Embed Size (px)
Citation preview

Mol Gen Genet (1991) 226:250-256 002689259100099N
© Springer-Verlag 1991
Cloning and nucleotide sequence of the hemA gene of Agrobacterium radiobacter M. Drolet and A. Sasarman
Department of Microbiology and Immunology, Universit~ de Montr6al, Montr6al, Qu6bec, Canada, H3C 3J7
Received August 31, 1990 / November 16, 1990
Summary. The hemA gene of Agrobacterium radiobacter ATCC4718 was identified by hybridization with a hemA probe from Rhizobium meliloti and cloned by comple- mentation of a hemA mutant of Escherichia coIi K12. E. coli hemA transformants carrying the hemA gene of Agrobacterium showed 6-aminolevulinic acid synthetase (6-ALAS) activity in vitro. The hemA gene was carried on a 4.4 kb EcoRI fragment which could be reduced to a 2.6 kb EcoRI-SstI fragment without affecting its complementing or 6-ALAS activity. The sequence of the hemA gene showed an open reading frame of 1215 nucle- otides, which could code for a protein of 44361 Da. This is very close to the molecular weight of the HemA pro- tein obtained using an in vitro coupled transcription- translation system (45000 Da). Comparison of amino acid sequences of the 6-ALAS of A. radiobacter and Bradyrhizobium japonicum showed strong homology be- tween the two enzymes; less, but still significant, homol- ogy was observed when A. radiobacter and human 6- ALAS were compared. Primer extension experiments en- abled us to identify two promoters for the hemA gene of A. radiobacter. One of these promoters shows some similarity to the first promoter of the hemA gene of R. meliloti.
Key words: cS-Aminolevulinic acid synthetase (6-ALAS) - hemA gene - Agrobacterium radiobacter - Tetrapyr- roles - Vitamin B12
Introduction
6-Aminolevulinic acid (6-ALA) is the first intermediate of tetrapyrrole synthesis in all organisms. Heine, chloro- phyll, and vitamin B12, which is produced only by some microbes, are all derivatives of this pathway. 6-ALA is synthesized either in three steps, via a five-carbon (5C) pathway from the intact carbon skeleton of glutamate (Castelfranco and Beale 1983) or in one step, by the condensation of glycine with succinyl-CoA (Granick and Beale 1978).
Offprint requests to .' A. Sasarman
In plants (Castelfranco and Beale 1983), algae (Mayer et al. 1987), cyanobacteria, methanogenic bacteria (Av- issar et al. 1989) and some purple bacteria, Escheriehia coli (Li et al. 1989a), probably Salmonella typhimurium (Elliot 1989) and Bacillus subtilis (Petricek et al. 1990), and many others (Avissar et al. 1989), 6-ALA is synthe- sized from glutamate. The hemA gene of E. coli K12 was the first gene of this pathway to be cloned and sequenced (Drolet et al. 1989; Li et al. 1989b; Verkamp and Chelm 1989). As shown by Avissar and Beale (1989), this gene codes for glutamyl-tRNA dehydrogenase, the second enzyme of the 5C pathway.
In animals, yeast, fungi and other bacteria the hemA gene encodes 6-aminolevulinic synthetase (6-ALAS) which catalyzes the condensation of glycine with succin- yl-CoA to form 6-ALA. Several hemA genes of this 4C pathway have been cloned and sequenced, among them the hemA gene of Rhizobium meliloti (Leong et al. 1982; 1985), Bradyrhizobium japonicum (Robertson McClung et al. 1987) and Rhodobacter eapsulatus (Biel et al. 1988; Hornberger et al. 1990). In R. sphaeroides two genes re- sponsible for 6-ALA synthesis (hemA and hemT) have been isolated Tai et al. 1988). In eucaryotic organisms, several hemA genes have been cloned and sequenced, namely the hemA genes of Saecharomyces cerevisiae (Ar- rese et al. 1983; Bard and Ingolia 1984; Urban-Grimal et al. 1984; 1986), chicken liver (Borthwick et al. 1984; 1985; Maguire etal. 1986; Pirola etal. 1984), mouse liver and anemic spleen (Schoenhaut and Curtis 1986), rat (Srivastava et al. 1988), and human (Bawden et al. 1987). Comparison of amino acid sequences of the var- ious cS-ALAS revealed close homology between them.
The regulation of the tetrapyrrole pathway has been studied mainly in the photosynthetic bacterium, R. sphaeroides (for a review see Lascelles 1975). In this bac- terium, heine (Burnham and Lascelles 1963) and Mg- protoporphyrin (Yubisui and Yoneyama 1972) inhibit the activity of 6-ALAS, while trisulfides activate this enzyme (Sandy et al. 1975). It seems likely that the main regulatory mechanisms of the tetrapyrrole pathway in most bacteria act at the level of 6-ALA synthesis (Las- celles 1975), but we know very little about them.
Agrobacterium radiobacter ATCC 4718 is a good pro- ducer of vitamin B12, as shown in our laboratory (un-

published). The presence of the cobalamin pathway in this organism allows the study of the relationship be- tween the regulation of vitamin B12 and of heme synthe- sis, without the interference of bacteriochlorophyll syn- thesis, as in Rhodobacter. In order to study this relation- ship, we decided to clone the hemA gene of A. radio- bacter and to determine its nucleotide sequence. As a preliminary step to the study of the regulation of the hemA gene of A. radiobacter, we also identified two pro- moters for this gene.
Materials and methods
Bacterial strains and plasmids. The bacterial strains and plasmids used in the present study are listed in Table 1.
Media. Strains of E. coli K12 were routinely maintained on brain heart infusion agar supplemented, when re- quired, with appropriate antibiotics (10 gg/ml tetracy- cline or 50 pg/ml ampicillin), or with 6-ALA at 50 btg/ml. Strains of A. radiobacter and R. meliloti were routinely maintained on NZ-amine (Miller 1972) or LB media. YT medium supplemented with 0.1 mM isopropyl fi-I)- thiogalactopyranoside (IPTG) and 0.004% X-Gal (5- bromo-4-chloro-indolyl-~-D-galactopyranoside), was used for cloning with M13mp18 and M13mp19 vectors.
Enzymes and chemicals. Restriction endonucleases and DNA modification enzymes were purchased from Beth- esda Research Laboratories (Gaithersburg, Md.), Phar- macia PL Biochemicals, and New England Biolabs (Bev- erly, Mass.). Various chemicals used in cloning, DNA
251
sequencing, and other molecular biology experiments were from Sigma (St. Louis, Mo.), Bethesda Research Laboratories, and Boehringer Mannheim Biochemicals (Indianapolis, Ind.). L-[3SS]methionine was purchased from Amersham International (Arlington Heights, Ill.), and ~-[32p]nucleotide triphosphates were from ICN Pharmaceuticals (Irvine, Calif.).
DNA isolation. Isolation of chromosomal DNA of A. radiobacter and R. meliloti was performed as described by Silhavy et al. (1984). Large scale isolation of plasmid DNA was performed by the alkaline lysis method (Man- iatis et al. 1982). For rapid analysis of recombinant plas- raids, the method of Holmes and Quigley (1981) was used.
DNA cloning. Restriction digests, ligations and transfor- mations were done essentially as described by Maniatis et al. (1982).
Hybridization. Southern blot experiments were per- formed essentially as described by Maniatis et al. (1982). For high stringency conditions, the last wash in 0.1 x SSC and 0.1% SDS was performed at 55 ° C for 30 min and under low stringency this wash was done at 37 ° C for 10 min.
Catalase test. The qualitative catalase tests were per- formed as previously described by Sasarman etal. (1970).
f -ALAS assay. Bacteria (E. coli strains) were grown aer- obically in brain heart infusion broth to an O.D. of
Table 1. Bacterial strains and plasmids Relevant characteristics Source
Strains
Eseherichia coli SASX475 SAS2025 SAS2026 SAS2027 Rhizobium meliloti Agrobacterium radiobacter
Plasmids
pUC18 pUC19 pSAS1421
pSASI423
pSAS1424
pSAS1425
pSAS1426
hemA, recA, Tc r SASX475/pSAS1424, HemA + SASX475/pSAS1425, HemA + SASX475/pSAS1426, HemA + Prototroph
Prototroph
Apr ap r NheI-- Sau3A HemA + fragment from E. coli K12,
inserted in pUC18 Sall HemA + fragment from R. meliloti,
inserted in pBR322 EcoRI HemA + fragment from A. radiobacter,
inserted in pUC19 SstI-EcoRI HemA ÷ fragment from A. radiobacter,
inserted in pUC 19 Sau3A HemA + fragment (1.4 kb) from A. radiobacter, inserted in pUCJ 8
This laboratory This work This work This work ATCC 10310
ATCC 4718
Drolet et al. (1989)
This work
This work
This work
This work
Tc, tetracycline; Ap, ampicillin

252
0.6 at 600 nm. a-ALAS activity in sonic extracts of bac- teria was assayed by the procedure of Burnham (1970). Protein content was determined by the biuret method using human serum albumin as standard.
DNA sequencing. Nucleotide sequencing by the dideoxy chain termination method of Sanger et al. (1977) was carried out using phages M13mp18 and M13mp19 and the synthetic 17mer oligonucleotide primer, purchased from Pharmacia PL Biochemicals.
RNA isolation. RNA was extracted and purified from E. coIi K12 and A. radiobacter as described by Aiba et al. (1981).
Primer extension analysis. For the mapping of the 5' end of the hemA mRNA, the following protocol was used. Fifty micrograms of RNA was resuspended in 20 gl of a solution of 20 mM TRIS-HC1 pH 8.3,200 mM NaC1, 0.1 mM EDTA and 1 pmol of an oligonucleotide. After heating for 3 rain at 100°C the samples were cooled slowly to room temperature over a period of 45 rain. The mixture was then precipitated with ethanol and resuspended in 50 mM TRIS-HC1 pH 8.3, 5 mM MgCI2, 5 mM DTT, 50 mM KC1, 50 units RNAguard (from Pharmacia PL), 1.25 gg actinomycin D, 2 mM each ofdATP, dTTP and dGTP, 40 gCi ofc~ [32p]-dCTP and 20 units of AMV reverse transcriptase (Pharmacia PL). After incubation for 60 rain at 45 ° C, 2 mM cold dCTP was added, and incubation continued for 1 h. Fi- nally, 17 gl of a stop solution from the Sequenase kit was added to stop the reaction. We used the same syn- thetic oligonucleotide (complementary to nucleotides 632-613, see Fig. 2) for primer extension and for DNA sequencing of the corresponding region of the hemA gene.
In vitro plasmid DNA-directed translation. Proteins en- coded by the different recombinant plasmids were de- tected using an in vitro coupled transcription-translation system of E. coli purchased from Amersham Internation- al. The proteins were separated in 12% acrylamide gels according to Laemmli (1970).
Results
Cloning the hemA gene of A. radiobacter
In order to clone the hemA gene of A. radiobacter, a DNA probe was obtained from the corresponding gene of R. meliloti isolated in our laboratory (unpublished). The probe was hybridized under low stringency to the chromosomal DNA of A. radiobacter digested with ei- ther BamHI, PstI, SaII, EcoRI or HindIII. A positive signal was obtained with an EcoRI fragment of about 4 kb and, in the case of the other endonucleases, with fragments larger than 10 kb (results not shown). Since only one hybridization signal was obtained for each of the restriction endonuclease digestions, it was concluded that only one hemA gene was present in A. radiobacter. A large quantity of chromosomal DNA of A. radiobacter
was subsequently digested with EcoRI and following electrophoresis in a low melting temperature agarose gel, the gel region containing DNA of around 4 kb in size was isolated and the DNA was extracted. The DNA was ligated to phosphatase-treated pUC19 and used to transform the E. coli K12 hemA mutant, SASX475. Ap- proximately 1000 ampicillin-resistant transformants were obtained, and analysis of the plasmids carried by 20 of them indicated that about two-thirds contained inserts of 4-5 kb. Only four transformants were able to complement the hemA mutant, as shown by the for- mation of large colonies and production of catalase (heme-deficient mutants of E. coli form tiny colonies on solid media and are not able to produce catalase since heme is the prosthetic group of this enzyme; see Sasarman etal. 1968a, b). The presence of the hemA gene in these transformants was confirmed by determi- nation of a-ALAS activity in cellular extracts (see be- low).
Analysis of the plasmids carried by the four HemA ÷ transformants showed that all possessed an EcoRI insert of 4.4 kb (Fig. 1) which was oriented in one direction in two plasmids and in the opposite orientation in the other two. The EcoRI insert of one of these, pSAS1424, was extracted from an agarose gel and used as a probe for Southern blot analysis. Hybridization under high stringency was done with chromosomal DNA of A. ra- diobacter, digested with the same enzymes as used in the first Southern blot experiment with the R. meIiloti probe. The same hybridization patterns were obtained as with the R. meliloti probe, but with stronger signals, as expected for homologous DNA (results not shown).
To determine the location of the hemA gene on the EcoRI fragment, subfragments obtained with either SstI, KpnI or HpaI were recloned into the pUC18 and pUC19 vectors and transformed into SASX475. The smallest
I I I I I I I I I I 0 1 2 3 4 4.4 kb
Hem A + I I
Hem A -+ I I
Hem A- I !
rt-
Fig. 1. Comptementation studies. HemA +, complementation of the hernA mutation by the corresponding fragment, regardless of its orientation with respect to the fl-galactosidase promoter of the pUC vectors; HemA +, complementation of the hemA mutation only when the gene is in the same orientation as the fl-galactosidase promoter of the pUC vectors; HemA-, absence of complementa- tion in both orientations
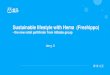
253
f ragment still able to complemen t the hemA m u t a n t after ~ i so TCGCGATAAT AGC/~TAGCC CGGCTCGGAA GCA'PI'GCCGA TC~GCACC GCCGACACCG 120 2 days i n c u b a t i o n was an EcoRI-SstI insert of 2.6 kb eeGATeGerG eGee~erGe eGeGeeGeGA AeAa"rAeeeG rGATGGeree Aee~T~eA
(Fig. 1), which gave a positive test in bo th or ientat ions . 18o Fur the r subc lon ing experiments with the EcoRI frag- GcGeeGG,,'a eGGCAeeGAT eG~GeGcce a~r~ecGTcr GeGTGeAGe'r TGe~GeGAe240
men t were per formed using par t ia l Sau3A digestion, i~IAGGCCGACGA GCGCAAATAT GATTGCC'I'*I~ ~TCATGAAAC T~CTCCAAC G~IACAATGA300
Two clones were ob ta ined that gave full complementa - GCTCTTCGGA TCGGTAGGCG TCCCATATTT ATCTGCGGCG ACTGCTACCT CCGACCGCCA360 t ion after i ncuba t i on for 1 day and accumula ted por- eAGAT~^ ACAA'I~CGAT GTCG~'I~AGG GAAGCCTCAT ~FI~CCAGGCA ~I~CCTGCGAG
420 phyrins. One of the clones was able to complemen t CGTa~,a~TCGA TTGTGTGCCA GACAACGGCG CGGTGCGAAA AAGGTTCCAA ATCTCGCGGT
P~-~ 480 SASX475 in both orientations, whereas the other, hav- ~TTGATGT GAATCAATTT TGTACTTTTC ATCAGAGTCT ATAAACGGTT TATTGG~ATG
540 ing the shortest insert (1.4 kb), failed to complemen t G'I~C~%.CA ACGGCG~.'I~ GACGCAGACG TGG~'~I~KAGC AGCTCGCa~.G GGTTGACGGT
~..~ 3 A ~ 600 when inserted in the reverse orientation (Fig. 1). This CCCGGTGTCG AGGCAGAAAA ACGATCTGGT CAATTCTGGA GCATGGCATG GACTTCGAGG
result al lowed us to determine the direct ion of t ranscrip- ,~ D F E 660 CATTTTTCAC AACGGAACTG CAAAGCCTGC ATTCTGAGGG ACGCTATCGC GTTTTTGCGG t i o n o f t h e h e m A g e n e o f A , radiobacter as being f rom ^ F F • T E L Q s ,. , s E G ~ Y R v F A
the r igh t -hand EcoRI site (Fig. 1) toward the KpnI and 720 ATATCGAACG CCAGCAGGGC AATTTTCCCC GCGCGACACG GTACAACGCC AATGGCCAAC
BglII sites. This result was conf i rmed subsequent ly by D ~ ~ ~ ~ ~ G ~ ~ ~ ~ A • ~ ~ ~ A ~ G 780
mapp ing the 5' end of the hemA m R N A (see below). GCAAGGACGT AACCGTCTGG TGCTCCAACG ACTATCTGGG CATGGGCCAG AACCCGAAAG
The presence of the hemA gene in t r ans fo rman t s was R K D V T V W e S N D Y L G M G Q N P K 840 by TCATCGAAGC CATGAAAGCC GCTATCGATC ACTGTGGCGC GGGTGCGGGA GGCACCCGGA
c o n f i r m e d t h e d e t e r m i n a t i o n o f g - A L A S activity i n v i E A M K A A I D H C e A e A e G T R
900 cellular extracts as described by B u r n h a m (1970). 6- A~C~GG TACCAACCAT TATCACGTCC TGCTTGAACA GGAACTTGCC GACCTCCATG
A L A S activity, which is absent in E. c o l i K ] 2 as this , ~ s G ~ ~ , ~ ~ v '~ ~ ~ ~ ~ '. A D ~ , 960
bac te r ium uses the g lu tamate pa thway for the biosynthe- GCAAGGAATC GGCGCTGATC TTCACCTCGG GTTATGTGTC AAACTGGGCC ACACTCGGCA sis of cS-ALA, was easy to determine in the hemA m u t a n t ~ ~ z s A ~. ~ ~ • s G V V S N W A ~ Z G 1020
hyb pl CGCTTGGCCA GAAAATTCCG GGCCTGATCA TTTTCTCGGA TGCGCTCAAC CATGCATCGA S A S X 4 7 5 h a r b o u r i n g v a r i o u s hemA + r i d a s m i d s • L G Q K I P e L I I F S D A L N H A S
1080 (Table 2). The higher ~5-ALAS activity observed for TGATCGAAGG CATCCGTTAT GGCCG'I~GCG AGCGGGTTAT CTGGAAACAC AACGATCTCG
SAS2027 is p robab ly due to the fact tha t the enzyme ~ I E G I R Y G R C E R V I W K H N D L 1140
in this s train appears to be synthesized unde r the cont ro l AAGATCTCGA AGCCAAGCTC AAGGCTGCCG ATCCGAACGC GCCGAAGCTG ATCGCTTTCG o f t h e l a c p r o m o t e r o f t h e p U C 1 8 v e c t o r . ~ ~ '" ~ A ~ ,~ K A A D ~ N A ,' ~ ~ ~ A 1200
by AATCCGTTTA TTCCATGGAT GGCGATATCG CGCCGATCAA GGAAATCTGC GATCTGGCCG Sequencing the hemA gene was per formed the di- ~ s v Y N M D G D I A P I K E I C D L A
1260 deoxy chain termination method of Sanger et al. (1977). ACCGTTATGG CGCCATGACC TATCTCGATG AAGTTCATGC CGTCGGCATG TATGGCCCGC
Both s t rands were sequenced f rom the NruI site, which D R Y G A M T Y L D E V H A V G M Y G P 1320
is 587 bp ups t ream f rom the start codon, to the Sau3A GCGGCGGCGG CATTGCCGAG CGCGAAGGTC TCATGGATCG CCTGACGATC ATCG/b~GGAA
site, which is 130 bp downs t r eam f rom the stop codon R G G G I A E R E G L M D R L T I I E G 1380 (1935 ) CGCTCGGCAA GGCCTTCGGC GTTATGGGCG GCTATATTAC CGGGTCCACG GCGGTCTGCG nucleotides. We sequenced along stretch of nucle- T L G K A F G V M U G Y I T G S T A V C
1440 otides ups t ream of the start codon to enable compar i son ACTTCATCCG TT~-FFI'CGCC TCCGGTTTCA TCTTCACGAC GGCCCTGCCG CCGTCGCTCG
w i t h t h e 5 ' region of the hemA gene of R. meliloti and D F I a S F A S e F I F T T A L P P S L 1500
jap a j ( O R F ) CCGCCGGCGC GATCGCCTCG ATCCAGCATT TGAAGGCAAG CCCCTTCGAG CGTGCCCGCC B. onicum. One m o r open reading frame A A G A I A S I Q H L K A S P F E R A R
star t ing f rom nucleot ide 588 and ending at nucleot ide ls6o (Fig 2) ATCAGGACCG GGTGCGCAAG CTGCGCGGCC TTCTCGATGC ACGTGGCATT CCGCATATGG
1802 was identif ied . . The or ien ta t ion of this O R F . ~ D R v ~ K ~ R G ~. ~ D A ~ ~ ~ ~ n
1620 corresponds to the o r ien ta t ion of the t ranscr ip t ion of AeA~eeCAG CCACATCGTG CCTGTCATGG TGGGTGATGC CGCC~GTGC ~TGGATTT
the hemA gene as deduced by the c loning exper iment D N P S H I V P V M V G D A A K C K W I 1680
dig CGGATATCCT GCTCGACAGT CACGGCGTCT ATGTGCAGCC GATCAACTAT CCGACCGTGC with p a r t i a l S a u 3 A estion. The amino acid sequence s D ~ z ~ ~ s H ~ v ~ v e ~ ~ ~ ~ ~ • v of the hemA gene, as deduced f rom the nucleot ide se- 1~40
CGCGCAAGAC CGAGCGCCTG CGCATTACCC CGACGCCGCT GCATAGCGAT GCCGACATCG quence, indicates a protein with a molecular weight of ~ R K T E R L R I T P T P L H S D A D I
1800 44361 Da, which is very close to that found for the c~- AGCATCTCGT CGGCGCGCTG CACCAGCTCT GGTCACATTG TGCACTGGCG CGTGCGGTGG
A L A S o f B . j a p o n i c u m ( 4 4 5 9 9 D a ; R o b e r t s o n M c C l u n g ~ " ~ v G ^ ~ ~ ~ Z W s H e A ~ A ~ A v 1860 et al. 1987). ACGTGAGGCCG~,** CAG~AAGAT AAA.AGAAGAC CGGGCCGCGT "/'~I'ACGCGGCC ~CT'I'T,l~G1920
TGGCGATCAA TTCACGGGCG TGACGACGGG CTTCCTCATC CGCTGCGAAC ACATCTTCCA 1935
TCGTGGCGGC AGATC ~.~3A Table 2. 6-Aminolevulinic acid synthetase (ALAS) activity of var-
ious hemA clones a
Time (rain) cS-ALAS activity (nmoles of 6-ALA formed per mg of protein)
SAS2025 SAS2026 SAS2027 SASX475
0 0 0 0 0 10 12 13 20 0 2O 22 24 40 0 30 32 39 58 0
a J-ALAS activity of cellular extracts in the presence of succinyI- CoA and glycine. See Table i for a description of the strains
Fig. 2. Nucleotide sequence of the hemA gene of Agrobacterium radiobacter and the deduced amino acid sequence of the HemA protein. A potential Shine-Dalgarno sequence is underlined. PI and P2 refer to the two transcriptional start sites mapped by primer extension (see Fig. 3)
Mapping o f the 5'end o f the h e m A m R N A
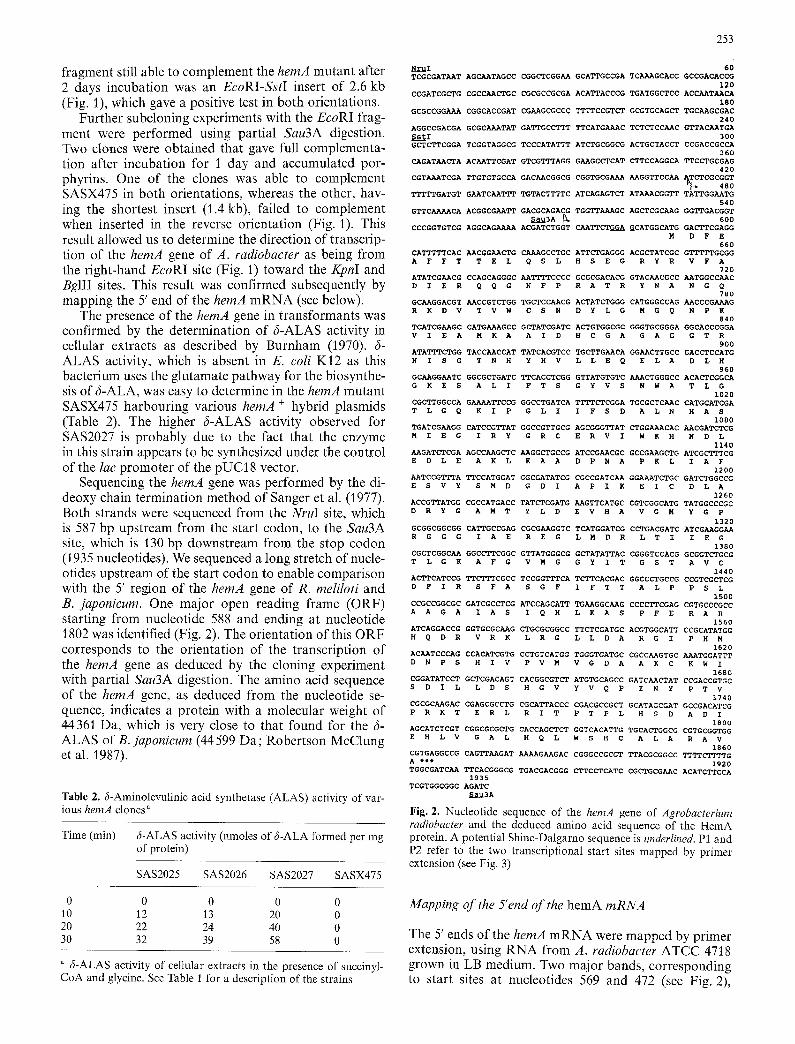
The 5' ends of the hemA m R N A were mapped by pr imer extension, us ing R N A f rom A. radiobacter A T C C 47] 8 grown in LB medium. Two major bands , cor responding to start sites at nucleotides 569 and 472 (see Fig. 2),
472
P E A T G C A a C D E F G
- 6 8
569
254
Fig. 3. Mapping of the 5' end of the hernA mRNA by primer extension. RNA was extracted from A. radiobacter grown in LB medium. The numbers refer to the position in the nucleotide sequence represented in Fig. 2. PE, primer extension
A. radiobacter GGGTTGAC.. GGTCCCGGTGTCGAGGCAGAAAAACGATCTGG I I I I I I I I I I I I I I I I I I I I I I I I I I
R. melill2ti GGGTTGACCAC TGATC GCT T TGAAGGAAGAAAGGCGA . CAGG
Fig. 4. Alignment of the P1 promoter region of the hemA gene of A. radiobacter and Rhizobium meliloti (Leong et al. 1985)
were obtained (Fig. 3). Analysis of the promoter regions deduced from this primer extension experiment, allowed us to identify a T T G A C sequence (starting at position 532; see Fig. 2), located 31 bp upstream from the first start (569) of transcription (P1). This sequence is very similar to the - 3 5 consensus sequence of the E. coli a70 promoters. In the - 1 0 region of the P1 promoter there is a purine-rich sequence (from nucleotides 550- 564; see Fig. 2) which is also found in the same region of the two promoters of the hemA gene of R. meliloti (Leong et al. 1985). Very weak homology is found be- tween these - 1 0 regions and the consensus - 1 0 region of the E. coli 0-70 promoters. An alignment of the PI promoters of the hemA gene of R. meliloti and A. radio- bacter is shown in Fig. 4.
Seven base pairs upstream from the P2 transcription start, a TATAAA sequence is found, which is very simi- lar to the consensus - 10 region of the E. coli ~ 70 pro- moters. However, the - 3 5 region of the P2 promoter
- 4 5 l>
- 2 4
- 18
Kd
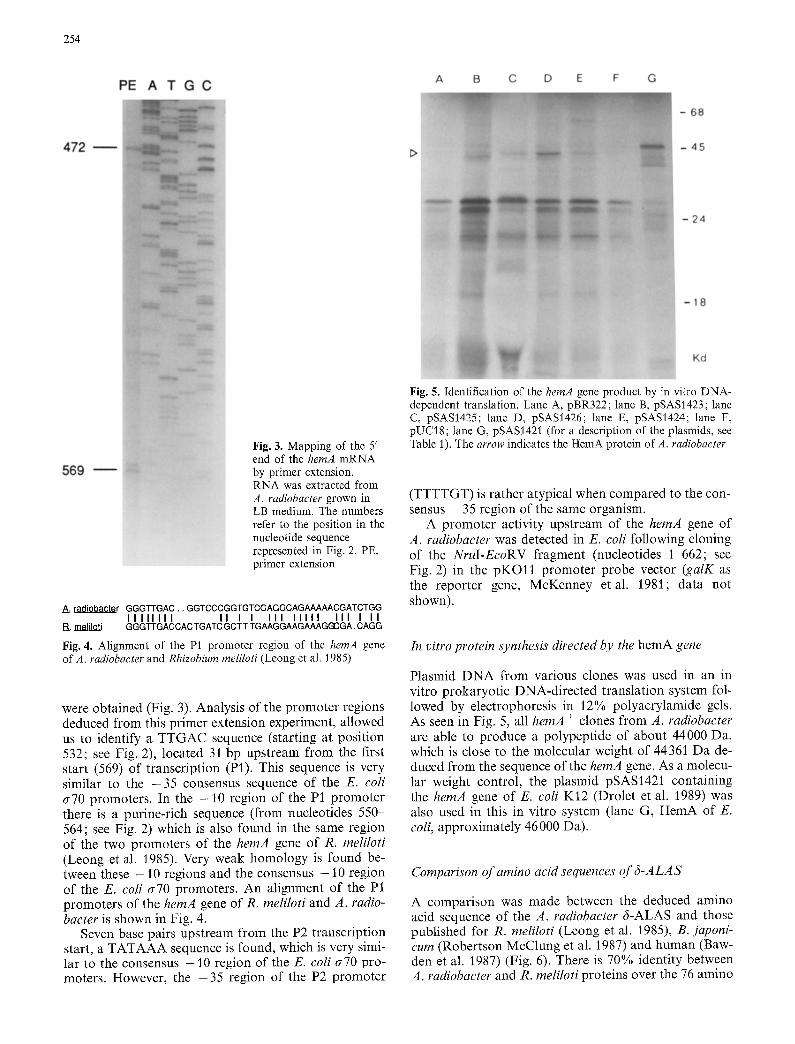
Fig. 5. Identification of the hemA gene product by in vitro DNA- dependent translation. Lane A, pBR322; lane B, pSAS1423; lane C, pSAS1425; lane D, pSAS1426; lane E, pSASI424; lane F, pUC18; lane G, pSAS1421 (for a description of the plasmids, see Table 1). The arrow indicates the HemA protein of A. radiobacter
(TTTTGT) is rather atypical when compared to the con- sensus - 3 5 region of the same organism.
A promoter activity upstream of the hemA gene of A. radiobacter was detected in E. coli following cloning of the NruI-EcoRV fragment (nucleotides 1-662; see Fig. 2) in the p K O l l promoter probe vector (galK as the reporter gene, McKenney etal . 1981; data not shown).
In vitro protein synthesis directed by the hemA gene
Plasmid DNA from various clones was used in an in vitro prokaryotic DNA-directed translation system fol- lowed by electrophoresis in 12% polyacrylamide gels. As seen in Fig. 5, all hemA + clones from A. radiobacter are able to produce a polypeptide of about 44000 Da, which is close to the molecular weight of 44361 Da de- duced from the sequence of the hemA gene. As a molecu- lar weight control, the plasmid pSAS1421 containing the hemA gene of E. coli K12 (Drolet et al. 1989) was also used in this in vitro system (lane G, HemA of E. coli, approximately 46000 Da).
Comparison of amino acid sequences o f 6 - A L A S
A comparison was made between the deduced amino acid sequence of the A. radiobacter 5-ALAS and those published for R. meIiloti (Leong et al. 1985), B. japoni- cum (Robertson McClung et al. 1987) and human (Baw- den et al. 1987) (Fig. 6). There is 70% identity between A. radiobacter and R. meliloti proteins over the 76 amino

255
1 50 A STFHY DRFFEKY.S_DEKN DDHTYRVFRTVNRRAHI FPMADDY S DS LI TK~Q~SVWCS NDY B . . MDYSQFFNSALDRLHTERRYRVFADLERMAGRFpHAIWH • SPKG. KRDVVIWCSNDY C . . MDFEAFFTTELQS~SEGRYRVFADIERQQ~NFPRATRY. NANGQRKDVTVWCSNDY D MMMDFE S FFKNELDGLHQEGRYRVFADLARI{RGS FPKATRY . TADG . AQEVTVWCSNDY
i00 A LGMSRHPRVCGAVMDTLKQHGAGAGGTR~I SGT S KFHVDLERE LADLHGKDAALLFS S C B LGMGQHPKVVGAMVETATRVGTGAGGTRNIAGTHHPLVQ_~_AELADLHGKEAALLFTS G C LGMGQNPKVl EAMKAAI DHCGAGAGGTRNI S GTNHYHVLLEQE LADLHGKE SALI FTS G D LGMGQC_PIVTEAMKNAI DE
150 A FV_AHDSTLFTLVKMMPGCE I YS DS GDHASMI QGIRNSRVPKYI FRHNDVSHLRELLQRS B YVSN_QTGIAT IAKLI PNCLI LS DELNHS SMIEGIRQSGCERQVFRHNDLADLEALLKAA C YVSNWATLGTLGQKI PGLI_I FS DALNHASMIEGIRYGRCERV_IWKHNDLE DLEAKLKAA
200 A DPSVPKIVAFETVHSVDGAVLPLEE LCDVAHEFGAITFVDEVHAWGLYGARGGGIGDRD B GANRPKLIACE S LYS MDGDVAPLAKI CDLAEKYNAMTYVDEVHAVGMYGPRGGGIAERD C DPNAPKLIAFE SVYSMDGDIAPIKE I CDLADRYGAMTYLDEVHAVGMYGPRGGGIAERE
250 A GVMPKMDI I SGTLGKAFGCVGGY IASTRS LMDTVRS YAAGFI FTTS LPPMLI~GALESV B GVMHRI DI LEGTLAKAFGCLGGY IAANGRI I DAVRS YAPGFI FTTALPPAI CSAATAAI C G LMDRLTI I EGTLGKAFGVMGGYITGSTAVCDF IRS FAS GFI FTTALPPS LAAGA IAS I
300 350 A RI LKSAEGRVV~_ Q~RN~_QMLMDAGLPVVHCPSHI I PVRVADAAKNTEVCDE_LM B KHLK.. T_S SWERERHQDRAARVKAI LNAAGLPVMS S DTHIVPLF I GDAEKCKQAS DLLL C QHLK.. AS PFERARHQDRVRKLRGLLDARG I PHMDNPS HIVPVMVGDAAKCKWI S DILL
40O A S _PJ~NI YVQAINY PTVPRGEE LLRIAPTPHHTPQMMNYFLENLLVTWKQVGLE LKPHS S A B EEHG I YI 0PINYPTVAKGS ERLRI TPS PYHDDGLI DQLAEALLQVWDRLGLPLKQKSLA_ C DSHGVYVQPI NY PTVPRKTERLRI TPTPLH_S DADIEHLVGALHOL_WSHCA_LARAVA*
A ECN B AE*
Fig. 6. Comparison of the deduced amino acid sequences of human (A, amino acids 197-612, Bawden et al. 1987), Bradyrhizobiumja- ponicum (B, Robertson McClung et al. 1987), A. radiobacter (C) and R. meliloti (D, the 76 known amino acids, Leong et al. 1985)
acid sequence published for R. meIiloti. For the same region, the B. japonicum and A. radiobacter proteins show 54% identity and those of R. meliloti and B. japoni- cum, 53%. When the entire length of the A. radiobacter protein was compared with the B. japonicum protein, we found 60% identity, while the same comparison be- tween the A. radiobacter and human proteins showed 48% amino acid identity. After including in this compar- ison the highly conserved amino acids as described by Dayhoff et al. (1972), the percentage homology between the HemA proteins of A. radiobacter and B. japonieum increased to 81% and that between the A. radiobacter and human proteins to 71%.
Discussion
A. radiobacter, a close relative of R. meliloti, uses for the synthesis of 6-ALA the C4 (succinate) pathway, in contrast to the C5 (glutamate) pathway used by E. coli. In spite of the presence of different pathways for the synthesis of 6-ALA, the hemA gene of A. radiobacter is able to complement a hemA mutant of E. coil K12. Similar complementation of E. coli hemA mutants has already been described in the case of the hemA gene of R. meliloti (Leong et al. 1982) and of mouse (Schoen- haut and Curtis 1986). The interesting point here is that these cases do not represent a classical genetic comple- mentation, where the complementing cistron is part of the same pathway as the deficient one.
We observed that the complementation of the hemA mutant of E. coli K12 was very slow (2 days) even when
the hemA gene was present on a very high copy number vector (in pUC, more than 200 copies per cell). We also observed that the fragment cloned in pKO11 vector al- lowed E. coli cells to metabolize galactose very slowly. Consequently, we have to conclude that these hemA pro- moters are poorly active in E. coli. Moreover, when the hemA gene of A. radiobacter was cloned in the low copy number vector pRK290 and used to transform a hemA mutant of E. coli K]2, only very weak complementation was observed after prolonged incubation (not shown). The same situation was observed with the hemA gene of R. meliloti cloned in the same vector (Leong et al. 1982) and used to transform the same E. coIi hemA mu- tant. It is of interest to note that the hemA gene of Rhodobacter capsulatus has been shown to be able to complement a hemA mutant of E. coli only when the gene was under the control of the lac promoter of the pGEM vector (Hornberger et al. 1990)
Comparison between the P1 promoters of the hemA genes of R. meliloti and A. radiobacter revealed a high degree of homology. The fact that the unusual " 1 0 re- gions (purine-rich) of these promoters are similar may reflect the presence of an alternative factor essential for efficient transcription of these hemA genes. The absence of such a factor in E. coli may explain why these hemA gene are so poorly expressed in this bacterium.
Homology studies between the first 76 amino acids of the HemA protein of R. meliloti with those of A. radiobacter and B. japonicum, revealed a higher degree of amino acid sequence identity between R. meliloti and A. radiobacter. This suggests that R. meliloti and A. ra- diobacter are more closely related than, at least, R. meli- loti and B. japonicum. Comparison between the entire length of HemA proteins revealed the presence of strong homology between such evolutionary distant organisms as bacteria and human. This suggests major structural constraints for the activity of 6-ALAS, as suggested by Robertson McClung et al. (1987) from comparisons be- tween 6-ALAS of chicken and B. japonicum.
Acknowledgments. We gratefully acknowledge the valuable help of D. Tardif in maintaining our collection of strains. We thank H. Bergeron for the preparation of figures. This study was supported by grant OGP0036561 from the Natural Sciences and Engineering Research Council of Canada and grant CAFIR-196 from Univer- sit6 de Montr6al to A. Sasarman. M. Drolet was the beneficiary of a studentship from the Medical Research Council of Canada.
References
Aiba H, Adhya S, de Crombrugghe B (1981) Evidence for two functional gal promoters in intact Escherichia coli cells. J Biol Chem 256:1190~11910
Arrese M, Carvajal E, Robison S, Sambunaris A, Panek A, Mat- toon J (]983) Cloning of the 5-aminolevulinic acid synthase structural gene in yeast. Curr Genet 7:175-183
Avissar YJ, Beale SI (1989) Identification of the enzymatic basis for 6-aminolevulinic acid auxotrophy in a hemA mutant of Eseheriehia coli. J Bacteriol 171:2919-2924
Avissar YJ, Ormerod JG, Beale SI (1989b) Distribution of ~-ami- nolevulinic acid biosynthetic pathways among phototrophic bacterial groups. Arch Microbiol 151:513-519

256
Bard M, Ingolia TD (1984) Plasmid-mediated complementation of a 5-aminolevulinic acid-requiring Saccharomyces cerevisiae mutant. Gene 28 : 195 199
Bawden M J, Borthwick IA, Healy HM, Morris CP, May BK, E1- liott WH (1987) Sequence of human 5-aminolevulinate synthase cDNA. Nucleic Acids Res 15 : 8563
Biel SW, Wright MS, Biel AJ (1988) Cloning of the Rhodobacter capsulatus hemA gene. J Bacteriol 170:4832-4834
Borthwick IA, Srivastava G, Hobbs AA, Pirola BA, Brooker JD, May BK, Elliott WH (1984) Molecular cloning of hepatic 5- aminolevulinate synthase. Eur J Biochem 144: 95-99
Borthwick IA, Srivastava G, Day RD, Pirola BA, Snoswell MA, May BK, Elliott WH (1985) Complete nucleotide sequence of hepatic 5-aminolaevulinate synthase precursor. Eur J Biochem 150:481-484
Burnham BF (1970) 5-aminolevulinic acid synthase (Rhodopseudo- monas spheroides). Methods Enzymol 17 A: 195-200
Burnham BF, Lascelles J (1963) Control of porphyrin biosynthesis through a negative feedback mechanism. Biochem J 87:462-472
Castelfranco PA, Beale SI (1983) Chlorophyll biosynthesis: recent advances and areas of current interest. Annu Rev Plant Physiol 34:241-278
Dayhoff MO, Eck RV, Park CM (1972) A model of evolutionary change in proteins. In: Dayhoff MO (ed) Atlas of protein se- quence and structure 1972, vol 5. National Biomedical Research Foundation, Washington, DC, pp 89-99
Drolet M, P61oquin L, Echeland Y, Cousineau L, Sasarman A (1989) Isolation and nucleotide sequence of the hemA gene of Escherichia eoli KI2. Mol Gen Genet 216:347 352
Elliott T (1989) Cloning, genetic characterization, and nucleotide sequence of the hemA-prfA operon of Salmonella typhimurium. J Bacteriol 171:3948-3960
Granick S, Beale SI (1978) Heroes, chlorophylls, and related com- pounds: biosynthesis and metabolic regulation. Adv Enzymol 46 : 33-203
Holmes DS, Quigley M (1981) A rapid boiling method for the preparation of bacterial plasmids. Anal Biochem 114:193-197
Hornberger U, Liebetanz R, Hans-Volker T, Drews G (1990) Clon- ing and sequencing of the hemA gene of Rhodobacter eapsulatus and isolation of a 6-aminolevulinic acid dependent mutant strain. Mol Gen Genet 221:371-378
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227 : 680-685
Lascelles J (1975) The regulation of heme and chlorophyll synthesis in bacteria. Ann NY Acad Sci 244:334-347
Leong SA, Ditta GS, Helinski DR (1982) Heine biosynthesis in Rhizobium. Identification of a cloned gene coding for 5-amino- levulinic acid synthetase from Rhizobium meliloti. J Biol Chem 257: 8724-8730
Leong SA, Williams PH, Ditta GS (1985) Analysis of the 5' regula- tory region of the gene for 5-aminolevulinic acid synthetase of Rhizobium meliloti. Nucleic Acids Res 13:5965 5976
Li JM, Brathwaite O, Cosloy SD, Russel CS (1989a) 5-aminolevu- linic acid synthesis in Escherichia coll. J Bacteriol 171:2547 2552
Li JM, Russel CS, Cosloy SD (1989b) Cloning and structure of the hemA gene of Escherichia eoli K-12. Gene 82:209 217
Maguire D J, Day AR, Borthwick IA, Srivastava G, Wigley PL, May BK, Elliott WH (1986) Nucleotide sequence of the chicken 5-aminolevulinate synthase gene. Nucleic Acids Res 14:1379 1391
Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Mayer SM, Beale SI, Weinstein JD (1987) Enzymatic conversion
of glutamate to 6-aminolevulinic acid in soluble extracts of Eug- lena gracilis. J Biol Chem 262:12541 12549
McKenney K, Shimatake H, Cored D, Schmeissner U, Brady C, Rosenberg M (1981) A system to study promoter and termina- tor signals recognized by Eseherichia coli RNA polymerase. In: Chirikjian JG, Pappas TS (eds) Gene amplification and analysis, vol 2, Structural analysis of nucleic acids. Elsevier, New York, pp 383-415
Miller J (1972) Experiments in molecular genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Petricek M, Rutberg L, Shroder I, Hederstedt L (1990) Cloning and characterization of the hemA region of the Bacillus subtilis chromosome. J Bacteriol 172:2250-2258
Pirola BA, Mayer F, Borthwick IA, Srivastava G, May BK, Elliott WH (1984) Structure of the 5-aminolevulinic acid synthase from chick embryo liver. Eur J Biochem 144:577-579
Robertson McClung C, Somerville JE, Guerinot ML, Chelm BK (1987) Structure of the Bradyrhizobium japonieum gene hemA encoding 5-aminolevulinic acid synthase. Gene 54:133-139
Sandy JD, Davies RC, Neuberger A (1975) Control of 5-aminole- vulinate synthase activity in Rhodopseudomonas spheroides. Bio- chem J 150:245-257
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 74: 5463-5467
Sasarman A, Surdeanu M, Horodniceanu T (1968a) Locus deter- mining the synthesis of &aminolevulinic acid in Escheriehia coli K12. J Bacteriol 96 : 1882-1884
Sasarman A, Surdeanu M, Szegli G, Horodniceanu T (1968b) He- rain-deficient mutants of Eseherichia coli K12. J Bacteriol 96:570-572
Sasarman A, Sanderson KE, Surdeanu M, Sonea S (1970) Heroin- deficient mutants of Salmonella typhimiurium. J Bacteriol 102:531-536
Schoenhaut DS, Curtis PJ (1986) Nucleotide sequence of mouse 5-aminolevulinic acid synthase cDNA and expression of its gene in hepatic and erythroid tissues. Gene 48:55-63
Silhavy TJ, Berman ML, Enquist LW (1984) Experiments with gene fusions. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Srivastava G, Borthwick IA, Maguire D J, Elferink CJ, Bawden M J, Mercer JFB, May BK (1988) Regulation of 5-aminolevulin- ate synthase mRNA in different rat tissues. J Biol Chem 263 : 5202-5209
Tai T-N, Moore MD, Kaplan S (1988) Cloning and characteriza- tion of the 5-aminolevulinate synthase gene(s) from Rhodo- bacter sphaeroides. Gene 70 : 139-151
Urban-Grimal D, Ribes V, Labbe-Bois R (1984) Cloning by genetic complementation and restriction mapping of the yeast heml gene coding for 5-aminolevulinate synthase. Curr Genet 8: 327- 331
Urban-Grimal D, Volland C, Garnier T, Dehoux P, Labbe-Bois R (1986) The nucleotide sequence of the heml gene and evi- dence for a precursor form of the mitochondrial 5-aminolevu- linate synthase in Saccharomyces cerevisiae. Eur J Biochem 156:511-519
Verkamp E, Chelm BK (1989) Isolation, nucleotide sequence and preliminary characterization of the Escherichia coli K-12 hemA gene. J Bacteriol 171:4728-4735
Yubisui T, Yoneyama Y (1972) Aminolevulinate synthase of Rho- dopseudomonas spheroides: purification and properties of the enzyme. Arch Biochem Biophys 150 : 77-85
C o m m u n i c a t e d by A. K o n d o r o s i