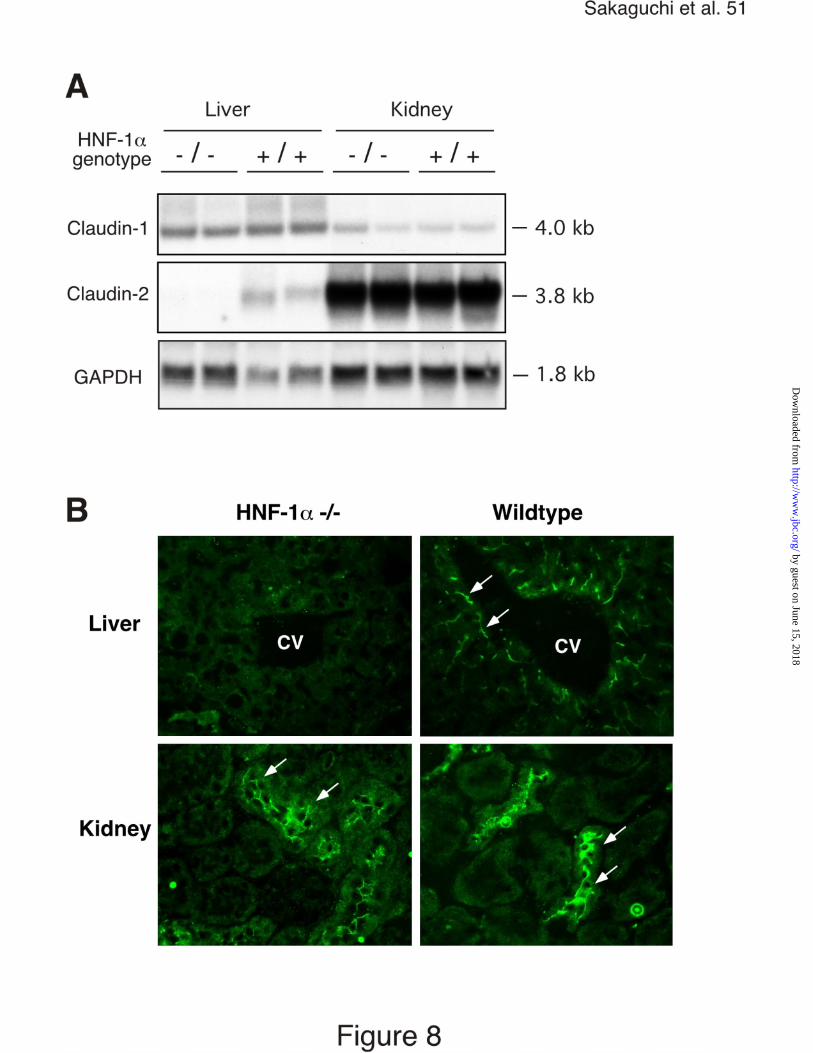
protein expression in these animals revealed that HNF-1α was required for
expression of claudin-2 in the liver (Fig. 8A). Claudin-2 mRNA and protein
expression was absent in the liver of HNF-1α deficient animals whereas claudin-
1 mRNA expression was unaffected (Fig. 8A and B). In contrast, HNF-1α was not
required for claudin-2 mRNA and protein expression in the kidneys (Fig. 8A and
B).
We next analyzed the expression of claudin-1 and claudin-2 along the
cephalo-caudal and crypt-villus axis in wildtype and HNF-1α deficient mice, since
the in vitro experiments suggest the ability of HNF-1α to regulate claudin-2
expression in the presence of Cdx homedomain proteins in intestinal epithelial
cells. Densitometric analysis of Northern blots after normalization to GAPDH
mRNA expression demonstrated that claudin-2 mRNA was differentially
expressed along the cephalo-caudal axis. In wildtype mice, claudin-2 mRNA was
expressed at 17.2 ± 1.7–fold higher levels in the ileum than in the jejunum (Fig.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 21
9A and B), in good agreement with the recent analysis of claudin-2 protein
expression in the rat intestine (2). In contrast, claudin-1 mRNA was expressed at
2.4 ± 0.4–fold higher levels in the jejunum than in the ileum (Fig. 9A and B).
Claudin-2 mRNA expression pattern correlated with Cdx1 mRNA expression in
the same intestinal segments, which increased 5 ± 0.5-fold from the jejunum to
the ileum (Fig.9A). However, Cdx2 mRNA expression levels were similar in the
jejunum and ileum (Fig. 9A). In the absence of HNF-1α, claudin-2 expression
decreased by 55 ± 10 % in the ileum (Fig. 9A and B). This regulation was specific
for claudin-2, since claudin-1 mRNA expression was not altered in the absence
of HNF-1α in the mouse jejunum and ileum (Fig. 9A and B).
The reduction of claudin-2 mRNA expression could be due to an overall
reduction of claudin-2 gene transcription or a reduced expression in specific
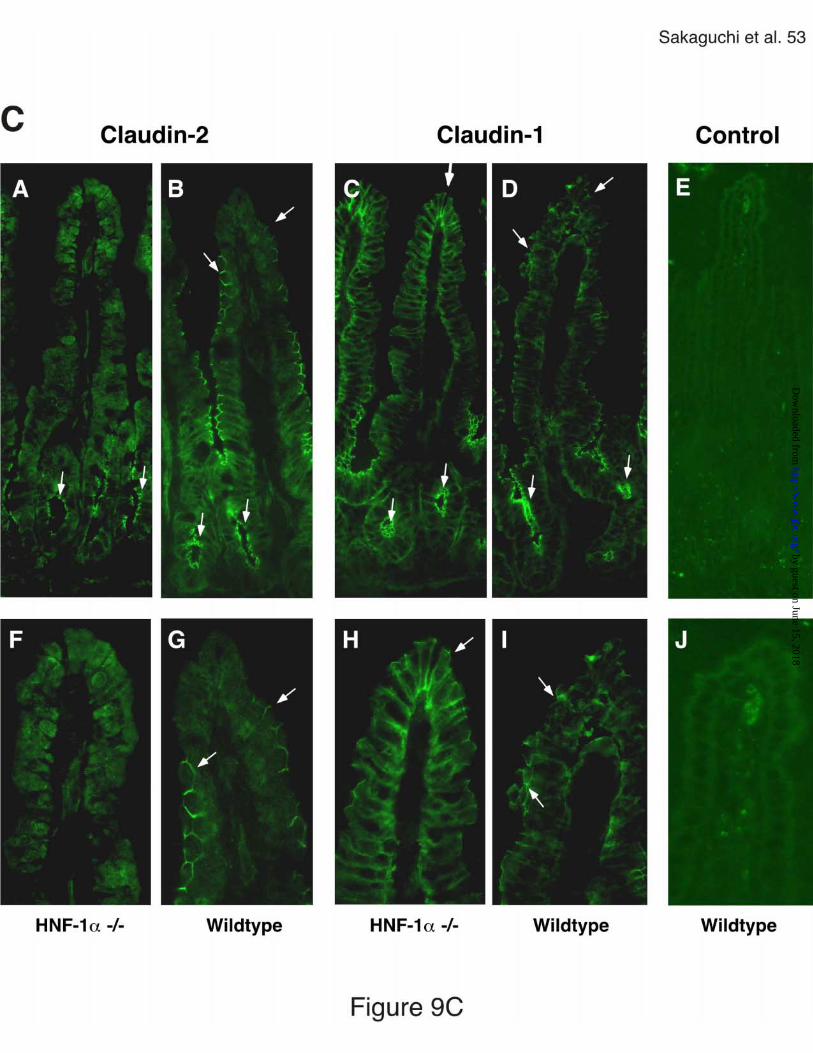
intestinal epithelial cell subsets. We therefore determined the expression and
subcellular distribution of claudin-1 and claudin-2 proteins in the ileum by
immunostaining along the crypt-villus axis in wildtype and HNF-1α deficient mice
(Fig. 9C). Claudin-2 protein was expressed in tight junctions of the crypt and
villus epithelium of the ileum in wildtype mice (Fig. 9C, panel B and G). In the
absence of HNF-1α, claudin-2 expression was restricted to the tight junctions of
the crypt epithelium (Fig. 9C, panel A and F). Claudin-1 expression was not
altered in the absence of HNF-1α, and was observed in tight junctions of the
crypt and villus epithelium of the ileum in both wildtype and HNF-1α deficient
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 22
mice (Fig. 9C, panel C, D, H and I). Incubation with rabbit control serum did not
result in detectable immunoreactivity (Fig. 9C, panel E and J).
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 23
Discussion
The diverse claudin family of tight junction associated proteins has the
potential of directing the variability of paracellular transport and barrier functions
within gastro-intestinal organs (5). Recent evidence demonstrated that claudins
are not only involved in the induction of tight junction formation, but are also able
to regulate water and ion specific paracellular transport mechanisms (9,10). Loss
of claudin-16 results in the inability to absorb magnesium in the thick ascending
limb of Henle (10). Claudin-4 expression resulted in the specific decrease in
absolute sodium permeability, whereas claudin-2 appeared to increase
paracellular conductance in kidney epithelial cells without changing the
paracellular transport of inert compounds (9). The molecular mechanisms
orchestrating the organ specific expression of claudin-2 are unknown. In this
report we provide the first insights into the transcriptional activation events, which
regulate the complex expression pattern of claudin-2.
We demonstrate that the mouse and human claudin-2 promoters contain
conserved binding sequences for Cdx homeodomain proteins and for the POU
homedomain family member HNF-1α. Cdx1 and Cdx2, intestine specific
homeobox proteins, play an important role for the transcription of the intestine
specific expression of several genes such as SI (15), LPH (21), and guanylyl
cyclase C (45). HNF-1α and HNF-1β were first identified as liver-enriched
transcription factors involved in the expression of several plasma proteins,
including albumin and clotting factors (49) and can act either as homo- or
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 24
heterodimers (26). There is increasing evidence that HNF-1α is crucial for the
transcription of the intestine-specific genes such as LPH (21) and SI (29,50).
Our experiments provide the first demonstration that Cdx homeodomain
proteins can initiate transcriptional activation of a TATA-less promoter. In
contrast HNF-family members have been demonstrated to activate tissue type
specific expression of Ksp cadherin (cadherin 16), which lacks TATA-boxes (48).
Similar to the SI and LPH genes, the claudin-2 promoter has two putative
Cdx binding sites. The Cdx binding site containing region of the claudin-2
promoter mediated basal transcriptional activation in intestinal epithelial cells, but
also had activity in fibroblasts and HepG2 cells. Similar to our results, Cdx
consensus binding site containing promoter have been demonstrated to induce
transcriptional activation in fibroblasts without Cdx proteins by undetermined
mechanisms (47). This promoter region may comprise a core promoter, which
contains transcriptional elements sensitive to activation by factors in none-
epithelial cells in the absence of Cdx and HNF-1α. In addition to tissue specific
expressed transcription factors like Cdx1 and Cdx2, silencer, binding upstream of
the investigated promoter region, may be necessary to direct tissue type specific
expression of claudin-2.
Both, Cdx1 and Cdx2 can interact with the Cdx consensus sites within the
claudin-2 promoter, although Cdx2 is the more potent activator of the claudin-2
gene transcription in Caco-2 cells. The stronger induction of claudin-2 promoter
activity by Cdx2 in comparison to Cdx1 was specific for Caco-2 cells, suggesting
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 25
that in these cells Cdx2 may co-operate with other factors enhancing its
transcriptional activity. Our experiments identified HNF-1α as a potential
candidate, since it was able to enhance Cdx2 but not Cdx1 induced claudin-2
promoter activity in Caco-2 cells.
Our results are consistent with the previous observation that Cdx2 is more
effective than Cdx1 in transcriptional activation of the clusterin gene promoter
(46). Although both Cdx binding sites were required for full transcriptional activity
of the human claudin-2 promoter in Caco-2 cells, Cdx2 binding occurred primarily
at the CdxA site. The second CdxB site may serve primarily to support Cdx2
homodimer or oligomer formation as has been proposed for the two Cdx sites in
the SI promoter (18). Alternatively, additional transcription factors may require
CdxB to bind and enhance Cdx2 mediated transcription. The involvement of
additional transcriptional activators may be particularly necessary in the
activation of the mouse claudin-2 promoter, in which the second Cdx-binding site
present in the human promoter is not completely preserved.
In Caco-2 cells, HNF-1α was able to enhanced claudin-2 promoter activity
only in the presence of overexpressed Cdx2. HNF-1α has been demonstrated to
synergize with Cdx2 to induce LPH gene transcription (21). However, whereas
the LPH promoter can be activated by the expression of HNF-1α alone,
activation of the claudin-2 promoter by HNF-1α in Caco-2 cells was dependent
on the recruitment of overexpressed Cdx2 to its binding site. This co-operation
was specific for HNF-1α since HNF-1β failed to enhance Cdx2-mediated
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 26
activation of the claudin-2 promoter. These results are similar to the previous
observations that HNF-1β was less potent than HNF-1α as a transactivator of
LPH (51), SI (29), and α1-antitrypsin (52) genes. Collectively, the promoter
analysis revealed the ability of Cdx homeodomain proteins and HNF-1α to bind
to their recognition sequences in the claudin-2 promoter and to regulate the
activation of this promoter in Caco-2 cells.
We analyzed wildtype and HNF-1α deficient mice to assess the role of
HNF-1α in the regulation of claudin-2 expression. These experiments indicate
that HNF-1α can regulate claudin-2 expression in an organ specific manner.
HNF-1α was required for claudin-2 expression in the liver. HNF-1α deficient mice
have enlarged fatty livers and dysregulated fatty acid homeostasis, which have
been traced in part to a reduced expression of liver fatty acid-binding protein
(53). It is currently not clear if the lack of claudin-2 contributes to the disturbed
liver function in HNF-1α deficient animals.
In contrast, claudin-2 mRNA and protein expression in proximal tubules of
the kidneys were not altered in the absence of HNF-1α. In the kidney HNF-3 may
compensate for the lack of HNF-1α in the activation of the claudin-2 promoter.
HNF-3 has recently been shown to mediate the kidney specific expression of
Ksp-cadherin through a motif similar to the HNF-3-CAAT box containing
sequence found in the claudin-2 promoter partially overlapping with the HNF-1
consensus sequence (48).
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 27
In the absence of HNF-1α, claudin-2 was still expressed in the small
intestine, although its expression was restricted to the crypt epithelium.
The loss of claudin-2 expression in intestinal villi epithelium may be due to the
lack of HNF-1α, which has been demonstrated to be predominantly expressed in
the intestinal epithelial cells of the small intestinal villi (28).
HNF-1α, Cdx1, and Cdx2, are differential expressed along the crypt-villus
axis of the small intestine (28,31). Cdx1 expression has been demonstrated to
localize to intestinal crypts, whereas Cdx2 expression was observed to extend
into small intestinal villi (31). However, recent experiments with antibodies
recognizing phosphorylated Cdx2 demonstrated activated Cdx2 in small
intestinal crypts (54). If the regulation of claudin-2 expression in mice
corresponds to its regulation in Caco-2 cells, HNF-1α may be required to
enhance Cdx2 mediated claudin-2 expression in the intestinal villi, whereas Cdx1
and/or Cdx2 may drive the remaining expression of claudin-2 in the crypt
epithelium of HNF-1α deficient mice.
The function of HNF-1α in the transcriptional regulation of claudin-2
expression was specific since claudin-1 expression was not regulated in the
absence of HNF-1α in the jejunum or ileum. The different transcriptional
regulation of claudin-1 was further apparent in the distinct expression pattern
along the cephalo-caudal axis and the unaltered expression along the in crypt-
villus axis in the absence of HNF-1α. The impact of HNF-1α gene disruption on
the gut has not been examined in detail. The loss of claudin-2 expression in the
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 28
ileal villi and the liver may contribute to the severe phenotype of the HNF-1α
deficient mice. HNF-1α gene disruption in mice leads to dwarfism due to a
reduced Insulin-like growth factor (IGF)-1 synthesis, and an early onset form of
type 2 diabetes mellitus due to impaired glycolytic signaling (35,44,55). However,
impaired intestinal and liver specific secretive or absorptive function may relate to
these phenotypes. Further analysis of the HNF-1α deficient mice should prove
valuable to uncover additional roles of claudin-2 in the regulation of organ
specific functions.
Our studies suggest that the expression of claudin-2 is under the
regulatory control of HNF-1α in the liver and small intestinal villi in mice. Whereas
in the liver HNF-1α is required for claudin-2 expression, in the intestine HNF-1α
may co-operate with additional factors to extend claudin-2 expression from the
crypt into the functionally distinct villus intestinal epithelial cell compartment. It
needs to be determined if the augmentation of claudin-2 gene expression by
HNF-1α in this compartment is dependent on Cdx2 as observed in Caco-2 cells.
Together our experiments support a model in which claudin-2 expression is
governed by distinct organ specific transcriptional mechanisms involving
homeodomain proteins.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 29
Acknowledgements
This work was supported by National Institutes of Health Grants DK51003,
DK54427 and DK33506 (H.-C.R.), by U. S. Public Health Service Grant DK54399
(DBR) and by March of Dimes Grant#1-FY99-221 (DBR). The authors gratefully
acknowledge Frank J. Gonzalez for providing the HNF-1α deficient mice, Taro
Akiyama for developing the PCR genotyping protocol, and Lihua Zhang for
technical assistance.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 30
References
1. Furuse, M., Fujita, K., Hiiragi, T., Fujimoto, K., and Tsukita, S. (1998)
J Cell Biol 141(7), 1539-50
2. Rahner, C., Mitic, L. L., and Anderson, J. M. (2001)
Gastroenterology 120(2), 411-22.
3. Kinugasa, T., Sakaguchi, T., Gu, X., and Reinecker, H. C. (2000)
Gastroenterology 118(6), 1001-11
4. Tsukita, S., and Furuse, M. (2000) J Cell Biol 149(1), 13-6.
5. Tsukita, S., Furuse, M., and Itoh, M. (2001)
Nature Reviews Molecular Cell Biology 2(4), 285-93
6. Schneeberger, E. E., and Lynch, R. D. (1992)
Am J Physiol 262(6 Pt 1), L647-61
7. Anderson, J. M., and Van Itallie, C. M. (1995)
Am J Physiol 269(4 Pt 1), G467-75
8. Madara, J. (1998) Ann. Rev. Physiol. 60, 143-159
9. Van Itallie, C., Rahner, C., and Anderson, J. M. (2001)
J Clin Invest 107(10), 1319-27.
10. Simon, D. B., Lu, Y., Choate, K. A., Velazquez, H., Al-Sabban, E.,
Praga, M., Casari, G., Bettinelli, A., Colussi, G., Rodriguez-Soriano, J.,
McCredie, D., Milford, D., Sanjad, S., and Lifton, R. P. (1999)
Science 285(5424), 103-6
11. Morita, K., Furuse, M., Fujimoto, K., and Tsukita, S. (1999)
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 31
Proc Natl Acad Sci U S A 96(2), 511-6
12. Charite, J., de Graaff, W., Consten, D., Reijnen, M. J., Korving, J., and
Deschamps, J. (1998) Development 125(22), 4349-58
13. Duprey, P., Chowdhury, K., Dressler, G. R., Balling, R., Simon, D.,
Guenet, J. L., and Gruss, P. (1988) Genes Dev 2(12A), 1647-54
14. James, R., Erler, T., and Kazenwadel, J. (1994)
J Biol Chem 269(21), 15229-37
15. Suh, E., and Traber, P. G. (1996) Mol Cell Biol 16(2), 619-25.
16. Soubeyran, P., Andre, F., Lissitzky, J. C., Mallo, G. V., Moucadel, V.,
Roccabianca, M., Rechreche, H., Marvaldi, J., Dikic, I., Dagorn, J. C., and
Iovanna, J. L. (1999) Gastroenterology 117(6), 1326-38.
17. Lynch, J., Suh, E. R., Silberg, D. G., Rulyak, S., Blanchard, N., and
Traber, P. G. (2000) J Biol Chem 275(6), 4499-506.
18. Suh, E., Chen, L., Taylor, J., and Traber, P. G. (1994)
Mol Cell Biol 14(11), 7340-51.
19. Lambert, M., Colnot, S., Suh, E., L'Horset, F., Blin, C., Calliot, M. E.,
Raymondjean, M., Thomasset, M., Traber, P. G., and Perret, C. (1996)
Eur J Biochem 236(3), 778-88
20. Troelsen, J. T., Mitchelmore, C., Spodsberg, N., Jensen, A. M., Noren, O.,
and Sjostrom, H. (1997) Biochem J 322(3), 833-8
21. Mitchelmore, C., Troelsen, J. T., Spodsberg, N., Sjostrom, H., and
Noren, O. (2000) Biochem J 346 (2), 529-35
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 32
22. Fang, R., Santiago, N. A., Olds, L. C., and Sibley, E. (2000)
Gastroenterology 118(1), 115-27.
23. Drummond, F., Sowden, J., Morrison, K., and Edwards, Y. H. (1996)
Eur J Biochem 236(2), 670-81
24. Drummond, F. J., Sowden, J., Morrison, K., and Edwards, Y. H. (1998)
FEBS Lett 423(2), 218-22
25. Lorentz, O., Duluc, I., Arcangelis, A. D., Simon-Assmann, P., Kedinger, M.,
and Freund, J. N. (1997) J Cell Biol 139(6), 1553-65.
26. Mendel, D. B., Hansen, L. P., Graves, M. K., Conley, P. B., and
Crabtree, G. R. (1991) Genes Dev 5(6), 1042-56
27. Kuo, C. J., Conley, P. B., Hsieh, C. L., Francke, U., and Crabtree, G. R.
(1990) Proc Natl Acad Sci USA 87(24), 9838-42
28. Serfas, M. S., and Tyner, A. L. (1993) Am J Physiol 265(3 Pt 1), G506-13
29. Boudreau, F., Zhu, Y., and Traber, P. G. (2001)
J Biol Chem 276(34), 32122-8
30. Silberg, D. G., Furth, E. E., Taylor, J. K., Schuck, T., Chiou, T., and
Traber, P. G. (1997) Gastroenterology 113(2), 478-86.
31. Silberg, D. G., Swain, G. P., Suh, E. R., and Traber, P. G. (2000)
Gastroenterology 119(4), 961-71.
32. Bach, I., and Yaniv, M. (1993) EMBO Journal 12(11), 4229-42
33. Awane, M., Andres, P. G., Li, D. J., and Reinecker, H. C. (1999)
J Immunol 162(9), 5337-44
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 33
34. Traber, P. G., Wu, G. D., and Wang, W. (1992) Mol Cell Biol 12(8), 3614-27
35. Lee, Y. H., Sauer, B., and Gonzalez, F. J. (1998)
Mol Cell Biol 18(5), 3059-68
36. Suzuki, Y., Tsunoda, T., Sese, J., Taira, H., Mizushima-Sugano, J., Hata,
H., Ota, T., Isogai, T., Tanaka, T., Nakamura, Y., Suyama, A., Sakaki, Y.,
Morishita, S., Okubo, K., and Sugano, S. (2001)Genome Res 11(5), 677-84.
37. Smale, S. T. (1997) Biochim Biophys Acta 1351(1-2), 73-88
38. Cereghini, S. (1996) FASEB J 10(2), 267-82
39. Wasylyk, C., Wasylyk, B., Heidecker, G., Huleihel, M., and Rapp, U. R.
(1989) Mol Cell Biol 9(5), 2247-50.
40. Gilmore, T. D. (1990) Cell 62(5), 841-3.
41. Orkin, S. H. (1992) Blood 80(3), 575-81.
42. Pontoglio, M. (2000) J Am Societ Nephrol 11 Suppl 16, S140-3
43. Chawengsaksophak, K., James, R., Hammond, V. E., Kontgen, F., and
Beck, F. (1997) Nature 386(6620), 84-7
44. Pontoglio, M., Barra, J., Hadchouel, M., Doyen, A., Kress, C., Bach, J. P.,
Babinet, C., and Yaniv, M. (1996) Cell 84(4), 575-85
45. Park, J., Schulz, S., and Waldman, S. A. (2000)
Gastroenterology 119(1), 89-96
46. Suh, E., Wang, Z., Swain, G. P., Tenniswood, M., and Traber, P. G. (2001)
Am J Physiol 280(1), G149-56
47. Xu, F., Li, H., and Jin, T. (1999) J Biol Chem 274(48), 34310-6.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 34
48. Whyte, D. A., Li, C., Thomson, R. B., Nix, S. L., Zanjani, R., Karp, S. L.,
Aronson, P. S., and Igarashi, P. (1999) Am J Physiol 277(4 Pt 2), F587-98
49. Mendel, D. B., Khavari, P. A., Conley, P. B., Graves, M. K., Hansen, L. P.,
Admon, A., and Crabtree, G. R. (1991) Science 254(5039), 1762-7
50. Wu, G. D., Chen, L., Forslund, K., and Traber, P. G. (1994)
J Biol Chem 269(25), 17080-5
51. Krasinski, S. D., Van Wering, H. M., Tannemaat, M. R., and Grand, R. J.
(2001) Am J Physiol 281(1), G69-84
52. Hu, C., and Perlmutter, D. H. (1999) Am J Physiol 276(5 Pt 1), G1181-94
53. Akiyama, T. E., Ward, J. M., and Gonzalez, F. J. (2000)
J Biol Chem 275(35), 27117-22
54. Rings, E. H., Boudreau, F., Taylor, J. K., Moffett, J., Suh, E. R., and
Traber, P. G. (2001) Gastroenterology 121(6), 1437-50.
55. Dukes, I. D., Sreenan, S., Roe, M. W., Levisetti, M., Zhou, Y. P., Ostrega, D.,
Bell, G. I., Pontoglio, M., Yaniv, M., Philipson, L., and Polonsky, K. S. (1998)
J Biol Chem 273(38), 24457-64
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 35
Legends
Figure 1. Sequence analysis of the 5’-flanking region of the human and mouse
claudin-2 gene. The nucleotides differing between two species are highlighted in
gray. Potential binding motifs are underlined. The regions used for the reporter
constructs are indicated by the arrows and the corresponding basepair (bp)
numbers.
Figure 2. Analysis of human claudin-2 promoter deletion constructs.
The reporter constructs containing sequentially deleted 5’-flanking fragments
were prepared and transfected into Caco-2, HepG2 and NIHT3T cells as
described in Materials and Methods. Results are expressed as relative luciferase
activity of three different experiments carried out in triplicate (mean ± SD). The
mean value of cells transfected with null pGL3B vector was set to 1.
Figure 3. Mutational analysis of the Cdx binding sites within the human claudin-
2 promoter. A, Sequences of wildtype and mutated Cdx consensus motifs. Cdx
consensus sequences are underlined and mutations are in bold. B, Reporter
gene assay. Transfection into Caco-2 cells and luciferase assay were done.
Results are expressed as relative luciferase activity of three different experiments
carried out in triplicate (mean ± SD). The mean value of cells transfected with
null pGL3B vector was set to 1.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 36
Figure 4. Cdx1- and Cdx2-overexpression activates the human claudin-2
promoter. A, Western blot analysis of nuclear Cdx1 and Cdx2 protein expression
in Caco-2 cells. Caco-2 cells were transfected with Cdx1- and/or Cdx2-
expression vectors and 10µg of nuclear proteins were analyzed. B and C,
Reporter gene analysis. Caco-2 and NIH3T3 cells were transfected with Cdx1-
and/or Cdx2-expression vectors in the presence of the indicated reporter
constructs and luciferase assays were performed. Results are expressed as
relative luciferase activity of three different experiments carried out in triplicate
(mean ± SD). The mean value of cells transfected with pGL3B vector in the
absence of expression vectors was set to 1.
Figure 5. Interaction of Cdx2 with the Cdx binding sites of the human claudin-2
promoter. A, Preferential binding of Cdx2 to the upstream Cdx site. EMSA was
performed using 4 µg of nuclear proteins from post-confluent Caco-2 cells.
Competitions were done with 100-fold excess of indicated oligonucleotides.
Supershift assays were done by addition of 1µl of either anti-Cdx1 or anti-Cdx2
antibody. The sequences of oligonucleotides (Wt, Mut 1, Mut 2, and Mut 1+2)
are given in Figure 3A. The DNA-protein complexes (A, B, and C) and the
supershifted bands are indicated by arrows and white arrowheads, respectively.
B, Concentration-dependent interaction between Cdx consensus sites and Cdx2.
Nuclear proteins from Cdx2-transfected Caco-2 cells were incubated with the
indicated labeled probes. DNA-protein complexes are indicated by black arrows.
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 37
The white arrow in lane 7 indicates the complex A. The supershifted bands in the
presence of anti-Cdx2 antibody are indicated by white arrowheads.
Figure 6. The effects of HNF-1α- and HNF-1β-overexpression on the human
claudin-2 promoter. A, Reporter gene analysis. Caco-2 cells were transfected
with 0.5 µg of indicated expression vectors in the presence of reporter constructs,
and luciferase assay was performed. Results are expressed as relative
luciferase activity of three different experiments carried out in triplicate (mean ±
SD). The mean value of cells transfected with pGL3B in the absence of
expression vectors was set at 1. B, Western blot analysis of Cdx2 and HNF-1α
proteins in the cytosol and the nuclear protein fractions. Caco-2 cells were
transfected with 0.5 µg of HNF-1α- and/or Cdx2-expression vectors. Two days
after transfection, cytosol and nuclear protein (NE) fractions were prepared.
Equal amounts of proteins (25 µg per lane) were analyzed.
Figure 7. Interactions between HNF-1α and HNF-1β with the HNF-1 binding site
in the human claudin-2 promoter. EMSA was performed using 4 µg of nuclear
proteins of Caco-2 cells transfected with the indicated vectors. Competitions
were done with a 100-fold excess of wild type (Wt) or mutant (Mut)
oligonucleotide. Supershift assays were done by addition of 1µl of anti-HNF-1α
or anti-HNF-1β antibody. The oligonucleotide sequences are given in Materials
and Methods. The specific DNA-protein complex is indicated by an arrow. The
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 38
supershifted bands by anti-HNF-1α and anti-HNF-1β antibodies are indicated by
black and white arrowheads, respectively.
Figure 8. Claudin-1 and claudin-2 expression in the liver and kidneys of HNF-1 α
deficient and wildtype mice. A, Northern blot analysis of claudin-1 and claudin-2
mRNAs in the liver and kidneys of HNF-1α deficient (-/-) and wild type littermates
(+/+). Total RNA (30 µg) was electrophoresed, transferred to a nylon membrane
and hybridized with the indicated probes. B, Immunostaining for claudin-2 protein
in the liver and kidneys of HNF-1α deficient (-/-) and wild type littermates (+/+).
Arrows indicate claudin-2 expression in tight junctions. CV, central vein of the
liver.
Figure 9. Claudin-1 and claudin-2 expression in the small intestine of HNF-1α
deficient and wildtype mice. A, Northern blot analysis of claudin-1, claudin-2,
Cdx1, Cdx2, and HNF-1α mRNA expression in the jejunum and ileum of HNF-1α
deficient (-/-) and wildtype (+/+) mice. B, Densitometric analysis of claudin-1
(open bars) and claudin-2 (black bars) mRNA expression in the presence or
absence of HNF-1α gene. Expression levels of claudin-1 and claudin-2 mRNAs
were normalized for GAPDH mRNA levels in the same RNA isolations and
expressed as relative density per area (mean ± SD, n = 3). C, Immunostaining of
claudin-2 and claudin-1 protein in the ileum of HNF-1α deficient (-/-) and wildtype
(+/+) mice. Frozen sections were stained with either anti-claudin-2 (panel A, B, F
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Sakaguchi et al. 39
and G) or anti-claudin-1 (panel C, D, H and I) antibody and FITC-labeled anti-
rabbit secondary antibody. Panel E and J, control staining with rabbit serum and
secondary antibody. Arrows indicate stainings of claudins in tight junctions
(Original magnification in panel A-E 40x, in panel F-J 100 x).
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from

Hans-Christian ReineckerTakanori Sakaguchi, Xiubin Gu, Heidi M. Golden, EunRan Suh, David B. Rhoads and
αproteins and hepatocyte nuclear factor-1with conserved binding sites in mouse and human for caudal-related homeodomain Cloning of the human claudin-2 5'-flanking region revealed a TATA-less promoter
published online April 4, 2002J. Biol. Chem.
10.1074/jbc.M110261200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on June 15, 2018http://w
ww
.jbc.org/D
ownloaded from