Embed Size (px)
Citation preview

Molec. gen. Genet. 177, 527 533 (1980) © by Springer-Verlag 1980
Cloning of DNA of the rpoBC Operon from the Chromosome of Escherichia coli K12
Andrew Newman and Richard S. Hayward Department of Molecular Biology, University of Edinburgh, Kings Buildings, Edinburgh EH9 3JR, Scotland
Summary. We provide evidence that, in terms of tran- scriptional organisation, the rpoBC operon carried by )tr/ff 18 accurately represents the corresponding re- gion of the E. coli K12 chromosome.
A restriction fragment of E. coli K12 chromoso- mal D N A carrying the genes rpoBC (encoding the fi and/V subunits of R N A polymerase) and rplL (cod- ing for r ibosomal proteins L7/L12) was cloned in a 2 vector, and the resulting phage tested for gene expression. In common with the corresponding frag- ment of 2rifa18 DNA, the chromosomal fragment has no strong promoter for rplL or rpoBC transcrip- tion. Another new phage was constructed by adding, to the restriction fragment carrying the rplL rpoBC structural genes from 2r/ff18, a sequence from the E. coli K12 chromosome which includes a promoter for these genes. As in 2 r ~ 1 8 itself, this promoter is shared with rplJ but not with rplKA.
The properties of the latter phage also show that the dominant rifampicin-resistance characteristic of )oriffl8 results from more than one mutation.
Introduction
The genes rpoBC, encoding the fi and fi' subunits of R N A polymerase (EC 2.7.7.6) in E. coli, lie togeth- er beside the ribosomal protein genes rplKAJL in the bacterial D N A carried by the defective transduc- ing phage 2r/f~18 (Lindahl et al., 1975; Lindahl et al., 1977). Several recent studies on the regulation of R N A polymerase synthesis have concentrated on de- termining the arrangement of units of transcription in 2rlf~18 D N A (Yamamoto and Nomura , 1978; Linn and Scaife, 1978; Newman et al., 1979; Post et al., 1979; Fiil et al., 1979). The simplest interpretation
Offprint requests to : Dr Richard Hayward
of the results is that the rpoBC genes on 2riff18 are co-transcribed with rplL and rplJ, as distal elements in a single operon, thus helping the cell to coordinate the production of its transcriptional and translational machineries.
All the above studies were performed using mu- tant D N A : 2rlf d 18 was isolated by virtue of its domi- nant expression of rifampicin-resistance (Kirschbaum and Konrad, 1973). The basis of the dominance of the rpoB allele carried by 2rif a 18 is obscure, and might involve coupling of a normal drug-resistance muta- tion in rpoB with a positive cis-regulatory alteration : the evidence on this point is contradictory (discussed in Newman et al., 1979). Accordingly, the transcrip- tional organisation of the rpoBC operon in 2rzfa18 might be anomalous. Furthermore, the generation of transducing phages is sometimes accompanied by structural alteration of the bacterial sequences (Rossi et al., 1979), as presumably in the cases of the related phages 2riff12 and ~b80 r ~ (Lindahl et al., 1977).
We therefore undertook to isolate the rpoBC genes and their regulatory elements directly from the E. coli K12 chromosome, avoiding any selection for their expression, and to compare their structure and func- tion with those of the sequences derived from 2r/fal 8.
Materials and Methods
Bacterial strains used are listed in Table 1. AJN10 was constructed in the same way as AJN1 (Newman et al., 1979) but using an isogenic rcs + starting strain. AJN20 is the source strain for phage carrying extensions of the bacterial DNA fragment present in AN2172 (Fig. 1) and was constructed as follows. The starting strain was a rifampicin-sensitive Arg + Pur + Rcs + Pl-transductant of X174 (Hayward, 1976). A Mal ÷ derivative was obtained by trans- duction with Pivot, purified, and checked for 2-sensitivity. It was then lysogenised with 2172 to produce a derivative carrying the prophage integrated at the chromosomal rpoBC region. ()A72 is integration-defective and, in the absence of helper, can only inte- grate into the E. coli chromosome by Rec-mediated recombina-
0026-8925/80/0177/0527/$01.40

528 A. Newman and R.S. Hayward: Cloning of E. coli rpoBC Operon
Table 1. Bacterial strains
Strain Relevant details Reference Source
C600 P2 lysogen Zissler et al., (P2) 1971
CR63 Rif-sensitive Appleyard, prototroph 1954
S159 uvrA Jaskunas et al., 1975
S159 uvrA 2ind lysogen Jaskunas et al., ().ind ) 1975
S159 uvrA 2imm21 lysogen Jaskunas et al., (2imm21 ) 1975
AJN1 Rif-sensitive Newman et al., rcs-40 recA56 m e t B 1979
AJN10 Rif-sensitive This work rcs + recA56 m e t B
AJN5 rpoB70 recA 1 m e t B This work
AJN20 rcs + recA56: This work carries AN2172 integrated into rpoB+ C + : Rif-R
N. Murray
N. Murray
M. Nomura
M. Nomura
T. Linn
tion.) A proportion of such lysogens were rifampicin-resistant. Indi- vidual Rif-R clones were purified, and made his + recA56 as de- scribed (Hayward, 1976). Hence AJN20 is a m e t B recA56 thi str mal + rcs + lysogen of AN2172.
AJN5 is a spontaneous Mal + derivative of E. coli strain AJ7 (Boyd et al., 1974).
The phage strains and methods used were as described previ- ously (Newman et al., 1979) or as detailed later, except for the following: electroelution of DNA fragments from agarose, as well as agarose gel electrophoresis of DNA, was carried out as in McDonell et al. (1977); cloning in pBR322 was as suggested by Bolivar et al. (1977); DNA was 32p-labelled by nick-translation essentially as described by Rigby et al. (1977) ; the plaque-hybridisa- tion technique was that of Benton and Davis (1977) ; and polyacryl- amide gradient gel analyses of DNA restriction fragments were based on Maniatis et al., (1975), with modifications by Boothroyd (1979).
Results and Interpretation
Isolation and Characterisation of rpoBC Genes from the E. coli Chromosome
A 'pool ' of hybrid phage produced by the ligation in vitro of mixed endoR. HindlII-digested DNA from E. coli K12 CR63 and NM4761 was kindly provided
by Dr N. Murray. It was screened for plaques hybridising to 32p-labelled DNA of the plasmid pAN219. This plasmid is a pBR322-derivative carry- ing an 11 kb HindIII fragment of 4rif a 18 DNA which includes the genes rpoBC, rpIL and part of rplJ (New- man et al., 1979; Post et al., 1979). DNA prepared from several phage isolates contained an 11 kb frag- ment of CR63 DNA. One such phage, AN263, was purified for further study. DNA restriction analyses (not shown) revealed that the HindIII fragment cloned in 463 has the same size and contains the same internal EcoRI targets as the 11 kb HindIII frag- ment of 4rif a18, which we had previously cloned in NM4761 to produce the phage designated AN417 (Newman et al., 1979; see also Fig. 1 : 4172 is a deriva- tive of 417, with an identical bacterial DNA segment). Are the transcriptional properties of the bacterial DNA carried by 463 and 417 also similar?
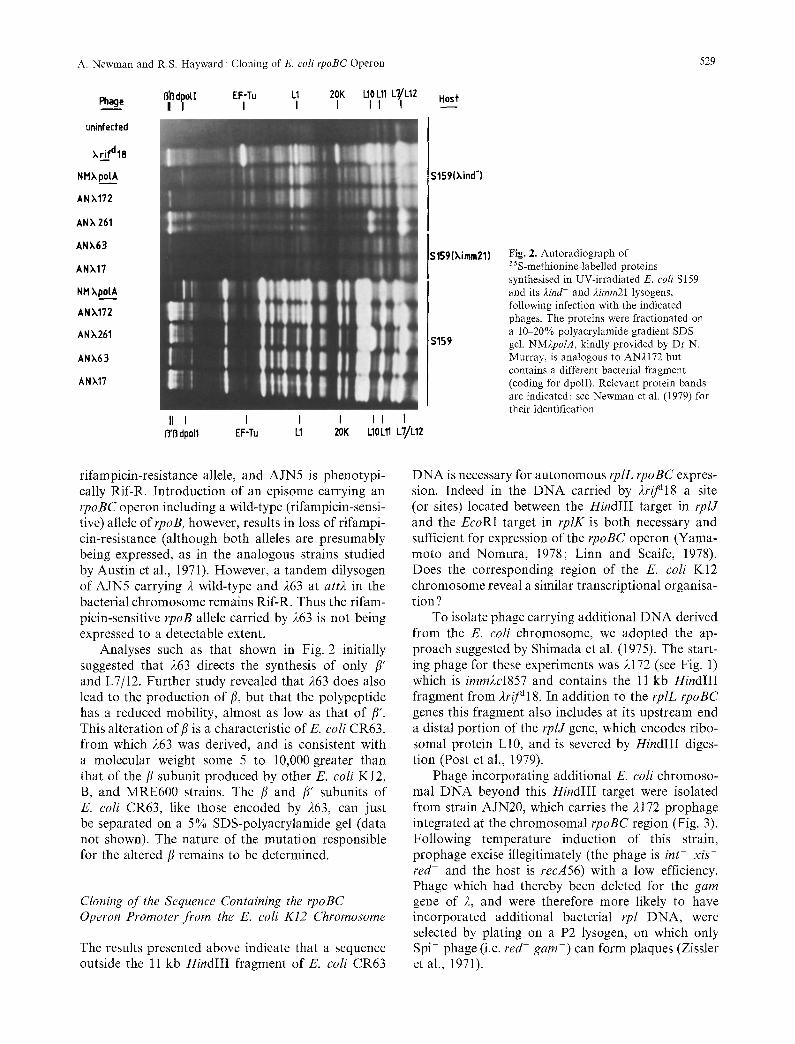
The bacterial genes carried by these phages can be analysed by aSS-methionine pulse-labelling of the proteins synthesised after infection of UV-irradiated E. coli S159, followed by SDS-polyacrylamide gra- dient gel electrophoresis, and autoradiography of the dried gel. Furthermore, synthesis of bacterial gene products in UV irradiated lysogenic E. coli is diagnos- tic of transcription from bacterial promoters within the cloned fragment, since all phage promoters which might serve the cloned genes are then repressed.
After infection of the non-lysogenic strain S159, 4PL is active, and both 417 and 263 direct the vigorous synthesis of/~, /~' and L7/12. In contrast infection of the S159 (4imm21) lysogen by these phages elicits a very weak synthesis of/3 and/~', while L7/12 syn- thesis is not detectably increased above background levels (see Fig. 2). We conclude that in the chromo- some ofE. coli CR63, as in the bacterial DNA carried by 4r/J~18, the transcription of rpoBC and rplL is effectively dependent upon sequences lying outside the cloned HindIII fragment. Although one or more promoters allowing expression of the rpo genes are present within the cloned DNA from either source, these function very weakly in the conditions tested.
We have confirmed that 463 does not carry the main promoter for the rpoBC operon, by constructing lysogens in a suitable strain. The K12 strain AJN5 is recA1 metB rpoB70 mal+: rpoB70 is a recessive
I A., I , , , 15o 1,.71 60
i i . bi .ol, l ,,.o
0"6
E. co[i genes: rpoC rp!
Transcription
Fig. 1. Genetic and physical map of phage AN2172, derived from Newman et al. (1979) and Hughes (1977). DNA fragment sizes are given in kilobase-pairs. The bacterial DNA segment is shown by the heavy line. The position and transcriptional orientation of the 2 promoter PL and of the E. coli genes are indicated
A. Newman and R.S. Hayward: Cloning of E. coli rpoBC Operon 529
Phage
uninfected
Xrifd18
NHX poi____A
ANX172
ANX261
ANX63
ANX17
NH XpoL_._AA
AN)d72
AN),261
ANX63
AN)d7
13'fl dpotI EF-Tu L1 II I I I
20K LIO Lll L?( L12 I I I Hos.~.t
II I I I fl'ft dpo[1 EF-Tu 1.1
I I I I 20K LIO Lll LT/L12
S159(Xind')
J s159(Ximm211
S159
Fig. 2. Autoradiograph of 35 S-methionine-labelled proteins synthesised in UV-irradiated E. coli S159 and its )Jnd- and 2imm21 lysogens, following infection with the indicated phages. The proteins were fractionated on a 10-20% polyacrylamide gradient SDS gel. NM)~polA, kindly provided by Dr N. Murray, is analogous to AN2172 but contains a different bacterial fragment (coding for dpolI). Relevant protein bands are indicated: see Newman et al. (1979) for their identification
rifampicin-resistance allele, and AJN5 is phenotypi- cally Rif-R. Introduction of an episome carrying an rpoBC operon including a wild-type (rifampicin-sensi- tive) allele ofrpoB, however, results in loss of rifampi- cin-resistance (although both alleles are presumably being expressed, as in the analogous strains studied by Austin et al., 1971). However, a tandem dilysogen of AJN5 carrying 2 wild-type and 263 at att2 in the bacterial chromosome remains Rif-R. Thus the rifam- picin-sensitive rpoB allele carried by 263 is not being expressed to a detectable extent.
Analyses such as that shown in Fig. 2 initially suggested that 263 directs the synthesis of only ]3' and L7/12. Further study revealed that 263 does also lead to the production of/~, but that the polypeptide has a reduced mobility, almost as low as that of/~'. This alteration of{/is a characteristic of E. coli CR63, from which 263 was derived, and is consistent with a molecular weight some 5 to 10,000 greater than that of the/~ subunit produced by other E. coli K12, B, and MRE600 strains. The /~ and /~' subunits of E. coli CR63, like those encoded by 263, can just be separated on a 5% SDS-polyacrylamide gel (data not shown). The nature of the mutation responsible for the altered/~ remains to be determined.
Cloning of the Sequence Containing the rpoBC Operon Promoter from the E. coli K12 Chromosome
The results presented above indicate that a sequence outside the 11 kb HindIII fragment of E. coli CR63
DNA is necessary for autonomous rplL rpoBC expres- sion. Indeed in the DNA carried by 2rlf18 a site (or sites) located between the HindIII target in rplJ and the EcoRI target in rplK is both necessary and sufficient for expression of the rpoBC operon (Yama- moto and Nomura, 1978; Linn and Scaife, 1978). Does the corresponding region of the E. coli K12 chromosome reveal a similar transcriptional organisa- tion?
To isolate phage carrying additional DNA derived from the E. coli chromosome, we adopted the ap- proach suggested by Shimada et al. (1975). The start- ing phage for these experiments was 2172 (see Fig. 1) which is imm2ci857 and contains the 11 kb HindIII fragment from 2rzfal8. In addition to the rplL rpoBC genes this fragment also includes at its upstream end a distal portion of the rp/J gene, which encodes ribo- somal protein L10, and is severed by HindIlI diges- tion (Post et al., 1979).
Phage incorporating additional E. coli chromoso- mal DNA beyond this HindIII target were isolated from strain AJN20, which carries the 2172 prophage integrated at the chromosomal rpoBC region (Fig. 3). Following temperature induction of this strain, prophage excise illegitimately (the phage is int- xis- red- and the host is recA56) with a low efficiency. Phage which had thereby been deleted for the gain gene of 2, and were therefore more likely to have incorporated additional bacterial rpl DNA, were selected by plating on a P2 lysogen, on which only Spi phage (i.e. red- gain-) can form plaques (Zissler et al., 1971).

530 A. Newman and R.S. Hayward: Cloning of E. coli rpoBC Operon
rp[ rif r K A JL rp~)B rpoC J
' ' / I I I | I I I
I I I I I I pAN38
rpl rif s A NgamL , r~oB
I I I
M72
i[[egifirnafe [ excision
, , rpoE
I t, l 11.1 01-1 ,0
[ e o., 1 i - [ i , t = ,o I HindTTr 23.z~ ] 110 5"5 I I 8"g
0-6 0'I
E.coli genes: rpoC rpoB LJA I rp[
rifr Fig. 3. Origin of phage AN2261, together with its genetic and restriction map. The relevant region of the chromosome of AJN20, containing )A72 ( in t - x i s - r e d A - B gain + cI857 ts) lysogenised via Rec into E. coli rec + rpoB + (Rif-S), is illustrated at the top. Open boxes represent chromosomal DNA, while filled boxes and lines represent respectively bacterial and lambda DNA of 2172. The extent of DNA carried by plasmid pAN38 is also indicated. The genetic and restriction map of phage 261, determined as described in the text, is given below. The source of the cloned bacterial DNA, where known, is illustrated by filled or open boxes: where unknown, by broken lines. DNA fragment sizes are given in kilobase-pairs
The Spi phage were then screened for plaque-hy- bridisation to 32p-labelled DNA of the plasmid pAN38 (constructed and kindly provided by Dr A. Nicolaidis). pAN38 is a derivative of pBR322 carrying a 1.9 kb fragment of 2r/f~18 DNA, extending from the HindIII site in rpU to the EcoRI site in rplK (see Fig. 4). Phage from a number of plaques which hybridised to pAN38 DNA were analysed by diges- tion of their DNA with restriction enzymes. One of these, designated AN2261, was purified for further study.
Digestion of DNA from 2172 and its derivative 2261 with EcoRI or HindIII reveals no major change in length of the restriction fragments obtained. After digestion with AvaI, however, 2261 DNA produced fragments of 12.6 kb and 1.8 kb in place of the 14.4 kb fragment produced from 2172 DNA (data not shown). From these results we can conclude that: (i) the additional bacterial DNA cloned in 2261 con- tains no sites for EcoRI or HindIII; (ii) the length of bacterial DNA added by illegitimate excision of the prophage in generating 2261 was closely matched by the length of 2 DNA lost; (iii) there is an AvaI target in the additional bacterial DNA. Note that there is an AvaI site in the corresponding region of 2r/fdl8 DNA, some 170 bp upstream of the HindIII site in rpU (Post et al., 1979: and see Fig. 4).
To define more accurately how much extra E. coli DNA has been cloned in 2261 (and to test its
structural identity with the sequences present in 2rifal8 DNA) the 8.0 kb EcoRI fragments of 2172 and 2261 DNA, which contain the region of interest, were purified by electroelution from a preparative agarose gel. The 1.9 kb fragment of 2rifal8 DNA was likewise prepared from an EcoRI/HindIII double digest of pAN38 DNA. All 3 fragments were then digested with endoR.HaelII, and analysed on a 3-10% polyacrylamide gradient gel alongside markers of defined size (Fig. 5). It is apparent that HaeIII- digestion of the 2261 8 kb fragment gives rise to new sub-fragments not present in the corresponding 2172 8 kb fragment digest. Significantly, new fragments of 237 bp, 207 bp, 72 bp and 166 bp appear, co-migrat- ing with bands produced from the 1.9 kb fragment of 2rifq8. Similar studies with EndoR.HhaI (data not shown) revealed the presence of several new bands in the 2261 DNA digest, co-migrating with bands derived from the 1.9 kb fragment of 2r/fall8.
Hence we have established a degree of structural similarity between the additional DNA cloned in 2261, derived from the chromosome of a rifampicin- sensitive E. coli, and the corresponding region of 2r/fal8. Furthermore, we deduce that the bacterial DNA in 2261 extends at least as far as the HaeIII site within rplA, at position 812 in the nucleotide sequence determined by Post etal. (1979): see Fig. 4. However, the HhaI mapping reveals that 2261 does not carry a complete copy of rpIA,

A. Newman and R.S. Hayward: Cloning of E. coli rpoBC Operon 531
Hap of rp..~[ genes in Xrifd18
EcoRI EcoRI 1 1 rptL I---I rplJ I [ rp[A H rplK
HaeTTr restriction map Hindllr AvaI EcoRI °f HindTTT-Ec°RI (l'9Kb) t f r a g m e n t of ~' t Xri._~18 622 II 237 I Z07 [72111 166 J 230 J ~01
EcoRI HindTIT X172 ~[I J XDNA
X261
EcoRI Hindll Hhal Hae~ Hhal
AvaI~ (876) ~121 1~21
ii [ l l[i ~ - - XDNA
Fig. 4. Extent of additional DNA cloned in phage261. The rpl region of 2rijm18, which is transcribed from right to left, is drawn to scale. The HaeIII subfragments of a HindIII/EcoRI-generated fragment from this region are shown, with their sizes given in basepairs. The requisite information is derived from Post et al. (1979). The extent of the bacterial DNA present in phages 172 and 261 is also shown, based on experiments described in the text. Only those HhaI sites which serve to define the endpoint of the bacterial DNA in 2261 are shown. Their numbered positions within the sequence of Post et al., are given in brackets
since its DNA stops short of the HhaI site at position 622.
Does the additional E. coli DNA in 2261 include a site allowing independent expression of the rpoBC operon? Figure 2 shows the results of an experiment designed to answer this question, involving infection of UV-irradiated E. coli S159 and its 2ind lysogen.
2172 directs the synthesis of L7/12,/~ and/~' when S159 is the host, but in S159 (2ind-) only a very weak synthesis of/~ and /~' is observed, and L7/12 synthesis is not detectably different from background. Thus in 2172 (as in 217) expression of the cloned rplL rpoBC genes is effectively dependent on the 2P L promoter (see Fig. 1).
In contrast, 2261 directs the synthesis of/~, /7', L7/12, and an additional protein co-migrating with L10 (the product of gene rpIJ) in both lysogenic and non-lysogenic hosts. The additional DNA cloned in 2261 must therefore include a site (or sites) capable of initiating transcription of all 4 genes. The simplest interpretation of this is that rpoBC are being co- transcribed with rplJL, presumably from a promoter located between rplJ and rplA. As expected, synthesis of protein L1 is not detectable in either host.
We have also constructed tandem dilysogens of 2261 (and of 2172) with 2imm21, the latter providing necessary integration functions, in the isogenic Rif-S, recA56 strains AJN1 and AJN10 (Table 1). AJN1 carries a mutation, rcs-40, which reverses the normal dominance of rpoB + (Rif-S) over rpoB (Rif-R) alleles in heterodiploids. Accordingly an AJN1/KLF10 rpoB70 merodiploid is phenotypically Rif-R, whereas the isogenic rcs + merodiploid (AJN10/KLF10 rpoB70) is Rif-S : Table 2. The construction of lyso- gens in both strains permits a test for dominance,
Table 2. Rif-phenotype of rpoBC-merodiploids
Host AJN10 (rcs +) AJN1 (rcs-40)
Phage or episome present
KLF10 rpoB70 S R )orij~18, 2imm21 R R )~172, ;timm21 S S k261, kirnm21 S R
as well as for expression in vivo, of the rpoB (Rif-R) genes carried by the phages.
The 2261, 2imm21 dilysogen of AJN1 (rcs-40) is Rif-R, growing normally at a rifampicin concentration of 200 gg/ml (Table 2). The 2172, 2imm21 derivatives of both AJN1 and AJN10 are Rif-S, while the 2rt.'] ~ 18 analogues are, as expected, Rif-R. Curing of 2261 and of 2rtf~18 from the above strains, by selection of survivors at 42 ° C, restores rifampicin-sensitivity. The survivors retain imm21, but are as expected irnm2- and 2-sensitive.
The expression of rifampicin-resistance by the )~261 prophage in strain AJN1 confirms that the E. coli DNA which it carries includes a bacterial pro- moter from which the rpoBC operon is strongly expressed in vivo: whereas 2172 and, as shown above, 263 contain no such promoter.
Surprisingly, the 2261, 2imm21 derivative of the rcs + strain AJN10 is Rif-S. This is discussed below.
Discussion
This analysis of rpoBC operon DNA cloned directly from the E. coli chromosome confirms the major con-

532 A. Newman and R.S. Hayward: Cloning of E. coli rpoBC Operon
1 2 3 4 5 Sizes (bp)
--622
-301
-- 237230 --207
--166
- -72
Fig. 5. DNA restriction analyses. The samples were fractionated on a 3-10% polyacrylamide gradient gel (modified from Maniatis et al., 1975), which was stained with ethidium bromide and pho- tographed under UV. DNA samples: 1, pBR322 cut with HpaII; 2, EcoRI (rpl-2) 8 kb fragment of 2172, recut with HaeIII; 3, as 2, but DNA from 2261 ; 4, HindIII-EcoRI (rplJAK) 1.9 kb frag- ment of 2r/fal8, recut with HaeIII; 5, pBR322 cut with HaeIII. Tracks 1 and 5 provide fragments of defined size (Sutcliffe, 1978). The sizes of the HaelII fragments in track 4 (derived from Post et al., 1979) are indicated on the right
clusion derived f rom experiments using 2rif a 18 D N A (cited in the In t roduct ion) : the rpoBC genes are co- transcribed, under normal growth condit ions in vivo, with the r ibosomal protein genes rplL and (probably) rplJ. In agreement with our results, Y a m a m o t o and N o m u r a (1979) showed that an EcoRI f ragment o f the E. coIi chromosomal D N A includes a p romoter for the rpU gene but not for the rplA gene which it carries.
The 2261, 2imm21 derivative o f the rcs + strain A J N 1 0 is Rif-S, despite the fact that the rpoB gene on 2261 should presumably be expressed (as in AJN1) and should carry the rifampicin-resistance muta t ion derived f rom 2r0~18; the chances of our having picked up a spontaneous muta t ion are remote (about 1 in 108), since we avoided any selection for rifampicin-resistance. Our interpretat ion o f this find- ing is that the Rif -D phenotype has a complex genetic basis. A rifampicin-resistance componen t has been recovered in 2261, but has been separated f rom some other feature which conferred dominance in ,~rzf~18. The present results do not reveal whether the domi- nance componen t lies outside rpoB, or within it (on the amino-terminal side o f the resistance mutat ion). In either case it might, for example, effectively in- crease the expression of the rpoB (Rif-R) allele in heterodiploids. We infer f rom the results o f Linn and Scaife (1978) that any relevant external muta t ion must lie between rpoB and the EcoRI site in rplK.
Our results suggest that the weak promoter (or promoters) for rpoB and rpoC, which we originally noted in the HindIII rplL-rpoBC fragment o f 2ri f ~ 18 D N A (Newman et al., 1979), are also present in the corresponding fragment o f E. coli CR63 D N A . We speculate that such weak promoters , whether consti- tutive or inducible, might contr ibute significantly to the expression of the polymerase genes during starva- tion, or under growth condit ions differing f rom those which we have so far tested. For example, activation of a normal ly weak independent p romoter for rpoC might explain some puzzling observations on the syn- thesis of fl and fl' during recovery o f E. coli f rom rifampicin t reatment (Nakamura and Yura, 1976).
Acknowledgements. We thank Drs J. Boothroyd, N. Murray and A. Nicolaidis and Ms K. Brown for advice and help, and Drs N. Murray and M. Smith for reading the manuscript. Rifampicin was a gift from Lepetit. The work was supported by a Project Grant and a Research Training Award (A.N.) from the Medical Research Council.
References
Appleyard, R.K.: Segregation of new lysogenic types during growth of a doubly lysogenic strain derived from Escherichia coli K12. Genetics 39, 440-452 (1954)
Austin, S.J., Tittawella, I.P.B., Hayward, R.S., Scaife, J.G. : Amber mutations of Escherichia coli RNA polymerase. Nature New Biol. 232, 133-136 (1971)
Benton, W.D., Davis, R.W.: Screening 2gt recombinant clones by hybridisation to single plaques in situ. Science 196, 180 182 (1977)
Bolivar, F., Rodriguez, R.L., Green, P.J., Betlach, M.G., Heynecker, H.L., Boyer, H.W., Crosa, J.H., Falkow, S. : Con- struction and characterisation of new cloning vehicles. II. A multipurpose cloning system. Gene 2, 95-113 (1977)

A. Newman and R.S. Hayward: Cloning of E. coli rpoBC Operon 533
Boothroyd, J.C. : PhD Thesis, University of Edinburgh (1979) Boyd, D., Zillig, W., Scaife, J.: Reference mutations for the fl
sub-unit of RNA polymerase. Mol. Gen. Genet. 130, 315 320 (1974)
Fiil, N.P., Bendiak, D., Collins, J., Friesen, J.D.: Expression of Escherichia coli ribosomal protein and RNA polymerase genes cloned on plasmids. Mol. Gen. Genet. 173, 39-50 (1979)
Hayward, R.S.: DNA blockade by rifampicin-inactivated E. coli RNA polymerase, and its amelioration by a specific mutation. Eur. J. Biochem. 71, 19 24 (1976)
Hughes, S.G. : A map of the cleavage sites for endonuclease AvaI in the chromosome of bacteriophage lambda. Biochem. J. 163, 503 509 (1977)
Jaskunas, S.R., Lindahl, L., Nomura, M., Burgess, R.R. : Identifi- cation of two copies of the gene for the elongation factor EF-Tu in E. coli. Nature 257, 458-462 (1975)
Kirschbaum, J.B., Konrad, E.B. : Isolation of a specialised lambda transducing bacteriophage carrying the beta subunit gene for Escherichia coil ribonucleic acid polymerase. J. Bacteriol. 116, 517 526 (1973)
Lindahl, L., Jaskunas, S.R., Dennis, P.P., Nomura, M.: Cluster of genes in Escherichia coli for ribosomal proteins, ribosomal RNA, and RNA polymerase subunits. Proc. Natl. Acad, Sci. U.S.A. 72, 2743-2747 (1975)
Lindahl, L., Yamamoto, M., Nomura, M., Kirschbaum, J.B., Allet, B., Rochaix, J.-D.: Mapping of a cluster of genes for components of the transcriptional and translational mechanisms of Escherichia coli. J. Mol. Biol. 109, 23 47 (1977)
Linn, T., Scaife, J.G. : Identification of a single promoter in E. coli for rplL and rpoBC. Nature 275, 33-37 (1978)
Maniatis, T., Jeffrey, A., van de Sande, H. : Chain length determi- nation of small double- and single-stranded DNA molecules by polyacrylamide gel electrophoresis. Biochemistry 14, 3787 3794 (1975)
McDonell, M.W., Simon, M.N., Studier, F.W. : Analysis of restric- tion fragments of T7 DNA and determination of molecular weights by electrophoresis in neutral and alkaline gels. J. Mol. Biol. 110, 119-146 (1977)
Nakamura, Y., Yura, T.: Effects of rifampicin on synthesis and functional activity of DNA-dependent RNA polymerase in Es- cherichia coll. Mol. Gen. Genet. 145, 227537 (1976)
Newman, A.J., Linn, T.G., Hayward, R.S. : Evidence for co-tran- scription of the RNA polymerase genes rpoBC with a ribosomal protein gene of Escherichia coli. Mol. Gen. Genet. 169, 195-204 (I979)
Post, L.E., Strycharz, G.D., Nomura, M., Lewis, H., Dennis, P.P. : Nucleotide sequence of the ribosomal protein gene cluster ad- jacent to the geue for RNA polymerase subunit/? in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 76, 1697 1701 (1979)
Rigby, P.W.J., Dieckmann, M., Rhodes, C., Berg, P.: Labelling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J. Mol. Biol. 113, 237-251 (1977)
Rossi, J.J., Ross, W., Egan, J., Lipman, D.J., Landy, A. : Structural organisation of Escherichia coli tRNA :ryr gene clusters in four different transducing bacteriophage. J. Mol. Biol. 128, 21-47 (1979)
Schweitzer, S.M., Matzura, H. :Transformation of Escherichia coli by a specific DNA restriction fragment. Mol. Gen. Genet. 155, 213-217 (1977)
Shimada, K., Weisberg, R.A., Gottesman, M.E. : Prophage lambda at unusual chromosomal locations. III. The components o~ the secondary attachment sites. J. Mol. Biol. 93, 415-429 (197.5)
Sutcliffe, J.G.: pBR322 restriction map derived from the DNA sequence: accurate DNA size markers up to 4361 nucleotide pairs long. Nucleic Acids Res. 5, 272i 2728 (1978)
Yamamoto, M., Nomura, M. : Co-transcription of genes for RNA polymerase sub-units fl and fl' with genes for ribosomal proteins in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 75, 3891-3895 (1978)
Yamamoto, M., Nomura, M. : Organisation of genes for transcrip- tion and translation in the rlf region of the Escherichia coli chromosome. J. Bacteriol. 137, 584 594 (1979)
Zissler, J., Signer, E., Schaefer, F.: The role of recombination in growth of phage 2. II. Inhibition of growth by prophage P2. In: The bacteriophage lambda (A.D. Hershey, ed.). New York: Cold Spring Harbor Laboratory 1971
C o m m u n i c a t e d b y A. B 6 c k
Received September 16/November 5, 1979





























