Embed Size (px)
Citation preview

BioIogical Conservation 36 (1986) 339-350
Climatic Effects on Calling and Spawning of the Natterjack Toad Bufo calamita: Discriminant
Analyses and Applications for Conservation Monitoring
Brian Banks* & Trevor J. C. Beebee
School of Biology, University of Sussex, Falmer, Brighton BNI 9QG, Great Britain
ABSTRACT
Unlike the other anurans native to Britain, the rare natterjack toad Bufo calamita has a protracted breeding season. Surviving populations are regularly monitoredfor assessment ofpopulation trendr and conservation requirements. We have examined the factors influencing callmg and spawning behaviour at two British sites and used discriminant analysis to identtfy climatic conditions associated with reproductive activity. These conditions include rainfall over a short period preceding spawning, though not necessarily on the day itself, and preferably following a spell of dry weather; maximum temperatures on the day before spawning of at least lo”C, and a series of preceding minimum temperatures of at least 5°C (on average) over 2-4 nights. We anticipate that this kind of climatic information, which is accessible without actually visiting toad sites, should maximise the eficiency of monitoring operations by allowing judicious use of limited resources.
INTRODUCTION
The natterjack toad Bufo calamitu is a widespread but locally distributed amphibian in western and north-central Europe (Beebee, 1979). How- ever, the specialised requirements of this species have led to dramatic declines over recent decades in some countries, such as Britain, where changes of land use have severely reduced the extents of suitable heathlands and sand dunes (Beebee, 1976, 1977). Monitoring and conservation of surviving populations have intensified in recent years, * Present address: c/o Nature Conservancy Council, South-East Regional Sub-Office, 73a High Street, Lewes, East Sussex, Great Britain.
339 Biol. Comerv. 0006-3207/86/$03-50 0 Elsevier Applied Science Publishers Ltd, England, 1986. Printed in Great Britain

340 Brian Banks. Trevor J. C. Beebee
the former activity relying heavily on assessing numbers of adults assembling to breed and subsequent survival of spawn and tadpoles. Unfortunately this assessment is more difficult with the natterjack than with species which breed ‘explosively’, i.e. those which spawn over brief and readily predictable timespans (such as Bufo bufo). The breeding season of B. cahmita extends from late March to July, during which period calling and spawning may occur at any climatically suitable time, though there is usually a large spawning effort early in the season (Banks & Beebee, 1986).
This extended breeding season poses problems for those monitoring toad population sizes. Counting numbers of spawn strings is generally regarded as the best approach for the rapid assessment of female numbers, on the assumption that each female deposits a single string every year (Smith & Payne, 1980). Mark and recapture methods, though potentially more accurate, are too time-consuming for wide-ranging surveys of the type commonly needed for conservation purposes. However, since spawn may be laid at any time over a period of up to 90 days or so (Beebee, 1985) it would be helpful to identify conditions within this period when breeding activity is most probable. To do so would effectively reduce the effort needed to carry out surveys, which in turn would make it more feasible to monitor the entire 40 or so colonies of natterjack toads remaining in Britain on a fairly regular basis. Such a data base has obvious implications for effective conservation planning.
We were therefore interested in relating calling and spawning activities of natterjack toads to particular climatic conditions. The hope was that by using simple macroclimatic measurements, which need not be made at the sites themselves, it would be possible to define and predict likely calling and spawning nights. The study involved two rather different toad populations; one was a small colony of less than 100 adults on heathland in Hampshire, and the second a much larger one with several thousand adults on sand dunes in Cumbria. Such a comparison should clarify the generality, or otherwise, of factors implicated as being important.
METHODS
During 1982 and 1983 alternate periods of a few days each were spent at the two study sites. Numbers of spawn strings deposited each night were recorded and calling activity of males was noted at the Hampshire

Climatic efects on Natterjack toad 341
site only. Data from Cumbria in 1982 and 1983 were not suitable for detailed analysis because, being such a large population, much spawn was deposited during periods of observer absence. In 1984 an entire season, excepting three days in April, was spent at the Cumbrian site and spawning data from this year were included in the analysis. Because the Cumbrian natterjack population was so large, calling by at least a few males occurred on most nights during the breeding season. This made studies of optimal calling conditions more difficult than at the Hampshire locality, where qualitative assessment could more readily be used. This is why calling was not monitored in Cumbria.
Meteorological information was obtained from the Meteorological Office, Bracknell, Berks. The relevant recording stations were both within 10 km of the study sites.
Discriminant analyses were carried out as described by Johnston (1978) using a SPSS package on the University of Sussex VAX computing system. Dependent variables were calling days (recorded as plus or minus) and spawning days (recorded similarly). Independent variables were: MIT = Minimum temperature on night before event; MAT = Maximum temperature on day of event; RF = Rainfall on day of event; SH = Sunshine hours on day of event; MT = Minimum temperature on night of event; MTA = accumulated minimum temperatures (sum) on nights - 4 to 0, where 0 = night of event; XTA = accumulated maximum temperatures on days -4 to 0; RFT = accumulated rainfall over days -4 to 0; X = days since last day on which rainfall = twice monthly average; Y = maximum temperature on day of event more (1) or less (0) than 12.5”C; Z = days since rainfall ‘burst’, defined as a day with at least as much rain as that in the accumulated four days previous to it; P = percentage of female population not yet spawned. This was calculated retrospectively for any particular day during the breeding season, after the end of it when the total number of strings deposited was known. Precise locations and fates of each string were noted to avoid duplication of counting.
RESULTS
General breeding patterns
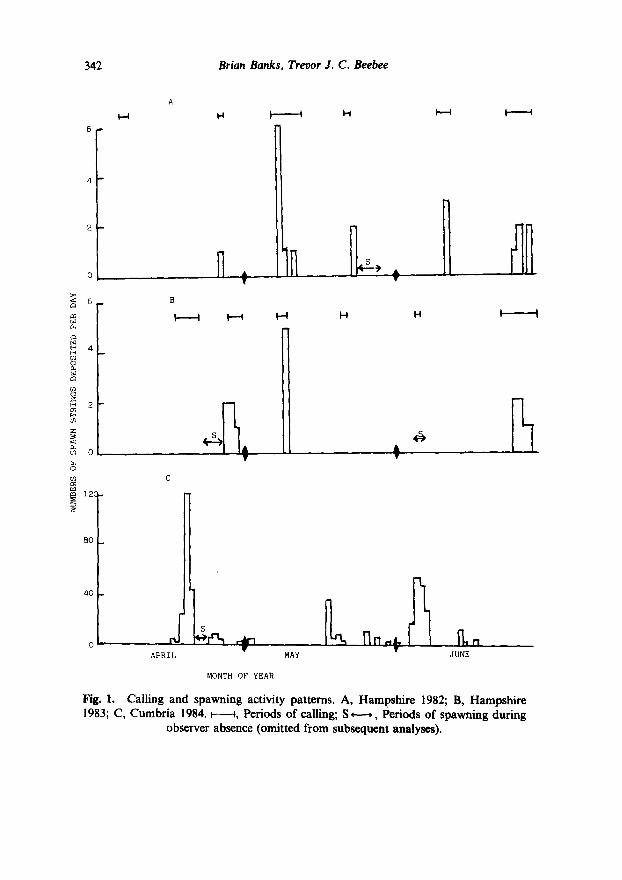
Calling and spawning activities observed in Hampshire in 1982 and 1983, and spawning in Cumbria during 1984, are summarised in Fig. 1.
342 Brian Banks, Treuor .I. C. Beebee
80
40
0 APRIL JUNE
MONTH OF YEAR
Fig. 1. Calling and spawning activity patterns. A, Hampshire 1982; B, Hampshire 1983; C, Cumbria 1984. M, Periods of calling; S - , Periods of spawning during
observer absence (omitted from subsequent analyses).

Climatic effects on Natterjack toad 343
Observations in Hampshire were made on 97 days over the two seasons; spawning (and presumably calling) which occurred during periods of observer absence were necessarily excluded from subsequent analysis. Observations in Cumbria during 1984 covered 73 days of the single season and very little activity was missed.
In general the breeding season was more protracted at the Cumbrian site, with spawning starting earlier and finishing later than in Hampshire in all three years (Cumbrian data for 1982 and 1983 not shown, see Methods). Breeding stopped unusually early at both sites in 1984 due to desiccation of the breeding ponds, and there was no late June spawning in Hampshire that year. Both sites were characterised by episodic bursts of activity; in Hampshire there were 37 calling days and 17 on which spawn was deposited out of the 97 studied. In Cumbria there were 36 spawning days, essentially half the total analysed. No spawn was laid either before or after the overall timespans of these studies, and spawning was almost invariably on calling days/nights at both sites. Calling, on the other hand, sometimes occurred over periods when no spawn was laid. This was particularly true at the start of the breeding period.
Climatic conditions and breeding activity
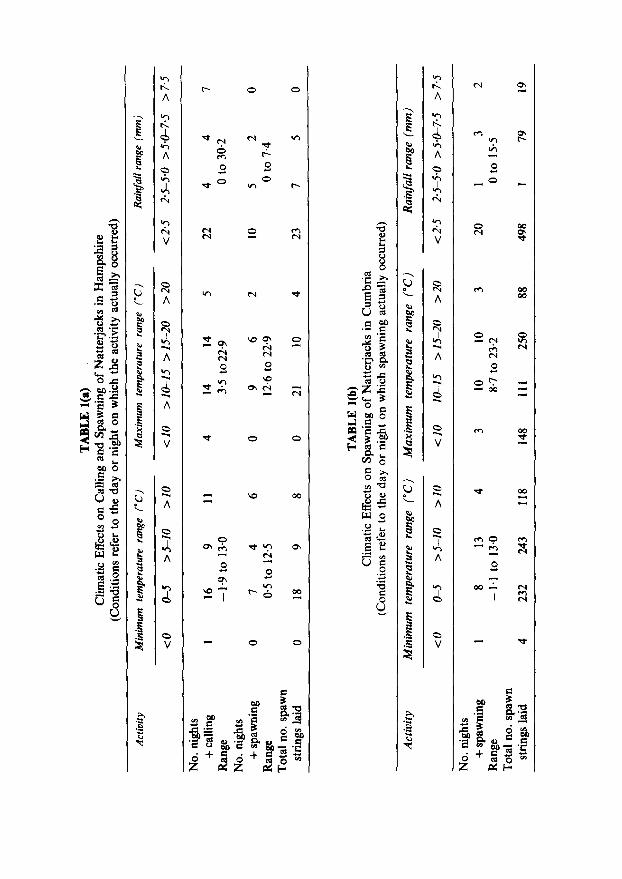
Both calling and spawning occurred over a wide range of environmental conditions (Table 1). Calling was surprisingly frequent on nights when temperatures eventually dropped to below 5”C, and exceptionally both calling and spawning occurred prior to a frost later the same night. At the Cumbrian site it was particularly noticeable that early in the season, when evening temperatures dropped rapidly, some pairs formed but delayed spawning until the following day when they could be seen active in bright sunshine. Later in the season spawning was almost always during the hours of darkness. Spawning was rare on nights following days with maximum temperatures of less than lO-12”C, though there was no obvious relationship with sunshine hours (data not shown). Calling, on the other hand, was more frequent after days with little sunshine, as has been noted before (Blankenhorn, 1972). A striking observation was the tendency to spawn during 24 h periods with little or no rainfall. In Hampshire, for example, 10 out of 17 spawning days were completely dry. Examination of meteorological data nevertheless suggested some link between rainfall and reproductive activity which
TABL
E l(n
)
Activ
ity
Clim
atic
Effe
cts
on C
allin
g an
d Sp
awni
ng o
f N
atte
rjack
s in
Ham
pshi
re
(Con
ditio
ns
refe
r to
the
day
or
nigh
t on
whi
ch t
he a
ctiv
ity a
ctua
lly o
ccur
red)
Min
imum
te
mpe
ratu
re
rang
e (“C
) M
axim
um
tem
pera
ture
ra
nge
(‘C)
Rainf
all
rang
e (m
m)
CO
o-5
>5-1
0 >l
O
<IO
>l
O-1
5 >
15-2
0 >2
0 <2
.5
2.5-
5.0
>5.0
-7.5
>
7.5
No.
nig
hts
+ ca
lling
1
16
9 11
4
14
14
5 22
4
4 I
Ran
ge
-1.9
to
13.
0 3.
5 to
22.9
0
to 3
0.2
No.
nig
hts
+ sp
awni
ng
0 7
4 6
0 9
6 2
10
5 2
0 R
ange
0.
5 to
12.
5 12
.6 to
22.
9 0
to 7
,4
Tota
l no
. sp
awn
strin
gs la
id
0 18
9
8 0
21
10
4 23
7
5 0
TABL
E l(b
) C
limat
ic E
ffect
s on
Spa
wni
ng o
f N
atte
rjack
s in
Cum
hria
(C
ondi
tions
re
fer
to t
he d
ay o
r ni
ght
on w
hich
spa
wni
ng a
ctua
lly o
ccur
red)
Activ
ity
Min
imum
te
mpe
ratu
re r
ange
(“C
) M
axim
um
tem
pera
ture
ran
ge
(‘C)
<O
&5
> 5-
10
>I0
Cl0
lo
-15
> 15
-20
>20
Rai
nfal
l ra
nge
(mm
)
~2.5
2.
5-5.
0 >5
@-7
.5
> 7.
5
No.
nig
hts
+ sp
awni
ng
Ran
ge
Tota
l no
. sp
awn
strin
gs la
id
1 8
13
4 3
10
10
3 20
1
3 2
-1.1
to
13.
0 8.
7 to
23.
2 0
to 1
5.5
4 23
2 24
3 11
8 14
8 11
1 25
0 88
49
8 1
79
19
TABL
E 2
Dis
crim
inan
t An
alys
is o
f Fa
ctor
s In
fluen
cing
C
allin
g an
d Sp
awni
ng
Activ
ity
% d
ays/
nigh
ts
x2
Deg
rees
P
A
B
co
rrect
ly
Of
clas
s$ie
d fre
edom
r5
‘ a
Cal
ling
(Han
ts)
72
234
7 <
o-00
2 O
-61
O-1
8 3
Cal
ling
(Han
ts) +
‘P
75
33.1
8
oGO
Oo1
O
-64
0.14
s
Spaw
ning
(H
ams)
66
IO
.0
7 o-
19
o-33
O
-04
OJ
Spaw
ning
(H
ants
) +
‘P
79
21.2
8
O-0
06
o-45
04
6 2
Spaw
ning
(C
umbr
ia)
77
22-3
7
O-0
02
0.67
0.
17
i?
Spaw
ning
(C
umbr
ia)
+ ‘P
77
22
.2
8 04
05
0.67
o-
17
9 a An
alys
es w
ere
on d
ata
rela
ting
to 9
7 da
ys (H
ams)
and
73
days
(Cum
bria
). ?v
Inde
pend
ent
varia
bles
use
d w
ere:
for
cal
ling,
MAT
, M
T,
SH,
RF,
X,
Y, Z
with
or
with
out
P, a
nd f
or s
paw
ning
, M
TA,
MAT
, M
IT,
RFI
, X,
Y,
Z w
ith o
r w
ithou
t P
(see
Met
hods
). A,
Pro
babi
lity
of c
allin
g or
spa
wni
ng a
ctua
lly h
appe
ning
on
day
pred
icte
d by
ana
lysi
s to
be
posi
tive.
B,
Pro
babi
lity
of c
allin
g or
spa
wni
ng o
ccur
ring
on a
n un
pred
icte
d da
y.

TABL
E 3
Furth
er
Feat
ures
of
Dis
crim
inan
t An
alys
es
Activ
ity a
naly
sed
% o
f day
s %
of
days
%
of f
alse
A
B
c
D
with
fals
e w
ith fa
lse
posi
tives
ne
gativ
e po
sitiv
e af
ter
15 M
ay
s C
allin
g (H
ants
) +
‘P
7.2
17.5
71
k 1
Spaw
ning
(H
ants
) +
‘P
4.1
16.5
62
11
/16
3116
0.
82
0.55
Sp
awni
ng (
Cum
bria
) 11
.0
12.3
77
51
9 O
-85
f Fa
lse
nega
tives
are
day
s w
here
the
act
ivity
occ
urre
d bu
t w
as n
ot p
redi
cted
by
disc
rimin
ant
anal
ysis
. Fal
se p
ositi
ves
are
days
3
whe
re th
e an
alys
is p
redi
cted
an
even
t whi
ch d
id n
ot a
ctua
lly h
appe
n.
9 A,
Fal
se-p
ositi
ve s
paw
ning
day
s w
hich
fel
l on
actu
al c
allin
g da
ys, a
s a
fract
ion
of t
otal
fal
se-p
ositi
ve s
paw
ning
day
s.
B, F
alse
-pos
itive
spa
wni
ng d
ays
whi
ch w
ere
one
or t
wo
days
afte
r ac
tual
spa
wni
ng d
ays,
as
a fra
ctio
n of
tot
al
fals
e-po
sitiv
e t $
spaw
ning
day
s.
cp
C,
Prob
abilit
y of
fin
ding
nat
teja
cks
eith
er c
allin
g or
spa
wni
ng o
n a
day/
nigh
t pr
edic
ted
posi
tive
for
spaw
ning
. D
, Pro
babi
lity
of fi
ndin
g sp
awn
with
in t
he tw
o da
ys a
fter
depo
sitio
n w
hen
visi
ting
on d
ays/
nigh
ts p
redi
cted
pos
itive
for
spaw
ning
.

Climatic efects on Natterjack toad 347
was not clear by considering days in isolation from the time preceding them, thus necessitating a more comprehensive form of data analysis.
. Dmriminant analysis of factors inihmtcing breeding activity
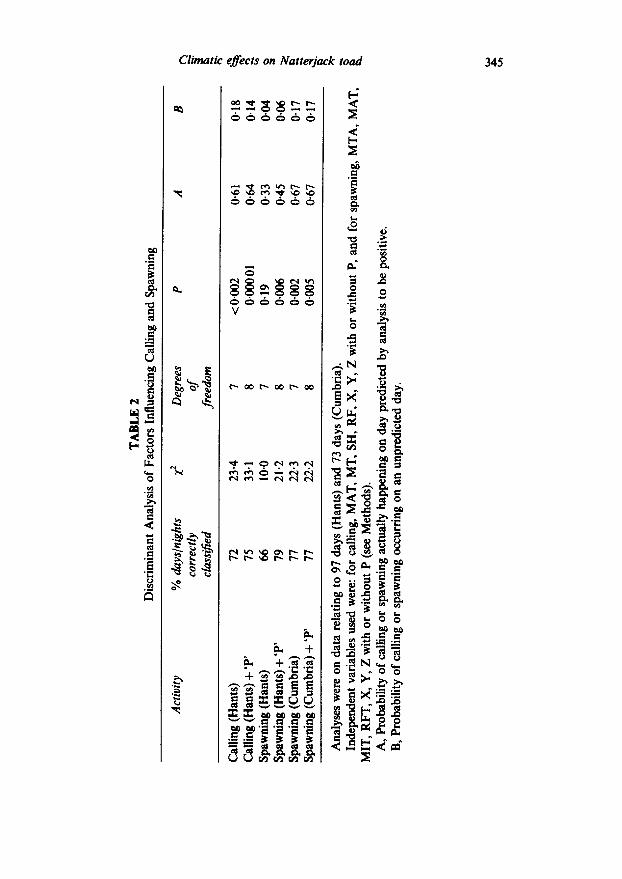
A variety of combinations of meteorological variables were tested, in a series of discriminant analyses, in relation to calling and spawning days. The most successful of these are summarised in Table 2. Calling days were classified with 72-75% accuracy, giving predictive powers of 0*61- 064. Since 37 out of 97 days were used for calling, these values compare with a random probability of O-38. Discriminant analysis only succeeded in making a significant separation of spawning and non-spawning days in Hampshire if the number of available females through the season (P) was included in the variables list. Under these circumstances nearly 80% of days were correctly classified with a predictive power for determining spawning days of 0.45, compared with the random probability of selecting a spawning day of O-17. In Cumbria the use of ‘P’ had no effect on the result, spawning and non-spawning days being classified with 77% accuracy. Predictive power for spawning days was 0.67, compared with the random figure of just under 0.5.
A further assessment of the discriminant analyses is summarised in Table 3. False positives were generally more often predicted than false negatives and most of the false positives fell in the latter part of the breeding seasons. A high proportion of spawning false positives in Hampshire coincided with days or nights when males were calling, all of which may indicate that P was not an accurate indicator of female availability at all times. The chances of finding natterjacks either calling or spawning on a night predicted positive for spawning were greater than 0.8. Moreover, quite a number of false positives for spawning fell within the two days after actual spawn deposition, which is also close enough for many monitoring purposes and could help improve the thoroughness of spawn string recording.
DISCUSSION
Circumstances triggering calling and spawning activities in anurans are certainly complex and probably hierarchical, and it may be unrealistic to expect definitive statements from meteorological data alone. Annual rhythms evolved over long periods of time no doubt define the broad

348 Brian Banks, Trevor J. C. Beebee
limits of the breeding season, for example. Nevertheless, climatic conditions seem likely to be involved in the fine control of breeding activity and this has been borne out by numerous studies with many different species. In the case of ‘explosive’ breeders, such as Bufo bufo, it seems that a threshold minimum temperature of 4-6”C must be exceeded for migration to occur, although the association of movement with rainfall is less clear (Gittins, 1983; Reading & Clark, 1983). It appears that there is some requirement for precipitation, but this can be a day or so previous to activity. Most work on this topic with natterjacks has thus far been casual or anecdotal, emphasising a general view that spawning occurs on cloudy or rainy nights (Kowalewski,’ 1974; Smith & Payne, 1980). A more rigorous study of factors affecting calling by Bufo calamita was carried out by Blankenhom (1972), who concluded that optimal conditions included rainfall up to day - 1, warm water (more than 7°C) at 1900 h and little sunshine on the day itself. No attempt was made to classify calling and non-calling conditions, and spawning was not investigated.
We have used a multivariate approach to address this problem, and successfully classified days/nights into calling or non-calling, spawning or non-spawning, in a statistically significant way by using combinations of meteorological variables. It was found essential to take account not just of conditions on particular days, but also of what was happening over periods previous to particular days. It was notable that the same mixtures of variables gave the best analyses at both study sites, though the discriminant functions derived from them were not identical. This is scarcely surprising since weather patterns at the two sites (one coastal, the other inland) differ considerably in most years. Many of the climatic variables no doubt act indirectly, e.g. via effects on water temperatures in the breeding ponds. The point of this analysis, of course, was to be able to predict toad activity using information widely available in weather forecasts and meteorological offices without having to visit the site first (which would negate the whole purpose of the study), so direct measurements of water temperatures could not be included.
An important difference between the two sites was toad population size. In Cumbria the numbers of females vastly outnumbered the total number of days available for spawning (by 6-8: 1) whereas in Hampshire the opposite was true, with a ratio of < 1:3. It was therefore much more likely in Hampshire than in Cumbria that suitable spawning conditions would arise without sexually receptive females being available to use them, a fact which we think explains the improvement seen in the

Climatic efects on Natterjack toad 349
Hampshire analysis when P was included. It may also happen that spawn production is sometimes constrained by the numbers of suitable spawning days/nights, though this would be difficult to demonstrate unequivocally. Practical use of this analysis therefore requires previous knowledge on the approximate size of the female population and, in the case of small colonies (say less than 100 females) a realisation that as the season progresses the chance of finding spawn even on a good day is likely to depend less on prevailing weather conditions than on the fraction of the population present that has not yet spawned. The former is now available for many natterjack colonies in Britain and the latter would thus be calculable approximately during any serious monitoring operation of a small colony, so these are not as restrictive features as they seem at first sight. Of course, monitoring aims to look for changes in population size, but so long as these are not dramatic in small colonies from one year to the next, use of discriminant analysis results and population estimates from the previous season should still facilitate detection of most toad activity through a breeding period.
Examination of the discriminant analyses allows circumscription of factors which combine to produce a high probability of toad activity, and hence optimal monitoring conditions. These are, in descending order of importance: high total rainfall (25-40mm) over the previous 2-4 days, preferably following a period of low precipitation; toad activity is maximal 2-4 days after the initial rainfall peak, often on days which themselves have little or no rain. Maximum temperature on the day before the spawning night should be at least lO”C, preferably up towards 18°C. A series of previous nights with above-average (for the time of year) minimum temperatures, preferably greater than 5°C over 3-4 nights, is also conducive. Such guidelines should give a high probability of encouraging calling or spawning activity, and should identify most breeding days/nights over a season, as long as female numbers and availability are also taken into account for small popu- lations. Accuracy will be greatest early in the season, which normally coincides with the majority of spawnings and thus the most critical time for monitoring.
ACKNOWLEDGEMENTS
We thank the Nature Conservancy Council and Natural Environment Research Council for financial support, and the Ministry of Defence

350 Brian Banks, Trevor J. C. Beebee
and Cumbria County Council for permissions to work on the study sites.
REFERENCES
Banks, B. & Beebee, T. J. C. (1986). Comparative fecundities of Bufo bufo and Bufo calamita in Britain. J. Zool. (Lond.), 208, 325-38.
Beebee, T. J. C. (1976). The natterjack toad Bufo calamita in the British Isles; a study of past and present status. Brit. J. Herpetol., 5, 515-21.
Beebee, T. J. C. (1977). Environmental change as a cause of natterjack toad Bufo calamita declines in Britain. Biol. Conserv., 11, 87-102.
Beebee, T. J. C. (1979). A review of scientific information pertaining to the nattejack toad Bufo calamita throughout its geographical range. Biol. Conserv., 16, 107-34.
Beebee, T. J. C. (1985). Geographical variations in breeding activity patterns of the natterjack toad Bufo calamita in Britain. J. Zool. (Lond.), 205, l-8.
Blankenhom, H. J. (1972). Meteorological variables affecting onset and duration of calling in Hyla arborea and Bufo calamita. Oecologia (Berl.), 9,223-34.
Gittins, S. P. (1983). The breeding migration of the common toad Bufo bufo to a pond in mid-Wales. J. Zool. (Lond.), 199, 555-62.
Johnston, R. J. (1978). Multivariate statistical analysis in geography. London, Longman.
Kowalewski, L. (1974). Observations on the phenology and ecology of amphibia in the region of Czestochowa. Acta Zool. Cracov., 19, 391-458.
Reading, C. J. & Clark, R. T. (1983). Male breeding behaviour and mate acquisition in the common toad Bufo bufo. J. Zool. (Lond.), 201, 237-46.
Smith, P. H. & Payne, K. R. (1980). A survey of natterjack toad Bufo calamita distribution and breeding success in the north Merseyside sand dune system, England. Biol. Conserv., 19, 27-39.





























