Embed Size (px)
Citation preview

R E S E A R C H L E T T E R
Cloning, characterizationand expressionanalysis of nucleotidemetabolism-relatedgenesofmycobacteriophageL5Bidisha Bhattacharya, Nabanita Giri, Mahasweta Mitra & Sujoy K. Das Gupta
Department of Microbiology, Bose Institute, Kolkata, India
Correspondence: Sujoy K. Das Gupta,
Department of Microbiology, Bose Institute,
P1/12 C.I.T. Scheme VIIM, Kolkata 700054,
India. Tel.: 191 33 23559416; fax: 191 33
2553886; e-mail: [email protected]
Received 24 August 2007; accepted
25 November 2007.
First published online February 2008.
DOI:10.1111/j.1574-6968.2007.01047.x
Editor: Dieter Jahn
Keywords
multienzyme complex; flavin-dependent
thymidylate synthase; phage infection.
Abstract
The genomes of mycobacteriophages of the L5 family, which includes the lytic phage
D29, contain several genes putatively linked to nucleotide-metabolizing functions.
Two such genes, 48 and 50, encoding thymidylate synthase and ribonucleotide
reductase (RNR), respectively, were overexpressed in Escherichia coli and the
recombinant proteins were biochemically characterized. It was established that
Gp50 was a class II RNR having properties similar to that of the corresponding
enzyme from Lactobacillus leichmanni, whereas Gp48 was a flavin-dependent
thymidylate synthase (ThyX) that resembled the Paramecium bursaria chlorella
virus-1 ThyX enzyme in its properties. That both these proteins play a role in phage
development was evident from the observation that they were detectable soon after
the lytic phase of growth commenced. Gp48 and 50 were also found to coimmuno-
precipitate, which indicates the possible existence of an L5 thymidylate synthase
complex. Thymidylate synthase assays revealed that during the intracellular stage of
phage growth, a significant decrease in the host thymidylate synthase (ThyA) activity
occurred. It appears that synthesis of the viral enzyme (ThyX) is necessary to
compensate for this loss in activity. In general, the results suggest that phage-
encoded nucleotide metabolism-related functions play an important role in the lytic
propagation of L5 and related mycobacteriophages.
Introduction
Mycobacteriophages have been considered to be corner-
stones of mycobacterial research (Hatfull & Jacobs, 1994).
Among the mycobacteriophages that have been well studied
are the closely related members of the L5 family, which
includes the lysogenic phages L1 (Doke, 1960; Lee et al.,
1991), L5 (Snapper et al., 1988; Hatfull & Sarkis, 1993) and
BxB1 (Jain & Hatfull, 2000) and the lytic phage D29 (Ford
et al., 1998). Although the L5 family of phages is considered
to be lysogenic or potentially lysogenic (D29) (Ribeiro et al.,
1997), they have features that strongly resemble lytic phages
(Hatfull & Sarkis, 1993). One such feature is the ability of
these phages to shut down host protein synthesis immedi-
ately following infection.
A large number of mycobacteriophage genome sequences
are currently available in the databases (Pedulla et al., 2003).
Comparative analysis of these sequences revealed that at
least half of the mycobacteriophage genes have no database
matches (NDMs), indicating thereby the possibility of the
existence of novel genes, whose functions are unknown.
Although several mycobacteriophage genome sequences are
available, only a limited number of genes have been char-
acterized, most of which belong to the L5 family. These are:
gene 71, the phage repressor (Donnelly-Wu et al., 1993); gene
33, integrase (Lee & Hatfull, 1993); and gene 36, excisionase
(Lewis & Hatfull, 2000). In addition, several other myco-
bacteriophage genes have been characterized, such as those
encoding a tape measure protein from TM4 (Piuri &
Hatfull, 2006), a polynucleotide kinase from Omega and
Cjw1 (Zhu et al., 2004) and a lysis protein from Ms6 (Garcia
et al., 2002).
In silico analysis (Hatfull & Sarkis, 1993; Ford et al., 1998)
of L5 and D29 genome sequences revealed the presence of
several genes that putatively code for enzymes involved in
DNA synthesis. One such gene (44) encodes a viral DNA
polymerase. Other DNA synthesis-related genes are 58, 59
and 65, which putatively encode primase, exonuclease and
FEMS Microbiol Lett 280 (2008) 64–72c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

helicase, respectively. Apart from genes encoding enzymes
related to DNA replication, there are several genes that
appear to code for nucleotide metabolism-related functions.
Two such genes are 48 (thymidylate synthase) and 50
[ribonucleotide reductase (RNR)]. In the case of D29, but
apparently not in the case of L5, there may be a gene for
dCMP deaminase also. All of these genes are clustered in the
central region of the phage genome, indicating that their
products have some related functions. If the entire scenario is
considered in its totality, it seems that, as in case of the lytic
phages such as T4, L5/D29 phage-encoded DNA polymeriza-
tion and deoxy-ribonucleotide synthesis-related enzymes act
in concert to meet the replicative demands of the viral DNA,
particularly during the early stage of infection.
In nature, RNR (Poole et al., 2002) accounts for the
acquisition of three (dCTP, dGTP and dATP) of the four
deoxyribonucleotides needed for DNA biosynthesis. The
fourth nucleotide, dTTP, is produced by two well-known
processes. Exogenous thymidine can be directly salvaged by
thymidine kinase or dTMP can be synthesized de novo from
dUMP, a reaction catalyzed by thymidylate synthase. Thy-
midylate synthases may be of two types (Myllykallio et al.,
2002) – the classical thymidylate synthase designated as
ThyA and the alternative class of thymidylate synthases
known as ThyX. Both these enzymes catalyze the conversion
of dUMP to dTMP, but the reductive mechanisms are
different. In ThyA-catalyzed reactions, CH2H4folate serves
both as a reductant and as a CH2– donor whereas in a ThyX-
catalyzed reaction CH2H4folate acts only as a CH2– donor,
reducing equivalents being donated by NADPH through the
intermediate reduction of the enzyme-bound FAD. The
protein encoded by L5 gene 48 contains the highly conserved
ThyX motif RHRX7S (Myllykallio et al., 2002). ThyX
proteins in general are found in many pathogenic bacteria
and it has been proposed that this protein is an attractive target
for developing drugs against such bacteria (Graziani et al.,
2004). Interestingly, mycobacteria possess both the versions,
ThyX as well as ThyA, but the significance of this is not clearly
understood (Myllykallio et al., 2002; Sampathkumar et al.,
2005).
Mycobacteriophage L5 gene 50 putatively codes for a class
II RNR (RNR II). RNRs of the class II family typically use
adenosylcobalamin as the cofactor for the generation of the
free radicals (Poole et al., 2002). It is generally considered
that Class II enzymes function under micro-aerophillic
conditions, whereas class I and III enzymes function under
aerobic and anaerobic conditions, respectively.
Enzymes catalyzing sequential metabolic reactions are
often organized in the form of multienzyme complexes.
Such multienzyme complexes also exist in case of dNTP
biosynthesis. Although the dNTP-synthesizing complexes
exist in several prokaryotic and eukaryotic systems, the
best-characterized system is the complex encoded by the
Escherichia coli lytic phage T4 (Tomich et al., 1974; Reddy
et al., 1977; Prem veer Reddy & Pardee, 1980; Allen et al.,
1983; Mathews, 1993). In case of T4, it has been demon-
strated that several phage-encoded nucleotide metabolism-
related proteins such as thymidylate synthase, dihydrofolate
reductase, ribonucleotide reductase, deoxycytidylate deami-
nase, dC/UTPase and deoxycytidylate hydroxymethylase form
a functional aggregate. It is believed that such an aggregate
acts as a ‘metabolome’, which plays an important role in
maintaining an adequate supply of deoxy-nucleotide precur-
sors for the rapid synthesis of viral DNA (Mathews, 1993).
The presence of genes for ThyX and RNR II in the
genome of L5 and related mycobacteriophages has been
predicted purely on the basis of in silico analysis. No
functional characterization of these proteins has been car-
ried out so far. The present work was thus initiated with the
dual objective of characterizing the translated products of
genes 48 and 50 and analyzing their expression status during
infection. The results, apart from confirming bio-chemically
the in silico predictions, shed significant light on the nature
of these enzymes, their expression status and their ability to
associate with each other.
Materials and methods
Phages, bacterial strains and plasmids
Escherichia coli XL1Blue was used for routine purposes.
The expression vector pQE 30 (Qiagen) was used for over-
expression of the phage genes. For phage induction experi-
ments, the thermoinducible lysogenic strain Mycobacterium
smegmatis L1c1ts (obtained from Dr N.C. Mandal) was used
(Chaudhuri et al., 1993; Chatterjee et al., 2000). This strain
harbors a thermo-inducible lysogenic mutant of mycobac-
teriophage L1 as a prophage. Although the complete gen-
ome sequence of L1 phage is not available, restriction maps
(Lee et al., 1991) and DNA sequences of several L1 genes
(Datta & Mandal, 1998; Chattopadhyay et al., 2003; Sau
et al., 2004; Ganguly et al., 2007) indicate that L1 is only a
minor variant of L5. Hence, the use of an L1 lysogen to study
the L5 developmental pathways is justified. Cloning and
expression of mycobacteriophage genes 48 and 50 were
performed using genomic DNA derived from mycobac-
teiophage L5 (Hatfull & Sarkis, 1993). In case of Gp50, both
enzymatic and immunological experiments were conducted
using the L5 protein. In case of Gp48, immunological
experiments were conducted using antisera raised against
the L5 protein, but for enzymological studies Gp48 of D29
(Ford et al., 1998) was used, as the L5 protein could not be
recovered in the soluble fraction. D29 is a lytic phage
belonging to the L5 family. Many of the polypeptides
encoded by D29 share extensive homology with those
encoded by L5. In case of Gp48, the homology is 470%.
FEMS Microbiol Lett 280 (2008) 64–72 c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
65Mycobacteriophage ThyX and RNR

Cloning of mycobacteriophage genes inexpression vectors
Mycobacteriophage DNA were isolated from L5 or D29 as
per standard methods using CsCl purified phages (Chatter-
jee et al., 2000). The phage DNA was then used for PCR
amplification of desired genes. For L5 gene 50, the primers
used were P1, forward primer (50-GGGGTACCTTGACTG
ACGAAATCC-30) and P2, reverse primer (50-CCCAAGCTT
CACTTAATGGGGCATGC-30), (the KpnI and HindIII sites
are underlined). The PCR product was digested with KpnI
and HindIII for cloning in the expression vector pQE 30
(Qiagen). The L5 gene 48 was amplified using primers P3,
forward primer (50-GAAGATCTCATATGAAAGCCAAACT
GATC-30), and P4, reverse primer (50-CCCAAGCTTCA
TATGTCAGTAGCTGTAG-30) (the BglII and HindIII sites
are underlined). The PCR product was digested with BglII
and HindIII for cloning in the expression vector pQE 30. D29
gene 48 was amplified using the forward primer (50-CG
GGATCCATGAAAGTCCAACTGATC-30) and the reverse
primer (50-CCCAAGCTTTCAGCCTCCGTAGCTG-30) (the
BamHI and HindIII sites are underlined). The amplicon was
cloned at the BamHI–HindIII site of pQE-30.
Expression and purification of recombinantproteins
Hexa-histidine-tagged recombinant protein purification
was performed using Ni-nitrilotriacetic acid (NTA) agarose
chromatography either under denaturing conditions or
under native conditions as per standard protocols (Qiagen).
Cells were harvested by centrifugation and sonicated in
buffer A (50 mM sodium phosphate pH 8.0, 300 mM NaCl,
20 mM imidazole). After centrifugation at 14 000 g, the
clear supernatant was loaded onto a 2-mL Ni21-NTA
agarose column, pre-equilibrated with buffer A. The
column was washed with 10 column volumes of buffer B
(50 mM sodium phosphate pH 8.0, 300 mM NaCl, 50 mM
imidazole) and the bound protein was eluted with buffer C
(50 mM sodium phosphate pH 8.0, 300 mM NaCl, 250 mM
imidazole) and analyzed by 12% sodium dodecyl sulfate-
polyacrylamide gel electrophoresis (SDS-PAGE). Fractions
containing pure protein were pooled and dialyzed over-
night at 4 1C in the dark against 50 mM Tris-HCl, pH 7.4,
containing 10% glycerol. For isolating proteins under
denaturing conditions, pellet was dissolved in lysis buffer D
(10 mM Tris, 50 mM sodium phosphate, 300 mM NaCl,
8 M urea, pH 8.0), sonicated, centrifuged and loaded onto
a Ni-NTA column. After washing with 10 column volume of
buffer E (10 mM Tris, 50 mM sodium phosphate, 300 mM
NaCl, 8 M urea, pH 6.3), protein was eluted in buffer F
(10 mM Tris, 50 mM sodium phosphate, 300 mM NaCl,
8 M urea, pH 4.5).
Induction of lysogen
The thermoinducible lysogen, M. smegmatis L1c1ts, was
grown to an OD of 0.8 at 28 1C in Middlebrook 7H9
(MB7H9) broth as described earlier (Chaudhuri et al.,
1993) and induced at 42 1C for 30 min, followed by growth
at 37 1C for 3 h. Aliquots were removed at definite intervals
(15, 30 and 60 min) after the flasks were immersed in the
42 1C water bath and processed for immunological and
enzymatic assays.
Immunodetection
Recombinant mycobacteriophage proteins were purified to
near homogeneity by affinity chromatography on Ni-NTA
agarose under denaturing conditions. The purified protein
was further separated from any contaminant proteins by
SDS-PAGE. The bands were excised and crushed before
injecting into rabbits according to previously standardized
protocols (Basu et al., 2002). Both immune and preimmune
sera were collected. The specificities of the derived antisera
were confirmed by Western blotting (1 : 1000 dilution) using
purified proteins. For immunodetection of phage-derived
proteins, cell-free extracts of thermo-induced M. smegmatis
L1c1ts lysogens were subjected to Western blotting using
specific antibodies (1 : 1000 dilution) according to standard
protocols (Sambrook et al., 1989).
Immunoprecipitation
Mycobacterium smegmatis L1c1ts lysogenic cells were grown
at 28 1C and then the temperature was shifted to 42 1C. After
15 min of induction, the proteins expressed were pulse label-
ed with 35S methionine (specific activity 3000 Ci mmol�1),
obtained from BRIT, Mumbai, India), for 3 min. Extracts
were prepared by sonicating the labeled cells in immuno-
precipitation buffer (50 mM Tris pH 7.4, 500 mM NaCl,
1 mM EDTA, 0.1% Nonidet P-40) and then incubated with
immune or preimmune sera (1 : 1000 dilution), overnight at
4 1C in the same buffer. Antibody-target complexes were
then pulled down using 25 mL of Protein-A Sepharose beads
(Amersham GE) (pre-equilibrated with the immunopreci-
pitation buffer containing 1% bovine serum albumin to
block nonspecific interactions) by centrifugation at 1000 g.
After washing five times with the immunoprecipitation
buffer, the beads were suspended in 1� SDS sample buffer,
boiled for 5 min and resolved by 13.5% SDS-PAGE. Bands
were visualized by autoradiography.
Determination of ThyX activity
The NADPH oxidase activity of ThyX was measured in a
total volume of 1 mL. The basal reaction components were
50 mM Tris-HCl, pH 7.4, 1 mM MgCl2, 10% glycerol. In
addition, dUMP, NADPH and FAD were added either alone
FEMS Microbiol Lett 280 (2008) 64–72c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
66 B. Bhattacharya et al.

or in specific combinations at desired concentrations.
Activity was monitored by net decrease of A340 nm using a
CARY 50 spectrophotometer (Varian). An extinction coeffi-
cient of 6400 cm�1 at 340 nm (e340) was used to quantify
absorption changes.
Thymidylate-synthesizing activity of ThyX was deter-
mined by monitoring the amount of tritium transferred to
water using [5-3H] dUMP (Movarek Biochemicals) as a
substrate as described previously (Griffin et al., 2005). The
standard reaction mixture (100mL) contained 50 mM
Tris-HCl, pH 7.4, 200 mM 5, 10-CH2H4folate, 500 mM
NADPH, 60mM FAD, 1 mM MgCl2 and 10% glycerol. The
specific activity of the [5-3H]dUMP stock used in the
experiments was 14.3 Ci mmol�1. The reaction was started
with the addition of 9 mg of purified ThyX. After 3 min, the
reaction was terminated by the addition of 300 mL of a
100 mg mL�1 activated charcoal suspension containing 2%
tricarboxylic acid, to remove the unused radiolabeled sub-
strate. Samples were mixed at room temperature for 1 h and
centrifuged at 12 000 g for 10 min to pellet the charcoal.
Radioactivity in the supernatant was determined by liquid
scintillation counting. Enzymatic activity of the protein was
expressed as nmol of dTMP formed min�1 mg�1.
Thymidylate synthase activity in cell extracts of induced
M. smegmatis L1c1ts cells was performed by inducing
200 mL of M. smegmatis L1c1ts culture at 42 1C. Fifty
milliliters of aliquots were removed at specified time points
and the harvested cells were sonicated. After removal of
debris by centrifugation, the extracts (60mL) were used in
the assays. The protein concentrations in the extracts were
nearly the same. If necessary, the activity data were normal-
ized with respect to the protein concentration. An assay for
ThyX was performed as described above. For ThyA activity,
the same buffer was used, except that NADPH and FAD
were omitted. The results were expressed as relative activity
taking the activity of the uninduced extract as unity.
Spectrophotometric detection of protein-boundFAD
Detection of FAD bound to protein was performed as
described earlier (Myllykallio et al., 2002) by incubating the
protein at 95 1C for 10 min in the dark. Precipitated protein
were pelleted by centrifugation at 10 000 g for 10 min. The
absorption spectra of the released flavin present in the
supernatant was determined spectrophotometrically from
250 to 750 nm in a 1-cm quartz cuvette.
RNR assay
RNR activity was determined by the diphenylamine proce-
dure of Blakley (1966) using ATP as the substrate. A
complete reaction mixture (0.5 mL) contained 0.2 mM ATP,
20 mM dithiothreitol, 10 mM 50-deoxyadenosylcobalamine
(coenzyme B12), 1 mM EDTA, 50 mM potassium phosphate
buffer, pH 7.3, and an appropriate amount of enzyme. The
reaction mixture was incubated for 1 h at 37 1C and the
reaction was terminated by placing the tubes in ice. The
reaction mixtures were treated with chloroacetamide and
subsequently with diphenylamine. Conversion of ATP to
dATP was monitored by recording the A595 nm. In some
cases, deoxyribonucleotides were incorporated in the assay
at desired concentrations to study stimulatory/inhibitory
effects.
Results
Immunodetection experiments
To test whether Gp48 and 50 were at all produced during
phage growth, the total protein extracted from M. smegmatis
L1c1ts cells induced for varying lengths of time was sub-
jected to Western blotting experiments using respective
antisera. The results (Fig. 1a and b) showed that Gp48 or
50 are present at detectable levels in the induced, but not
uninduced, extracts.
Following the demonstration of the presence of these
proteins in extracts of induced lysogenic cells by Western
blotting, immunoprecipitation of 35S-methionine pulse-
labeled proteins was attempted using the respective anti-
bodies. The rationale for this was twofold – first, a positive
CtrlkDa97664329
21
14
Antisera
kDa
kDa
976643292114
9766
43
29
21
0′ 15′ 30′ 60′ Ctrl 0′ 15′ 30′ 60′
Gp48
Gp48
Gp48
Gp49
Gp49
Gp50
Gp50
Gp50
I P I P I P
(a) (b)
(c)
Fig. 1. (a and b) Immunodetection of polypeptides synthesized from
genes 48 and 50 (a and b, respectively) using the corresponding antisera.
(c) Immunoprecipitation using antisera against Gp48, 50 and 49 as
indicated. P and I are abbreviated forms of preimmune and immune,
respectively. The identities of the immuno-precipitated proteins are
indicated by solid arrows. Bands representing unidentified proteins are
indicated by broken arrows. L5 Gp49 was expressed and processed for
the antibody essentially as described in the case of Gp48 and Gp50. In
each blot, the purified protein was incorporated as a control (Ctrl). The
time periods (mins) for which the inductions were performed are shown
on the top. The positions of molecular weight standards are shown.
FEMS Microbiol Lett 280 (2008) 64–72 c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
67Mycobacteriophage ThyX and RNR
immunoprecipitation would indicate that these proteins are
synthesized actively during the induced phase of phage
growth, and second, such an experiment might lead to
important associated proteins. The pulse-labeled proteins
obtained from cells induced for 15 min were thus subjected
to immunoprecipitation using either immune or preim-
mune sera against Gp48 and 50, respectively (Fig. 1c). As a
control experiment, immunoprecipitation of a third protein
Gp49 expressed from gene 49 was performed using the
corresponding antisera. The results showed that antisera
against Gp48 immunoprecipitated the corresponding
28 kDa polypeptide (Fig. 1c marked by an arrow). No
immunoprecipitation of this polypeptide, or for that matter
any other polypeptide, was observed using the preimmune
sera. In case of anti-Gp50, similar immunoprecipitation of
the specific target was observed. Interestingly, anti-Gp48
and 50 cross-immunoprecipitated each other’s targets and
also a couple of other proteins (Fig. 1c, marked by broken
arrows). This result indicates that in vivo, Gp48, 50 and
several other unidentified proteins associate to form a
complex. The fact that none of these bands were precipitated
in case of Gp49 gives additional support to the claim that the
precipitation is not artifactual. Moreover, it is unlikely that
coimmunoprecipitation is due to epitope sharing because in
the Western blots, performed at the same dilution of antisera
(Fig. 1a and b) no evidence of aberrant reactivity was found.
Gp48-a mycobacteriophage flavin-dependentthymidylate synthase (FDTS)
In order to characterize Gp48 biochemically, attempts were
first made to express the L5 gene in E. coli. Although a high
expression level was obtained, the protein could not be
recovered in the soluble state. In order to overcome this
difficulty, the D29 gene 48 that shares extensive homology
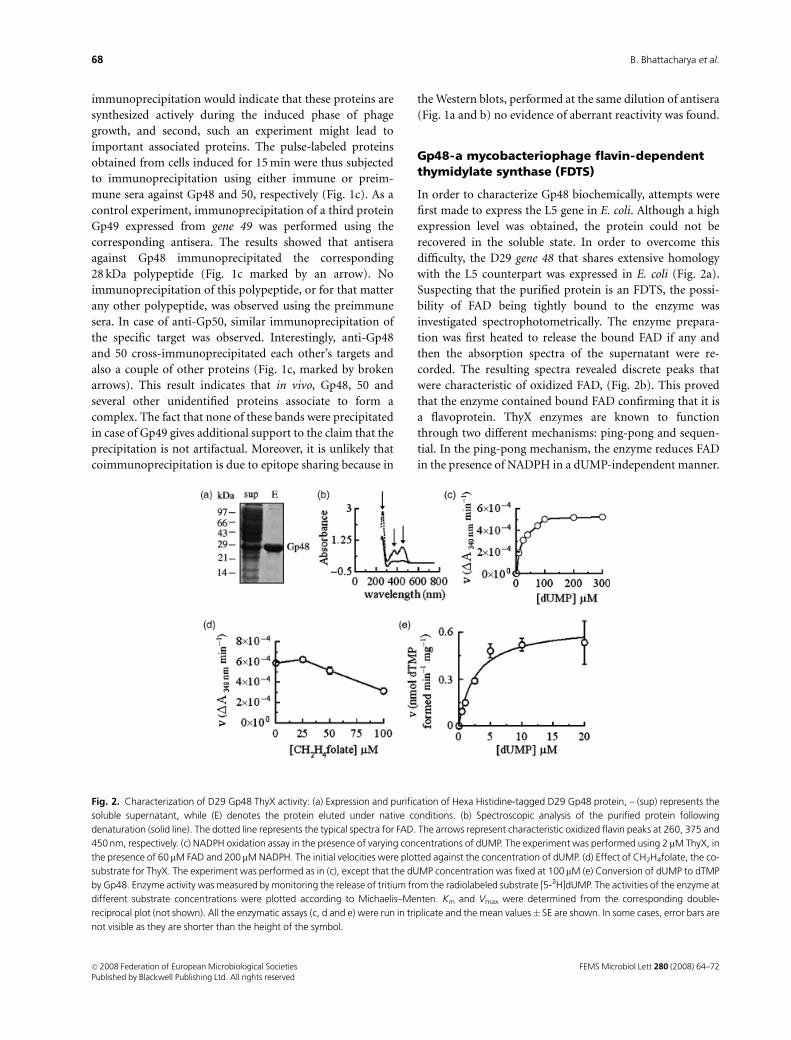
with the L5 counterpart was expressed in E. coli (Fig. 2a).
Suspecting that the purified protein is an FDTS, the possi-
bility of FAD being tightly bound to the enzyme was
investigated spectrophotometrically. The enzyme prepara-
tion was first heated to release the bound FAD if any and
then the absorption spectra of the supernatant were re-
corded. The resulting spectra revealed discrete peaks that
were characteristic of oxidized FAD, (Fig. 2b). This proved
that the enzyme contained bound FAD confirming that it is
a flavoprotein. ThyX enzymes are known to function
through two different mechanisms: ping-pong and sequen-
tial. In the ping-pong mechanism, the enzyme reduces FAD
in the presence of NADPH in a dUMP-independent manner.
Fig. 2. Characterization of D29 Gp48 ThyX activity: (a) Expression and purification of Hexa Histidine-tagged D29 Gp48 protein, – (sup) represents the
soluble supernatant, while (E) denotes the protein eluted under native conditions. (b) Spectroscopic analysis of the purified protein following
denaturation (solid line). The dotted line represents the typical spectra for FAD. The arrows represent characteristic oxidized flavin peaks at 260, 375 and
450 nm, respectively. (c) NADPH oxidation assay in the presence of varying concentrations of dUMP. The experiment was performed using 2 mM ThyX, in
the presence of 60 mM FAD and 200mM NADPH. The initial velocities were plotted against the concentration of dUMP. (d) Effect of CH2H4folate, the co-
substrate for ThyX. The experiment was performed as in (c), except that the dUMP concentration was fixed at 100 mM (e) Conversion of dUMP to dTMP
by Gp48. Enzyme activity was measured by monitoring the release of tritium from the radiolabeled substrate [5-3H]dUMP. The activities of the enzyme at
different substrate concentrations were plotted according to Michaelis–Menten. Km and Vmax were determined from the corresponding double-
reciprocal plot (not shown). All the enzymatic assays (c, d and e) were run in triplicate and the mean values� SE are shown. In some cases, error bars are
not visible as they are shorter than the height of the symbol.
FEMS Microbiol Lett 280 (2008) 64–72c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
68 B. Bhattacharya et al.
In the sequential mechanism, however, both dUMP and
NADPH must bind to the enzyme for the reduction of FAD.
In other words, the enzyme is expected to have dUMP-
dependent NADPH oxidase activity. In order to obtain an
insight into which of these mechanisms operates in case of
D29 ThyX-catalyzed reactions, the NADPH oxidase activity
of the protein was assessed in the absence or in the presence
of increasing concentrations of dUMP, maintaining NADPH
at a saturating concentration of 200mM. The results show
that the enzyme is a dUMP-dependent NADPH oxidase
(Fig. 2c). From the corresponding double-reciprocal plot,
the Km could be determined to be about 1mM for dUMP.
The ability of a ThyX to act as a dUMP-dependent NADPH
oxidase has been reported in case of the Paramecium
bursaria chlorella virus (PBCV)-I enzyme. In the above,
assay CH2H4folate was absent. If the same NADPH oxida-
tion assay was performed in the presence of CH2H4folate, a
significant dose-dependent inhibition of the NADPH oxi-
dase activity was observed (Fig. 2d). This indicates that as in
the case of the PBCV-1 enzyme, the CH2H4folate-binding
site of D29 ThyX overlaps with that of the NADPH-binding
site. The ability of the enzyme to convert dUMP to dTMP in
the presence of NADPH and CH2H4folate (200mM each)
was then assayed by the tritium release assay. From the
resulting curve (as shown in Fig. 2e), the Km for dUMP was
estimated to be about 3 mM, which is comparable to the
1 mM value obtained using the NADPH assay. The results,
taken together, indicate that Gp48 acts as an FDTS by the
sequential mechanism as proposed in case of the PBCV-1
enzyme (Graziani et al., 2004).
These investigations were performed with the D29 en-
zyme. However, a soluble glutatione-S-transferase-tagged
version of the L5 protein was studied. The properties of this
fusion protein were similar to that of the D29 protein.
Gp50- a Class II ribonucleotide reductase
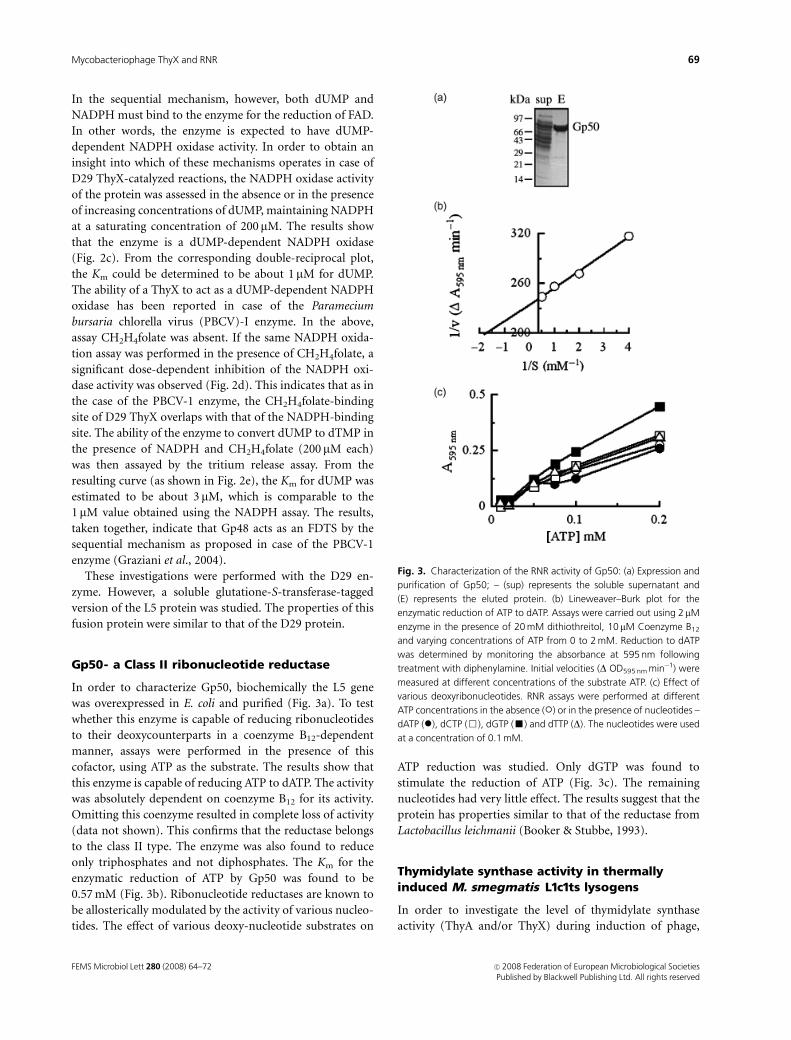
In order to characterize Gp50, biochemically the L5 gene
was overexpressed in E. coli and purified (Fig. 3a). To test
whether this enzyme is capable of reducing ribonucleotides
to their deoxycounterparts in a coenzyme B12-dependent
manner, assays were performed in the presence of this
cofactor, using ATP as the substrate. The results show that
this enzyme is capable of reducing ATP to dATP. The activity
was absolutely dependent on coenzyme B12 for its activity.
Omitting this coenzyme resulted in complete loss of activity
(data not shown). This confirms that the reductase belongs
to the class II type. The enzyme was also found to reduce
only triphosphates and not diphosphates. The Km for the
enzymatic reduction of ATP by Gp50 was found to be
0.57 mM (Fig. 3b). Ribonucleotide reductases are known to
be allosterically modulated by the activity of various nucleo-
tides. The effect of various deoxy-nucleotide substrates on
ATP reduction was studied. Only dGTP was found to
stimulate the reduction of ATP (Fig. 3c). The remaining
nucleotides had very little effect. The results suggest that the
protein has properties similar to that of the reductase from
Lactobacillus leichmanii (Booker & Stubbe, 1993).
Thymidylate synthase activity in thermallyinduced M. smegmatis L1c1ts lysogens
In order to investigate the level of thymidylate synthase
activity (ThyA and/or ThyX) during induction of phage,
Fig. 3. Characterization of the RNR activity of Gp50: (a) Expression and
purification of Gp50; – (sup) represents the soluble supernatant and
(E) represents the eluted protein. (b) Lineweaver–Burk plot for the
enzymatic reduction of ATP to dATP. Assays were carried out using 2 mM
enzyme in the presence of 20 mM dithiothreitol, 10mM Coenzyme B12
and varying concentrations of ATP from 0 to 2 mM. Reduction to dATP
was determined by monitoring the absorbance at 595 nm following
treatment with diphenylamine. Initial velocities (D OD595 nm min�1) were
measured at different concentrations of the substrate ATP. (c) Effect of
various deoxyribonucleotides. RNR assays were performed at different
ATP concentrations in the absence (�) or in the presence of nucleotides –
dATP (�), dCTP (&), dGTP (’) and dTTP (D). The nucleotides were used
at a concentration of 0.1 mM.
FEMS Microbiol Lett 280 (2008) 64–72 c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
69Mycobacteriophage ThyX and RNR
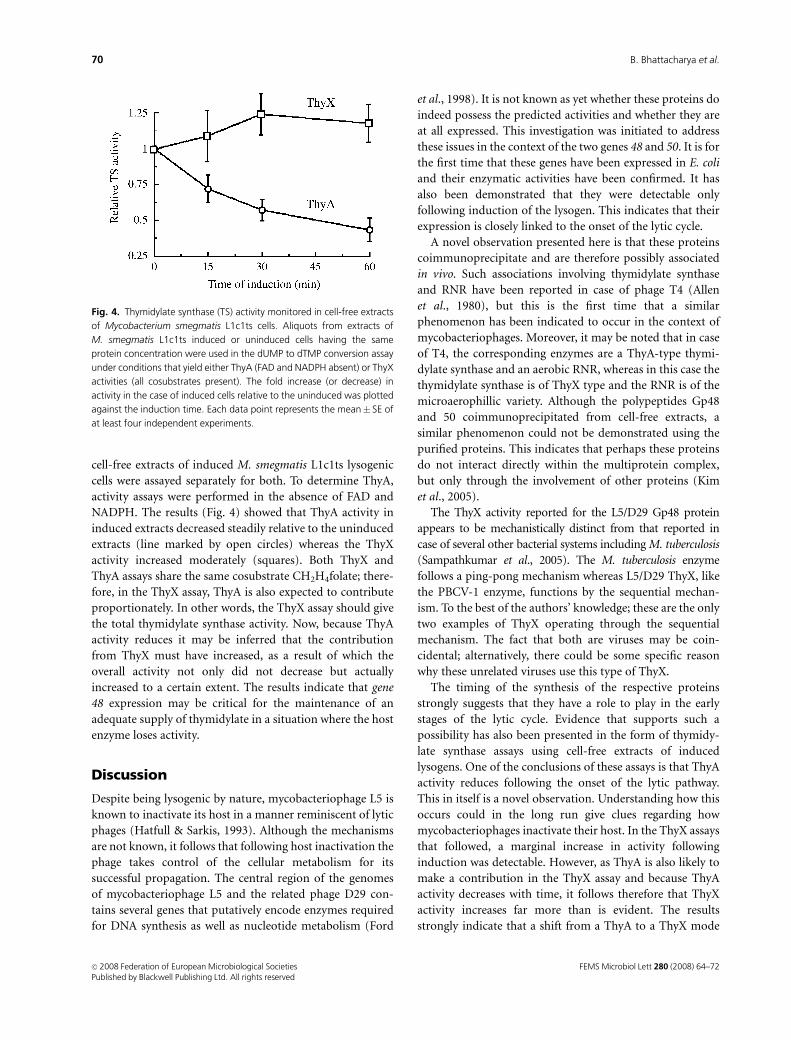
cell-free extracts of induced M. smegmatis L1c1ts lysogenic
cells were assayed separately for both. To determine ThyA,
activity assays were performed in the absence of FAD and
NADPH. The results (Fig. 4) showed that ThyA activity in
induced extracts decreased steadily relative to the uninduced
extracts (line marked by open circles) whereas the ThyX
activity increased moderately (squares). Both ThyX and
ThyA assays share the same cosubstrate CH2H4folate; there-
fore, in the ThyX assay, ThyA is also expected to contribute
proportionately. In other words, the ThyX assay should give
the total thymidylate synthase activity. Now, because ThyA
activity reduces it may be inferred that the contribution
from ThyX must have increased, as a result of which the
overall activity not only did not decrease but actually
increased to a certain extent. The results indicate that gene
48 expression may be critical for the maintenance of an
adequate supply of thymidylate in a situation where the host
enzyme loses activity.
Discussion
Despite being lysogenic by nature, mycobacteriophage L5 is
known to inactivate its host in a manner reminiscent of lytic
phages (Hatfull & Sarkis, 1993). Although the mechanisms
are not known, it follows that following host inactivation the
phage takes control of the cellular metabolism for its
successful propagation. The central region of the genomes
of mycobacteriophage L5 and the related phage D29 con-
tains several genes that putatively encode enzymes required
for DNA synthesis as well as nucleotide metabolism (Ford
et al., 1998). It is not known as yet whether these proteins do
indeed possess the predicted activities and whether they are
at all expressed. This investigation was initiated to address
these issues in the context of the two genes 48 and 50. It is for
the first time that these genes have been expressed in E. coli
and their enzymatic activities have been confirmed. It has
also been demonstrated that they were detectable only
following induction of the lysogen. This indicates that their
expression is closely linked to the onset of the lytic cycle.
A novel observation presented here is that these proteins
coimmunoprecipitate and are therefore possibly associated
in vivo. Such associations involving thymidylate synthase
and RNR have been reported in case of phage T4 (Allen
et al., 1980), but this is the first time that a similar
phenomenon has been indicated to occur in the context of
mycobacteriophages. Moreover, it may be noted that in case
of T4, the corresponding enzymes are a ThyA-type thymi-
dylate synthase and an aerobic RNR, whereas in this case the
thymidylate synthase is of ThyX type and the RNR is of the
microaerophillic variety. Although the polypeptides Gp48
and 50 coimmunoprecipitated from cell-free extracts, a
similar phenomenon could not be demonstrated using the
purified proteins. This indicates that perhaps these proteins
do not interact directly within the multiprotein complex,
but only through the involvement of other proteins (Kim
et al., 2005).
The ThyX activity reported for the L5/D29 Gp48 protein
appears to be mechanistically distinct from that reported in
case of several other bacterial systems including M. tuberculosis
(Sampathkumar et al., 2005). The M. tuberculosis enzyme
follows a ping-pong mechanism whereas L5/D29 ThyX, like
the PBCV-1 enzyme, functions by the sequential mechan-
ism. To the best of the authors’ knowledge; these are the only
two examples of ThyX operating through the sequential
mechanism. The fact that both are viruses may be coin-
cidental; alternatively, there could be some specific reason
why these unrelated viruses use this type of ThyX.
The timing of the synthesis of the respective proteins
strongly suggests that they have a role to play in the early
stages of the lytic cycle. Evidence that supports such a
possibility has also been presented in the form of thymidy-
late synthase assays using cell-free extracts of induced
lysogens. One of the conclusions of these assays is that ThyA
activity reduces following the onset of the lytic pathway.
This in itself is a novel observation. Understanding how this
occurs could in the long run give clues regarding how
mycobacteriophages inactivate their host. In the ThyX assays
that followed, a marginal increase in activity following
induction was detectable. However, as ThyA is also likely to
make a contribution in the ThyX assay and because ThyA
activity decreases with time, it follows therefore that ThyX
activity increases far more than is evident. The results
strongly indicate that a shift from a ThyA to a ThyX mode
Fig. 4. Thymidylate synthase (TS) activity monitored in cell-free extracts
of Mycobacterium smegmatis L1c1ts cells. Aliquots from extracts of
M. smegmatis L1c1ts induced or uninduced cells having the same
protein concentration were used in the dUMP to dTMP conversion assay
under conditions that yield either ThyA (FAD and NADPH absent) or ThyX
activities (all cosubstrates present). The fold increase (or decrease) in
activity in the case of induced cells relative to the uninduced was plotted
against the induction time. Each data point represents the mean� SE of
at least four independent experiments.
FEMS Microbiol Lett 280 (2008) 64–72c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
70 B. Bhattacharya et al.

of synthesis occurs as phage growth commences. Whatever
is true for ThyX is also probably true for the phage-encoded
RNR II, although this has not been verified. The results
of this investigation indicate that like other well-known
lytic phages, L5 (and related phages) manipulate host
nucleotide metabolism in such a way as to benefit itself at
the cost of the host.
Acknowledgements
The authors thank N.C. Mandal, A.K. Tyagi and R. McNerney
for the phages L1, L5 and D29, respectively. The project was
funded by a grant from DBT, Government of India. B.B.
and N.G. are grateful to CSIR and UGC, Government of
India for their fellowships. The authors thank P. Halder for
technical assistance.
References
Allen JR, Reddy GP, Lasser GW & Mathews CK (1980) T4
ribonucleotide reductase. Physical and kinetic linkage to other
enzymes of deoxyribonucleotide biosynthesis. J Biol Chem 255:
7583–7588.
Allen JR, Lasser GW, Goldman DA, Booth JW & Mathews CK
(1983) T4 phage deoxyribonucleotide-synthesizing enzyme
complex. Further studies on enzyme composition and
regulation. J Biol Chem 258: 5746–5753.
Basu A, Chawla-Sarkar M, Chakrabarti S & Das Gupta SK (2002)
Origin binding activity of the Mycobacterial plasmid pAL5000
replication protein RepB is stimulated through interactions
with host factors and coupled expression of repA. J Bacteriol
184: 2204–2214.
Blakley LR (1966) Cobamides and ribonucleotide reduction.
J Biol Chem 241: 176–179.
Booker S & Stubbe J (1993) Cloning, sequencing, and expression
of the adenosylcobalamin-dependent ribonucleotide reductase
from Lactobacillus leichmannii. Proc Natl Acad Sci USA 90:
8352–8356.
Chatterjee S, Mitra M & Das Gupta SK (2000) A high yielding
mutant of mycobacteriophage L1 and its application as a
diagnostic tool. FEMS Microbiol Lett 188: 47–53.
Chattopadhyay C, Sau S & Mandal NC (2003) Cloning and
characterization of the promoters of temperate
mycobacteriophage L1. J Biochem Mol Biol 36: 586–592.
Chaudhuri B, Sau S, Datta HJ & Mandal NC (1993) Isolation,
characterization, and mapping of temperature-sensitive
mutations in the genes essential for lysogenic and lytic growth
of the mycobacteriophage L1. Virology 194: 166–172.
Datta HJ & Mandal NC (1998) Identification of an early positive
regulatory gene of mycobacteriophage L1. J Gen Virol 79:
205–210.
Doke S (1960) Studies on mycobacteriophages and mycobacteria.
J Kumamoto Med Soc 34: 1360–1373.
Donnelly-Wu MK, Jacobs WR Jr & Hatfull GF (1993)
Superinfection immunity of mycobacteriophage L5:
applications for genetic transformation of mycobacteria. Mol
Microbiol 7: 407–417.
Ford ME, Sarkis GJ, Belanger AE, Hendrix RW & Hatfull GF
(1998) Genome structure of mycobacteriophage D29:
implications for phage evolution. J Mol Biol 279: 143–164.
Ganguly T, Bandhu A, Chattoraj P, Chanda PK, Das M, Mandal
NC & Sau S (2007) Repressor of temperate mycobacteriophage
L1 harbors a stable C-terminal domain and binds to different
asymmetric operator DNAs with variable affinity. Virol J 4: 64.
Garcia M, Pimentel M & Moniz-Pereira J (2002) Expression of
Mycobacteriophage Ms6 lysis genes is driven by two
sigma(70)-like promoters and is dependent on a transcription
termination signal present in the leader RNA. J Bacteriol 184:
3034–3043.
Graziani S, Xia Y, Gurnon JR et al. (2004) Functional analysis of
FAD-dependent thymidylate synthase ThyX from Paramecium
bursaria Chlorella virus-1. J Biol Chem 279: 54340–54347.
Griffin J, Roshick C, Iliffe-Lee E & McClarty G (2005) Catalytic
mechanism of Chlamydia trachomatis flavin-dependent
thymidylate synthase. J Biol Chem 280: 5456–5467.
Hatfull GF & Jacobs WR Jr (1994) Mycobacteriophages: corner-
stones of mycobacterial research. Tuberculosis: Pathogenesis,
Protection, and Control (Bloom BR, ed), pp. 165–183.
American Society for Microbiology, Washington, DC.
Hatfull GF & Sarkis GJ (1993) DNA sequence, structure and gene
expression of mycobacteriophage L5: a phage system for
mycobacterial genetics. Mol Microbiol 7: 395–405.
Jain S & Hatfull GF (2000) Transcriptional regulation and
immunity in mycobacteriophage Bxb1. Mol Microbiol 38:
971–985.
Kim J, Wheeler LJ, Shen R & Mathews CK (2005) Protein-DNA
interactions in the T4 dNTP synthetase complex dependent on
gene 32 single-stranded DNA-binding protein. Mol Microbiol
55: 1502–1514.
Lee MH & Hatfull GF (1993) Mycobacteriophage L5 integrase-
mediated site-specific integration in vitro. J Bacteriol 175:
6836–6841.
Lee MH, Pascopella L, Jacobs WR Jr & Hatfull GF (1991) Site-
specific integration of mycobacteriophage L5: integration-
proficient vectors for Mycobacterium smegmatis,
Mycobacterium tuberculosis, and bacille Calmette-Guerin. Proc
Natl Acad Sci USA 88: 3111–3115.
Lewis JA & Hatfull GF (2000) Identification and characterization
of mycobacteriophage L5 excisionase. Mol Microbiol 35:
350–360.
Mathews CK (1993) The cell-bag of enzymes or network of
channels? J Bacteriol 175: 6377–6381.
Myllykallio H, Lipowski G, Leduc D, Filee J, Forterre P & Liebl U
(2002) An alternative flavin-dependent mechanism for
thymidylate synthesis. Science 297: 105–107.
Pedulla ML, Ford ME, Houtz JM et al. (2003) Origins of highly
mosaic mycobacteriophage genomes. Cell 113: 171–182.
FEMS Microbiol Lett 280 (2008) 64–72 c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
71Mycobacteriophage ThyX and RNR

Piuri M & Hatfull GF (2006) A peptidoglycan hydrolase motif
within the mycobacteriophage TM4 tape measure protein
promotes efficient infection of stationary phase cells. Mol
Microbiol 62: 1569–1585.
Poole AM, Logan DT & Sjoberg BM (2002) The evolution of the
ribonucleotide reductases: much ado about oxygen. J Mol Evol
55: 180–196.
Prem veer Reddy G & Pardee AB (1980) Multienzyme complex
for metabolic channeling in mammalian DNA replication.
Proc Natl Acad Sci USA 77: 3312–3316.
Reddy GP, Singh A, Stafford ME & Mathews CK (1977) Enzyme
associations in T4 phage DNA precursor synthesis. Proc Natl
Acad Sci USA 74: 3152–3156.
Ribeiro G, Viveiros M, David HL & Costa JV (1997)
Mycobacteriophage D29 contains an integration system
similar to that of the temperate mycobacteriophage L5.
Microbiology 143: 2701–2708.
Sambrook J, Fritsch EF & Maniatis T (1989) Molecular Cloning: A
Laboratory Manual, 2nd edn. Cold Spring Harbor Laboratory,
Cold Spring Harbor, NY.
Sampathkumar P, Turley S, Ulmer JE, Rhie HG, Sibley CH & Hol
WG (2005) Structure of the Mycobacterium tuberculosis flavin
dependent thymidylate synthase (MtbThyX) at 2.0A
resolution. J Mol Biol 352: 1091–1104.
Sau S, Chattoraj P, Ganguly T, Lee CY & Mandal NC (2004)
Cloning and sequencing analysis of the repressor gene of
temperate mycobacteriophage L1. J Biochem Mol Biol 37:
254–259.
Snapper SB, Lugosi L, Jekkel A, Melton RE, Kieser T, Bloom BR &
Jacobs WR Jr (1988) Lysogeny and transformation in
mycobacteria: stable expression of foreign genes. Proc Natl
Acad Sci USA 85: 6987–6991.
Tomich PK, Chiu CS, Wovcha MG & Greenberg GR (1974)
Evidence for a complex regulating the in vivo activities of early
enzymes induced by bacteriophage T4. J Biol Chem 249:
7613–7622.
Zhu H, Yin S & Shuman S (2004) Characterization of
polynucleotide kinase/phosphatase enzymes from
Mycobacteriophages omega and Cjw1 and vibriophage
KVP40. J Biol Chem 279: 26358–26369.
FEMS Microbiol Lett 280 (2008) 64–72c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
72 B. Bhattacharya et al.





























