Embed Size (px)
Citation preview

PERSPECTIVE
Climate change frames debate over theextinction of megafauna in Sahul(Pleistocene Australia-New Guinea)Stephen Wroea,b, Judith H. Fielda,1, Michael Archera, Donald K. Graysonc, Gilbert J. Priced, Julien Louysd,J. Tyler Faithe, Gregory E. Webbd, Iain Davidsonf, and Scott D. MooneyaaSchool of Biological, Earth, and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia; bSchool ofEngineering, University of Newcastle, NSW 2308, Australia; cDepartment of Anthropology and Quaternary Research Center, University ofWashington, Seattle, WA 98195; dSchool of Earth Sciences, The University of Queensland, St. Lucia, QLD 4072, Australia; eSchool of SocialScience, The University of Queensland, St. Lucia, QLD 4072, Australia; and fSchool of Humanities, University of New England, Armidale, NSW2351, Australia
Edited by James O’Connell, University of Utah, Salt Lake City, UT, and approved April 9, 2013 (received for review February 12, 2013)
Around 88 large vertebrate taxa disappeared from Sahul sometime during the Pleistocene, with the majority of losses (54 taxa) clearly takingplace within the last 400,000 years. The largest was the 2.8-ton browsing Diprotodon optatum, whereas the ∼100- to 130-kg marsupial lion,Thylacoleo carnifex, the world’s most specialized mammalian carnivore, and Varanus priscus, the largest lizard known, were formidablepredators. Explanations for these extinctions have centered on climatic change or human activities. Here, we review the evidence and argu-ments for both. Human involvement in the disappearance of some species remains possible but unproven. Mounting evidence points to theloss of most species before the peopling of Sahul (circa 50–45 ka) and a significant role for climate change in the disappearance of thecontinent’s megafauna.
megafauna extinction | Pleistocene extinctions | archaeology | human colonization | faunal turnover
Explaining Pleistocene faunal extinctionsremains one of the most challenging prob-lems in the prehistory of Sahul (1–3). Thevast majority of extinctions across geologicaltime are wholly attributable to climate-related factors (4), but claims that some, oreven all, Pleistocene extinctions of large-gigantic vertebrates (Fig. 1) in Sahul were theconsequence of human activity have gen-erated particularly robust debate. Polarizedviews have emerged to account for the modeand timing of these events (2, 5–11). A pau-city of empirical data; shortfalls in radiomet-ric dating; and, until recently, a limitedappreciation of the paleoenvironmental re-cord (7, 12) have placed considerable con-straints on the ability to resolve “who orwhat” was responsible for these extinctions.Given these limitations, assertions such as “...the question is no longer if, but rather how,humans induced this prehistoric extinctionevent” (ref. 13, p. 563) are premature.Sahul—mainland Australia, Tasmania,
and New Guinea—comprised up to ∼11 mil-lion km2 at glacial maxima. Although domi-nated by an expansive desert core (14),environments ranged from periglacial in Tas-mania to tropical in New Guinea (Fig. 2). Itwas against a backdrop of deteriorating con-ditions leading up to the Last Glacial Maxi-mum [LGM; in Marine Isotope Stage (MIS)
2], ∼28–19 ka, that the first people crossedthe biogeographic divide (Wallacea) to enterSahul at ∼50–45 ka (15, 16). It seems likelythat the size, distribution, and density of hu-man populations in pre-LGM times havebeen overestimated (17), although thesewere evidently behaviorally modern people(18, 19).
Late Pleistocene Fauna and ExtinctionChronologiesAs commonly used in the context of extinc-tions in Sahul, the term megafauna refers toan arbitrary compilation of relatively largemammalian, reptilian, and avian taxa, rang-ing in size from∼10 kg or less up to>2,000 kg(20–23). In addition to Diprotodon opta-tum and Thylacoleo carnifex (Fig. 1), otherwell-known marsupial “giants” included the230-kg kangaroo Procoptodon goliah (24),the tapir-like Palorchestes azael, and thebull-sized Zygomaturus trilobus. Amongnonmammalian megafaunal species werethe massively built flightless bird Genyornisnewtoni (25), the anaconda-like madtsoiidsnake Wonambi naracoortensis, and the5-m-long mekosuchine crocodile Pallim-narchus pollens (26).
It is important to note that extinctions inPleistocene Sahul were not restricted to thelarge-bodied species described above. Extinc-tions of small-bodied species, including frogs,
bandicoots, dasyurids, and rodents (27), alsooccurred. Furthermore, a range of medium-and small-bodied species disappeared orunderwent major geographic range shifts ontime frames similar to those of larger taxa(28–33). Also notable is the fact that not allmegafauna went extinct: Some underwentdwarfing, whereas others appear to havesurvived relatively unchanged, such as emus,cassowaries, wombats, salt-water crocodiles,and many species of large kangaroos, in-cluding reds (Macropus rufus), grays (Mac-ropus giganteus and Macropus fuliginosus),and wallaroos (Macropus robustus,Macropusantilopinus, and Macropus bernardus). Taxafrom the whole gamut of body size dis-tributions, not just the largest, were affectedby extrinsic factors during the Pleistocene.Only some of these factors led to extinctions.There is no reason to assume that all specieslumped together as extinct megafauna were
Author contributions: S.W. and J.H.F. designed research; S.W., J.H.F.,
G.J.P., and J.L. performed research; G.J.P., J.L., J.T.F., and S.D.M.
analyzed data; and S.W., J.H.F., M.A., D.K.G., J.T.F., G.E.W., I.D.,
and S.D.M. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/
lookup/suppl/doi:10.1073/pnas.1302698110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1302698110 PNAS | May 28, 2013 | vol. 110 | no. 22 | 8777–8781
PERS
PECT
IVE
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 20
, 202
0

sufficiently similar ecologically or entangled(or contemporaneous, see below) to enableone simultaneous event to account for allthese extinctions.
The last appearance dates in Sahul for thesuite of taxa traditionally referred to as mega-fauna are consistent with a staggered extinc-tion process that was in train well beforethe arrival of humans (3, 10, 12) (Fig. 3). In-cluding new middle Pleistocene species re-corded from south-central Australia (2), asmany as 50 of the 88 known extinct mega-faunal taxa are absent from fossil recordspostdating the Penultimate Glacial Maxi-mum (PGM;MIS6) (3) at∼130 ka (Fig. 3 andTable S1). Additional taxa disappeared at∼85–80 ka (3, 6, 12). There is firm evidencefor only ∼8–14 now-extinct megafaunal spe-cies overlapping with human presence on thecontinent. Nearly half of these late-survivingspecies are from New Guinea (3, 34), andmost are identified from single occurrences.
The uncertainty that currently existsaround establishing accurate chronologies formegafaunal decline and extinction, particu-larly the paucity of securely dated fossil ma-terial (5), has been one of the major obstaclesto resolving the question of what caused theseextinctions. The absence of so many of thespecies in question following the MIS6 glacialmaximum, a time of undoubtedly severeclimate, with further attenuation throughMIS5–MIS3 is consistent with a staggeredextinction chronology. However, it has beensuggested that this appearance of staggeredloss could be a sampling artifact, referred to asthe Signor–Lipps effect (13). Technically, thisis conceivable, because the last occurrences inthe fossil record are unlikely to documentprecisely the actual time of extinction. How-ever, given the actual record and the fact thatso many of the extinct taxa are not knownto have existed within tens of thousands ofyears of human arrival, themost parsimonious
explanation of the data are that the extinc-tions were indeed staggered over a periodthat began long before human arrival. Astaggered extinction event is further sup-ported by recent rigorous statistical tests ofstratigraphically intact prehuman fossil se-quences (Fig. 4) that show progressivelosses of diversity over time (12), stronglysuggesting that temporally progressive mid-dle to late Pleistocene declines in diversityare not sampling artifacts.
Extinctions of a comparable magnitude tothose of megafauna have been observed forsmall-bodied species in a range of specificlocalized studies. For example, small-sizedspecies (<3 kg) from Mount Etna in north-eastern Australia (Fig. 2) experienced a 50%reduction in species richness (from 28 to 35species to 16 to 12 species) over the courseof the middle to late Pleistocene (35). Theselosses track progressive environmental changefrom rainforest to less mesic habitats. Like-wise, the eastern Darling Downs recordsmiddle to late Pleistocene extinctions amongboth small- and large-bodied species con-comitant with progressively drying envi-ronments (30–33).
Human-Mediated Extinction ProcessesThe argument for a primary human role inthe extinction of megafauna has been basedon the presumption that the arrival ofhumans on Sahul was synchronous with thedisappearance of all now-extinct megafauna(36). A link between the two seemed obviousto advocates of a human-driven process (37),and a role for climate change was subse-quently discounted. The case for a humanrole was buttressed by claims that MIS3 (60–28 ka) was a time of relatively stable climaticconditions (13). As such, human activitieswere therefore the only credible explanationfor the extinctions (2, 11, 38).
Proponents of early human-mediatedextinctions in Sahul have based their argu-ments and conclusions on a number of as-sumptions. Key among these are that all ormost now-extinct megafauna survived thePGM and the ensuing 80,000 y to be presentwhen people arrived, that the extinctionscould largely or wholly be attributed to a sin-gle cause within a relatively narrow timeframe, and that climate changes within thelast two glacial cycles were unremarkablerelative to those of previous cycles (3, 9).
The case for a human role was bolsteredfollowing the observation of a broad overlapbetween human arrival and terminal dates fora small number of late-surviving megafaunalspecies. A hypothetical “extinction window”at 51–39 ka was proposed during which, itwas argued, all or most species of now-extinct
megafauna disappeared (6). Modeling studiesbased on the same dataset suggested thathuman activities could have accounted forthe decline and disappearance of all mega-fauna within 600 y (39). Various publicationsbefore and since have offered proxy data tosupport the notion of a primary human role(8, 40).
Human activities as a primary extinctiondriver are not, however, supported by thepaleontology or the archeology. The com-plete lack of evidence for predation on, oreven consumption of, megafauna by peopleaside, the extinction window noted above isbased on a statistical analysis of just sevensites (7). Four of these have no published dataand cannot be scrutinized. A fifth site,Menindee Lake (Fig. 2), is 10,000 y youngerthan originally proposed but still 10,000 yolder than the archeological materials fromthe same site (7). The remaining two locali-ties are >100 ka in age (7), and thus earlierthan the arrival of people by at least 50,000 y.Only two sites in Sahul have secure exca-vated contexts with co-occurrence of extinctmegafauna and people: Cuddie Springs insoutheastern Australia and Nombe Rock-shelter in the New Guinea highlands (7, 34)(Fig. 2). Two other sites have yielded evi-dence for single instances of megafaunapersisting after the LGM. These fauna wereidentified in archeological sites but were re-covered from noncultural (prehuman) hori-zons (Cloggs Cave and Seton Rockshelter)(7). No kill-sites are known (3, 7). Althoughmegafauna are known from Tasmania, mostextinct species had vanished before humansarrived. There is a single example of a short
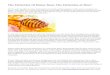
Fig. 1. Thylacoleo carnifex, the 100- to 130-kg mar-supial lion with massive “bolt cutter-like” cheek teeth andthe most powerful bite for its size of any mammaliancarnivore, was a formidable predator of large animals.(Drawing by Peter Schouten.)
Fig. 2. Sites mentioned in the main text. The continentallimits of Sahul are defined by the −130-m bathymetry line,and Sahul incorporates Australia, New Guinea, and Tas-mania. The Nullarbor Plain caves contain middle Pleisto-cene fossil deposits. Kings Creek is located in the easternDarling Downs region.
8778 | www.pnas.org/cgi/doi/10.1073/pnas.1302698110 Wroe et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 20
, 202
0

temporal overlap between people and Pro-temnodon anak, but the bones of P. anakhave never been identified in the rich fau-nal assemblages recovered from the nu-merous Pleistocene archeological sites inves-tigated (41).
Climate Change and MegafaunaMiddle to late Pleistocene faunal extinctionsand human arrival in Sahul occurred againsta backdrop of significant climatic change.Until recently, our capacity to unravel thepotential impacts of climatic flux on plantsand animals, including humans, has beenlimited by the relatively coarse resolution ofthe fossil records. Numerous independentstudies, including analyses based on high-resolution Antarctic ice core data, now allowus to reevaluate the climatic record, the long-term trends spanning some 800 ka, and themore detailed shorter term fluctuations atcritical periods through time.
There is a growing consensus that Sahulwas subject to stepwise and progressive dry-ing beginning as early as 700 ka (42), andnotably within the last 350–400 ka (43–46).In recent years, Antarctic ice core records(47–49) have revealed a distinct change inglacial-interglacial cycles after ∼450 ka. Thelast five interglacial stages (MIS11.3, MIS9.3,MIS7.5, MIS5.5, and MIS1) were, on average,warmer than preceding interglacial stages, asindicated by Deuterium (δD), a proxy fortemperature, in the European Project forIce Coring in Antarctica (EPICA) Dome C(EDC) record. Concurrently, from ∼450 ka,the isotopic minima, representing the coldestor full glacial stages in the EDC δD record,show an obvious downward trend. Thesetrends mean that isotopic changes recordedfor the last glacial cycle (from MIS5.5–MIS2)are almost double the amplitude of the in-terglacial-glacial maximum cycles between800 and 500 ka (Fig. 5). In addition, δD(hence, temperature) is conspicuously low inthe EDC ice core data from ∼70–63 ka andduring MIS2. Importantly, the occurrence oftwo extended and severe cold periods in theone glacial cycle is unprecedented in the800,000 y Antarctic ice core record (48).
The high-resolution EDC record alsodocuments considerable millennial-scalevariability. There are four distinct AntarcticIsotopic Maxima between 60 ka and 35 ka,and it is now clear that there was markedaridification from ∼50–45 ka (50, 51), theinterval during which the first humans ar-rived in Sahul.
In short, the temperature swings of the lastfew glacial cycles were without precedent andthe last glacial cycle exhibits unusual vari-ability. The climate patterns in the 100,000 y
Fig. 3. Extinction chronology of Pleistocene megafaunal taxa in Sahul (88 species). For many species, terminaldates are debated, as is the definition of a “secure” date. If the selection criteria of Roberts et al. (9) are applied, asfew as 8 species clearly survived to the time of human colonization of Sahul. Species reported to overlap withhumans but lacking contextual information linking the dates to the taxa in question are illustrated in gray. Themajority of taxa cannot be placed within the time frame of human arrival, and many lack reliable radiometric datesaltogether, with some of the taxa claimed to have been the victims of human-mediated extinction known only fromthe Pliocene (e.g., Euryzygoma dunense). Of those taxa known to overlap with people, some persist well beyonda previously proposed 51- to 39-ka “extinction window” (9), whereas most of those falling within this window arenot securely linked to the dates (six of nine taxa). Of those taxa potentially surviving until more recently than ∼50 ka,almost half are from New Guinea.
Wroe et al. PNAS | May 28, 2013 | vol. 110 | no. 22 | 8779
PERS
PECT
IVE
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 20
, 202
0

preceding the LGM were anything but be-nign. Rather than MIS3 being a mild andperhaps humid interstadial in Sahul (e.g., ref.13), paleoenvironmental records demonstratesignificant climatic inconstancy. For example,it has recently been shown that climatic var-iability was a key factor in the extinction ofthe large flightless bird G. newtoni near LakeEyre by ∼50 ka (51), and the small mammalsfrom Tight Entrance Cave show a markedincrease in turnover at this time (10).With anincreasing number of reports presentinghigh-resolution data through critical periods,further correlations between the disappear-ance of other megafauna and climate changeseem likely to emerge.
Further support for a climatically drivenprocess comes from analyses demonstratingthat small-bodied species were susceptible tosignificant impacts resulting from climatechange during the middle to late Pleistocene,before human arrival. Bottom-up, ecosystem-changing processes triggered by the disap-pearance of particular small-bodied speciesare increasingly being described for NorthAmerican and Eurasian ecosystems (52, 53).Given trophic interdependencies, ecosystemchanges of this kind in small keystone speciescould have reduced the resilience of larger-bodied species in those same ecosystems.
Paleoenvironmental Proxies and theRole of HumansPaleoenvironmental records have also beenused as important proxies for human ac-
tivity in arguments for a human role in theextinctions. For example, increases in fireactivity reconstructed from charcoal analy-ses of sediment samples recovered fromLynch’s Crater (northeast Queensland) havebeen assumed to be a proxy for humanactivities in particular (54). However, otherrecords show increased burning long before
people arrived (10). A recent comprehen-sive overview of charcoal work in Austral-asia (55) determined that fire histories morestrongly correlate with climate than humanactivities.
Other proxies include dung-fungus (Spor-ormiella). In a recent study, also located atLynch’s Crater, declining Sporormiella spores(a proxy for megaherbivores) at ∼41 ka wereinterpreted as evidence for human-mediatedextinction of megafauna (40). Subsequentpeaks in Sporormiellawere attributed to extantkangaroos. The authors did not explore otherequally plausible scenarios. Variation inSporormiella peaks could as easily be attrib-uted to changes in spore abundance correlatedwith changes in climate, especially moistureavailability. Furthermore, significant con-tributions to all the Sporormiella peaks, notjust those after 41 ka, could have been madeby still-extant kangaroos or cassowaries.
Paleontological and isotopic data frommiddle Pleistocene fossil deposits in southerncentral Australia (2) have demonstrated aridadaptations in some now-extinct fauna. Theimplication is that subsequent increased ar-idity was unlikely to have led to late Pleis-tocene extinctions (2). A human role wasinferred based on the unproven assumptionthat the extinctions occurred after peoplearrived in the region at ∼40 ka. However, asalready noted, there is no evidence that manyof these species survived the middle Pleisto-cene. Paleoenvironmental data show that themiddle to late Pleistocene was particularly
Fig. 4. Late Pleistocene temporal occurrences of megafauna from the intensively sampled Kings Creek Catchment,Darling Downs, southeastern Queensland (13), in relation to the hypothesized continental megafaunal extinctionwindow and earliest evidence of human colonization. Rarefaction, bootstrapping, and associated statistical analysesdemonstrate that the apparent temporally progressive loss of megafauna was a real phenomenon and cannot simplybe explained as a result of taphonomic or sampling biases (14). Importantly, the data clearly point to prehumanclimate changes as a driver of the losses and do not support the hypothesis that there was a local mass extinction of allmegafauna at or around the hypothesized extinction window.
Fig. 5. The EDC δD (‰) record over the last 900 ka (45–47) illustrates the trend toward increasingly warm inter-glacials (red line) and increasingly cold glacial maxima (blue line) beginning at around 400 ka. (Upper) Graphic depictsthe number of species that are not represented thereafter in the fossil record in the following temporal bins: >400 kaor undated, 400–126 ka, 126–51 ka, 51–39 ka, and <39 ka. Species with last appearance dates before human arrival(50–45 ka) are highlighted in green, and those with last appearance dates after this are highlighted in blue. There islittle evidence for any extinction pulse associated with human arrival.
8780 | www.pnas.org/cgi/doi/10.1073/pnas.1302698110 Wroe et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 20
, 202
0

arid and variable and, as noted above, thatclimate deteriorated markedly from ∼50–45ka (42–45, 48, 50, 51). It is clear that at leastsome arid-adapted species can be insulatedagainst aridity. They are advantaged in timesof water stress by increased mobility andwater conservation strategies. However, thereis a tradeoff. Large species must exist at lowerdensities and, of necessity, require greaterhome ranges to maintain viable populations.Further, adaptation to aridity does not in-sulate against hyperaridity (5). Even thesurviving red kangaroo (M. rufus), superblyadapted as it is to arid conditions, has sufferedmassive population crashes during rela-tively benign climate intervals within his-toric times (23).
ConclusionMany questions remain. As we work towarda better understanding of the biology anddistribution of extinct species, a complexpattern of faunal extinctions across time andSahul’s diverse landscape is emerging.Tracking individual histories of faunal species(56), although possible in many northernhemisphere contexts, is severely constrainedin Sahul by the patchy fossil record (5), yet itis one area that would certainly help to re-solve the nature of faunal turnover throughtime. Advances in our knowledge of pale-oclimates are steadily improving our un-derstanding of the complexities of theextinction processes. Mounting evidence in-creasingly points to climate change as the
primary driver of Pleistocene faunal extinc-tions. Many species of megafauna did notpersist into the late Pleistocene, and othersubsequent extinctions postdated the PGMbut were completed before humans arrived.Importantly, a role for humans in the dis-appearance of any surviving taxa, althoughpossible, is yet to be demonstrated. Extinctspecies that persisted until or beyond ∼50–45ka finally disappeared within the context ofa rapidly deteriorating climate.
ACKNOWLEDGMENTS. We thank Jim Allen for incisiveand constructive comments on an early draft of the manu-script. This work was funded by the University of NewSouth Wales and the Australian Research Council: GrantsDP0666374 and DP0987985 (to S.W.), Grant DP0557923(to J.T.F.), Grants DE120101533 and DP120101752 (toG.J.P.), and Grants LP100200486 and LP0989969 (to M.A.).
1 Wroe S, Field J, Grayson DK (2006) Megafaunal extinction:Climate, humans and assumptions. Trends Ecol Evol 21(2):61–62,author reply 63–64.2 Prideaux GJ, et al. (2007) An arid-adapted middle Pleistocenevertebrate fauna from south-central Australia. Nature 445(7126):422–425.3 Wroe S, Field J (2006) A review of the evidence for a human role inthe extinction of Australian megafauna and an alternativeinterpretation. Quat Sci Rev 25(21-22):2692–2703.4 Barnosky AD, et al. (2011) Has the Earth’s sixth mass extinctionalready arrived? Nature 471(7336):51–57.5 Field J, Wroe S (2012) Aridity, faunal adaptations and AustralianLate Pleistocene extinctions. World Archaeol 44(1):56–74.6 Roberts RG, et al. (2001) New ages for the last Australianmegafauna: Continent-wide extinction about 46,000 years ago.Science 292(5523):1888–1892.7 Field J, Fillios M, Wroe S (2008) Chronological overlap betweenhumans and megafauna in Sahul (Pleistocene Australia-New Guinea):A review of the evidence. Earth Sci Rev 89(3-4):97–115.8 Miller GH, et al. (1999) Pleistocene extinction of Genyomisnewtoni: Human impact on australian megafauna. Science 283(5399):205–208.9 Wroe S, Field J, Fullagar R, Jermiin LS (2004) Megafaunal extinctionin the late Quaternary and the global overkill hypothesis. Alcheringa28(1):291–331.10 Faith JT, O’Connell JF (2011) Revisiting the late Pleistocenemammal extinction record at Tight Entrance Cave, southwesternAustralia. Quaternary Research 76(3):397–400.11 Ayliffe LK, et al. (2008) Age constraints on Pleistocenemegafauna at Tight Entrance Cave in southwestern Australia. QuatSci Rev 27(17–18):1784–1788.12 Price GJ, et al. (2011) Dating megafaunal extinction on thePleistocene Darling Downs, eastern Australia: The promise and pitfallsof dating as a test of extinction hypotheses. Quat Sci Rev 30(7-8):899–914.13 Brook BW, et al. (2007) Would the Australian megafauna havebecome extinct if humans had never colonised the continent?Comments on “A review of the evidence for a human role in theextinction of Australian megafauna and an alternative explanation”by S. Wroe and J. Field. Quat Sci Rev 26(3–4):560–564.14 Hesse PP, Magee JW, van der Kaars S (2004) Late Quaternaryclimates of the Australian arid zone: A review. Quat Int 118–119:87–102.15 Allen J, O’Connell JF (2008) Getting from Sunda to Sahul.Colonisation, Seafaring and the Archaeology of MaritimeLandscapes, eds Clark G, Leach BF, O’Connor S (ANU Press,Canberra, Australia), pp 31–46.16 Summerhayes GR, et al. (2010) Human adaptation and plant usein highland New Guinea 49,000 to 44,000 years ago. Science330(6000):78–81.17 O’Connell J, Allen J (2012) The restaurant at the end of theuniverse: Modelling the colonisation of Sahul. AustralianArchaeology 74:5–17.18 Davidson I (2013) Peopling the last new worlds: The firstcolonisation of Sahul and the Americas. Quat Int 285:1–29.19 O’Connell JF, Allen J (2007) Pre-LGM Sahul (Pleistocene Australia-New Guinea) and the archaeology of early modern humans.Rethinking the Human Revolution: New Behavioural and Biological
Perspectives on the Origins and dispersal of Modern Humans, edsMellars P, Boyle K, Bar-Yosef O, Stringer C (McDonald Institute forArchaeological Research, Cambridge, UK), pp 395–410.20 Wroe S, Crowther M, Dortch J, Chong J (2004) The size of thelargest marsupial and why it matters. Proc Biol Sci 271(Suppl 3):S34–S36.21 Fry BG, et al. (2009) A central role for venom in predation byVaranus komodoensis Genyoris newtoni (Komodo Dragon) andthe extinct giant Varanus (Megalania) priscus. Proc Natl Acad SciUSA 106(22):8969–8974.22 Wroe S, Argot C, Dickman C (2004) On the rarity of big fiercecarnivores and primacy of isolation and area: Tracking largemammalian carnivore diversity on two isolated continents. Proc BiolSci 271(1544):1203–1211.23 Horton DR (1984) Red kangaroos: Last of the Australianmegafauna. Quaternary Extinctions: A Prehistoric Revolution, edsMartin PS, Klein RG (Univ of Arizona Press, Tucson, AZ), pp 639–690.24 Helgen KM, Wells RT, Kear BP, Gerdtz WR, Flannery TF (2006)Ecological and evolutionary significance of sizes of giant extinctkangaroos. Aust J Zool (1987) 54(4):293–303.25 Garvey J, Cochrane B, Field J, Boney C (2011) Modern emu(Dromaius novaehollandiae) butchery, economic utility andanalogues for the Australian archaeological record. EnvironmentalArchaeology 16(2):97–112.26 Wroe S (2002) A review of terrestrial mammalian and reptiliancarnivore ecology in Australian fossil faunas and factors influencingtheir diversity: The myth of reptilian domination and its broaderramifications. Aust J Zool 50(1):1–24.27 Van Dyck S (1982) Antechinus puteus (Marsupialia: Dasyuridae),a new fossil species from the Texas Caves, Southeastern Queensland.Australian Mammalogy 5(1):59–68.28 Cramb J, Hocknull S (2010) Two new species of AntechinusMacleay (Dasyuridae: Marsupialia) from mid-Pleistocene cavedeposits in eastern central Queensland. Australian Mammalogy32(2):127–144.29 Lundelius EL, Jr. (1989) The implications of disharmoniousassemblages for Pleistocene extinctions. J Archaeol Sci 16(4):407–417.30 Price GJ (2004) Fossil bandicoots (Marsupialia, Peramelidae) andenvironmental change during the Pleistocene on the Darling Downs,southeastern Queensland, Australia. Journal of SystematicPalaeontology 2(4):347–356.31 Price GJS, Sobbe IH (2005) Pleistocene palaeoecology andenvironmental change on the Darling Downs, southeasternQueensland, Australia. Memoirs of the Queensland Museum 51(1):171–201.32 Price GJ (2012) Plio-Pleistocene climate and faunal change incentral eastern Australia. Episodes 35(1):160–165.33 Price GJ, Tyler MJ, Cooke BN (2005) Pleistocene frogs fromthe Darling Downs, south-eastern Queensland, and theirpalaeoenvironmental significance. Alcheringa 29(1):171–182.34 Field J, Wroe S, Trueman CN, Garvey J, Wyatt-Spratt S (2013)Looking for the archaeological signature in Australian megafaunalextinctions. Quat Int 285:76–88.35 Hocknull SA, Zhao J-X, Feng Y-X, Webb GE (2007) Responses ofQuaternary rainforest vertebrates to climate change in Australia.Earth Planet Sci Lett 264(1-2):317–331.36 Flannery TF (1990) Pleistocene faunal loss: Implications ofthe aftershock for Australia’s past and future. Archaeology inOceania 25(2):45–67.
37 Barnosky AD, Lindsey EL (2010) Timing of Quaternarymegafaunal extinction in South America in relation to human arrivaland climate change. Quat Int 217(1-2):10–29.38 Turney CSM, et al. (2008) Late-surviving megafauna in Tasmania,Australia, implicate human involvement in their extinction. Proc NatlAcad Sci USA 105(34):12150–12153.39 Brook BW, Bowman DMJS (2004) The uncertain blitzkrieg ofPleistocene megafauna. J Biogeogr 31(4):517–523.40 Rule S, et al. (2012) The aftermath of megafaunal extinction:Ecosystem transformation in Pleistocene Australia. Science 335(6075):1483–1486.41 Cosgrove R, et al. (2010) Overdone overkill—The archaeologicalperspective on Tasmanian megafaunal extinctions. J Archaeol Sci37(10):2486–2503.42 Prescott GW, Williams DR, Balmford A, Green RE, Manica A(2012) Quantitative global analysis of the role of climate and peoplein explaining late Quaternary megafaunal extinctions. Proc Natl AcadSci USA 109(12):4527–4531.43 Hesse PP (1994) The record of continental dust in Tasman Seasediments. Quat Sci Rev 13(3):257–272.44 Kershaw P, Moss P, Kaars S (2003) Causes and consequences oflong-term climatic variability on the Australian continent. Freshw Biol48(7):1274–1283.45 Peerderman FM, Davies PJ, Chivas AR (1993) The stable oxygenisotope signal in shallow-water, upper-slope sediments off the GreatBarrier Reef (Hole 820A). Proceedings of the Ocean Drilling Program- Scientific Results 133:163–173.46 Hope G, et al. (2004) History of vegetation and habitat change inthe Austral-Asian region. Quat Int 118-119:103–126.47 Jouzel J, et al. (2007) Orbital and millennial Antarctic climatevariability over the past 800,000 years. Science 317(5839):793–796.48 Wolff EW, Chappellaz J, Blunier T, Rasmussen SO, Svensson A(2010) Millennial-scale variability during the last glacial: The ice corerecord. Quat Sci Rev 29(21-22):2828–2838.49 Barbante C, Fischer H, Masson-Delmotte V, Waelbroeck C,Wolff EW (2010) Climate of the last million years: New insights fromEPICA and other records. Quat Sci Rev 29(1–2):1–7.50 Cohen TJ, et al. (2012) Late Quaternary megalakes fed by thenorthern and southern river systems of central Australia: Varyingmoisture sources and increased continental aridity. PalaeogeogrPalaeoclimatol Palaeoecol 356-357:89–108.51 Murphy BP, Williamson GJ, Bowman DMJS (2012) Did centralAustralian megafaunal extinction coincide with abrupt ecosystemcollapse or gradual climate change? Global Ecology andBiogeography 21(2):142–151.52 Blois JL, McGuire JL, Hadly EA (2010) Small mammal diversity lossin response to late-Pleistocene climatic change. Nature 465(7299):771–774.53 Brace S, et al. (2012) Serial population extinctions in a smallmammal indicate Late Pleistocene ecosystem instability. Proc NatlAcad Sci USA 109(50):20532–20536.54 Turney CSM, et al. (2001) Redating the onset of burning atLynch’s Crater (North Queensland): Implications for human settlementin Australia. Journal of Quaternary Science 16(8):767–771.55 Mooney SD, et al. (2011) Late Quaternary fire regimes ofAustralasia. Quat Sci Rev 30(1–2):28–46.56 Grayson DK (2007) Deciphering North American Pleistoceneextinctions. J Anthropol Res 63(2):185–214.
Wroe et al. PNAS | May 28, 2013 | vol. 110 | no. 22 | 8781
PERS
PECT
IVE
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 20
, 202
0