Embed Size (px)
Citation preview

YEAST VOL. 11: 425433 (1995)
Cloning and Sequencing of the URA5 Gene from the Yeast Yarrowia lipolytica MANUEL SANCHEZ, MARCIANO PRADO, FRANCISCO J. IGLESIASt AND ANGEL DOMINGUEZ*
Departamento de Microbiologia y Genhtica, tDepartamento de Fisica Aplicada, Universidad de Salamanca, 37071 Salamanca, Spain
Received 28 October 1994; accepted 18 December 1994
The lJRA.5 gene of Yarrowia lipolytica encoding the orotate phosphoribosyl transferase (OPRTase, EC2.4.2.10) was isolated by target integration in a mutant strain originally named ura2.21. The nucleotide sequence of the gene predicts a protein with high similarities with the OPRTases from Saccharomyces cerevisiae, Podospora anserina and Escherichia coli and to a lesser extent with that of Dictyostelium discoideum. The transcription start point has been mapped by primer extension analysis and indicates the existence of a long leader sequence in the corresponding mRNA. Northern-blot hybridization revealed the URA.5 transcript to be approximately 0.94 kb. Deletion of the URA.5 gene in Y. lipolytica produced a leaky phenotype similar to the one described for the uraS mutation in S. cerevisiae. The URAS gene of Y lipolytica was able to complement functionally the ura5 mutation of S. cerevisiae. The sequence presented here has been submitted to the EMBL data library under Accession Number 222571.
KEY WORDS - Yarrowia lipolytica; orotate phosphoribosyl transferase; nucleotide sequence; transcription
INTRODUCTION The organization of the pyrimidine pathway differs from prokaryotes to eukaryotes. Most prokaryotes have six structural genes which code for the six enzymes required for biosynthesis of UMP (Poulsen et al., 1983). In Saccharomyces cerevisiae and Neurospora crassa, there are only five struc- tural genes and one of them, URA2 in S. cerevisiae and PYR3 in N. crassa, codes for a bi-functional polypeptide which has carbamyl phosphate synthetase (CPSase, EC 2.7.29) and aspartate transcarbamylase (ATCase, EC 2.1.3.2) activities (Makoff and Radford, 1978; Souciet et al., 1989). In higher eukaryotes, the six enzymes appear to be encoded by only three structural genes. One gene encodes the first three enzymatic activities that are located in a multifunctional protein (CSPase, ATCase) and dihydroorotase (EC 3.5.2.3); the second gene codes for the dihydroorotate dehydro- genase (EC 3.5.2.3) and the third gene codes for a polypeptide with the other two enzyme activities, orotate phosphoribosyl transferase OPRTase, EC 2.4.2.10) and orotidine-5'-phosphate decarboxy- lase (OMPdecase, EC 4.1.1.23) (for a review, see
*Corresponding author
CCC 0749- 503W95/050425-09 J; 1995 by John Wiley & Sons Ltd
Jones, 1980). Also in Dictyostelium discoideum, another primitive eukaryotic organism (McCarroll et al., 1983), at least two enzymatic activities of the pathway, OPRTase and OMPdecase, are located in a bifunctional protein coded by the DdPYR5-6 gene, which contains an open reading frame (ORF) of 478 codons, as occurs similarly in mam- mals (Boy-Marcotte et al., 1984; Jacquet et al., 1988).
One mutation, ura2.21, in the pyrimidine pathway has been isolated in Yarrowia lipolytica that is not complemented functionally by the gene products of the URA3 or URA2 (either from Y. lipolytica or S. cerevisiae; A. Dominguez, un- published work). Since Y. lipolytica is unusual in both its rRNA sequence evolution and in its relationship to other yeasts (Barns et al., 1991), we decided to clone and characterize the gene that complements the ura2.21 mutation. After its isolation and sequencing, we demonstrated that the gene is homologous to the URAS gene of S. cerevisiae. Moreover, the Y. lipolytica gene isolated functionally complements the ura.5 mutation in S. cerevisiae. In the present work, we show that the OPRTase in Y. lipolytica is encoded by a single locus and shows strong similarities with the

426 M. SANCHEZ ET AL.
EmRl
Figure 1. Restriction maps of the plasmid used and constructed in this work pINA62 (Gaillardin and Ribet, 1987), pMP47, pMS16, pMS18 (our work, see text)
OPRTases from other microorganisms, and we propose renaming the mutation as ura5-21 and the gene as YIURA.5.
MATERIALS AND METHODS Strains and media
The Y. lipolytica strains W29 MAT A (wild- type); 21601-2 MAT A leu2 lys1.13; 11606.1B MAT B lys 3.2 and 21501-13 MAT A leu2 lys.5-12 ura 2.21 were kindly supplied by C. Gaillardin, Laboratoire de Genetique-INRA, Thiverval- Grignon, France. Strain MPY73, MAT A leu2 ura 2.21 was selected after sporulation of a diploid strain constructed by a cross between 11606.1B x 21501-13. They were grown in YED (1% yeast extract, 1% glucose) or in appropriately supplemented minimal media (0.67% yeast nitro- gen base, 1% glucose). The media and procedures used for mating, sporuiation and transformation of Y. lipolytica have already been described (Barth and Weber, 1985; Xuan el al., 1988).
The S. cerevisiae strain 5573-7c, MATa leu2-3 his4-712 trpl was kindly supplied by Dr F. del Rey (from our department). Strain FL476-1c, MATa um.5 was obtained from the Yeast Genetic Stock Center. S. cerevisiae strain MS-1 l e d - 3 his4-712
trpl ura.5 was selected by us after asci dissection of a diploid strain constructed by crossing between 5573-7c and FL476-lc. Media, procedures used for mating, sporulation and asci dissection for S. cerevisiae were as described by Sherman et al. (1977). Transformation of S. cerevisiae was carried out by electroporation (Becker and Guarente, 1991).
The Escherichia coli strains used for transfonna- tion and amplification of recombinant DNA were: DH5a supE44 AlacUI69((~80 IacZ AM15)hsdRI 7 recAl endAlgyrA96 thi-1 relAl (Hanahan, 1983); MC1061 hsdR mcrB araD139 A(araABC- leu)7679AlacX74 gulU galK rpsL thi (Meissner et al., 1987) and MV1190 A(1ac-pro AB) thi supE A(srl-recA)306:: TnlO(tet7 [F':tra 0 3 6 pro AB IacFZ AM151 (Bio-Rad). E. coli strains were grown on LB broth.
Screening by complementation of Ura' transformunts
The Y. lipolytica gene bank constructed in the plasmid pINA62 contains, as selectable marker, the LEU2 gene of this yeast. Unique sites, ApaI and Not1 have been localized outside the LEU2 gene (see Figure 1). As Not1 places are scattered at low frequencies in the yeast genome, we decided to

URAS GENE OF Y. LIPOLYTICA 427
linearize the gene bank by Not1 digestion prior to the transformation experiments for increased efficiency (Davidow et al., 1985).
After transformation of Y. lipolytica, the cells were plated onto the selective MM containing no leucine. The Leu+ transformants obtained were replica-plated on to MM to screen for Ura+ transformants.
After transformation of S. cerevisiae, the cells were plated alternatively onto the selective MM containing no tryptophan. The Trp+ transfor- mants obtained were replica-plated onto MM to screen for Ura+ transformants.
Plasmid The URAS gene was isolated as a 9.9 kb insert
from a DNA library constructed from Y. lipolytica W29 partially digested with Sau3A and cloned in the BamHI site of the pINA62 vector (Figure 1; Xuan et al., 1988). Sub-clones were constructed by internal deletions within the 9-9 kb insert present in the pMP47 (Figure 1). pMS-16 (Figure 1) contains a 1.3 kb, BamHI-NcoI fragment inserted in the BamHI site of plasmid pINA62. pMS-18 (Figure 1) contains a 2.7 kb fragment BamHI- HindIII (Figure 3) inserted in the BamHI- HindIII sites of the Tet' gene of YRp7 (Struhl et al., 1979).
DNA manipulations Restriction enzyme digestions and DNA liga-
tions were performed according to the recommen- dations of the manufacturer. Isolation of plasmid DNA from E. coli was performed using standard procedures (Sambrook et al., 1989). DNA frag- ments to be used as probes were labelled by random priming with digoxygenin-dUTP. Southern blotting, hybridization and immuno- logical detection were carried out according to the instructions of the manufacturer (Boehringer).
Sequence analysis of the URAS locus A cloned DNA restriction fragment harboring
the URAS gene was isolated from plasmid pMP47 as a 1.3 kb BarnHI-NcoI fragment. This fragment was sub-cloned into the BamHI-EcoRV sites of pBluescript plasmids (SK' and KS' , Stratagene), and a nested set of closely spaced delections was created by using exonuclease 111. Templates were sequenced on both strands with Sequenase enzyme (Boehringer) using the dideoxynucleotide chain- termination sequencing reactions (Sanger et al.,
1977). The products of sequencing reactions were resolved on buffer gradient polyacrylamide-urea sequencing gels (Biggin et al., 1983) and exposed to Kodak XAR-5 X-ray film.
RNA preparations, Northern analysis and transcript mapping
RNA was prepared from exponentially growing cultures on YED by the method of Percival-Smith and Segall(l984). Poly (A)+ RNA was isolated by affinity chromatography on oligo(dT)-cellulose. Prehybridization and hybridization were per- formed according to Sambrook et al. (1989). In all Northern analysis experiments, the RNA concen- tration was normalized by hybridization with the LEU2 gene from Y. lipolytica. Primer extension analysis was carried out essentially as described by Sambrook et al. (1989), by using a synthetic oligo- nucleotide (5'-3') complementary to nucleotide positions +15 to +44 of the URAS coding sequence.
DNA and protein sequences were analysed by using the DNASIS and PROSIS programs (Pharmacia-LKB, Hitachi). The amino-acid se- quence of URAS was compared with the SWISS- PROT data bank by using the FASTA program (Pearson and Lipman, 1988). Alignments of protein sequences were done with CLUSTAL programs (Higgins and Sharp, 1988).
RESULTS
Isolation and sequencing of the URAS gene From 104 000 colonies isolated as described in
Materials and Methods (transformation fre- quency; 6000 transformants per pg of DNA), we obtained four transformants capable of growth in the absence of uracil.
In order to obtain the physical map of the inserted DNA, we performed digestion of the DNAs of MP73, MP73 transformed with pINA62 and of the four Ura+ transformants. After electro- phoresis and Southern transfer, these DNAs were hybridized with the LEU2 probe. This probe hy- bridizes in the wild type with a BamHI-BamHI fragment of 11 kb or with a SphI-SphI fragment of 8 kb which contains the LEU2 gene (see also Xuan et al., 1988). The same probe hybridized in the case of one transformant with two BamHI-BamHI fragments of 8 and 13.7 kb and two SphI-SphI fragments of 21.5 and 6.kb respectively (Figure 2). The same results were obtained with the other
428
a b c d
M. SANCHEZ ET AL.
A
23.1-
9.4-
6.5-
4.3-
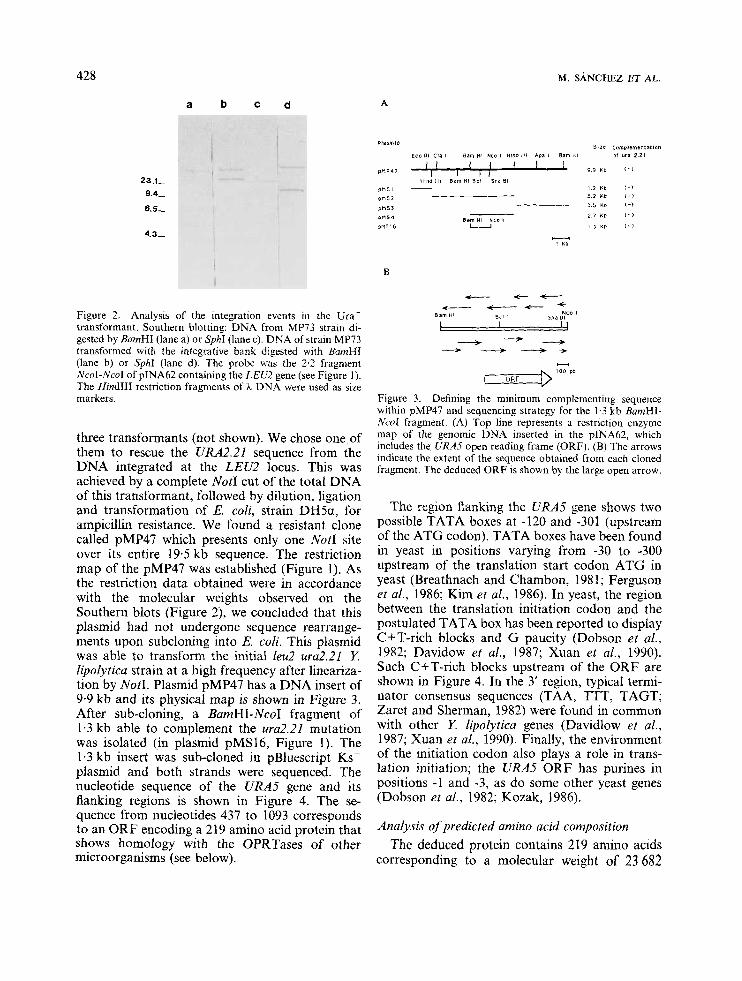
Figure 2. Analysis of the integration events in the Ura' transformant. Southern blotting: DNA from MP73 strain di- gested by BamHI (lane a) or Sphl (lane c). DNA of strain MP73 transformed with the integrative bank digested with BnrnHI (lane b) or SphI (lane d). The probe was the 2.2 fragment NcoI-NcoI of pINA62 containing the LEU2 gene (see Figure 1). The Hind111 restriction fragments of h DNA were used as size markers.
three transformants (not shown). We chose one of them to rescue the URA2.21 sequence from the DNA integrated at the LEU2 locus. This was achieved by a complete NotI cut of the total DNA of this transformant, followed by dilution, ligation and transformation of E. coEi, strain DHSa, for ampicillin resistance. We found a resistant clone called pMP47 which presents only one NotI site over its entire 19.5 kb sequence. The restriction map of the pMP47 was established (Figure 1). As the restriction data obtained were in accordance with the molecular weights observed on the Southern blots (Figure 2), we concluded that this plasmid had not undergone sequence rearrange- ments upon subcloning into E. coli. This plasmid was able to transform the initial leu2 ura2.21 Y. lipolytica strain at a high frequency after lineariza- tion by NotI. Plasmid pMP47 has a DNA insert of 9.9 kb and its physical map is shown in Figure 3. After sub-cloning, a BamHI-NcoI fragment of 1.3 kb able to complement the ura2.21 mutation was isolated (in plasmid pMS16, Figure 1). The 1.3 kb insert was sub-cloned in pBluescript Ks' plasmid and both strands were sequenced. The nucleotide sequence of the URA.5 gene and its flanking regions is shown in Figure 4. The se- quence from nucleotides 437 to 1093 corresponds to an ORF encoding a 219 amino acid protein that shows homology with the OPRTases of other microorganisms (see below).
B
-+--- c - t - t- +-----+
Barn HI NCO I - - - - +
- - + - - - - + + Y
,-FF--D loo pD
Figure 3. Defining the minimum complementing sequence within pMP47 and sequencing strategy for the 1.3 kb BarnHI- NcoI fragment. (A) Top line represents a restriction enzyme map of the genomic DNA inserted in the pINA62, which includes the URAS open reading frame (ORF). (B) The arrows indicate the extent of the sequence obtained from each cloned fragment. The deduced ORF IS shown by the large open arrow.
The region flanking the URAS gene shows two possible TATA boxes at -120 and -301 (upstream of the ATG codon). TATA boxes have been found in yeast in positions varying from -30 to -300 upstream of the translation start codon ATG in yeast (Breathnach and Chambon, 1981; Ferguson et al., 1986; Kim et al., 1986). In yeast, the region between the translation initiation codon and the postulated TATA box has been reported to display C+T-rich blocks and G paucity (Dobson et al., 1982; Davidow er al., 1987; Xuan et al., 1990). Such C+T-rich blocks upstream of the ORF are shown in Figure 4. In the 3' region, typical termi- nator consensus sequences (TAA, TTT, TAGT; Zaret and Sherman, 1982) were found in common with other Y. lipolytica genes (Davidlow et al., 1987; Xuan et al., 1990). Finally, the environment of the initiation codon also plays a role in trans- lation initiation; the URAS ORF has purines in positions -1 and -3, as do some other yeast genes (Dobson et al., 1982; Kozak, 1986).
Analysis of predicted amino acid composition The deduced protein contains 219 amino acids
corresponding to a molecular weight of 23 682

URAj GENE OF Y. LIPOLYTICA 429
1
80
C CAT CCC TCC CTC ACC TCC AAG CCT TAT T m OCA CCT CTC GCG TCT ?&A M T TTA TCT TCT CCT T U CTT CCA TAT TCA
TTA CCT CTA CCT TTC TGT ADA CAA TAT CAD GGT CTT C K CTT CCC TCT TTA ACT ?AT* CTG TCT ACA M T C M TCT CCC v
161 AAG TAA AM AM AM AM AAA ACC TCA ACC AM GGT ACT ACA ACT ACT TCC MT AAT cAC TCA AAG TCC CTA TTC C M CCA
242 GAA TAT TCA CAC AAG AAG TTC CTA CCA C K TTT G M ATG ACC CCC CTA CTG TCA CAT ACT ATT TGA CCT TCA AAA TAT AAC
323 ccc C M ATA TCC TK TCT ACC CU TCT CCA TCT CTT CAC CAT OK AAC TCT AAT GGC AGA AM A m ACT TAC CTT CAG CAT
1 404
Met Cln A h T y r L y s L y s A s p Phe L e u Asp L e u A h T h r Ly. T F G l U CCC ACT TTC CTA TCT CTT A X ACT ACT ACC ACA AT0 CAD CCT TAC AAG AAG CAC TTT CTG CAT CTG GCT ACC AAG TAC GAG
17 A h L e u L y s P h e Gly Ser Pha T h r L e u Lye sex c l y A r g T h r sex P z o Tyr P h e P h e As" L e u G l y L e u P h e Asn T h r GlY 485 GCC CTC AAG TTT CCC TCC T K ACC CTC AAG TCC GGC CCA ACT TCA CCC TAC TTC TTC MC C K GCT C K TTC AAC ACC CCA
44 5 6 6
71 647
Val 1\11 L e u Sex T h r Val Cly All Ser P h e A h Cln Val Ile Ile Asn SeT Gly V a l Glu P h a Asp Val I h Phe C l Y PTO CTC CCT CTG TCC ACT CTC CCA CCT TCC TTT CCC CAG CTG ATC ATC M C TCC CCA CTC CAD TTT CAC CTC ATT TTC CCT CCC
Ala T y r L y a Cly 11- Pro L e u Alr Ala Val T h r A l a A h L y a Ile Ala Clu L e u Gly C l y Clu L y s T y r A h T h r LYS ClU GCC TAC AAG CCT ATT CCT CTC CCC CCC CTC ACC GCC CCC AAC ATT GCC CAA TTG GCC CGA GAG AAG TAC GCT ACC
98 728
T y r All P h e Asn Arg L y s Clu Ale, L y s A s p Hi. G l y Clu c 1 y c ly A m Ile Val Cly Aln ser L e u L y s G l y L y s Lys Val TAC CCT TTC AAC CCA AM CAC CCC AAG CAC CAC CCT CAD GCA CCA M C ATT CTC CGA CCT TCT CTC AAG CGC AAG AAG GTC
125 809
152 890
L e u Ile Ile A a p Asp Val 11. T h r A h C l y T h r A h 11. Ly. clu A l a P h e See 11. 11. Asp Ala A#n C l y A h T h r Val CTC ATT ATC GAC CAC CTT A K ACC CCT CGT ACT CCT ATT AAG GAG GCC TTC TCA ATC ATT CAC CCA MC CCC GCT ACC CTC
Ser A h Val Val 11e Aln L e u Asp Arg C l n Clu T h r T h r Val Asp Ser P r o L y e Ser A h Val Gln Val Val Sex G h T h r TCT CCT GTG CTC ATT CCA CTC CAT CCA CAG CAD ACC ACT GTT CAC ACC CCC AAG TCT CCC G W CAC CTT CTT TCC C M ACC
179 971
206 1052
1133
1214 TCA ACT TCT ACC CAT TGA TCT TAA ATC TAT CCC CTA TAG CCA ATA ACC ATG 0
T y r A m Val P r o Val L e u A m 11. Phe A m L e u As- Asp val 11. C l n T y r T h r Asp C l y Ile L e u T b r Clu ASP Clu LY. TAC AAC CTC CCT CTT C K AAC ATC TTC AAC CTC MC CAC CTC ATC CAD TAC ACC CAT CCC A K TTG ACC GAG CAC GAG
LYS LYS Ile CIY C l n T y r * g Asp Cln T y r Sar P r o Ly. Clu *** AAG AAG ATT GAG CAG TAC CCA CAT CAC TAC TCC CCT AAG GAG TAG ATC CGT CCG ACA CTA CGA GTA T M AAG ACT ACA ATA
ATT GAA CCC TGA GTT CCA TCT A X TCA TCC TTA TTT CTA CTT ACG CTA C M CTA GAG TAC ATA CAG TAC ATA CGT ACA TAC
- -
79
160
241
322
( 0 3
16 484
43 565
70 646
97 727
124 808
151 889
178 970
205 1051
220 1132
1213
1265
Figure 4. Nucleotide sequence of the yeast Y. lipolyrira URA5 gene region isolated from wild-type strain W29 (plasmid pMS-16). The sequence of one DNA strand and the deduced amino acid sequence for OPRTase are shown. Nucleotides are numbered from the 5' end to the 3' end of the sequenced fragment. Amino acids are numbered from the first putative ATG in the large open reading frame (ORF) at the left of the figure. (V) represents the position of the major transcription start. The signals discussed in the text are underlined
daltons. Using the method of Bennetzen and Hall (1982), we found a codon bias value of 0.49, which corresponds to an intermediate protein abundancy.
We have compared the deduced protein se- quence (219 amino acids) with the OPRTases of E. coli (Poulsen et al., 1983), Podospora anserina (Turcq and Bequeret, 1987), D. discoideum (Jacquet et al., 1988) and S. cerevisiae (de Montigny et al., 1989). The results are shown in Figure 5. The sequences are significantly similar, 58% identical amino acids (91% conservative sub- stitutions), with the URAS gene of S. cerevisiae and 49% identical amino acids (84.5% conservative substitutions) with the pyrE gene of E. coli, indi- cating that the isolated gene corresponds to the OPRTase.
Transcription of the URAS gene Northern blotting using the 0.6 kb BclI-SnaBI
fragment as a a probe revealed one single band of about 0.94 kb (Figure 6) in both the poly(A)+ (lane a) and total (lane b) RNA from a wild-type strain. These results indicate that the transcript is a polyadenylated RNA and that the BclI-SnaBI fragment does not include another transcriptional
YlUF'.AS s c m 5 PaURA5 E c p y r E Ddpyr56G
Y 1 m 5 s c m 5 P a m S ECPYXE Ddpyr56G
I1-5 s c m 5 P a m 5 ECpyT-E Ddpyr56G
Figure 5. Predicted amino acid sequences of the OPRTase of Yurrowiu lipolytica (YIURA5) , Succlzarom,vces cerevisiae (ScURASj. Escherichiu coli (EcpyrE), Podospora anserina (PuURAS) and Dictyostelium discoideum (Ddpyr56G). Gaps have been introduced to give the best alignment. Identical residues (asterisks) in the five polypeptides and conservative amino acid substitutions (dots) are indicated. Shadows indicate identities betwen Y. lipolyricu and S. rerevisiue
430 M. SANCHEZ ET AL.
a b c
9.49- 7.4 6 -
4.40-
2.37-
1.35-
0.94 -
0.24-
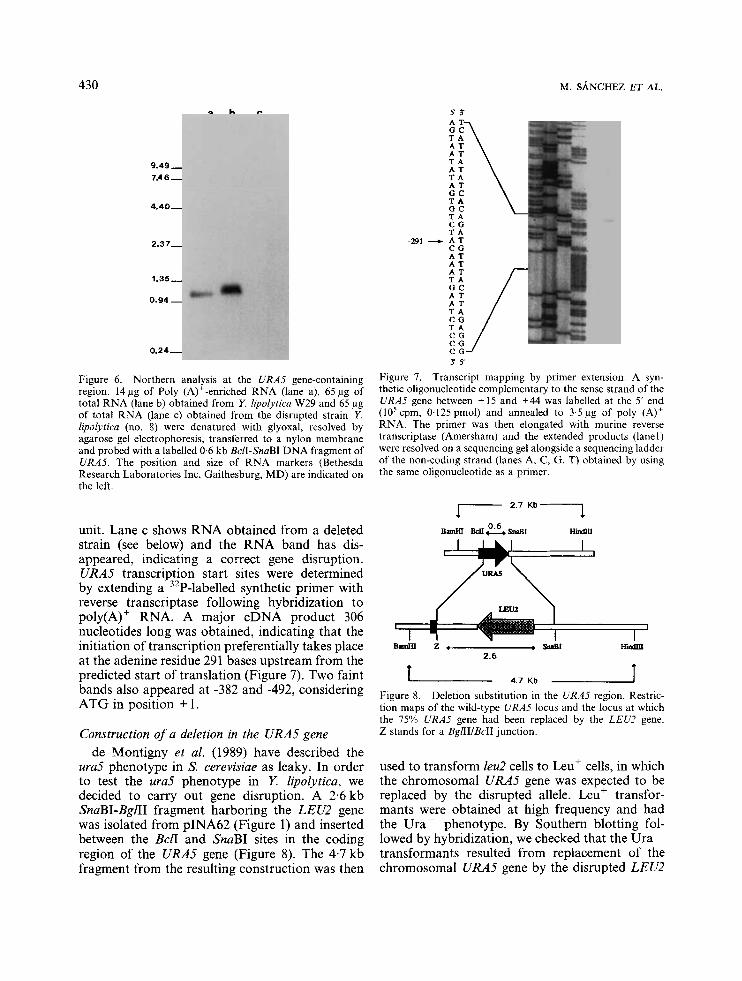
Figure 6. Northern analysis at the URAS gene-containing region. 14 pg of Poly (A)'-enriched RNA (lane a), 65 pg of total RNA (lane b) obtained from Y lipolytica W29 and 65 pg of total RNA (lane c) obtained from the disrupted strain Y. lipolytica (no. 8) were denatured with glyoxal, resolved by agarose gel electrophoresis, transferred to a nylon membrane and probed with a labelled 0.6 kb BclI-SnaBI DNA fragment of URAS. The position and size of RNA markers (Bethesda Research Laboratories Inc, Gaithesburg, MD) are indicated on the left.
unit. Lane c shows RNA obtained from a deleted strain (see below) and the RNA band has dis- appeared, indicating a correct gene disruption. URAS transcription start sites were determined by extending a 32P-labelled synthetic primer with reverse transcriptase following hybridization to poly(A)' RNA. A major cDNA product 306 nucleotides long was obtained, indicating that the initiation of transcription preferentially takes place at the adenine residue 291 bases upstream from the predicted start of translation (Figure 7). Two faint bands also appeared at -382 and -492, considering ATG in position + 1.
Construction of a deletion in the URAS gene de Montigny et al. (1989) have described the
ura.5 phenotype in S. cerevisiae as leaky. In order to test the ura.5 phenotype in Y. lipolytica, we decided to carry out gene disruption. A 2.6 kb SnaBI-BglII fragment harboring the LEU2 gene was isolated from pINA62 (Figure 1) and inserted between the BclI and SnaBI sites in the coding region of the URAS gene (Figure 8). The 4-7 kb fragment from the resulting construction was then
-29 1
5 ' s
T A A T G C T A G C T A C G T A A T C G A T
C G T A C G C G C G T5'
Figure 7. Transcript mapping by primer extension. A syn- thetic oligonucleotide complementary to the sense strand of the URAS gene between +15 and +44 was labelled at the 5' end (lo5 cpm, 0.125 pmol) and annealed to 3.5 pg of poly (A)' RNA. The primer was then elongated with murine reverse transcriptase (Amersham) and the extended products (lanel) were resolved on a sequencing gel alongside a sequencing ladder of the non-coding strand (lanes A, C, G, T) obtained by using the same oligonucleotide as a primer.
/- \ BrmHI 2 , e Snael Kmdm
2.6 - 4.7 Kb t Figure 8. Deletion substitution in the URAS region. Restric- tion maps of the wild-type URAS locus and the locus at which the 75% URAS gene had been replaced by the LEU2 gene. Z stands for a BgnIlBcII junction.
used to transform leu2 cells to LeuC cells, in which the chromosomal URA.5 gene was expected to be replaced by the disrupted allele. Leuf transfor- mants were obtained at high frequency and had the Ura- phenotype. By Southern blotting fol- lowed by hybridization, we checked that the Ura transformants resulted from replacement of the chromosomal URA.5 gene by the disrupted LEU2

URA5 GENE OF Y. LIPOLYTICA 43 1
a b c d e f g
23.1-
9.4- 6.5-
4.3-
2.3- 2.0-
Figure 9. Southern blot of total DNA from the deleted URA.5 strain. Wild-type DNA (lane b), disrupted strain no. 8 (lane c), disrupted strain no. 11 (lane d) digested with BamHI-HindII1. The probe was the 2.2 LEU2 (fragment NcoI-NcoI of pINA62, see Figure 1). Wild-type DNA (lane e), disrupted strain no. 8 (lane f). disrupted strain no. 11 (lane g) digested with BarnHI- Hindlll. The probe was the 0.6 kb URAS (fragment BclT-SnaBI of pMSl6, see Figure 3). The Hind111 restriction fragments of h DNA were used as size markers (lane a).
Figure 10. Phenotypes of ura5 mutants.The ura5 mutants and control strains were grown on complete medium (without uracil) at 28°C. Photographs were taken after 2 days of growth. ( I ) S. cerriisiae strain MSI; (2) Y. lipolytica strain MP-73; (3) deleted strain (no. 8) of Y. lipolytica transformed with pMS16, (4) deleted strain (no. 8) of Y. lipolytica; ( 5 ) S. cerevisiae strain MS1 transformed with pMS18.
construct (Figure 9). A clear Ura- phenotype was observed much tighter than for ura5 mutants of S. cerevisiue (de Montigny et al., 1989; Figure 10).
Expression of the Y. lipolytica URA5 in S. cerevisiae
If structural homology is representative of func- tional homology, the U M 5 homolog from Y. lipolytica should be expressed in S. cerevisiae. A BamHI-Hind111 DNA fragment of 2-7 kb (see Figure 3 ) , which contains the entire U M 5 gene, was cloned into YRp7 (Struhl et al., 1979, giving
rise to pMS18; Figure 1). This plasmid, together with YRp7 (as control), was used to transform the S. cerevisiae strain MS-I (leu2-3 his4-712 trpl ura5).
The desired transformants were selected on the basis of their expected Trp' phenotype. From ten Trp' transformants obtained with YRp7, none was Ura', as would be expected. From ten Trp' transformants obtained with pMS 18 derived from MS-1, all were Ura+, indicating that the URA.5 gene from Y. lipolytica was functionally expressed in S. cerevisiae, thus complementing the ura5 mutation (Figure 10). In order to confirm these results, plasmid segregation experiments were carried out as previously described (Rose et al., 1987; Shnchez et al., 1993). In all cases (50 trans- formants selected randomly), loss of the Trp+ character was accompanied by loss of the Ura+ character, confirming the functionality of the URA.5 gene of Y. lipolytica in S. cerevisiae.
DISCUSSION
By integrative transformation and complementa- tion of one Y. lipolytica uracil auxotroph uru2.21, a 1.3 kb genomic fragment has been cloned and sequenced. The protein sequence from the gene product shows similarities with the genes of other microorganisms coding for OPRTase (Figure 4). Thus, we propose renaming the mutation as ura5-21 and the gene as YLURAS. The presence of the peptide sequences Asp Asp Val and Thr Ala Gly Thr in the positions + 128 and + 132, strongly conserved among other phosphoribosyl trans- ferases (Hershey and Taylor, 1986), indicates an important role for these residues.
The signals for translation, transcription, termi- nation and polyadenylation agree with the consen- sus sequences for Y. lipolytica (Xuan et al., 1990) and for yeast in general (Zaret and Sherman, 1982). Only one RNA has been found to hybridize to our DNA fragment, either from poly(A)' RNA or from total RNA. Primer extension indicates that the initiation of the translation starts -291 bases upstream from the predicted AUG. This defines a presumptive 5' non-translated leader se- quence higher than the average found in yeast (Yoon and Donahue, 1992) but similar to the mRNA described for the URAS of S. cerevisiae (de Montigny et al., 1989). The full length of the messenger deduced (about 0.9 kb from the posi- tions of the start and stop signals for transcription)

432 M. SANCHEZ ET A L .
is consistent with the 0.94 kb determined by Northern blotting (Figure 6).
A leaky phenotype for the ura5 strains has been described in S. cerevisiae (de Montigny et a/., 1989). In order to analyse the ura5 phenotype in Y. lipolytica and to confirm the cloning of the gene, we have replaced the wild-type gene with a deleted allele. Several strains showing a ura5 phenotype in which a deletion in the URA5 gene occurs (see Figure 9) were isolated. The ura5 phenotype is also leaky in Y. lipolytica, although the mutation is less leaky than in S. cerevisiae (Figure 10). There are very few reports about the behaviour of Y. lipo1.vtica genes in other yeasts. Lopez et a/. (1994) have shown that SEC 14pyL is able to complement the lethality of secll null mutations in S. cerevi- siae. Our data also show clearly that URA5pYL was able to substitute for URASp in S. cerevisiae (Figure 10).
ACKNOWLEDGEMENTS We wish to thank C. Gaillardin for providing the Y. lipolytica gene bank and Y. lipolytica strains, F. del Rey for S. cerevisiae 5573-7c strain, S . Frisbie for revising the English version of this manuscript, C. Gancedo for critical reading of the manuscript and M. C. Lopez and J. C. Torres-Guzman for helpful discussions. This work was partially sup- ported by grants from the DGICYT (BI092-0304) and CEE (BIOT-CT 91-0267 DSCN).
REFERENCES Barns, S. M., Lane, D. J., Sogin, M. L., Bibeau, C. and
Weisburg, W. G. (1991). Evolutionary relationships among pathogenic Candida species and relatives. J. Bacteriol. 173, 2250-2255.
Barth, G. and Weber, H. (1985). Improvement of sporu- lation in the yeast Yarrowia lipolytica. Ant. van Leeuwenhoek J. Microbiol. Serol. 51, 167-1 77.
Becker, D. M. and Guarente, L. (1991). High efficiency transformation of yeast by electroporation. Methods in Enzymology 194, 182-1 87.
Bennetzen, J. L. and Hall, B. D. (1982). Codon selection in yeast. J. Biol. Chem. 257, 3026-3031.
Biggin, M. D., Gibson, T. J. and Hong, G. F. (1983). Buffer gradient gels and 35S label as an aid to rapid DNA sequence termination. Proc. Natl. Acad. Sci. USA 80, 3963-3965.
Boy-Marcotte, E., Vilaine, F., Camonis, J. and Jacquet, M. (1984). A DNA sequence from Dictyostelium discoideum complements ura3 and ura5 mutations of Saccharomyces cerevisiae. Mol. Gen. Genet. 193, 40-13.
Breathnach, R. and Chambon, P. (1981). Organization and expression of eukaryotic split genes coding for proteins. Annu. Rev. Biochem. SO, 349 -383.
Davidow, L. S. , Apostolakos, D., O'Donnell, M . M.. Proctor, A. R., Ogrydziak. D. M., Wing, R. A,. Stasko. 1. and DeZeeuw, J. R. (1985). Integrative transformation of the yeast Yarrowin Iipolj~rica. Curr. Genet. 10, 3 9 4 8 .
Davidow, L. S., Kaczmarek, F. S.. DeZeeuw, J . R., Conlon, S. W., Lauth, M. R., Pereira, D. A. and Franke, A. E. (1987). The Yarrowin Iipolrticrr LEI12 gene. Curr. Genet. 11, 377-383.
de Montigny, J., Belarbi, A,. Hubert, J. L. and Lacroute. F. (1989). Structure and expression of the URA5 gene of Saccharomyces cerevisiae. Mol. Gen. Gcnct. 215, 455462.
Dobson, M. J., Tuite, M. F., Roberts, N. A., Kingsman. A. J . and Kingsman, S. M. (1982). Conservation of high efficiency promoter sequences in Scrc.c,/iciroriilc.c.\ cerevisiae. Nucl. Acids Res. 10, 2625-2631.
Ferguson, J., Ho, J. Y . , Peterson. T. A. and Reed, S. I . (1986). Nucleotide sequence of the yeast cell division cycle start genes CDC28. CDC36. and CDC3Y. and ii structural analysis of the predicted products. " V 4 c . l . Acids Res. 14, 6681-6697.
Gaillardin, C., Ribet, A-M. (1987). LEU2 directed expression of beta-galactosidase activity and phleo- mycin resistance in Yarrowin lipolj~ricn. Curr. Gu7t~t.
Hanahan, D. (1983). Studies on transformation o f Escherichia coli with plasmids. J . Mol. Biol. 166,
Hershey, H. V. and Taylor, M. W. (1986). Nucleotide sequence and deduced amino-acid sequence of Escherichia coli adenine phosphoribosyltransferase and comparison with other analogous enzymes. Genc,
Higgins, D. G. and Sharp, P. M. (1988). Clustal: ;I
package for performing multiple sequence alignments on a microcomputer. Gene 73, 237-244.
Jacquet, M., Guilbaud, R. and Garreau. H . (1988). Sequence analysis of the DdPYR5-6 gene coding for U M P synthase in Dictyosteliurn di.rroide~it/i in coni- parison with orotate phosphoribosyl transferases and OMP decarboxylases. Mol. Gen. Gener. 211, 441 4 4 5 .
Jones, M. E. (1980). Pyrimidine nucleotide bioaynthesis in animals: genes, enzymes and regulation of UMP biosynthesis. Annu. Rev. Biochetn. 49, 253-219.
Kim, S., Mellor, J., Kingsman. A. J . and Kingsman. S. M. (1986). Multiple control element in the TRPI promoter of Saccharomyes cerevisine. Mol. Cell. Biol. 6,42514258.
Kozak, M. (1986). Bifunctional messenger RNAs in eukaryotes. Cell 44, 283-292.
Lopez, M. C., Nicaud, J. M., Skinner, H. B., Vergnolle. C., Kader, J. C., Bankaitis, V. A. and Gaillardin, C. (1994). A phosphatidylinositol/phosphatidylcholine transfer protein is required for the diferentiation of
11, 369-375.
557-580.
43, 287-293.

URAi GENE OF Y. LIPOLYTICA 433
the dimorphic yeast Yarrowia lipolytica from the yeast to the mycelial form. J. Cell. Biol. 124, 113-127.
Makoff, A. J. and Radford, A. (1978). Genetics and biochemistry of carbamoyl phosphate biosynthesis and its utilization in the pyrimidine biosynthetic path- way. Microbiol. Rev. 425, 307-328.
McCarroll, R., Olsen, G. J., Stahl, Y. D., Woese, C. R. and Sogin, M. L. (1983). Nucleotide sequence of the Dictyostelium discoideum small-subunit ribosomal ri- bonucleic acid inferred from the gene sequence: evo- lutionary implications. Biochemistry 22, 5858-5868.
Meissner, P. S., Sisk, W. P. and Berman, M. L. (1987). Bacteriophage cloning system for the construction of directional cDNA libraries. Proc. Natl. Acad. Sci. USA 84,41714175.
Pearson, W. R. and Lipman, D. J. (1988). Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85, 2444-2448.
Percival-Smith, A. and Segall, J. (1984). Isolation of DNA sequences preferentially expressed during sporulation in Saccharomyces cerevisiae. Mol. Cell. Biol. 6, 2443-245 1.
Poulsen, P., Jensen, K. F., Valentin-Hansen, P., Carlsson, P. and Lundberg, L. G . (1983). Nucleotide sequence of the Escherichia coli PyrE gene and of the DNA in front of the protein coding region. Eur. J . Biochem. 135, 223-229.
Rose, M. D., Novick, P., Thomas, J. H., Botstein, D. and Fink, G. R. (1987). A Saccharomyces cerevisiae genomic plasmid bank based on a centromeric- containing shuttle vector. Gene 60, 237-243.
Sambrook, J. E., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual, 2nd edn. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Sanchez, M., Iglesias, F. J., Santamaria, C. and Dominguez, A. (1993). Transformation of Kluyvero-
myces lactis by electroporation. Appl. Environ. Micro- biol. 59, 2087-2092.
Sanger, F., Nicklen, S. and Coulson, A. R. ( 1 977). DNA sequencing with chain-terminating inhibitor. Proc. Natl. Acad. Sci. USA 74, 5463-5467.
Sherman, F., Fink, G. R. and Lawrence, C. (1977). Methods in Yeast Genetics. Cold Spring Harbor Labo- ratory, Cold Spring Harbor, NY.
Souciet, J. L., Nagy, M., Le Govar, M., Lacroute, F. and Potier, S. (1989). Organization of the yeast URA2 gene: identification of a defective dihydroorotase-like domain in the multifunctional carbamoylphosphate synthase-aspartate transcarbamylase complex. Gene
Struhl, K., Stinchcomb, D. T., Scherer, S. and Davis, R. W. (1979). High-frequency transformation of yeast: Autonomous replication of hybrid DNA mol- ecules. Proc. Natl. Acad. Sci. USA 76, 1035-1039.
Turcq, B. and Bequeret, J. (1987). The URAS gene of the filamentous fungus Podosporu anserina: nucleotide sequence and expression in transformed strains. Gene 53, 201-209.
Xuan, J. W., Fournier, P. and Gaillardin, C. (1988). Cloning of the LYSS gene encoding saccharopine dehydrogenase from the yeast Yarrowia lipolytica by target integration. Curr. Genet. 14, 15-21.
Xuan, J. W., Fournier, P., Declerek, N., Chasles, M. and Gaillardin, C. (1 990). Overlapping reading frames at the LYSS locus in the yeast Yarrowia lipolvtica. Mol. Cell. Biol. 10, 4795-4806.
Yoon, H. and Donahue, T. F. (1992). Control of translation initiation in Succharomyces cerevisiae. Mol. Microbiol. 6, 1413-1419.
Zaret, K. S. and Sherman, F. (1982). DNA sequence required for efficient transcription termination in yeast. Cell 28, 265-273.
79, 59-70.
![Optimization of lipid production with a genome-scale model of ......reconstructions for P. pastoris and P. stipitis [8, 9] and for the oleaginous yeast Yarrowia lipolytica, for which](https://img.pdfslide.us/doc/110x75/60af29819c322d48513532a7/optimization-of-lipid-production-with-a-genome-scale-model-of-reconstructions.jpg)







![Lipid production by the oleaginous yeast Yarrowia lipolytica ......minerals, and vitamins, which are crucial for fermenta-tion [18]. Moreover, molasses is used as the main sub-strate](https://img.pdfslide.us/doc/110x75/612199446e91810e306d6787/lipid-production-by-the-oleaginous-yeast-yarrowia-lipolytica-minerals-and.jpg)




















