Embed Size (px)
Citation preview

CL- "3 c
ANNALS OF SURGERYVol. 220. No. 1.10-18@ 1994 J. B. Lippincott Company
N. Scott Adzick, M.O., and H. Peter Lorenz, M.O.
From the UCSF Fetal Treatment Center and the Department of Surgery, University of California,San Francisco. California
ObjectiveThis review updates the surgeon about the cellular, matrix, and growth factor components of
scarless fetal wound repair.
Summary Background DataFetal ski n wound healing is characterized by the absence 01 scar tissue lormation. This uniquerepair process is not dependent on the sterile, aqueous intrauterine environment. The differencesbetween letal and adult skin wound healing appear to reflect processes intrinsic to letal tissue,such as the unique letal libroblasts, a more rapid and ordered deposition and turnover 01 tissuecomponents, and, particularly, a markedly reduced inflammatory inliltrate and cytokine prolile.Scarless letal wounds are relatively delicient in the inflammatory cytokine, translorming growthlactor,a (TGF-,a). In contrast, the librosis characteristic 01 adult wound repair may be associatedwith TGF-,a excess. Recent experimental studies suggest that specilic anti- TGF-,a therapeuticstrategies can ameliorate scar lormation in adult wound repair and librotic diseases. Inhibitors 01
TGF-,a may be important luture drugs to control scar.
ConclusionsBased on the scarless letal wound repair model, a number 01 ways in which the matrix and cellularresponse 01 the healing adult wound might be manipulated to reduce scarring are reviewed.
Fetal surgical skin wounds heal rapidly and withoutthe scarring and inflammation that accompany adultskin wounds. In all species examined (mice, rats, rabbits,pigs, sheep, and monkeys), the prenatal wound healingprocess is faster and more efficient than adult repair andproduces new tissue rather than scar.1 Similarly, humanfetal surgery has shown that the younger the fetus is at
the time of surgery, the less likely he/she will be bornwith surgical scars. How does this happen? A summaryof some of the phenomenologic differences betweenadult and fetal repair is shown in Table 1.
We propose that scarless fetal repair is a consequenceof a unique extracellular matrix produced by the fetalfibroblast in the absence of an adult-like inflammatoryresponse to injury. Unraveling the biology offetal repairhas led to novel strategies for the prevention and treat-ment of scarring and fibrosis.
THE FETAL ENVIRONMENT
There are multiple differences between the fetal andadult environment that can influence wound repair.
Supported by Nationallnstitutes ofHealth grants HD 25505 and GM27345.
Address reprint requests to N. Scott Adzick, M.D., The fetal Treat-ment Center, University ofCalifornia, San Francisco, 3rd and Par-nassus A venues, Room 1601 HSW, San Francisco, CA 94143-0570.
Accepted for publication October 10,1993.
10

11Cells, Matrix, Growth Factors, and the SurgeonVo/220oNo1
Table 1. COMPARISON OFADUL T AND FETAL SKIN WOUND
HEALlNG CHARACTERISTICS
Wound HealingCharacteristics Adult Fetus
Sca,Gel! ProliferationSpeed to closure
ScabOxygen tensionFluid environmentSterile environment
Skin temperatureAcute inflammationMatrix depositionTGFp' & bFGF
\ngiogenesisEpithelializationKeratinization
PresentSlowerSlowerPresentGreaterAbsentAbsentCoolerGreater
Slower. disorganizedGreaterGreaterSlowerPresent
AbsentFasterFasterAbsentLesserPresentPresentWarmerLesser
Faster, organizedLesserLesserFasterImmature
First, fetal skin wounds are continuously bathed inwarm, sterile amniotic fluid that is rich in growth factors.Amniotic fluid also is a fertile source of extracellular ma-trix molecules, such as hyaluronic acid and fibronectin,which are important components in fetal skin wounds.2Second, fetal tissue oxygenation is much less than that ofadult tissue. We have measured tissue pO2 in fetal sheepat midgestation, a time at which scarless skin healing oc-curso Using a miniaturized oximeter probe, we foundthat fetal sheep tissue pO2 is only 16 mm Hg, whereasadult tissue pO2 is 45 mm Hg to 60 mm Hg. These find-ings seem paradoxical because adult wound studies haveshown that wound hypoxia can result in delayed healing,impaire~ leucocyte function, and increased infection.3Finally, the profile of growth factors in fetal and adultserum are different -e.g., fetal serum contains muchhigher levels ofinsulin-like growth factor 11 and hyaluro-nic acid stimulating factor.4.5
Basically, there are two ways to evaluate the role oftheenvironment in the fetal wound healing process -eitherpul adult skin in the fetal environment, or place fetalskin in the adult environment, then determine the effectofthe environment on healing. To investigate the influ-ence of the fetal environment in modulating postnatalwound healing, we transplanted adult sheep ski n anta60-day gestation (term = 145 days) fetallambs.6 In ibisexperiment, the engrafted adult skin is bathed in amni-otic fluid, perfused by fetal blood, and the immature fetalimmune system will not reject the adult skin graft. Theadult grafts were wounded 40 days later (100 days gesta-lían), at a time at which scarless repair occurs in fetalsheep. The wounds were analyzed by collagen immuno-histochemistry and were shown to heal with scar forma-
tion. Neither an amniotic fluid environment nor perfu-sion by fetal blood prevented scar formation in thewounded adult skin graft. This study suggests that scar-less fetal skin healing properties are intrinsic to fetal ski~and are not caused by the fetal environment.
Support for ibis concept comes [rom the flip sirle ex-periment -fetal tissue healing in the adult environ-mento Ferguson andcolleagues at the U niversity ofMan-chester in England have done these experiments using anopossum called theMonodelphus domesticus. At birtb,the opossum is physiologically and functionally a fetusand the pouch young remain attached to the mother'snipple for 4 weeks after birth. So ~he "fetal" developmentof ibis marsupial continues in ibis nonsterile environ-ment and in the absence ofamniotic fluido Wounded 2-day old pouch young heal rapidly and scarlessly outsideof the sterile, fluid uterine environment, whereas olderanimals show extensive scarring.' Similarly, using novelin vitro systems, both lhara and Martin have shown thatisolated fetal rat or mouse tissue grown in organ culturemedia can heal wounds without scar formation.8.9 Thus,amniotic fluid and fetal blood components, such asplatelets, are not required for scarless repair.
Despite a relatively constant intrauterine environ-ment, fetuses heal without scar early in gestation and be-gin to scar late in ges~ation.IO We have studied the tem-poral sequence ofrepair outcomes in early, mid, and lategestation fetal rhesus monkeys. Early gestation primatelip wounds heal with regeneration of all skin elements,including collagen, hair follicles, sebaceous glands, andeven muscle in the deeper tissue layers. However, as ges-tation proceeds, the primate fetus first loses its ability toregenerate normal hair follicle and other appendage pat-terns, but remains able to resto re a normal, reticular col-lagen pattern after wounding. This pattern of "transi-tion" wound repair is not regeneration or classic scar be-cause the wound collagen organization is unchangedfrom skin that is not wounded. By the early third trimes-ter, a complete switch to adult-type repair occurs withwounds showing densely packed; disorganized collagendeposition characteristic of scar .11
The experiments described in ibis section led to thehypothesis that fetal healing must involve different cellu-lar and connective tissue events iban ádult repair, andthat ibis process is independent ofthe unique fetal envi-ronment. In addition, we have learned thatthe type ofhealing result in fetal animals algo depends on 1) the ex-tent of tissue damage -large excisional fetal skinwounds scar earlier in gestation iban incisionalwoundsl2; 2) the type oftissue that is injured -internalfetal tissues, such as the diaphragm muscle, stomach,and peritoneum, heal with scarformationl3; and 3) thespecies of animal -fetal rabbits do n<>:t healexcisionalskin wounds.14
~
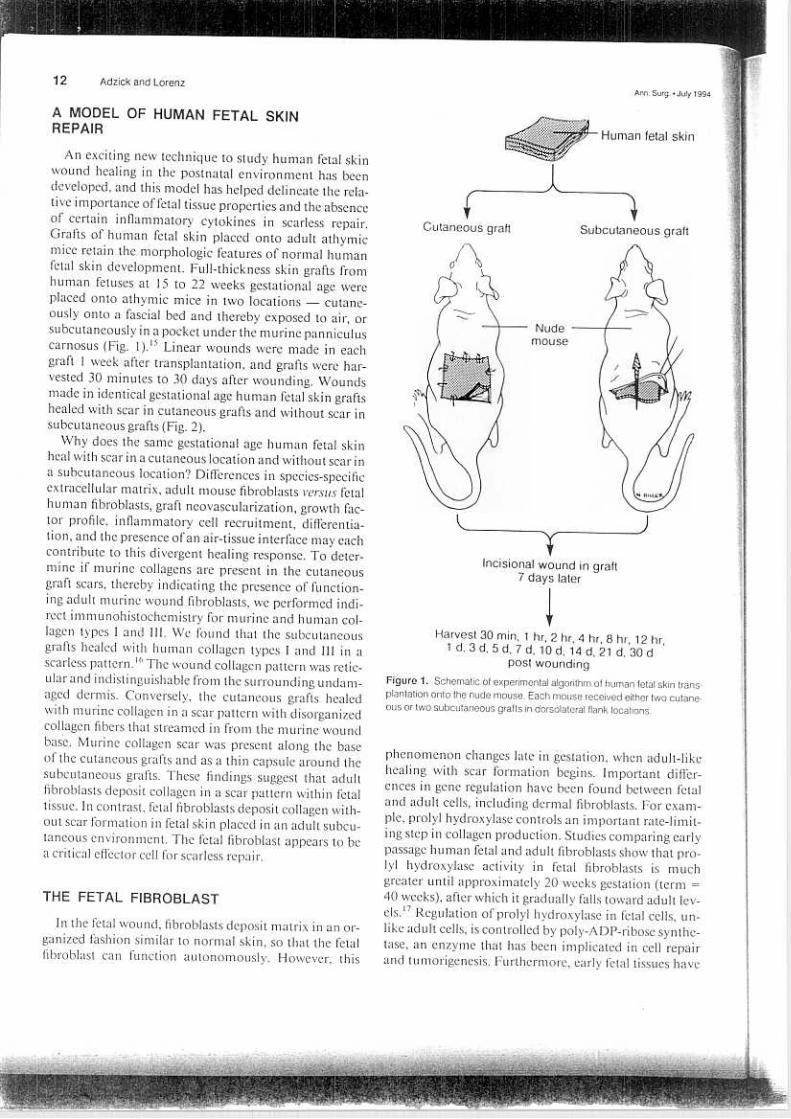
12 Adzick and LorenzAnn Sur9 o July 1994
A MODEL OF HUMAN FETAL SKINREPAIR Human fetal skin
( ' Cutaneous graft Subcutaneous graft
~~
~
---Nude
mouse
/ A/
'\'\\
!
An exciting new technique to study human fetal skinwound healing in the post natal environment has beendeveloped, and ibis model has helped delineate the rela-tive importance of fetal tissue properties and the absenceof certain inflammatory cytokines in scarless repair.Grafts of human fetal skin placed onto adult athymicmice retain the morphologic features of normal humanfetal skin development. Full-thickness skin grafts fromhuman fetuses at 15 to 22 weeks gestational age wereplaced onto athymic mice in two locations -cutane-ously onto a fascial bed and thereby exposed to air, orsubcutaneously in a pocket under the murine panniculuscarnosus (Fig. 1).15 Linear wounds were made in eachgraft l week after transplantation, and grafts were har-vested 30 minutes to 30 days after wounding. Woundsmade in "identical gestational age human fetal skin graftshealed with scar in cutaneous grafts and without scar insubcutaneous grafts (Fig. 2).
Why does ~he same gestational age human fetal skinheal with scar in a cutaneous location and without scar ina subcutaneous location? Differences in species-specificextracellular matrix, adult mouse fibroblasts verSltS fetalhuman fibroblasts, graft neovascularization, growth fac-tor profile, inflammatory cell ret:ruitment, differentia-tion, and the presence oran air-tissue interface mar eachcontrib~te to ibis divergent healing response. To deter-mine if murine collagens are pr~sent in the cutaneousgraft scars, thereby indicating the presence of function-ing adult murine wound fibroblasts, we performed indi-rect immunohistochemistry fQr murine and human col-lagen types I and 111. We found that the subcutaneousgrafts healed with human collagen types I and 111 in ascarless pattern.16 The wound collagen pattern was retic-ular and indistinguishable from the surrounding undam-aged dermis. Conversely, the cutaneous grafts healedwith murine collagen in a scar pattern with disorganizedcollagen fibers that streamed in from the murine woundbase. Murine collagen scar was present along the baseQfthe cutaneous grafts and as a thin capsule around thesubcutaneous grafts. These findings suggest that adultfibroblasts deposit collagen in a scar pattern within fetaltissue. In cpntrast, fetal fibroblasts deposit collagen with-out scar formation In fetal skin placed in an adult subcu-taneous environment. The fetal fibroblast appears to bea critical effector cell for scarless repair.
L~_-t-~-~Incisional wound in graft
7 days later
Harvesl 30 min, 1 hr, 2 hr, 4 hr, 8 hr, 12 hr,1 d. 3 d, 5 d. 7 d, 10 d, 14 d, 21 d, 30 d
post woundingFigure 1. Schematic 01 experimental algorithm 01 human letal skin trans-
plantation anta the nude mouse. Each mouse received either two cutane-ous or two subcutaneous gralts in dorsolateral flank locations.
phenomenon changes late in gestation, when adult-likehealing with scar formation begins. Important differ-ences in gene regulation have been found between fetaland adult cells, including dermal fibroblasts. For exam-pIe, prolyl hydroxylase controls an important rate-limit-ing step in collagen production. Studies comparing earlypassage human fetal and adult fibroblasts show that pro-lyl hydroxylase activity in fetal fibroblasts is muchgreater until approximately 20 weeks gestation (term =40 weeks), after which it gradually falls toward adult lev-els.17 Regulation ofprolyl hydroxylase in fetal cells, un-like adult cells, is controlled by poly-ADP-ribose synthe-tase, an enzyme that has been implicated in cell repairand tumorigenesis. Furthermore, early fetal tissues have
THE FETAL FIBROBLAST
In the fetal wound, fibroblasts deposit matrix in an or-ganized fashion similar to normal skin, so that the fetalfibroblast can function autonomously. However, this
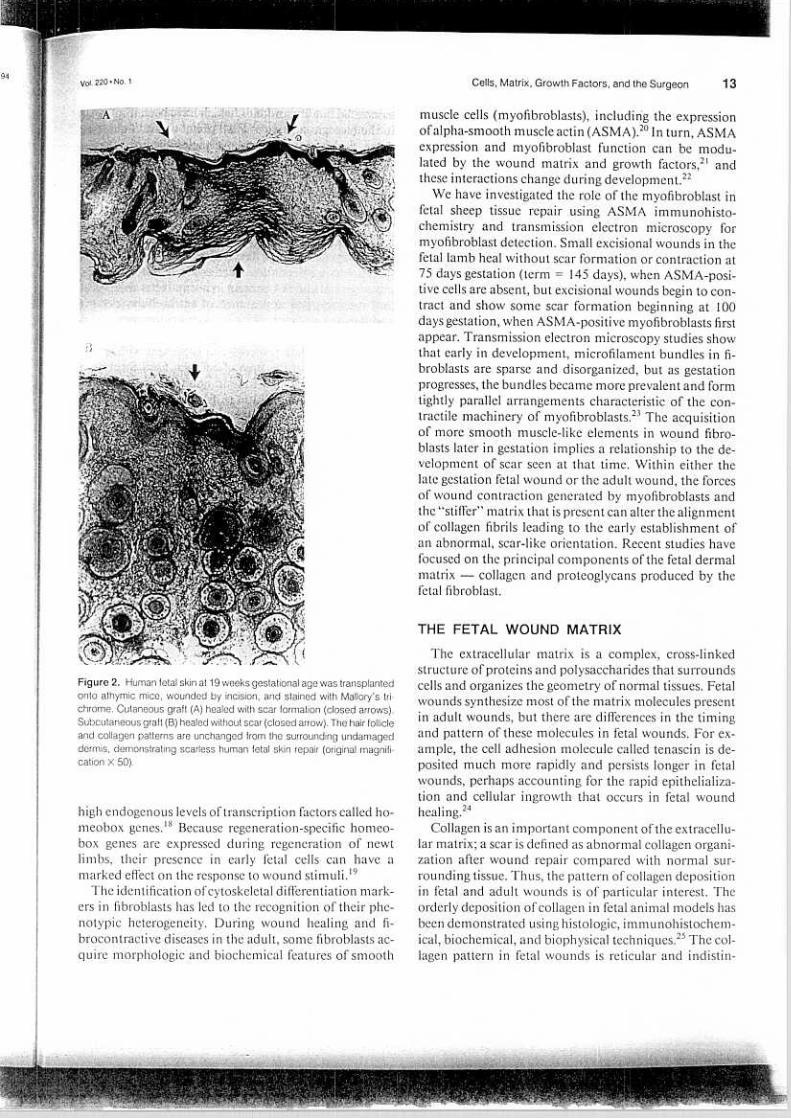
Vol. 220' No. 1 13Cells, Matrix, Growth Factors, and the Surgeon
~~
muscle cells (myofibroblasts), includirig the expressionof alpha-smooth muscle actin (ASMA)!O In turn, ASMAexpression and myofibroblast function can be modu-lated by the wound matrix and growth factors,21 andthese interactions change during development.22
We have investigated the role ofthe myofibroblast infetal sheep tissue repair using ASMA immunohisto-chemistry and transmission electron microscopy formyofibroblast detection. Small excisional wounds in thefetallamb heal without scar formation or contraction at75 days gestation (term = 145 days), when ASMA-posi-tive celIs are absent, but excisional wounds begin to con-tract and show some scar formation beginning at 100days gestation, when ASMA-positive myofibroblasts firstappear. Transmission electron microscopy studies showthat early in development, microfilament bundles in fi-broblasts are sparse and disorganized, but as gestationprogresses, the bundles became more prevalent and formtightly paralIel arrangements characteristic of the con-tractile machinery of myofibroblasts.23 The acquisitionof more smooth muscle-like elements in wound fibro-blasts later in gestation implies a relationship to the de-velopment of scar seen at that time. Within either thelate gestation fetal wound or the adult wound, the forcesof wound contraction generated by myofibroblasts andthc "stiffer" matrix that is present can alter the alignmentof colIagen fibrils leading to the early establishment ofan abnormal, scar-like orientation. Recent studies havefocused on the principal components ofthe fetal dermalmatrix -collagen and proteoglycans produced by thefetal fibroblast.
~~¡3~
'. :-
Figure 2. Human fetal skin at 19 weeks gestational age was transplantedanta athymic mice, wounded by incision, and stained with Mallory's tri-chrome. Cutaneous graft (A) healed wilh scar formation (closed arrows).Subcutaneous graft (8) healed without scar (closed arrow). The hair follicleand collagen patterns are unchanged from the surrounding undamaged
dermis, demonstrating scarless human fetal ski n repair (original magnifi-cation X 50).
THE FETAL WOUND MA TRIX
The extracellular matrix is a complex, cross-linkedstructure ofproteins and polysaccharides that surroundscells and organizes the geometry of normal tissues. Fetalwounds synthesize most of the matrix molecules presentin adult wounds, but there are differences in the timingand pattern of these molecules in fetal wounds. For ex-ample, the cell adhesion molecule calIed tenascin is de-posited much more rapidly and persists longer in fetalwounds, perhaps accounting for the rapid epithelializa-tion and celIular ingrowth that occurs in fetal woundhealing.24
ColIagen is an important component ofthe extracelIu-lar matrix; a scar is defined as abnormal COlIagen organi-zation after wound repair compared with normal sur-rounding tissue. Thus, the pattern of colIagen depositionin fetal and adult wounds is of particular interest. Theorderly deposition of colIagen in fetal animal models hasbeen demonstrated using histologic, immunohistochem-ical, biochemical, and biophysical techniques.25 The col-lagen pattern in fetal wounds is reticular and indistin-
high endogenous levels oftranscription factors called ho-meobox genes.18 Because regeneration-specific homeo-box genes are expressed during regeneration of newtlimbs, their presence in early fetal cells can have amarked effect on the response to wound stimuli.19
The identification of cytoskeletal differentiation mark-ers in fibroblasts has led to the recognition of their phe-notypic heterogeneity. During wound healing and fi-brocontractive diseases in the adult, some fibroblasts ac-quire morphologic and biochemical features of smooth

14 Adzick and Lorenz Ann Surg
~
guishable from adjacent normal tissue, whereas the adultwound contains large, parallel collagen bundles that areoriented perpendicular to the wound surface.IO Thus,scarless fetal wound healing must reflect the organizationof collagen, not the absence of collagen in the fetalwound matrix.
Although collagen types 1, 111, V, and VI are present inboth fetal and adult wounds,IO.26 there are a few knowndifferences in dermal collagen between the fetus and theadulto The presence ofaminopropeptides oftype I colla-gen in heterogeneous collagen fibrils is more common inthe fetus compared with the adult!7 The fetal dermis alsocontains a preponderance of type 111 collagen,28 and asthe fetus develops, the ratio of type lII/type I collagendecreases, which may influence collagen fibril size. Smalldiameter collagen I fibrils with a high turnover and a sim-ilar configuration to fetal collagen (in terms of attachedtype III collagen and presence of type I aminopropep-tide) occur adjacent to the epidermal/dermal junction inadult skin,27 which is the region that shows mini mal scar-ring in adult wounds. These minar differences betweencollagen in the fetal and adult dermis may be importantin modulating the nature ofthe wounding response.
Alterations in the synthesis ofproteoglycans and theirconstituent glycosaminoglycans correlate with the cellproliferation, migration, and collagen synthesis that ac-company adult wound healing!9 Proteoglycans are aheterogeneous group of polyanionic macromoleculesthat consist of a protein CaTe, to which a variable numberof linear, sulfated glycosaminoglycan chains are boundcovalently. These macromolecules include versican, alarge chondroitin sulfate proteoglycan; decorin, a smalldermatan sulfate proteoglycan; and heparan sulfate pro-teoglycan. Proteoglycans and glycosaminoglycans ha vebeen shown to affect wound collagen organization andfibrillogenesis, and by binding to specific binding sites oncollagen, control its rate of degradation.30 The sulfatedglycosaminoglycans temporally follow hyaluronic acidin the adult healing process, and the decline of hyaluro-nic acid levels and the appearance ofsulfated glycosami-noglycans have been shown in several embryologic sys-tems to correlate with the onset of cytodifferentiation.31
The fetal wound matrix is rich in glycosaminogly-cans -in fetal rabbit wounds, the glycosaminoglycancontent is approximately 3 times that ofthe adult woundduring the same time period, and approximately lOtimes that found in fetal skin that is not wounded.32 Hy-aluronic acid (HA), the principal glycosaminoglycanpresent in fetal wounds, is a large molecule composed ofalternating units of glucuronic acid and N-acetylglucos-ami ne. An HA-rich matrix is permissive for cell motilityand proliferation, and a prolonged presence ofHA in fe-tal wounds may pro vide the matrix signal that orches-trates healing by regeneration. Mammalian HA al so has
associated binding proteins, which have been implicatedin the biologic activity of HA.33 Fetal wound fluid, fetalurine, and amniotic fluid have the ability to stimulateand sustain HA synthesis because of a unique hyaluronicacid-stimulating activity. Recent support for the impor-tant role of HA in the fetal wound healing process is thatlevels ofboth HA and hyaluronic acid-stimulating activ-ity in fetallamb wound fluid decrease significantly dur-ing the transition period from fetal-like to adult healingat 120 days gestation in fetal sheep:4 Similarly, there is agestational age-dependent decrease in both total glycos-aminoglycan and HA content in normal fetal sheep skin,and the temporal appearance of extracelIular decorinand heparan sulfate proteoglycan coincides with the on-set of scarring that begins during late gestation.34 FinalIy,chondroitin sulfate proteoglycan is present within fetalmouse wounds at the time of colIagen fibril formation,but it is absent at that time in adult mouse wounds.26Thus, HA and chondroitin sulfate are likely importantfor scar-free repair, whereas other sulfated glycosamino-glycans may playa role in scar formation.
The primary wound "scaffolding" into which fibro-blasts migrate has a very important influence on colIagenfibrilIogenesis. In the fetal wound, fibroblasts can mi-grate rapidly into a loose honeycomb matrix containinghigh levels ofhyaluronic acid. In contrast, adult woundsexhibit slow fibroblast migration into a denser, more re-sistant wound matrix where fibroblast migration occursmore easily along the wound margins. These initialdifferences in the migration and orientation of fibro-blasts may establish the pattern of collagen fibrils de pos-ited in the respective wounds, with a loose reticular struc-ture in the fetus and a closely packed, disorganized ar-rangement in the adulto
THE ROLE OF INFLAMMA TION ANOCYTOKINES
There are majar differences in the degree of inflam-mation in fetal and adult wounds. Until mid-gestation,the fetus is significantly neutropenic and has not devel-oped self/non-self immunologic identity.36 Histologi-cally, there are very few, ifany, polymorphonuclear leu-kocytes in fetal wounds,37 and there may be a defect inimmature polymorphonuclear leukocyte chemotacticability.38 Recent studies have correlated the absence ofscarring in fetal wounds with the sparse inflammatoryresponse, as evidenced by markedly reduced macro-phage and monocyte infiltrates,7 absence of endogenousimmunoglobulins at the wound site,IO.2S reduced angio-genesis, and altered levels of peptide growth factors.39The transition of the fetal healing phenotype to a scar-ring, adult phenotype in the marsupial correlates directlywith the amount ofinflammatory reaction at the wound

15Cells, Matrix, Growth Factors, and the SurgeonVol. 220.No 1
site.40 These studies suggest that immature fetal immunecells do not respond to the wounding stimulus in a sim-ilar fashion to adult cells.
Peptide growth factors are released by inflammatorycells and help provide a cell-to-cell and cell-to-matrixcommunicationsystem. These cytokines can affect ma-trix synthesis, matrix degradation, cell proliferation, andcell recruitment to the wound site. Because ofthe prom-inent role that inflammation plays in adult tissue repair,the characteristic inflammatory mediators of adultwound healing may be absent or modified in fetalwounds. A different cytokine profile in fetal wounds, aconsequence of the relative lack of inflammatory cells,can cause matrix molecule differences that lead to scar-free healing.
TRANSFORMING GROWTH FACTOR-BET A (TGF-fJ)
Of the many cytokines that have been implicated inwound healing, TGF-{:J affects all phases of the healingprocess, including the inflammatory response and ma-trix accumulation.41 In the adult wound, the macro-phage is a crucial inflammatory cell that releases TGF-{:J and other cytokines. The mammalian TGF-{:J familyconsists ofthree known isoforms, TGF-{:J 1, TGF-{:J2, andTGF-{:J3, that are structurally and functionally closely re-lated to one another. Through autocrine and paracrinemechanisms, TGF-{:J stimulates the deposition of colla-gen and other matrix components by fibroblasts, inhibitscollagenase, blocks plasminogen inhibitor, enhances an-giogenesis, and is chemotactic for fibroblasts, monocytesand macrophages.42 Thus, TGF-{:J is capable ofstimulat-ing fibrogenesis by the fibroblast.
Transforming growth factor-{:J may provide the linkbetween injury, inflammation, and fibrosis. After tissueinjury, activated macrophage-derived TGF-{:J disturbsthe balance of synthesis and degradation of collagens andcauses accumulation of extracellular matrix. Transform-ing growth factor-{:J induces alpha-smooth muscle actinexpression in fibroblasts, and this finding correlates withscar formation and pathologic wound contraction.43 Theability of TGF-{:J to induce its own production may becrucial for the development of progressive scarring inchronic diseases that lead to eventual obliteration of nor-mal tissue architecture. The correlation between TGF-fJlevels and scar formation holds true for a variety of fi-brotic diseases. Experimental intraperitoneal admin-istration of TGF-{:J results in adhesion formation.44There is enhanced expression of TGF-{:Jl at the site ofscar in the rat brain after localized cerebral injury.45 Inproliferative vitreoretinopathy, total TGF-{:J levels in in-traocular fluid increase as the retinal scarring progressesfrom mild to moderate to severe.46 Markedly increased
amounts of TGF-fJ are present in fibrogenic diseasessuch as cirrhosis,47 interstitial pulmonary fibrosis,48 glo-merulonephritis,49.S0 and scleroderma.SI
Exogenously applied TGF-fJ promotes scar formationin both adult and fetal wounds. Specifically, the additionofTGF-fJ to polyvinyl alcohol sponges implanted in fetalrabbits produces fibrosis.S2 We have demonstrated thatwhen TGF-fJ is added to human fetal skin wounds via aslow release disk, scar formation results.S3 In vitro studieshave shown that exposure of fetal dermal fibroblasts toTGF-fJ results in marked upregulation of collagen geneexpression.S4 Thus, the cellular and matrix machinerythat is necessary for scar formation exists in fetalwounds.
Fibroblasts are not only capable of responding to thenumerous cytokines produced by the immune system,but they can synthesize and secrete, on their own, growthfactors with pleotrophic effects. The technique of reversetranscription-polymerase chain reaction was used to de-tect differences in TGF-fJ-specific mRNA production byfetal and adult sheep fibroblasts.SS To test whether fetaland adult fibroblasts have different responses to the lowtissue pO2 in the fetal wound environment, we exam-ined TGF-fJI gene expression in these cells under nor-moxic and hypoxic conditions. Both fetal and adult fi-broblasts showed the potential to produce TGF-fJ 1 atnormoxia, but fetal fibroblasts responded to hypoxiawith a decrease in TGF-fJl transcription, whereas adultfibroblasts were stimulated by hypoxia to increase TGF-fJ transcription. Thus, the low fetal wound pO2 maymarkedly downregulate TGF-fJI gene expression.
This series of studies led to the hypothesis that scarlessfetal wounds may be relatively TGF-fJ deficient. Whitbyhas performed TGF-fJ immunostaining studies in fetalmouse wounds and has found that TGF-fJ was absent,whereas TGF-fJ staining was abundant in neo natal andadult mouse wounds.39 We have shown an absence ofimmunostaining for either TGF-fJI or TGF-fJ2 from lhour to 28 days after wounding human fetal skin.S3 Incontrast, wounds in adult human ski n grafts demon-strated TGF-fJ 1 at the wound edge at 6 hours through 21days and TGF-fJ2 at 12 hours through 7 days. It is possi-ble that TGF-fJ levels in fetal wounds are below thethreshold for detection by immunohistochemical tech-niques, but compared with adu1t wounds, fetal woundsare at least relatively TGF-fJ deficient. Although we havedetected significant amounts of TGF-fJ in fetal sheepwound fluid, there may be an alteration or limitation ofTGF-fJ biologic activity by the local fetal wound en vi-ronment.S6
Macrophages are the principal source of TGF-fJ inadult wounds, and the reduced TGF-fJ level in fetalwounds can reflect the absent or minimal macrophageinfiltrate in these wounds. In adult wounds, TGF-fJ also

16 Adzick and Lorenz AM. Surg oJuly 1994
is released from the alpha-granules of activated platelets,so it also is possible that fetal platelets mar not synthesizeor release TGF-fl at the fetal wound site. The relative lackof TGF-fl, a cytokine known to induce fibrosis, mar bean important reason the fetus heals by regenerationrather iban by scarring. Transforming growth factor-flmar be important in adult tissue repair, but excessiveaction of ibis cytokine mar be responsible for the tissuedamage caused by scarring in many serious diseases.These findings suggest that anti- TGF-fl therapeutic strat-egies mar ameliorate scar formation in adult fibrotic dis-eases.
Table 2. ANTI- TRANSFORMING GROWTHFACTOR-BETA THERAPEUTIC STRATEGIES
Neutralizing antibodies to TGF{31 and {32
TGF{33TGF{3-binding proteoglycans
BiglycanDecorin
Soluble receptors or receptor antagonistsPrevent TGF{3 activation
Block IGF-II/mannose-6-phosphate receptorAdd latent TGF{3 binding protein
Antisense oligonucleotides
Anti-macrophage strategies
CYTOKINE EXCESS IN ADUL T WOUNDS
Shah and colleagues mimicked this fetal wound situa-tion within the healing adult rat wound by using an anti-TGF-,B polyclonal neutralizing antibody to experimen-tally reduce TGF-,B levels.S7 This manipulation resultedin markedly diminished scarring in adult wounds. Theneutralizing antibody-treated wounds had normal ten-sile strength and a nearly normal dermal architecturecompared with untreated wounds, and this salutaryeffect was accompanied by deposition of less collagenand fibronectin and infiltration by fewer macrophagcsand blood vessels. Injection ofTGF-,B alone had the op-po si te effects. Application of TGF-,B neutralizing anti-body at the time ofwounding (and not later) was essen-tial to reduce active TGF-,B levels, prevent auto-induc-tion ofTGF-,B mRNA, and limit macrophage infiltrationand further TGF-,B release. The effectiveness ofthis ap-proach has been demonstrated in another fibrotic pro-cess, beca use admi nistration of either TGF-,B 1 antiserumor decorin to neutralize TGF-,B biologic activity sup-presses the pathologic increase in matrix synthesis thatoccurs in an animal model of glomerulonephritis.49.soTh us, the relationship ofTGF-,B to scar formation fulfillsKoch's postulates -TGF-,B is absent in scarless fetalwounds, addition of TGF-,B to fetal wounds results inscar formation, the presence ofTGF-,B after adult injurycorrelates with the degree offibrosis observed, and block-ing ofTGF-,B in adult wounds has a potent anti-scarringeffect.
Subsequent studies by Shah and Ferguson have shownthat neutralization of both TGF-,Bl and TGF-,B2 iso-forros has a much greater, synergistic anti-scarring effectthan neutralization of either isoform alone.s8 Neutraliz-ing antibodies against platelet-derived growth factor alsoha ve some anti-scarring effect, but antibodies to epider-mal growth factor ha ve no effect on dermal scarring.S9Although injection of cytokine antibodies into woundshas very limited clinical potential because of antigenicityproblems, there are other promising ways to reducewound levels of TGF-,B 1 and TGF-,B2. Addition of the
TGF-,83 isoform downregulates TGF-,8 1 and ,82 levelsand leads to a pronounced anti-scarring resulto The ap-plication to rodent wounds of the inexpensive andreadily available sugar mannose-6-phosphate also limitsscar formation, presumably by blocking the insulin-likegrowth factor-lI/mannose-6-phosphate receptor that isimportant for TGF-,8 activation.S9 Various other theo-retical anti- TGF-,8 therapeutic strategies, such as flood-ing the wound with soluble TGF-,8 receptors to competeeffectively with cellular TGF-,8 binding sites or addingantisense oligonucleotides to inhibit TGF-,8 gene expres-sion, al so can make adult wounds heal in a fetal-likemanner (Table 2). Inhibitors of TGF-,8 may be impor-tant future drugs for the control offibrosis.
Basic fibroblast growth factor is another example ofcytokine excess in postnatal wounds. Using immunohis-tochemical techniques, basic fibroblast growth factor ispresent in neonatal and adult mouse lip wounds, but isnot detected in fetal mouse lip wounds.39 Basic fibroblastgrowth factor is a powerful angiogenesis stimulator, andincreased neovascularization is a normal component ofadult wound repair.60 Immunostaining for collagen IVand laminin, normal components of the endothelialbasement membrane, shows profuse angiogenesis inadult wounds, whereas fetal wounds ha ve a diminishedvascular pattern similar to adjacent undamaged fetal tis-sue.IO.26 A more rapid rate of adult wound neovasculari-zation would bring earlier wound perfusion by adult se-rum and inflammatory cells, thereby contributing to ascar repair pattern.
In evolutionary terms, it appears that adult woundsmay be optimized for speed of healing under adverseconditions (dirt, foreign bodies, etc.) and the result is anexcessive inflammatory infiltrate and cytokine profile.The potential fetal regenerative response in adultwounds may be overrun by an inflammation-inducedcytokine surplus leading to scar. This phenomena is anexample of"cytokine poisoning," in which the inappro-priate reparative response of the patient may prove det-

17Cells, Matrix, Growth Factors. and the SurgeonVol. 220oNo. 1
~
rimental to outcome. This paradox is analogous to thepathophysiology of toxic host mediators associated withdisease states such as systemic sepsis and multiple organfailure.6l,62
Fl.;TURE DJRECTIONS
Fetal wound healing studies can help surgeons un-derstand what accounts for scarring and, perhaps moreimportantly, how scar formation can be prevented.There are a number ofways in which the matrix and cel-lular response ofthe healing adult wound might be ma-nipulated to reduce sGarring -i.e., inhibiting the woundinflammatory response by blocking inflammatory cyto-l;ines such as TGF-fJ, basic fibroblast growth factor, a~dpiatelet-derived growth factor; adding exogenous tenas-cin to facilitate keratinocyte and fibroblast migrationinto the wound; transplanting fibroblasts with fetal char-acteristics to the adult wound site; providing a more po-rous wound scaffold by addition of hyaluronic acid orhyaluronic acid-stimulating activity to enhance fibro-blast migration and promote regeneration of a normalreticular collagen organization in the wounded dermis.Although all of these potential therapeutic strategies re-quire rigorous scientific testing, adult wound healingmay be altered toward a scar-free, fetal-like phenotypeby modifying one or more of the components that aredifferent between fetal and adult repair.
References
strate rapid collagen deposition without scar formation. J PediatrSurg 1990; 25:63-69.
11. Lorenz HP, Whitby DJ, Longaker MT, Adzick NS. The ontogenyofscar formation in the non-human primate. Ann Surg 1993; 217:391-396.
12. Longaker MT, Burd DAR, Gown AM, et al. Midgestation fetallamb excisional wounds contract in utero. J Pediatr Surg 1991; 26:942-947.
13. Longaker MT, Whitby DJ, Jennings RW, et al. Fetal diaphrag-matic wounds heal with scar formation. J Surg Res 1991; 50:375-385.
14. Krummel TM, Ehrlich HP, Nelson JM,et al. In vitro and in vivoanalysis ofthe inability offetal rabbit wounds to contract. WoundReRReg 1993; 1:15-21.
15. Lorenz HP, Longaker MT, Perkocha LA, et al. Scarless woundrepair: a human fetal skin modelo Development 1992; 114:253-259.
16. Lorenz HP, Longaker MT, Whitby DJ, et al. Scarless fetal skinrepair is int~insic to the fetal fibroblast. Surg Forum 1992; 43:696-697.
17. Duncan BW, Qian J, Liu X, Bhatnagar R. Regulation of prolylhydroxyl~e activity in fetal and adult fibroblasts. In Adzick NS,Longaker MT, eds. Fetal Woun~ Healing. New York: ElsevierSci-entific Press, 1992, pp 303-323.
18. Mackenzie A, Leeming GL, Jowett A, et al. The homeobox geneHa,\" 7. J has specific regional and temporal expression patternsdur-ing early murine craniofacial embryogenesis, especially duringtooth development in vivo and in vitro. Development 1991; 111:269-285.
19. Brown R, Brockes JP. Identification and expression ora regenera-tioij-specific homeobox gene in thc ncwt limb blastema. Develop-mcnt 1991; 111:489-496.
20. Sappino AP, Schurch W, Gabbiani G. Dilfcrcntiation repertoire offibroblastic cclls: cxprcssion of cytoskclctal proteins as markers ofphcnotypic modulations. Lab Invcst 1990; 63: 144-161.
21. Darby 1, Skalli O, Gabbiani G. Alpha-smooth muscle actin is tran-sicntly cxprcsscd by myofibroblasts during experimental woundhcaling. Lab Invest 1990; 63:21-29.
22. Sappino AP, Dictrich Py, Skalli O, et al. Colonic pericryptal fi-broblasts: dilferentiation pattern in embryogenesis and phenotypicmodulation in epithclial proliferation lesions. Virchows Archiv APathol Anat Histopathol 1990; 415:551-557.
23. Estes JM, Vande Berg JS, Adzick NS, et al. Phenotypic and func-tional features of myofibroblasts in fetal wounds. Differentiation
(in presS).24. Whitby DJ, Longaker MT, Harrison MR, et al. Rapid epitheliza-
tion offetal wounds is associated with early deposition oftenascin.J Cell Sci 1991; 99:583-586.
25. Adzick N-5, Longaker MT.Scarless wound healing in the fetus: therole ofthe extracellular matrix. Prog Clin Biol Res 1991; 365: 177-192.
26. Whitby DJ, Ferguson MWJ. The extracellular matrix of lipwóunds in fetal, neonatal, and adult mice. Developm'ent 1991;112:651-668.
27. FIeischmajer R, Perlish JS, Burgeson RE, et al. Type 1 and type I1Icollagen interactions during fibrillogenesis. Ann NY Acad Sci
1990;580:161-175.28. Merkel JR, DiPaolo BR, Hallock GG, Rice DC. Type 1 and type
I11 cóllagen content ofhealing wounds in fetal and adult rats. ProcSoc E~p Biol Med 1988; 187:493-497.
29. YeoTK, Brown L, Dvorak HF. Alterations in proteoglycan syn-thesis com~on to healing wounds and tumors. Amer J Path 1991;138: 1437-1450.
l. Adzick NS. Longaker MT. eds. Fetal Wound Healing. New York:Elsevier Scientific Press, 1992.
2. Longaker MT, Adzick NS. Hall JL. et al. Studies in fetal woundhealing VII: fetal wound healing mar be modulated byelevatedhyaluronic acid stimulating activity in amniotic nuid. J PediatrSurg 1991; 25:430-433.
3. Jonsson K. Jensen JA, Goodson WH, et al. Tissue oxygenation,anemia. and pcrfusion in relation to wound healing in surgical pa-tients. Ann Surg 1991; 214:605-613.
4. Longaker MT. Chiu ES, Harrison MR, et al. Studies in fetal woundhealing IV: hyaluronic acid stimulating activity distinguishes fetalfrom adult wound nuid. Ann Surg 1989; 210:667-672.
5. Estes JM. Spcncer EM, Longaker MT, Adzick NS. Insulin-likegrowth factor-11 in ovine wound nuid: evidence for developmentalregulation. Surg Forum 1991; 42:659-:661.
6. Longaker MT, Whitby OJ, Ferguson MWJ, et al. Adult skinwounds in the fetal environment heal with scar formation. AnnSurg i994;219:65-72.
7. Ferguson MWJ, Howarth GF. Marsupial models ofscarless fetalwound healing. In Adzick NS, Longaker MT, eds. Fetal WoundHealing. New York: Elsevier Scientific Press, 1992, pp 95-124.
8. Ihara 1, Motobayashi Y. Wound closure in foetal rat skjn. Oevel-opment 1992; 114:573-582.
9. Martin P, Lewis J. Actin cables and epidermal movement in em-bryonic wound healing. Nature 1992; 360: 179-182.
10. Longaker MT, Whitby DJ, Adzick NS,etal. Studiesin fetal woundhealing VI: second and early third trimester fetal wounds demon-

18 Adzick and LorenzAnn Surg °July 1994
and transforming growth factor-{J type 2 le veis in the eye. J Clin
Invest 1989;83:1661-1666.47. Castilla A, Prieto J, Fausto N. Transforming growth factors beta 1
and alpha in chronic liverdiscase: Elfects ofinterferon alfa therapy.N Engl J Med 1991; 324:933-940.
48. Broekelmann TJ, Limper AH, Colby TV, McDonald JA.
Transforming growth factor {JI is prescnt at sites of extracellularmatrix gene expression in human pulmonary fibrosis. Proc NatlAcad Sci U S A 1991; 88:6642-6646.
49. Border WA, Okuda S, Languino, et al. Suppression ofexperimen-tal glomerulonephritis by antiscrum against transforming growth
factor{Jl. Nature 1990; 346:371-374.50. Border WA, Noble NA, Yamamoto T, et al. Natural inhibitor of
transforming growth factor-{J protects against scarring in experi-mental kidiley discase. Nature 1992; 360:361-364.
51. Peltonen J, Kahari L, Jaakkola S, et al. Evaluation oftransforming
growth factor {J and type 1 procollagen gene expression in fibroticdiscases by in situ hybridization. J Invest Dermatol 1990; 94:365-371.
52. Krummel TM, Michna BA, Thomas BL, et al. TGF{J induces fi-brosis in a fetal wound modelo J Pediatr Surg 1988; 23:647-652.
53. Sullivan KM, Lorenz HP, Adzick NS. The role of transforming
growth factor beta in human fetal wound healing. Surg Forum
1993;44:625-627.54. Lorenz HP, Chang J, Longaker MT, Banda MJ. Transforming
growth factors (,'1 and (,'2 synergistically incrcasc collagen gene ex-pression in fetal fibroblasts but not in adult fibroblasts. Surg Forum
1993;44:723-725.55. Chang J, Longaker MT, Lorcnz HP, ct al. Fctal and adult shccp
fibroblast TGF-{J 1 gene cxprcssion in V;//'II: effects of hypoxia and
gcstational agc. Surg Forum 1993; 44:720-722.56. Longakcr MT, Bouhana KS, Robcrts AB, ct al. Rcgulation offcta!
wound hcaling. Surg Forum 1991: 42:654-655.57. Shah M, Forcman DM. Fcrguson MWJ. Control of scarring in
adult wounds by ncutralizingantibody to transforming growth Cacotorbcta. Lancct 1992: 1:213-214.
58. Shah M. Forman D, Fcrguson M. Rcduction of scar tissuc forma-tion in adult rodcnt wound hcalingand manipulation ofthcgrowthfactor profilc. Prcsentcd at thc Sccond Annual Mccting of thcWound HcalingSocicty. Apri11992; Richmond. VA.
59. Shah M, Robcrts AB, Gold U, Ferguson MWJ. Immunolocaliza-
tion ofTGF(3 isoforms in normal and experimentally modulatcdincisional wounds in adult rodents. Wound Rep Rcg 1993; 2: 124.
60. Folkman J, Shing Y. Angiogcnesis. J Biol Chem 1992; 267: 10931-
10934.61. Baue AE. Thc horror autotoxicus and multiplc-organ failure. Arch
Surg 1992: 127:1451-1462.62. Oeitch EA. Multiple organ failure: pathophysiology and potential
future therapy. Ann Surg 1992; 216: 117-134.
30. Ruoslahti E. Yamaguchi Y. Proteoglycans as modulators ofgrowth factor activities. Cell 1991; 64:867-869.
31. Bertolami CN. Berg S. Frymiller EG. Glycosaminoglycan process-ing during tissue repair: degradation ofhylauronic acid./n AdzickNS. Longaker MT. eds. Fetal Wound Healing. New York: ElsevierScientific Press. 1992. pp 215~226.
32. DePalma RL. Krummel TM. Durham LA. et al. Characterizationand quantification of wound matrix in the fetal rabbit. Matrix1990; 9:224-231.
33. Toole BP. Hyaluronan and its binding proteins. the hyaladherins.Curr Op Cell Biol 1990; 2:839-844.
34. Estes JM. Adzick NS. Longaker MT. et al. Hyaluronate metabo-lism undergoes ontogenetic transition during fetal development:Implic~tions for scarless wound healing. J Pediatr Surg 1993; 28:1227-1231.
35. Freund RM. Siebert JW. Cabrera J. et al. Serial quantification ofhyaluronan and sulfated glycosaminoglycans in fetal sheep skin.Biochem MolBiollnt 1993; 29:813-816.
36. Flake A W. Harrison MR. Adzick NS. Zanjani ED. Transplanta-tion of fetal hematopoietic stem cells in utero: the creation of he-matopoietic chimeras. Science 1986; 223:776-778.
37. Adzick NS. Harrison MR. Glick PL. et al. Comparison offetal.newborn. and adult rabbit wound healing by histologic. enzyme-histochemical. and hydroxyproline determinations. J Pediatr Surg1985;20:315-319.
38. Jennings R W. Adzick NS. Longaker MT. et al. Ontogeny of fetalsheep polymorphonuclear leukocyte phagocytosis. J Pediatr Surg1991;26:853-855.
39. Whitby DJ. Ferguson MWJ. Immunohistochemicallocalizationofgrowth factors in fetal wound healing. Dev Bio11991; 147:207-215.
40. Armstrong J. Healing charactcristics of dermal wounds on thepouch young of the short tailed grey opossum. Wound Rcp Rcg1993; 2:96.
41. Barnard JA. Lyons RM. Moscs HL. Thc ccll biology oftr"dnsform-ing growth factor fl. Biochim Biophys Acta 1990; 1032:79:"87.
42. Sporn MB. Robcrts AB. Transforming growth factor-fl: rcccntprogrcss and ncw challcnges. J Cell Bio11992; 119: 1017-1021.
43. Dcsmoulicrc A. Gcinoz A. Gabbiani F. Gabbiani G. Transforminggrowth factor-fll induces alpha-smooth muscle actin cxprcssion ingranulation tissue myofibroblasts and in quicsccnt and growing f.-broblasts. J Ccll Bio11993; 122: 103-111.
44. Logan A. Frautschy SA. Gonzalez AM. et al. Enhanced expressionof transforming growth factor fll in the rat brain after a localizcdcerebral injury. Brain Res 1992; 587:216-225.
45. Williams RS. Rossi AM. Chcgini N. Schultz G. Effect oftransform-inggrowth factorfl on postoperative adhesion formation and intactperitoneum. J Surg Res 1992; 52:65-70.
46. Connor TB. Roberts AB. Sporn MB. et al. Correlation offibrosis
~II~~~~~~~~,.~~~





















![H O Cl][Cl]](https://img.pdfslide.us/doc/110x75/625a722790c4ca087c5c7d67/h-o-clcl.jpg)
![James Dashner - [Captiv in Labirint] 3 Tratament Letal [v.2.0]](https://img.pdfslide.us/doc/110x75/563db86e550346aa9a939dcb/james-dashner-captiv-in-labirint-3-tratament-letal-v20.jpg)





