-
8/22/2019 Cigarette Induce COPD
1/21
doi: 10.1152/ajplung.00390.2007294:L612-L631, 2008. First
published 25 January 2008;Am J Physiol Lung Cell Mol Physiol
Andrew Churg, Manuel Cosio and Joanne L. Wrightfrom animal
modelsMechanisms of cigarette smoke-induced COPD: insights
You might find this additional info useful...
198 articles, 53 of which you can access for free at:This
article
citeshttp://ajplung.physiology.org/content/294/4/L612.full#ref-list-1
15 other HighWire-hosted articles:This article has been cited
byhttp://ajplung.physiology.org/content/294/4/L612#cited-by
including high resolution figures, can be found at:Updated
information and
serviceshttp://ajplung.physiology.org/content/294/4/L612.full
can be found at:Molecular PhysiologyAmerican Journal of
Physiology - Lung Cellular andaboutAdditional material and
information
http://www.the-aps.org/publications/ajplung
This information is current as of July 31, 2013.
ISSN: 1040-0605, ESSN: 1522-1504. Visit our website at
http://www.the-aps.org/.
Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright
2008 the American Physiological Society.components of the
respiratory system. It is published 12 times a year (monthly) by
the American Physiologicalthe broad scope of molecular, cellular,
and integrative aspects of normal and abnormal function of cells
and
publishes original research coveringAmerican Journal of
Physiology - Lung Cellular and Molecular Physiology
http://ajplung.physiology.org/content/294/4/L612#cited-byhttp://ajplung.physiology.org/content/294/4/L612#cited-byhttp://ajplung.physiology.org/content/294/4/L612#cited-byhttp://ajplung.physiology.org/content/294/4/L612#cited-byhttp://ajplung.physiology.org/content/294/4/L612#cited-by
-
8/22/2019 Cigarette Induce COPD
2/21
Invited Review
Mechanisms of cigarette smoke-induced COPD: insights from animal
models
Andrew Churg,1 Manuel Cosio,2 and Joanne L. Wright1
1Department of Pathology, University of British Columbia,
Vancouver, British Columbia; and 2Respiratory Division, Royal
Victoria Hospital, and Meakins-Christie Laboratories, McGill
University, Montreal, Quebec, Canada
Churg A, Cosio M, Wright JL. Mechanisms of cigarette
smoke-induced COPD:insights from animal models. Am J Physiol Lung
Cell Mol Physiol 294: L612L631, 2008.First published January 25,
2008; doi:10.1152/ajplung.00390.2007.Cigarette smoke-induced animal
models of chronic obstructive pulmonary disease support
theprotease-antiprotease hypothesis of emphysema, although which
cells and proteasesare the crucial actors remains controversial.
Inhibition of either serine or metallo-proteases produces
significant protection against emphysema, but inhibition
isinvariably accompanied by decreases in the inflammatory response
to cigarettesmoke, suggesting that these inhibitors do more than
just prevent matrix degrada-tion. Direct anti-inflammatory
interventions are also effective against the develop-ment of
emphysema, as are antioxidant strategies; the latter again decrease
smoke-induced inflammation. There is increasing evidence for
autoimmunity, perhapsdirected against matrix components, as a
driving force in emphysema. There is
intriguing but controversial animal model evidence that failure
to repair/failure oflung maintenance also plays a role in the
pathogenesis of emphysema. Cigarettesmoke produces small airway
remodeling in laboratory animals, possibly by directinduction of
fibrogenic growth factors in the airway wall, and also
producespulmonary hypertension, at least in part through direct
upregulation of vasoactivemediators in the intrapulmonary arteries.
Smoke exposure causes goblet cellmetaplasia and excess mucus
production in the small airways and proximal trachea,but these
changes are not good models of either chronic bronchitis or
acuteexacerbations. Emphysema, small airway remodeling, pulmonary
hypertension,and mucus production appear to be at least partially
independent processes that mayrequire different therapeutic
approaches.
animals; emphysema; small airway remodeling; pulmonary
hypertension; chronicobstructive pulmonary disease
CHRONIC OBSTRUCTIVE PULMONARY DISEASE (COPD) is now thefifth
leading cause of death worldwide (126). A meta-analysisusing data
from 28 countries suggests that the prevalence ofCOPD based on
spirometric measurements is 9 10% in adultsover age 40. It has been
estimated that there are 1517 millionindividuals with COPD in the
United States alone (62, 160).
By far, the most important risk factor for COPD in thedeveloped
world is cigarette smoking; exposures to dusts,fumes, air pollution
particles, and, in the developing world,biomass fuels, are also
believed to cause COPD (62), but thereis much less information
available about these etiologies.Genetic predisposition seems to
play an important role in thedevelopment of COPD, and a variety of
genetic polymor-phisms related to levels of antiproteases
(1-antitrypsin, thebest established and the most clearly
important), metallopro-teases, proinflammatory and profibrotic
cytokines, and variousantioxidant enzymes and detoxifying enzymes
have beenlinked to COPD (reviewed in Refs. 43, 81, 147, 178).
This review will concentrate on mechanistic insights
fromlaboratory animal models of COPD caused by cigarettesmoke, both
because of the overwhelming role of cigarette
smoke as a causative agent, and because of a lack of
animalmodels based on dusts, air pollution particles, and
biomassfuels. We will emphasize in vivo models and draw on datafrom
tissue culture and explant studies where these appeardirectly
relevant; for reviews that focus more on in vitroexperiments,
molecular mechanisms, potential targets, andnon-smoke-induced COPD
models, see Refs. 7, 8, 18, 40,41, 99, 156, 197. There are plans to
cover, in a future reviewin this journal, the pros and cons of
various different animalmodels of COPD, including cigarette
smoke-induced, elas-tase-induced, and genetic manipulation-induced
models, andso these are not addressed here.
Because human COPD really consists of four anatomiclesions
(emphysema, small airway remodeling, vascular re-modeling with
pulmonary hypertension, mucus overproductionand chronic bronchitis)
and one functional lesion [acute exac-erbation (13, 26)], we have
separated our discussion into thesebroad categories. In some
senses, this separation is artificial,since many patients have
airflow limitation because of bothemphysema and small airway
remodeling, and they may havepulmonary hypertension and chronic
bronchitis and developacute exacerbations as well. However, as will
become appar-ent, the mechanisms behind these anatomic lesions may
wellbe different, so separating them is useful in
understandingpathogenesis and in devising therapeutic
approaches.
Address for reprint requests and other correspondence: A. Churg,
Dept. ofPathology, Univ. of British Columbia, 2211 Wesbrook Mall,
Vancouver, BC,Canada V6T 2B5 (e-mail:
[email protected]).
Am J Physiol Lung Cell Mol Physiol 294: L612L631, 2008.First
published January 25, 2008; doi:10.1152/ajplung.00390.2007.
1040-0605/08 $8.00 Copyright 2008 the American Physiological
Society http://www.ajplung.orgL612
-
8/22/2019 Cigarette Induce COPD
3/21
Mechanisms Related to the Protease-Antiprotease Hypothesisof
Emphysema
Chronic (generally 6 mo or longer) exposure of mice orguinea
pigs to cigarette smoke produces lesions that morpho-logically and
physiologically resemble a mild form of centri-lobular human
emphysema (184) (Fig. 1), and emphysema is
the lesion that has received the most study, although
theanatomic changes differ from those in humans in a subtle
waybecause these animals do not normally have respiratory
bron-chioles, the locus of initial destruction in human
smoke-induced centrilobular emphysema.
The classic theory of emphysema is the protease
antiproteasehypothesis. This hypothesis was formulated from the
observa-tion that humans deficient in 1-antitrypsin (A1AT)
developedearly emphysema, particularly if they smoked (85), and
fromthe experiments of Gross and colleagues (59) showing
thatinstillation of elastolytic enzymes produced emphysema
inexperimental animals. These observations lead to the idea
thatsmoke evokes an inflammatory cell reaction and that these
cellsrelease proteases that overwhelm the antiproteolytic
defenses
of the lower respiratory tract, leading to matrix destruction
andemphysema. Despite a variety of new theories, the
protease-antiprotease hypothesis (now expanded to include metallo
andcysteine proteases as well as the original serine
proteases)remains the generally accepted basis for the destruction
ofmatrix that leads to emphysema, and is the basis of
mostexperimental smoke exposure models, but exactly what
cells/proteases are crucial to this process is a complex and
contro-versial issue.
That smoke evokes an inflammatory response in both hu-mans
(reviewed in Ref. 165) and animals is clear. Everyexperimental
animal study that has looked at lavage/tissueneutrophils,
macrophages, and lymphocytes in guinea pigs,rats, and mouse strains
that develop emphysema has found an
increase, although the details of cell types, timing, and
mag-nitude of the effect vary (Tables 1, 2, 3), and smoke
inducesproinflammatory cytokine release from cultured
macrophages,epithelial cells, and fibroblasts (90, 195, 198). Most
but not all
(68, 97) authors report an increase in gene expression/proteinin
whole lung/bronchoalveolar lavage (BAL) of chemoattrac-tant and
proinflammatory mediators including TNF, IL-1,IL-8, MIP-2, MCP-1,
MIP-1, MIP-1, MCP-3, KC, PGE2,IL-12, IL-18, RANTES, and IP-10 (28,
30, 32, 61, 78, 83, 92,98, 166).
Interference with/manipulation of serine proteases. The
original formulation of the protease-antiprotease
hypothesispostulated the neutrophil, and in particular, neutrophil
elastase,as the important effectors in emphysema (76). In recent
years,the role of the neutrophil has become controversial;
somereports have shown localization of neutrophils in areas of
tissuedestruction in human emphysema (39) but others have failed
tofind any correlations between neutrophil numbers in
tissuesections and the severity of lung destruction (47, 49).
Table 1 summarizes interventions related to serine
proteases.Shapiro et al. (155) showed that mice lacking
neutrophilelastase were 59% protected against emphysema, strong
evi-dence for a role for neutrophil elastase (see below). In
acutesmoke exposure studies, levels of lavage neutrophils
correlated
with levels of lavage desmosine, a marker of elastin break-down,
and lavage hydroxyproline, a measure of collagenbreakdown (30, 44,
186), and administration of anti-neutrophilantibodies before smoke
exposure reduced both neutrophilsand matrix breakdown (44). In
chronic studies, inhaled (128) orinjected (30) A1AT or the
synthetic serine elastase inhibitorZD0892 (186) provided partial
protection against emphysema(Table 1). Takubo et al. (163) and
Cavarra et al. (24) showedthat pallid mice, which are naturally
deficient in A1AT, devel-oped earlier emphysema than strains with
normal A1AT levels.
While these reports support a role of the neutrophil/neutro-phil
elastase in the genesis of emphysema, they produced thesurprising
result that all interventions decreased the inflamma-tory response
and A1AT suppressed smoke-induced elevations
of TNF as well (30). There is extensive evidence fromalveolar
epithelial cell and alveolar macrophage cultures aswell as whole
mouse and human lung tissue that smoke directlyevokes an
inflammatory response by activating NF-B (112,
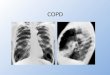
Fig. 1. Cigarette smoke-induced emphysema inguinea pigs.
Photomicrographs of lung from a con-trol (A) and a 6-mo
smoke-exposed (B) animalshowing the typical pattern of
centrilobular-like em-physema seen in small laboratory animals
chroni-cally exposed to smoke. Note that the process of airspace
enlargement primarily affects alveolar ducts.
Invited Review
L613ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
4/21
195), and possibly the aryl hydrocarbon receptor (167)
andToll-like receptor-4, at least early on (98). Recent data
alsosuggest that smoke inactivates histone deacetylases, leading
to
prolonged (and non-steroid-sensitive) inflammation (9, 74,112).
TNF production is driven by NF-B and TNF isgenerally presumed to be
a driver of inflammatory cell influx insmokers. Thus, a priori, one
would expect continuing cigarettesmoke exposure to generate a
continuing inflammatory re-sponse, and serine protease inhibitors
should provide protec-tion against emphysema without decreasing
inflammation; sup-pression of the inflammatory response thus
implies additionalanti-inflammatory mechanisms are at work (see
below).
Interference with/manipulation of metalloproteases. In thelast
15 years, there has been an increasing interest in
metallopro-teases (MMP) as mediators of emphysema. This has stemmed
inpart from the recognition that a number of metalloproteases,
including MMP-9 and MMP-12 (124), can degrade elastin; in
partfrom reports of increased levels of MMPs including MMP-1,
-2,-9, -14 (50, 72, 120, 150), and in some studies, MMP-12 (42,
60,
72, 111, 182), in BAL fluid, alveolar macrophage supernatant,
orwhole lung tissue from smokers with emphysema compared withthose
without; and in particular, from the report (63) that micewith a
targeted deletion of MMP-12 (MMP-12/) failed todevelop emphysema
after cigarette smoke exposure.
Cigarette smoke causes increased whole lung or
alveolarmacrophage levels of MMP-2, -9, -12, -13, and -14 in
mice(32) and MMP-1 in guinea pigs (151). Table 1 lists
experi-mental studies using genetically targeted mice or MMP
inhib-itors in smoke exposure models. Several broad
conclusionsstand out. First, MMP inhibition or deletion can
significantly oreven totally abrogate the development of emphysema,
indicat-ing a clear role for MMPs in this process. In fact, with
broad
Table 1. Effects of interference with proteases in chronic smoke
exposure studies
Serine Protease Inhibition or Manipulation
Report (Ref. no.) Species/Strain Intervention Inflammatory
Response% Protection Against
Emphysema Comments
Wright (186) Guinea pig ZD0892 (serine proteaseinhibitor)
Decreased 45% Decreased BAL desmosine,hydroxyproline; treatment
for last 2
mo of smoke exposure did notprotect against emphysema
Churg (30) C57Bl/6 mouse A1AT injected (prolastin) Decreased 67%
Decreased BAL desmosine,hydroxyproline; treatment roughlydoubled
serum A1AT levels andsuppressed smoke-mediatedincreases in serum
TNF
Shapiro (155) C57Bl/6 mouse Neutrophil elastaseknockout
Decreasedneutrophils,macrophages
59% MMP-12 and neutrophil elastasedegrade each others
inhibitors
Pemberton (128) Mouse A1AT inhaled (recombinanthuman A1AT)
Decreased 72% Highest dose increased BAL PMN andgave less
protection againstemphysema
Takubo (163) Pallid mouse A1AT deficient Only CD4
cellssignificantlyincreased in tissue
Early onsetemphysema
Pan-lobular emphysema
Cavarra (24) Pallid mouse A1AT deficient Not reported Early
onset
emphysema
Pan-lobular emphysema
Metalloprotease and Cysteine Protease Inhibition or
Manipulation
Report (Ref. no.) Species/Strain Intervention Inflammatory
Response% Protection Against
Emphysema Comments
Hautamaki (63)Shapiro (155)
129 mouseC57Bl/6 mouse
MMP-12 knockout Decreasedmacrophages butnot BALneutrophils
100% MMP-12 degrades A1AT andneutrophil elastase
degradesTIMP-1
Mahadeva (99) Mouse MMP-9 knockout Not reported 0%Pemberton
(127) Mouse Broad spectrum MMP
inhibitor GM6001Decreased 90% 90% average value for 2
highest
dosesMartin (105) Mouse Broad spectrum MMP
inhibitor RS113456Not reported 100% Started after 3 mo of smoke
exposure;
no increase in mean air space sizebetween 3 and 6 mo
Martin (105) Mouse Broad spectrum MMPinhibitor RS132908
Not reported 75%
Selman (152) Guinea pig Broad spectrum MMPinhibitor CP471474
Decreased 30% Measured as alveolar area at 4 mo;decreased MMP-9
levels at 4 mo
Churg (34) Guinea pig MMP-9/-12 inhibitorAZ11557272
Decreased 68% Decreased BAL desmosine; strongcorrelation BAL
desmosine withmean air space size; inhibitor alsoprevents small
airway remodeling
Kang (78) Mouse (C57) IL-18R knockout Decreased 51% Decreased
apoptosis; decreasedcytokines; decreased cathepsin B, S
All studies use 6 mo of smoke exposure, except as noted.
Invited Review
L614 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
5/21
spectrum MMP inhibitors, it is sometimes possible to
achievegreater protection against emphysema than with serine
elastaseinhibitors.
Second, the choice of which MMP to target is crucial.
Micelacking MMP-12 are completely protected against emphysema(63,
155), whereas mice lacking MMP-9 show no protection atall (99).
This latter observation is of particular interest, since it
has been suggested from studies of cultured alveolar
humanmacrophages that MMP-9 is the major mediator of emphysemain
humans (143, 144), and some have denied a role forMMP-12 in humans
(72). Selman et al. (152) found littleprotection with a broad
spectrum MMP inhibitor in guineapigs, but we (34) found 70%
protection in guinea pigs givena combined MMP-9/-12 inhibitor,
AZ11557272, indicatingthat MMPs are not just confined to murine
emphysema models,and lending support to the idea that one or both
of these MMPsmay be central players in humans. Animals exposed to
smokeand AZ11557272 had 70% protection against decreases inairflow
compared with animals exposed to smoke alone, thusshowing that
prevention of anatomic changes in animal modelsconfers a
corresponding physiological benefit.
However, as is true of serine proteases, the exact role ofMMPs
in the pathogenesis of emphysema is unclear, particu-larly since
neutrophils/serine proteases and MMPS appear tointeract, and MMP
inhibition reduces smoke-induced neutro-phil and macrophage influx
(29, 34, 128, 155). In an acutesmoke model using MMP-12/ mice, we
(29) showed thatMMP-12 was required for smoke-induced neutrophil
influxand neutrophils were required for matrix breakdown.
Thislatter idea was also supported by the finding that the
broadspectrum MMP inhibitor RS113456 acutely inhibited neutro-phil
influx and matrix breakdown (44). But in a 6-mo smokemodel, Shapiro
et al. (155) found that MMP-12/ micedeveloped a BAL neutrophilia
comparable to that of wild-
type animals, implying that proinflammatory mechanismschange
over time; we have seen a similar late neutrophilrecruitment effect
in animals treated with A1AT and in TNFreceptor (p55 p75/) knockout
mice (30, 32). It is of interestin this regard that Stevenson et
al. (161), using gene microar-rays, recently reported that gene
expression patterns in ratschange from an early proinflammatory to
a later pattern ofenhanced acquired immunity, although these data
must beviewed with great caution, since rats are extremely
susceptibleto (and the Stevenson paper indeed found) the
phenomenonknown as particle overload (reviewed in Ref. 119), and
particleoverload leads to prolonged generation of inflammatory,
fibro-genic, and probably mutagenic mediators by alveolar
macro-phages.
Additional evidence for interactions of neutrophils
andmacrophages comes from Shapiro et al. (155) who showedthat
neutrophil elastase activates pro-MMP-12 and destroystissue
inhibitor of metalloprotease (TIMP)-1; conversely,MMP-12 degrades
A1AT. Thus neutrophil elastase andMMP-12 cooperate to increase each
others proteolyticpotential.
There appear to be a variety of mechanisms by which serineand
metalloprotease inhibitors exert anti-inflammatory effects.We (186)
found that the serine elastase inhibitor ZD0892inhibited acute
smoke-mediated increases in gene expressionof the
neutrophil/macrophage chemoattractants MIP-2 andMCP-1 in mice. We
also observed that alveolar macrophages
from MMP-12/ mice did not increase TNF secretion aftersmoke
exposure, apparently because MMP-12 functions as aform of TNF
converting enzyme that liberates active TNF,(Figs. 2 and 3) and
lack of TNF appeared to be one reason fora lack of neutrophil
infiltration in the lungs of MMP-12/animals (31). As well, A1AT
suppresses smoke-mediated in-creases in TNF release by inhibiting
the serine proteases
thrombin and plasmin that leak into the air spaces after
smokeexposure (31, 35); thrombin and plasmin in turn
activateproteinase-activated receptor-1 (PAR-1), thereby
causingMMP-12 and TNF release (32, 133) (Fig. 2). This whole setof
observations would remove MMP-12 from a matrix destruc-tive role to
a signaling role in the context of cigarette smokeexposure, and
signaling roles are now recognized as majorfunctions of MMPs
(125).
An alternate explanation for decreases in inflammatorycell
influx is that inhibition of matrix destruction by eitherneutrophil
or macrophage-derived proteases decreases thegeneration of
chemotactic matrix fragments (1, 69, 129,153, 154). Houghton et al.
(68) observed that the lavagefluid of wild-type mice contained
elastin fragments thatwere chemoattractants for monocytes and that
this chemoat-tractant activity was absent from the lavage of
MMP-12/mice. They suggested that matrix breakdown in emphysemais
driven by both neutrophil elastase and MMP-12, with thelatter the
major player because of the relatively large num-bers of monocytes
that migrate into the lung after smokeexposure. In practice, MMP-12
may well play both a sig-naling and a direct matrix destructive
role.
More recently, Maeno et al. (97) have proposed that thiswhole
process of smoke-driven matrix destruction is initiatedby CD8
lymphocyte-mediated production of IFN/IP-10,with resulting
neutrophil and macrophage infiltration and in-
Fig. 2. Postulated mechanisms of cigarette smoke-induced MMP-12
release.Smoke directly activates Toll-like receptor-4 (TLR4) and
probably other (asyet unidentified) cell surface receptors that
drive MMP-12 release. In addition,smoke causes leakage of plasma
proteins into the alveolar spaces. Theseinclude prothrombin and
plasminogen, which are converted to thrombin andplasmin by alveolar
macrophage-derived tissue factor (TF) and plasminogenactivator
(PA); thrombin and plasmin in turn activate
proteinase-activatedreceptor-1 (PAR-1), which causes MMP-12
release. IP-10 derived fromsmoke-evoked T cells also causes
increased macrophage production ofMMP-12 (see Fig. 3). Modeled from
Refs. 35, 97, 98, 133.
Invited Review
L615ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
6/21
creases in MMP-12 production (Fig. 3). Of note, these data
aresimilar to findings reported in human lungs with emphysemaby
Grumelli et al. (60).
Cantor et al. (20, 21) showed that inhaled hyaluronan (HA)can
bind to elastin fibers in vivo and in vitro and preventselastolysis
by elastases. HA reduced air space enlargementcaused by porcine
pancreatic elastase (20), provided 100%
protection against emphysema in smoke-exposed DBA/2Jmice, and
reduced smoke-mediated increases in lavage desmo-sine. These
findings support matrix attack as a fundamentalfeature of
smoke-induced emphysema.
Cysteine proteases. Kang et al. (78) showed that smokeinduces
production of the cysteine proteases cathepsins B andS in mice via
a mechanism involving IL-18, and, probably,interferon-. These
proteases can degrade matrix componentsand thus might play a role
in emphysema. The same reportfound greater levels of cathepsins B
and S in alveolar macro-phages from cigarette smokers compared with
controls. IL-18R/ mice were significantly protected against
emphy-sema and had decreases in smoke-induced lavage neutrophilsand
macrophages as well as a variety of chemokines andcathepsins, along
with decreased levels of MMP-12. Thesedata thus present the same
mix of possible effectors seen withstudies inhibiting/deleting
serine and metalloproteases.
Anti-inflammatory interventions. If the
protease-antiproteasehypothesis (i.e., that the smoke-driven
inflammatory response
leads to proteolytic matrix destruction) is correct, one
couldinhibit smoke-mediated inflammatory responses instead of
pro-teases, and a number of studies have directly or
indirectlytargeted the production and/or signaling of
proinflammatorymolecules (Table 2).
TNF is consistently elevated in human smokers (10, 165).In mice,
knockout of TNF receptors 1 and 2 greatly reduced
inflammatory infiltrates, emphysema, levels of some MMPs,and
gene expression of proinflammatory cytokines (28, 32, 46)after
smoke exposure; TNF receptor 2 appeared to be themajor mediator of
the inflammatory response (46). Conversely,inducible overexpression
of TNF produced an increase inneutrophils, parenchymal B cell
nodules, MMP-12, cathepsinK, and emphysema in the absence of smoke
(176). There wasan extremely strong correlation (R 0.89, P
0.0001)between serum TNF levels and air space size after 6 mo
ofsmoke exposure in guinea pigs (34). While these reports implyan
important role for TNF, two studies in humans usingTNF antagonists
failed to show a benefit (10, 134, 172),suggesting that translation
of anti-inflammatory therapies from
animal models to humans is not straightforward.Using a different
anti-inflammatory approach, Thatcher et al.(166) prevented
neutrophil influx by administration of SCH-N,a CXCR2 inhibitor, to
mice, with an 50% reduction inparenchymal cells after a 3-day smoke
exposure. SCH-N didnot decrease smoke-mediated increases in TNF,
IL-6, orPGE2, and levels of the neutrophil chemoattractant
CXCR2ligands KC and MIP-2 were considerably elevated. Maes et
al.(98) showed that, in mice, Toll-like receptor-4 plays a role
inthe early but not the chronic inflammatory response to
cigarettesmoke.
Phosphodiesterase-4 degrades the anti-inflammatory nucle-otide
cyclic 35-adenosine monophosphate. In C57Bl/6 miceexposed to smoke
for 7 mo, Roflumilast, a phosphodiesterase-4
inhibitor, provided 100% protection against
smoke-inducedincreases in air space size, reversed smoke-induced
loss of lungdesmosine, reduced neutrophil and especially macrophage
in-flux, and more than doubled levels of the
anti-inflammatorycytokine, IL-10 (106). One of the effects of
Roflumilast is todecrease macrophage production of TNF, and in a
humantrial, 4 wk of Roflumilast decreased blood/sputum TNF,
IL-8,and neutrophil elastase, and improved FEV1 (58).
IFN appears to drive many proinflammatory cytokines viaCCR5
(92). IFN/ or CCR5/ mice exposed to smokefor 6 mo were completely
protected against emphysema andshowed decreased inflammatory cells,
apoptosis, levels ofMIP-1, MIP-1, and RANTES (16, 92). Protection
may have
been mediated through decreased MMP-12, since transgenicmice
overexpressing IFN increased MMP-12 production (92).Administration
of the 3-hydroxy-3-methyl-glutaryl-coen-
zyme-A reductase inhibitor, simvastatin (87), to rats
com-pletely abolished smoke-induced emphysema and
preventedperibronchial and perivascular accumulation of
lymphocytes.This report is difficult to interpret: the authors
reported aremarkable degree of air space enlargement (80% increase
inmean air space size) after a relatively short exposure period
(16wk). Furthermore, statins have been reported to increase
mac-rophage production of MMP-12 (4), which should make em-physema
worse. However, there are reports of beneficial ef-fects of statins
in patients with COPD (102, 159) and reports
Fig. 3. Postulated mechanisms of matrix attack and emphysema. In
this model,smoke causes release of a variety of chemokines from
structural cells andalveolar macrophages, leading to an influx of
neutrophils (PMN), macro-phages, lymphocytes, and dendritic cells.
MMP-12 causes conversion ofpro-TNF to active TNF, thereby
amplifying the inflammatory process.Neutrophil elastase and
macrophage-derived MMP-12, as well as other pro-
teases, can directly attack the alveolar wall, leading to both
matrix destructionand liberation of elastin fragments, which are
chemoattractants for macro-phages, further amplifying the
inflammatory response. The scheme shown hereencompasses the
findings of Lee et al. (94) that COPD patients have anti-elastin
antibodies, so that protease-derived elastin fragments evoke an
auto-immune response, leading to further matrix attack and
production of IFN/IP-10, which itself increases MMP-12 release by
macrophages and also increasesmatrix destruction. Other matrix
fragments or smoke-modified non-matrixproteins might function in a
similar fashion. In addition, neutrophil elastasedegrades TIMPs and
MMP-12 degrades 1-antitrypsin (A1AT), thus removinganti-proteolytic
defenses, and oxidants in the smoke may inhibit A1AT andother
antiproteases such as SLPI as well. This scheme thus incorporates
aninnate immune response to smoke, a protease feedback
amplification systemdriven by matrix destruction, and an acquired
immune response, also driven bymatrix destruction. The relative
importance of each process probably changesover time. Modeled from
Refs. 31, 38, 88, 92, 97, 155.
Invited Review
L616 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
7/21
that statins enhance clearance of apoptotic cells (114), so
thistype of compound may be worth further investigation.
In aggregate, both interventions directed against
proteases(serine, cysteine, or metalloproteases) and interventions
di-rected against the development of an inflammatory influxprovide
significant protection against emphysema in animalmodels and thus
support the protease-antiprotease hypothesis,although the
mechanism(s) involved are complex and notentirely clear.
Effects of Smoke on Different Mouse Strains: Evidence for
aGenetic Propensity to Emphysema
As noted above, susceptibility to COPD likely results
frommultiple genetic and environmental effects. Cavarra et al.
(24)showed that mouse strains with differing
antioxidant/antipro-tease capacity reacted differently to cigarette
smoke (see Mech-anisms Related to Oxidative Stress in Emphysema).
Guerassi-mov et al. (61) exposed five different strains of mice to
smokein an attempt to define genetically susceptible and
resistantvarieties. The strains were chosen based on differences in
theMHC haplotype, a major determinant of the inflammatoryresponse
in mice. After 6 mo of smoking, NZWLac/J mice had
no increase in mean air space size (Lm), whereas AJ,
SJL,C57BL/6, and AKR mice had 17.9%, 23.8%, 13.2%, and
38%increases, respectively. Because, as noted by Henson
andVandivier (64), it is remarkably easy to induce alveolar
en-largement with a very wide variety of manipulations in themouse,
Guerassimov et al. (61) defined emphysema as anincrease in Lm along
with an increase in lung compliance(Fig. 4). By these criteria,
only the AKR strain had significantemphysema, whereas A/J, SJL, and
C57BL6 strains appeared
to be mildly susceptible to smoke and NZW were resistant.Hoshino
and Cosio (unpublished data) then investigated the
genetic response to cigarette smoking in resistant (NZW)
andsusceptible (AKR) strains utilizing expression microarrays.There
were striking constitutive differences between the twostrains,
mainly in the expression of genes that encode forproteins with
immune function. The NZW mice had higherconstitutive expression of
genes that inhibit differentiation andproliferation of T and B
cells and protect against apoptosis andT cell activation (Ifi203,
CD72, C4, Klra-1, 8, and 13), alongwith higher levels of several
antioxidant genes that were notprominently expressed in AKR mice.
In contrast, the constitu-tive inflammatory genes expressed in AKR
mice were proin-
Table 2. Effects of manipulation of the immune/inflammatory
response in chronic smoke exposure studies
Report (Ref. no.) Species/Strain Intervention Inflammatory
Response% Protection Against
Emphysema Comments
Churg (32) Mouse (C57Bl/6) TNF R1 R2/ Decreased BALmacrophages,
neutrophils
71% Increased MMP-2, -9, -12, -13, -14with smoke exposure;
knockoutmice: decreased MMP-12, -13,-14; decreased gene
expression
of TNF, MCP-1, MIP-2Dhulst (46) Mouse (C57) TNF R1/ Not affected
NoneDhulst (46) Mouse (C57) TNF R2/ Decreased BAL neutrophils,
macrophages,lymphocytes
100% Decreased CD4 and CD8 cells
Martorana (106) Mouse (C57) PDE4 inhibitorRoflumilast
Decreased 100% 7-mo smoke exposure; increasedIL-10
Lee (87) Rat (Sprague- Dawley) Simvastatin Decreased
peribronchiallymphoid aggregates
100% 4-mo smoke exposure; alsodecreased pulmonary
arterypressure; possible technicalproblems with study
Cantor (21) Mouse (DBA/2) Hyaluron aerosol Not reported 100%
Putative protection of elastin fibers;possible developmental
problems
Bracke (13) Mouse (C57Bl/6) CCR6/ Decreased 67% No protection
against small airwayremodeling; decreased MMP-12,BAL PMN, T
lymphocytes,macrophages
Ma (92) Mouse (C57Bl/6) IFN/ Not reported 100% Decreased
apoptosis; MMP-12probably involved
Ma (92) Mouse (C57Bl/6) CCR5/ Decreased (primarilydecreased
macrophages)
100% Decreased cytokines, apoptosis
Bracke (16) Mouse (C58Bl/6) CCR5/ Decreased 25% Decreased PMN,
macrophages,dendritic cells, lymphocytes; noprotection against
small airwayremodeling
Maeno (97) Mouse (C57Bl/6) CD8/ Decreased 100% Some persisting
elevation in BALPMN
Maeno (97) Mouse (C57Bl/6) CD4/ Not decreased NoneDHulst (45)
SCID mouse No acquired immune
responseIncreased neutrophils,
macrophages, dendriticcells
None No T or B lymphocytes; nolymphoid follicles
Maes (98) Mouse (C3H/HeJ) TLR4/ Increased BAL PMNmacrophages at
26 wk
None Decreased BAL lymphocytes anddendritic cells at 26 wk
van der Deen (172) Mouse (FVB) Multi-drug resistanceprotein/
Decreased inflammation andinflammatory mediators No emphysema
produced in wildtype or knockout
All studies use 6 mo of smoke exposure, except as noted.
Invited Review
L617ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
8/21
flammatory, and several were related to adaptive immunity.
With smoke exposure there was a striking difference in
generesponse: the NZW strain demonstrated decreased expressionof 77
of the 82 genes that were significantly changed inresponse to
smoke, and 44% of the genes attenuated by smokeexposure influence
immune and/or inflammatory processesincluding immunoglobulins,
complement, chemokines, cyto-kines, and other proinflammatory
factors that enhance thefunctions of neutrophils, macrophages, and
T and B cells.NZW mice also upregulated antioxidant genes. In
contrast,AKR mice showed increased expression of 52 of the 57
genesthat were changed by smoke exposure, and 25% of the
genesincreased after smoking had functions related to the
immuneresponse; 13% were proapoptotic (Fig. 5).
In another study aimed at investigating the potentiation of
inflammation by cigarette smoke, Reynolds et al. (137)
inves-tigated the expression of Egr-1 (early growth response gene
1)in NZW and AKR mice. Egr-1 gene induces IL-1 and TNF,cytokines
that contribute to the recruitment of inflammatorycells after smoke
exposure. Egr-1 expression was marginallydetected by
immunochemistry in the lungs of nonsmokingmice, but increased
markedly in the susceptible AKR strainafter smoke exposure.
However, Egr-1 was only minimallyinduced in the lungs of the
resistant NZW.
These studies suggest that resistant animals do not increaseor
actively decrease the proinflammatory response to smokewhile
increasing the antioxidant response, thus preventingmatrix
breakdown and a potential acquired immune response to
matrix fragments. In contrast, inflammation in the
susceptible
strain is progressive, probably due to ongoing stimulation
ofinnate and adaptive immunity by the expression of genesinvolved
in antigen presentation and T and B cell activation.These findings
support the possibility of an autoimmune pro-cess triggered by
smoke exposure as a factor important in themaintenance of the
inflammation and the development ofemphysema.
Evidence for Acquired Immunity in Animal Modelsof Smoke-Induced
Emphysema
An abnormal inflammatory response is an important com-ponent in
the definition of COPD (131). Besides neutrophilsand alveolar
macrophages, both CD4 (T helper) and CD8
(cytotoxic) T cells are increased in the airways and
lungparenchyma of patients with COPD with a predominance ofCD8 T
cells, and Finkelstein et al. (49) showed that in humanlungs, there
is a correlation between the number of T lympho-cytes/mm3 of lungs
and the extent of emphysema. This infil-tration with T cells, seen
in smokers who develop COPD, butnot in normal smokers, represents
an activation of the adaptiveimmunity that presumably follows from
the initial and thensustained innate immune response characterized
by increasednumbers of macrophages and neutrophils.
The T cells found in patients with COPD are fully activated(60,
146), expressing a large array of Th1 chemokines andcytokines. This
inflammatory process most likely also involves
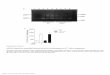
Fig. 4. Pressure volume curves from control and cigarette
smoke-exposed during 6 mo NZW, C57Bl/6, Pallid, and AKR/J mice. The
animals were anesthetizedand paralyzed, and lung mechanics were
measured at PEEP levels of 19 cmH2O by delivering a broad-band
volume perturbation to the lungs for a period of16 s, and the fast
Fourier transforms of the data windows were used to calculate the
input impedance of the lung. The parameter Htis is equal to lung
elastance(1/compliance) at a frequency of 1 rad/s 0.16 Hz, and we
used Htis vs. PEEP to calculate an equivalent pressure-volume curve
for the lungs between1 and 9 cmH2O. Only the Pallid and AKR/J show
a significant compliance increase. [From Guerassimov et al.
(61).]
Invited Review
L618 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
9/21
the migration of dendritic cells, since it has been
recentlyreported that T cells in humans with emphysema are
beingpresented with antigens derived from the breakdown of
elastin(88). These findings support a role for autoimmunity in
thepathogenesis of COPD.
Guerassimov et al. (61) investigated the inflammatory re-sponse
to long-term cigarette smoking in the mice with differ-ent
susceptibilities to emphysema described in EFFECTS OFSMOKE ON
DIFFERENT MOUSE STRAINS: EVIDENCE FOR A GENETICPROPENSITY TO
EMPHYSEMA. To approximate human studies, theyused morphometric
methods to quantitate the percentage ofimmunostained inflammatory
cells in the alveolar walls. After6 mo of smoking, only the
susceptible AKR strain had a floridcellular inflammatory infiltrate
comprising CD4, CD8, and
T cells, along with macrophages and neutrophils. Neitherthe
resistant nor the mildly susceptible strains exhibited
aninflammatory infiltrate containing T cells other than some cells
in the NZW strain (Fig. 6). Similar results were found byTakubo et
al. (163) in the Pallid mouse, a strain that alsodevelops marked
emphysema after 6 mo of smoking exposure.Analysis of inflammatory
chemokines and cytokines in thelungs (Fig. 7) in the three groups
with different susceptibilitiesconfirmed that a true Th1
inflammatory response, secondary toT cell activation, had developed
in the susceptible AKR micebut was not present in the resistant or
mildly susceptiblestrains. These findings are of interest since
they mimic the inflam-matory changes that smokers develop, and, as
in the human
smokers, only the susceptible animals develop the adaptive
im-mune reaction seen in humans.
The importance of CD8 T cell inflammation in the devel-opment of
cigarette-induced emphysema in mice has beenrecently demonstrated
by Maeno et al. (97). In contrast towild-type mice that showed
macrophage, neutrophil, and lym-phocyte inflammation and emphysema
after 6 mo of exposure,
CD8/ mice had a blunted inflammatory response and didnot develop
emphysema when exposed to long-term cigarettesmoke. They showed
that the CD8 T cell product IFN-inducible protein-10 induces
production of MMP-12 that de-grades elastin, causing lung
destruction and generating elastinfragments that attract more
macrophages and probably act asan antigenic stimulus to maintain
the adaptive immune reactionbringing more T cells into the
lung.
It should be noted that some of the literature in this area
iscontradictory. SCID mice, which completely lack B and Tcells,
develop emphysema of a degree comparable to that of thewild type
(Balb/C), along with an inflammatory influx ofneutrophils,
macrophages, and dendritic cells, and increasedlevels of MMP-12
(45). However, some workers have foundthat SCID mice develop
hemorrhage after smoke exposure,making emphysema difficult to
measure (Shapiro SD, personalcommunication). As well, reported
laboratory to laboratoryvariations in response to a given mouse
strain seem to translateinto different molecular responses. For
example, two of theauthors of this review (Churg and Wright) have
consistentlyfound an 3540% increase in air space size and a
smoke-induced elevation in gene and protein expression of TNF
andother proinflammatory mediators in C57Bl/6 mice (28, 30), butthe
third author of this paper (Cosio) finds much smallerincreases in
air space size and has not observed such inflam-matory responses in
C57Bl/6 mice. The mice used by Maenoet al. (97), which developed a
CD8 T cell response (see
above), were also C57Bl/6 based. It is unclear if these
findingsindicate the presence of small genetic differences in the
samemouse strain obtained from different sources, or mean
thatdifferent smoke exposure conditions and smoke exposure sys-tems
can force animals of the same notional strain into the typeof
proinflammatory and adaptive immune response just de-scribed.
Mechanisms Related to Oxidative Stress in Emphysema
Cigarette smoke is an extremely concentrated source ofreactive
oxygen species (ROS) and reactive nitrogen species(130). The
inflammatory response to smoke potentially aug-ments oxidative
stress, since neutrophils and macrophages
release ROS, and those from smokers release even greateramounts
(17, 66, 91, 95, 96).
There is considerable biochemical evidence of oxidativestress in
cigarette smokers and greater levels in those withCOPD (reviewed in
Refs. 14, 95, 96). These changes includeincreased exhaled H2O2 and
8-isoprostane, decreased plasmaantioxidants, and increased plasma
and tissue levels of oxi-dized proteins, various lipid peroxidation
products such as4-hydroxynonenal, as well as protein tyrosine
residues/3-nitro-tyrosine, indicators of attack by reactive
nitrogen species.Antioxidant enzyme levels are also altered.
Oxidative attackmay inactivate antiproteases such as SLPI (25), and
oxidativedamage is believed to decrease production of/inactivate
some
Fig. 5. Overall differences in gene expression profiles in mice
resistant, NZW,and susceptible, AKR, to the development of
emphysema after 6 mo ofcigarette smoke exposure. Gene expression
profiles were investigated in 3animals per control and exposed
groups using Affymetrix Microarray Analy-sis. Compared with their
controls, NZW mice largely downregulated geneexpression, whereas
AKR mice showed the opposite pattern.
Invited Review
L619ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
10/21
histone deacetylases (9, 74), leading to a prolonged
inflamma-tory response.
Evidence of oxidant attack is also seen in animal models.Aoshiba
et al. (2) showed that even a 1-h exposure of mice tocigarette
smoke resulted in immunochemically detectable4-hydroxynonenal and
8-hydroxy-2-deoxyguanosine, a markerof oxidative DNA damage, in the
airway and alveolar epithelialcells. We (27) found that a 10-min
smoke exposure producedlipid peroxidation in rat tracheal explants.
Rangasamy et al.
(132), using microarrays, showed upregulation of a variety
ofantioxidant enzymes after smoke exposure of ICR mice.McCusker and
Hoidal (109) found increases in superoxidedismutase and catalase
expression in alveolar macrophagesfrom smoke-exposed hamsters,
whereas Cavarra et al. (25)showed depletion of protein thiols,
ascorbic acid, and glutathi-one, all antioxidant substances, in
smoke-exposed C57Bl/6mice.
Interference with the antioxidant response by deletion ofNrf2
(Nrf2/) (71, 132), a redox-sensitive transcriptionfactor that
upregulates a variety of detoxification and antioxi-dant genes,
caused increased smoke-induced inflammation andearlier and more
severe emphysema than in wild-type (ICR orICR/Balb/c) mice, and
Nrf2/ mice had impaired upregula-
tion of antioxidant enzymes (157). As well, Nrf2/ micewere
deficient in A1AT and secretory leukoprotease inhibitor(SLPI).
These findings link a lack of antioxidant protection tothe
protease-antiprotease hypothesis (Fig. 3).
Conversely, several investigators have acutely
increasedantioxidant protection (Table 3) with consequent decreases
ininflammatory cell influx, protein oxidation, inflammatory
me-diators, and airway squamous metaplasia after short-termsmoke
exposure (5, 116, 158). Rubio et al. (142) found that
theantioxidant N-acetyl cysteine ameliorated elastase-induced
em-physema in rats.
Cavarra et al. (24) reported that ICR mice, which
increasedantioxidant levels in BAL after smoke exposure, were
pro-
tected against emphysema, whereas C57Bl/6 and DBA/2,which did
not increase antioxidant levels, developed emphy-sema; of note,
levels of antioxidant protection appeared to bemore important than
BAL levels of elastase inhibitory capacityin determining the
severity of emphysema. Foronjy et al. (52)showed that chronically
smoke-exposed transgenic CuZnSODanimals had much smaller increases
in neutrophil and macro-phage infiltration than did wild-type
animals, did not showincreases in production of a variety of MMPs
including MMP-
12, were protected against generation of lipid
peroxidationproducts, and were 100% protected against the
development ofemphysema. There was substantial protection against
elastase-induced emphysema as well.
These studies thus support a role for oxidant attack in
thegenesis of smoke-induced emphysema, but it is noteworthythat the
immediate mechanism of attack appears to be throughincreases in
inflammatory cells, and, probably, decreases inantiprotease
protection, thus linking oxidant attack to theprotease-antiprotease
hypothesis.
Mechanisms Related to Repair of Alveolar Structurein
Emphysema
Retinoids accumulate in the lung during embryogenesis andare
essential to alveolar septation; mice lacking retinoid recep-tors
have lower levels of elastin and enlarged alveolar spaces(108).
Massaro and Massaro (107) reported that treatment withall-trans
retinoic acid reversed the emphysematous changesseen in rats after
instillation of elastase, and this was confirmedin rats by Belloni
et al. (12). However, reports in otherspecies and with cigarette
smoke have been discouraging.March et al. (103) did not find any
protective effects ofretinoids against smoke-induced emphysema in
either A/J orB6C3F1 mice, and Meshi et al. (110) did not observe
anybenefit in smoke-exposed guinea pigs. A human trial ofretinoid
therapy did not show any clear improvement in
Fig. 6. Inflammation in the alveolar wall inmouse strains with
different susceptibilitiesto the development of emphysema after 6
moof cigarette smoke exposure. In these exper-iments, only the
susceptible AKR miceshowed a full inflammatory profile. PMN,
neu-trophil; AM, alveolar macrophage; CD4,CD4 T cell; CD8, CD8 T
cell. [FromGuerassimov et al. (61).]
Invited Review
L620 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
11/21
emphysema (141). But this issue still needs further
investi-gation because the dose of retinoic acid may be
crucial.Stinchcombe and Maden (162) have recently observed
thatretinoic acid will restore alveolar architecture that is
dis-rupted by steroid treatment of newborn mice if a highenough
dose is used.
Mechanisms Related to Failure to Repair/Failure of Lung
Maintenance in Emphysema
While there appears to be overwhelming evidence thatcigarette
smoke-induced damage to the alveolar wall matrix asa result of
inflammatory cell-derived protease, and, probably,oxidant, attack
(but, again, with oxidant-driven increases in
inflammatory cells) is the driving force behind emphysema,one of
the curious features of emphysema is that the alveolarwall largely
fails to regenerate new matrix. This is in sharpcontradistinction
to the small airways and the intrapulmo-nary arteries, where the
response to smoke is a markedincrease in matrix and/or structural
cells (see Mechanisms ofSmall Airway Remodeling and Mechanisms of
VascularRemodeling and Pulmonar y Hype rtension), despite the
factthat these anatomic compartments are separated by only afew
micrometers.
A variety of theories, conveniently grouped as failure
torepair/failure of lung maintenance have been advanced toaccount
for this phenomenon (reviewed in Refs. 134, 170,171). Increases in
apoptotic cells, sometimes accompanied bydecreases in proliferating
cells, have been found in severelyemphysematous compared with
nonemphysematous humanlungs (73, 100, 170, 196). Administration of
agents that causeendothelial or epithelial cell apoptosis to
animals results inrapid air space enlargement, although, unlike
smoke-inducedemphysema, this process is also rapidly reversible and
is notaccompanied by an inflammatory response (3, 79). In
tissueculture models, smoke exposure interferes with cell
prolifera-tion, chemotaxis, and production/remodeling of matrix
com-ponents by fibroblasts (23, 134) and depresses the
productionand activity of lysyl oxidase (53, 86), an enzyme crucial
to theformation of stable insoluble elastin and collagen.
Fibroblastsfrom emphysematous human lungs proliferate more
slowly
than those from nonemphysematous lungs (67, 117) and
showincreased expression of a variety of
senescence-associatedmarkers (118, 169).
Only a few animal models have looked at this issue usingsmoke
exposure. Mice lacking senescence-associated marker30, an
anti-aging calcium binding protein, show increased
Fig. 7. Cytokine and chemokine gene expression in mouse strains
withdifferent susceptibilities to the development of emphysema. In
these experi-ments, NZW mice tend to downregulate gene expression,
C57Bl/6 mice tendnot to change expression patterns, and AKR mice
tend to upregulate expres-sion. [From Guerassimov et al. (61).]
Table 3. Effects of antioxidant manipulation in acute and
chronic smoke exposure studies
Report (Ref. no.) Species/Strain Intervention Inflammatory
Response Effect on Emphysema Comments
Nishikawa (116) Guinea pig rhSOD Decreased Acute study only
Decreased IL-8 gene expression,NF-B activation
Smith (158) Rat Catalytic antioxidant
AEOL 10150
Decreased Acute study only Decreased MIP-2, ICAM-1,
squamous metaplasia inairways
Banerjee (5) Guinea pig Black tea (antioxidant)in drinking
water
Not reported Acute study only Decreased oxidation of
lungproteins
Cavarra (24) ICR, C57, DBA/2 mice ICR mice have
increasedantioxidant protection,cf. to C57, DBA/2
Not reported 5% increase in meanair space size inICR, not
significant;C57 and DBA/2 getemphysema
Elastin content decreased in C57and DBA/2 but not in ICR
Rangasamy (32) M ouse (ICR) Nrf2 knockout Increased Earlier and
moresevere
Decreased A1AT; increasedapoptosis
Iizuka (71) Mouse (Balb/c) Nrf2 knockout Increased Earlier and
moresevere
Decreased SLPI
Foronjy (52) Mouse Tg ZnCuSOD Decreased 100% protection Protects
against smoke-mediatedincrease in lipid peroxidation
Invited Review
L621ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
12/21
emphysema when exposed to smoke (148) and also showincreased
numbers of apoptotic lung structural cells (113). Inelastase
instillation emphysema, concurrent smoke exposureinhibits new
elastin synthesis, whereas with elastase alone,there is brisk
production of new protein (121). We haveobserved, using laser
capture microdissection, that the paren-chyma initially responds to
smoke by upregulation of genes
that produce matrix components, but that over time most ofthese
genes are downregulated, suggesting that failure to repairis not a
simple yes or no proposition, but a process that changeswith
increasing exposure (Churg, unpublished data). Kanget al. (78)
showed that smoke activates IL-18 and causesapoptosis by direct or
IL-18R-mediated activation of a num-ber of caspases. IFN/ mice
exposed to smoke for 6 mohad decreased levels of inflammatory
cells, apoptosis, andemphysema (92). In Nrf2/ mice (132), there was
an in-crease in apoptotic parenchymal cells with smoke
exposure.Smoke disrupts lung VEGF signaling in rats, and
VEGFappears to be an important trophic mediator of lung
endo-thelial cell survival (79).
Tuder et al. (171) have proposed that oxidative damage
andapoptosis are the primary events in emphysema and
thatinflammation is secondary, but this idea is contradicted
byevidence that smoke directly drives inflammation, even
inmonolayer cultures where smoke causes cytokine release with-out
apoptosis (see Mechanisms Related to the Protease-Anti-protease
Hypothesis of Emphysema). Guinea pigs and mostgenetically intact
mice that develop emphysema do not developincreased apoptosis, even
with prolonged smoke exposure(Ref. 51 and Churg, unpublished data),
although Bartalesi et al.(11) reported focal areas of increased
apoptosis in DBA/2 butnot C57Bl/6 mice.
The question of whether apoptosis is a primary or secondaryevent
in emphysema (and even whether it occurs in excess) is
unresolved and is not straightforward, because apoptosis
ofepithelial cells (anoikis) may also occur as a result of
destruc-tion of the underlying matrix (55). It should also be noted
thatthere is not a total absence of repair in emphysema, sincehuman
centrilobular emphysema is associated with increasedamounts of
collagen (22). Nonetheless, the data suggest thatthere is
ineffective repair in the parenchyma after long-termsmoke
exposure.
Mechanisms of Small Airway Remodeling
Small airway remodeling (increases in bronchiolar wallfibrous
tissue, muscle, inflammatory cells, and luminal mucus)(SAR) is now
recognized as an important cause of airflow
obstruction in cigarette smokers (65, 122). There are only a
fewreports that have examined smoke-induced SAR in animals.The
changes found are subtle and easily missed on casualexamination,
and some investigators deny that SAR occurs(104). However, careful
morphometric studies from severallaboratories have confirmed that
there is indeed SAR, largelymanifest as increases in airway wall
collagen, in both guineapigs (34) and mice (15, 33, 16) after
chronic smoke exposure(Fig. 8). Wright and colleagues (192)
reported an increase inthick collagen fibers (Fig. 8), an effect
that probably increasesairway stiffness, in the small airways of
guinea pigs exposedto cigarette smoke for 6 mo. They found negative
correla-tions of the amount of thick collagen fibers with peak
expiratory flow and FEV0.1/FVC, and positive correlationswith
airway resistance, thus indicating that smoke-inducedairway
remodeling in animals is associated with abnormalphysiology.
In humans, the usual assumption has been that, since em-physema
is driven by inflammatory cells and their proteases,SAR should
follow the same pathways (77). In fact, little is
known about the pathogenesis of SAR in humans. One piece
ofevidence against a primary role of inflammation in SAR arereports
(15, 16) that mice lacking CCR6 and CCR5 (receptorsfor various
chemoattractant chemokines) had decreased in-flammation and
decreased emphysema after smoke exposure,but no decrease in SAR
compared with wild-type animals.SAR was also prevented in
smoke-exposed guinea pigs by anMMP-9/-12 inhibitor, again evidence
that mechanisms otherthan inflammation are important (34).
Using laser capture microdissection of small airways
(bron-chioles) from the lungs of smoke-exposed C57Bl/6 mice,Churg
et al. (33) found that smoke persistently upregulatedgene
expression of type I procollagen and profibrotic cyto-kines,
particularly those related to TGF- signaling. With asingle acute
exposure, elevations in gene expression were seenwithin 2 h of
starting smoke exposure and mostly decreasedover 24 h, as opposed
to numbers of lavage inflammatory cellsthat increased slowly over
24 h (44), implying that, at least inthe short term, upregulation
of the fibrotic response is inde-pendent of inflammation.
In rat tracheal explants, an airway model system that is freeof
smoke-evoked inflammatory cells, a very brief (15-min)exposure to
smoke resulted in increases in the same set ofgenes mentioned
above, along with collagen (hydroxyproline)by 24 h, and these
increases could be prevented with aninhibitor of TGF- receptor 1
(Churg, unpublished observa-tions) or a TGF- competitor, fetuin
(179). The smoke-ex-
posed explants released increased amounts of TGF-1 via
anoxidant-driven mechanism (Fig. 9).In short, the animal models
suggest that small airway re-
modeling is driven by direct smoke induction of fibrogenicgrowth
factors, and the role of inflammatory cells is uncertain;rather,
the initial driving force appears to be oxidant-mediatedactivation
of TGF-. However, since neutrophils and macro-phages release
oxidants in response to smoke (17, 66, 91, 95,96), such cells might
potentiate TGF- release and hencepotentiate small airway remodeling
(Fig. 9).
Both B and T lymphocytes are increased in the small airwaywalls
in human COPD (37, 65, 145, 173). Whether they play adirect role in
the pathogenesis of SAR or reflect colonization ofthe small airways
by infectious agents in patients with ad-
vanced COPD is unclear (65). In cigarette smoke-exposedanimals,
peribronchial lymphoid aggregates increase in num-ber (Fig. 10),
but in CCR6/ and CCR5/ mice, a lack ofperibronchial lymphoid
aggregates with chronic smoke expo-sure did not protect against SAR
(15, 16).
Mechanisms of Vascular Remodeling and PulmonaryHypertension
Pulmonary hypertension (PHT) is a relatively common formof
cigarette smoke-induced lung disease and develops in 6%of subjects
with COPD, but is present in 40% of patients withan FEV1 of less
than 1 l (6, 19, 93, 94, 180). PHT is animportant complication
since it is a significant predictor of
Invited Review
L622 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
13/21
mortality, and is a major cause of morbidity, in patients
withCOPD (36, 168, 189).
The mechanism(s) of PHT in smokers is not known. Althoughit is
often stated that PHT arises as a result of loss of vascular
bedsecondary to emphysematous lung destruction and/or
hypoxicvasoconstriction, recent data indicate that this is not true
(re-viewed in Ref. 75). Guinea pigs exposed to cigarette
smokedevelop an 25% increase in mean pulmonary artery
pressure(191). Using plastic vascular casts in such animals, we
showedthat PHT is not associated with significant alveolar
capillary
destruction (193, 194). Likewise, although there is a
correlationbetween PHT and PO2, PO2 has not been found to be an
indepen-dent predictor of pulmonary arterial pressure (149).
In guinea pigs, smoke-increased arterial muscularization(Fig.
11) correlates with pulmonary arterial pressure, andinterestingly,
also correlates with increased mRNA and proteinlevels of the
vasoactive mediators endothelin and VEGF inthese vessels (198).
Smoke-induced vascular remodeling alsoappears to be related to
matrix reorganization, since in guineapigs, increases in pulmonary
arterial pressure and vascular
Fig. 8. Small airway remodeling in guineapigs. Photomicrographs
of Picrosirius Red-stained small airways (membranous bronchi-oles)
from control (A and B) and 6-mo smoke-exposed (C and D) guinea
pigs. Note thedistinct increase in airway wall collagen.
B and D are taken with polarized light; thebright white
birefringence indicates that this isthick fiber collagen, and
increases in thick fibercollagen cause airway wall stiffness,
leading toincreased resistance to airflow (see Ref. 192).
Fig. 9. Postulated mechanisms of small airway remodel-ing. In
this model, oxidants in cigarette smoke, and perhapsoxidants
released by smoke-evoked inflammatory cells,cause activation of
latent TGF- on the airway epithelialcell surface and also TGF-
bound to matrix, leading toTGF- signaling through Smads and
connective tissuegrowth factor (CTGF), and increased collagen
production.In addition, oxidant attack may activate MMP-9, which
inturn can activate latent TGF-. Modeled from Refs. 33,34, 179.
Invited Review
L623ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
14/21
muscularization could be reduced by administration of theserine
elastase inhibitor ZD0892 (188), and in smoke-exposedmice, there
was an early TNF-dependent upregulation ofMMP-2, -9, -12, and -13
in the small intrapulmonary arteries,
with only MMP-12 persisting over a 6-mo period (190).However, an
MMP-9/-12 inhibitor did not prevent PHT inguinea pigs (34).
In rats (164), simvastatin reduced pulmonary arterial pres-sure,
perhaps through suppressing inflammation and MMP-9induction, or
perhaps, as in other models of pulmonary hyper-tension (56),
through mechanisms involving increased apopto-
sis or the Rho-kinase pathway. Figure 12 shows some of
thefactors that appear to drive PHT in COPD.
Mechanisms of Goblet Cell Metaplasia, MucusHypersecretion, and
Exacerbations
In humans, goblet cell metaplasia is a frequent finding in
thelarge and small airways of cigarette smokers (115, 183), as
areincreases in the number and size of mucus-producing glands inthe
large airways. Indeed, the amount of intraluminal mucus inthe small
airways has been shown to represent a major differ-ence between
subjects with significant [GOLD Stage 3 and 4(48)] COPD compared
with subjects with a lesser degree ofairflow obstruction [GOLD
Stage 1 and 2 (48)] (65), and theexcess mucus production that
defines chronic bronchitis is alsothought to be important in the
pathogenesis of acute exacerba-tions of COPD (reviewed in Ref.
13).
As opposed to humans, bronchial glands are concentratedin the
proximal trachea in rats and mice, whereas in guineapigs, they are
more diffusely distributed but relatively sparsebeyond the proximal
trachea (57, 181). This anatomic distri-
bution means that it is difficult to produce a model of
smoke-induced chronic bronchitis in these animals, and for
thisreason, the literature tends to emphasize instead goblet
cellmetaplasia. However, goblet cell metaplasia, mucin produc-tion,
mucin secretion, and bronchial gland hypertrophy areprobably
independently regulated processes (reviewed in Ref.140). There are
also a variety of types of mucin (140), andthese may not correspond
exactly between humans and ani-mals. All of these factors make
pathological smoke-inducedchanges related to mucin production
somewhat difficult tomodel in animals.
In the guinea pig model, chronic cigarette smoke exposureinduces
secretory cell metaplasia of the small non-cartilaginousairway
epithelium (185) (Fig. 13), a finding reasonably analo-
gous to that seen in humans, but does not produce the
luminalmucous plugs seen in patients with COPD (185).
Smokingcessation reduces the degree of metaplasia (187). By
contrast,in mice, tobacco smoke has a relatively minor effect, with
onlya few secretory cells appearing in the small airways (11).
Some studies report significant degrees of goblet cell
meta-plasia in smoke-exposed rats (84, 138, 139, 177), and this
can
Fig. 10. Peribronchiolar lymphoid aggregate in a mouse exposed
to smoke for6 mo. Note the pigmented smokers macrophages (arrows),
a finding similar tothat seen in human smokers.
Fig. 11. Vascular remodeling. Smooth muscle actinstains of small
intrapulmonary arterial branchesfrom control (A) and animal exposed
to smoke for 6mo (B). Note the marked increase in smooth
muscle.Such increases correlate with increased pulmonaryarterial
pressure.
Invited Review
L624 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
15/21
be prevented, or postexposure regression enhanced, with
theantioxidant/mucolytic agent N-acetyl cysteine, as well as
avariety of nonsteroidal anti-inflammatory agents, and
steroids(138, 139). In rats, the gastro-protective agent
rebamipide(OPC-12759) decreased TNF production and reducedMUC5AC
production, reduced inflammatory cells in BAL, andinhibited
smoke-induced goblet cell metaplasia in the trachea(89). Tracheal
goblet cell metaplasia in guinea pigs was in-duced by a 2-wk smoke
exposure and could be attenuated by
use of a platelet-activating factor antagonist (82). By
contrast,a PDE4 inhibitor had no significant effect on the
minimalgoblet cell metaplasia induced by cigarette smoke in the
mousemodel (106). These findings suggest that goblet cell
metapla-
sia/mucin production is driven by a variety of mechanisms.Some
of these have been elucidated in tissue culture systems(reviewed in
Refs. 140, 175).
Investigation of exacerbations of COPD is an area of
greatinterest, as exacerbations are a major (poor) prognostic
featurein human smokers and are a major health care burden
(174).This phenomenon is complicated by a lack of a universally
accepted definition (13, 101), and the triggers of
exacerbationare not definitively known; bacterial and viral agents
aresuggested instigators. A recent human model of virus
infectionusing rhinovirus recapitulated the symptoms and signs of
anexacerbation (123). Regardless of etiology, there is evidence
ofupper and lower respiratory tract inflammation during
exacer-bations (70).
Since chronic bronchitis is a clinical definition,
involvingcough and sputum production, it is difficult to establish
ananimal model as sputum production is difficult to monitor
inanimals and probably sparse; the lack of diffuse increases
inmucus production likewise makes it difficult to make models
ofacute exacerbations. Potential models of exacerbation
includeadministration of lipopolysaccharide or administration of
bac-terial or viral agents with or without cigarette smoke.
Forrecent reviews on models of acute exacerbations, the reader
isreferred to Refs. 54, 80, 1017.
Conclusions
Taken as a whole, the data on proteases and anti-inflamma-tory
agents still support the idea that inflammatory
cell-derivedproteases are the major mediators of emphysema in
animalmodels. However, which cells/proteases are most important
inthis process remains uncertain, and there are clearly
complexinteractions among them (Fig. 3). Antioxidant protection
isimportant as well, and oxidative stress is linked to increases
ininflammation and decreases in antiproteolytic protection,
thereby connecting oxidative damage to the
protease-antipro-tease hypothesis (Fig. 3). As opposed to these
various initiatingfactors, failure of the parenchyma to properly
repair alsoappears to play a role in the genesis of emphysema, but
the data
Fig. 12. Postulated mechanisms involved in the development of
pulmonaryhypertension in COPD. Smoke (probably oxidant)-driven
increases in vaso-proliferative and vasoconstrictive agents
(endothelin and VEGF) and TNF-driven increases in MMP activation
result in vascular remodeling and endo-thelial dysfunction. The
normally vasorelaxant endothelial nitric oxide syn-
thase (eNOS) pathway is disrupted by smoke-generated oxidants
and alsoby TNF, thus increasing the vasoconstrictive effects and
endothelial dysfunc-tion. Vascular remodeling and endothelial
dysfunction act in a vicious circleproducing pulmonary
hypertension. Modeled from Refs. 189, 190, 191.
Fig. 13. Goblet cell metaplasia. Periodic acid
Schiff/diastase-stained sections of small airways from
controlguinea pig (A) and guinea pig exposed to smoke for 6mo (B).
Note the increased numbers of mucin-contain-ing gobletcells in the
epithelium of thesmoke-exposedanimal.
Invited Review
L625ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
16/21
are more controversial and most failure-to-repair models havenot
used cigarette smoke.
This wealth of data in fact produces the interesting conun-drum
that there are too many potential actors, and sorting outwhich
processes are fundamental to the development of em-physema and
which are epiphenomena is a crucial and difficultproblem; in this
regard, it appears that MMP-12 is consistently
implicated (when looked for) in the murine models and
mayrepresent a central chokepoint (Fig. 3). Whether this is true
ofhumans is an important question.
One of the puzzling facts about the role of proteases
andoxidative stress has always been why only some smokers,mouse or
human, develop emphysema while most are spared.Part of the answer
probably lies in the genotypic ability ofindividuals to mount an
inflammatory/antioxidant response.Recent data on the role of T
cells and the adaptive immuneresponse suggest a new paradigm that
could partially explainthis question, namely that COPD (at least
emphysema) is aconsequence of an autoimmune reaction to antigenic
peptidesderived from the breakdown of elastin and perhaps other
lungcomponents (37, 38, 98, 97, 100). Thus proteases along
withreactive oxygen species would be the initial culprits
initiatingthe breakdown of elastic and collagen tissue and
producing thenecessary antigenic peptides to potentially trigger
the autoim-mune reaction. However, only genetically predisposed
hu-mans, and mice, would develop the adaptive immune
responsenecessary to enhance and maintain the initial
inflammatoryresponse to cigarette smoke and eventually produce
disease.
The animal data suggest a variety of therapeutic approachesto
human emphysema [indeed, thus far animal models haveaddressed only
a few out of many potential targets (40, 41)],but translation to
the human setting is not straightforward, asevidenced by the
apparent lack of efficacy of anti-TNFtherapy in humans (10, 135,
174) compared with the clear role
of TNF in murine emphysema (32, 176). The reason for
thisdiscrepancy is unclear but probably relates in large part to
theinability to produce in laboratory animals severe COPD
withcigarette smoke; i.e., the smoke-induced animal models
arereproducing GOLD Stage 1 and 2 disease, but symptomaticCOPD
patients have GOLD 3/4 disease, and thus far no modelreproduces
either the anatomic or functional changes or thesmoke-independent
progression (136), seen in such patients.This finding implies that
the mechanisms driving human earlystage and late stage disease may
be quite different and that theanimal models are limited to
relatively early stage disease, butthat does not make the animal
models useless, since effectiveearly intervention is probably much
easier to design thaneffective late stage intervention.
Studies on SAR and PHT suggest that these processes are atleast
partially independent of the factors that drive emphysema,although
there are clearly areas of overlap, such as the role ofoxidants in
SAR and serine elastases in vascular remodeling,and the same is
true of increased mucus production and gobletcell metaplasia. This
situation implies that a therapeutic ap-proach to human COPD may
have to target each anatomiccompartment and a single therapeutic
agent may not be able toprevent all of the separate
manifestations.
GRANTS
This work was supported by Canadian Institutes of Health
Research Grants42539 and 81409.
REFERENCES
1. Adair-Kirk TL, Atkinson JJ, Broekelmann TJ, Doi M,
TryggvasonK, Miner JH, Mecham RP, Senior RM. A site on laminin
alpha 5,AQARSAASKVKVSMKF, induces inflammatory cell production
ofmatrix metalloproteinase-9 and chemotaxis. J Immunol 171:
398406,2003.
2. Aoshiba K, Koinuma M, Yokohori N, Nagai A.
Immunohistochemicalevaluation of oxidative stress in murine lungs
after cigarette smoke
exposure. Inhal Toxicol 15: 10291038, 2003.3. Aoshiba K,
Yokohori N, Nagai A. Alveolar wall apoptosis causes lung
destruction and emphysematous changes. Am J Respir Cell Mol Biol
28:555562, 2003.
4. Arikan MC, Shapiro SD, Mariani TJ. Induction of macrophage
elas-tase (MMP-12) gene expression by statins. J Cell Physiol 204:
139145,2005.
5. Banerjee S, Maity P, Mukherjee S, Sil AK, Panda K,
ChattopadhyayD, Chatterjee IB. Black tea prevents cigarette
smoke-induced apoptosisand lung damage. J Inflamm 4: 3, 2007.
6. Barbera JA, Peinado VI, Santos S. Pulmonary hypertension in
chronicobstructive pulmonary disease. Eur Respir J 21: 892905,
2003.
7. Barnes PJ, Shapiro SD, Pauwels RA. Chronic obstructive
pulmonarydisease: molecular and cellular mechanisms. Eur Respir J
22: 672688,2003.
8. Barnes PJ. Emerging targets for COPD therapy. Curr Drug
Targets
Inflamm Allergy 4: 675683, 2005.9. Barnes PJ. Reduced histone
deacetylase in COPD: clinical implications.
Chest 129: 151155, 2006.10. Barnes PJ. Unexpected failure of
anti-tumor necrosis factor therapy in
chronic obstructive pulmonary disease. Am J Respir Crit Care Med
175:866867, 2007.
11. Bartalesi B, Cavarra E, Fineschi S, Lucattelli M, Lunghi B,
Mar-torana PA, Lungarella G. Different lung responses to cigarette
smokein two strains of mice sensitive to oxidants. Eur Respir J 25:
1522,2005.
12. Belloni PN, Garvin L, Mao CP, Bailey-Healy I, Leaffer D.
Effects ofall-trans-retinoic acid in promoting alveolar repair.
Chest 117: 235S2341S, 2000.
13. Berge S, Wedzicha JA. COPD exacerbations: definitions and
classifi-cations. Eur Respir J 21, S41: 46S53S, 2003.
14. Bowler RP, Barnes PJ, Crapo JD. The role of oxidative stress
in
chronic obstructive pulmonary disease. COPD 1: 255277, 2004.15.
Bracke KR, Dhulst AI, Maes T, Moerloose KB, Demedts IK, Leb-ecque
S, Joos GF, Brusselle GG. Cigarette smoke-induced
pulmonaryinflammation and emphysema are attenuated in
CCR6-deficient mice.
J Immunol 177: 43504359, 2006.16. Bracke KR, Dhulst AI, Maes T,
Demedts IK, Moerloose KB, Kuziel
WA, Joos GF, Brusselle GG. Cigarette smoke-induced
pulmonaryinflammation, but not airway remodelling, is attenuated in
chemokinereceptor 5-deficient mice. Clin Exp Allergy 37: 14671479,
2007.
17. Bridges RB, Fu MC, Rehm SR. Increased neutrophil
myeloperoxidaseactivity associated with cigarette smoking. Eur J
Respir Dis 67: 8493,1985.
18. Brusselle GG, Bracke KR, Maes T, Dhulst AI, Moerloose KB,
JoosGF, Pauwels RA. Murine models of COPD. Pulm Pharmacol Ther
19:155165, 2006.
19. Budev MM, Arroliga AC, Wiedemann HP, Matthay RA. Cor
pulmo-nale: an overview. Semin Respir Crit Care Med 24: 233243,
2003.
20. Cantor JO, Shteyngart B, Cerreta JM, Liu M, Armand G,
TurinoGM. The effect of hyaluronan on elastic fiber injury in vitro
andelastase-induced airspace enlargement in vivo. Proc Soc Exp Biol
Med225: 6571, 2000.
21. Cantor JO, Cerreta JM, Ochoa M, Ma S, Chow T, Grunig G,
TurinoGM. Aerosolized hyaluronan limits airspace enlargement in a
mousemodel of cigarette smoke-induced pulmonary emphysema. Exp Lung
Res31: 417430, 2005.
22. Cardoso WV, Sekhon HS, Hyde DM, Thurlbeck WM. Collagen
andelastin in human pulmonary emphysema. Am Rev Respir Dis
147:975981, 1993.
23. Carnevali S, Nakamura Y, Mio T, Liu X, Takigawa K,
RombergerDJ, Spurzem JR, Rennard SI. Cigarette smoke extract
inhibits fibro-blast-mediated collagen gel contraction. Am J
Physiol Lung Cell MolPhysiol 274: L591L598, 1998.
Invited Review
L626 ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
17/21
24. Cavarra E, Bartalesi B, Lucattelli M, Fineschi S, Lunghi B,
GambelliF, Ortiz LA, Martorana PA, Lungarella G. Effects of
cigarette smokein mice with different levels of 1-proteinase
inhibitor and sensitivity tooxidants. Am J Respir Crit Care Med
164: 886890, 2001.
25. Cavarra E, Lucattelli M, Gambelli F, Bartalesi B, Fineschi
S, SzarkaA, Giannerini F, Martorana PA, Lungarella G. Human SLPI
inacti-vation after cigarette smoke exposure in a new in vivo model
ofpulmonary oxidative stress. Am J Physiol Lung Cell Mol Physiol
281:L412L417, 2001.
26. Celli BR, Barnes PJ. Exacerbations of chronic obstructive
pulmonarydisease. Eur Respir J 29: 12241238, 2007.
27. Churg A, Cherukupalli K. Cigarette smoke causes rapid lipid
peroxi-dation of rat tracheal epithelium. Int J Exptl Pathol 74:
127132, 1993.
28. Churg A, Dai J, Tai H, Xie C, Wright JL. Tumor necrosis
factor-alphais central to acute cigarette smoke-induced
inflammation and connectivetissue breakdown. Am J Respir Crit Care
Med 166: 849854, 2002.
29. Churg A, Zay K, Shay S, Xie C, Shapiro SD, Hendricks R,
Wright J.Acute cigarette smoke induced connective tissue breakdown
requiresboth neutrophils, and macrophage metalloelastase in mice.
Am J RespirCell Mol Biol 27: 368374, 2002.
30. Churg A, Wang RD, Xie C, Wright JL. 1-Antitrypsin
amelioratescigarette smoke-induced emphysema in the mouse. Am J
Respir CritCare Med 168: 199207, 2003.
31. Churg A, Wang RD, Tai H, Wang X, Xie C, Dai J, Shapiro
SD,Wright JL. Macrophage metalloelastase mediates acute cigarette
smoke-
induced inflammation via tumor necrosis factor-alpha release. Am
JRespir Crit Care Med 167: 10831089, 2003.
32. Churg A, Wang RD, Tai H, Wang X, Xie C, Wright JL.
Tumornecrosis factor-alpha drives 70% of cigarette smoke-induced
emphysemain the mouse. Am J Respir Crit Care Med 170: 492498,
2004.
33. Churg A, Tai H, Coulthard T, Wang R, Wright JL. Cigarette
smokedrives small airway remodeling by induction of growth factors
in theairway wall. Am J Respir Crit Care Med 174: 13271334,
2006.
34. Churg A, Wang R, Wang X, Onnervik PO, Thim K, Wright JL.
AnMMP-9/-12 inhibitor prevents smoke-induced emphysema and
smallairway remodeling in guinea pigs. Thorax 62: 706713, 2007.
35. Churg A, Wang X, Wang RD, Meixner SC, Pryzdial ELG,
WrightJL. 1-Antitrypsin suppresses TNF and MMP12 production by
ciga-rette smoke-stimulated macrophages. Am J Respir Cell Mol Biol
37:144151, 2007.
36. Cooper R, Ghali J, Simmons BE, Castaner A. Elevated
pulmonaryartery pressure. Chest 99: 112120, 1991.
37. Cosio MG, Majo J, Cosio MG. Inflammation of the airways and
lungparenchyma in COPD: role of T cells. Chest 121, Suppl 5:
160S165S,2002.
38. Cosio MG. Autoimmunity, T-cells and STAT-4 in the
pathogenesis ofCOPD. Eur Respir J 24: 35, 2004.
39. Damiano VV, Tsang A, Kucich U, Abrams WR, Rosenbloom
J,Kimbel P, Fallahnejad M, Weinbaum G. Immunolocalization of
elas-tase in human emphysematous lungs. J Clin Invest 78: 482493,
1986.
40. de Boer WI, Alagappan VK, Sharma HS. Molecular mechanisms
inchronic obstructive pulmonary disease; potential targets for
therapy. Cell
Biochem Biophys 47: 131148, 2007.41. de Boer WI. Perspectives
for cytokine antagonist therapy in COPD.
Drug Discov Today 10: 93106, 2005.42. Demedts IK, Morel-Montero
A, Lebecque S, Pacheco Y, Cataldo D,
Joos GF, Pauwels RA, Brusselle GG. Elevated MMP-12 protein
levelsin induced sputum from COPD patients. Thorax 61: 1962015,
2006.
43. Demeo DL, Hersh CP, Hoffman EA, Litonjua AA, Lazarus
R,Sparrow D, Benditt JO, Criner G, Make B, Martinez FJ, ScanlonPD,
Sciurba FC, Utz JP, Reilly JJ, Silverman EK. Genetic determi-nants
of emphysema distribution in the National Emphysema TreatmentTrial.
Am J Respir Crit Care Med 176: 4248, 2007.
44. Dhami R, Gilks B, Xie C, Zay K, Wright J, Churg A. Acute
cigarettesmoke-induced connective tissue breakdown is mediated by
neutrophilsand prevented by 1-antitrypsin. Am J Respir Cell Mol
Biol 22: 244252,2000.
45. Dhulst AI, Maes T, Bracke KR, Demedts IK, Tournoy KG, Joos
GF,Brusselle GG. Cigarette smoke-induced pulmonary emphysema in
scid-mice. Is the acquired immune system required? Respir Res 6:
147, 2005.
46. Dhulst AI, Bracke KR, Maes T, De Bleecker JL, Pauwels RA,
JoosGF, Brusselle GG. Role of tumour necrosis factor-alpha receptor
p75 incigarette smoke-induced pulmonary inflammation and emphysema.
Eur
Respir J 28: 102112, 2006.
47. Eidelman D, Saetta MP, Ghezzo H, Wang NS, Hoidal JR, King
M,Cosio MG. Cellularity of the alveolar walls in smokers and its
relation toalveolar destruction. Functional implications. Am Rev
Respir Dis 141:15471552, 1990.
48. Fabbri L, Pauwels RA, Hurd SS. Scientific Committee GOLD.
Globalstrategy for the diagnosis, management, and prevention of
chronicobstructive pulmonary disease: GOLD executive summary
updated 2003.COPD 1: 105141, 2003.
49. Finkelstein R, Fraser RS, Ghezzo H, Cosio MG. Alveolar
inflamma-tion and its relation to emphysema in smokers. Am J Respir
Crit Care
Med 152: 16661672, 1995.50. Finlay GA, ODriscoll LR, Russell KJ,
DArcy EM, Masterson JB,
FitzGerald MX, OConnor CM. Matrix metalloproteinase
expressionand production by alveolar macrophages in emphysema. Am J
RespirCrit Care Med 156: 240247, 1997.
51. Foronjy RF, Mercer BA, Maxfield MW, Powell CA, DArmiento
J,Okada Y. Structural emphysema does not correlate with lung
compli-ance: lessons from the mouse smoking model. Exp Lung Res
31:547562, 2005.
52. Foronjy RF, Mirochnitchenko O, Propokenko O, Lemaitre V, Jia
Y,Inouye M, Okada Y, DArmiento JM. Superoxide dismutase expres-sion
attenuates cigarette smoke- or elastase-generated emphysema inmice.
Am J Respir Crit Care Med 173: 623631, 2006.
53. Gao S, Chen K, Zhao Y, Rich CB, Chen L, Li SJ, Toselli P,
Stone P,Li W. Transcriptional and posttranscriptional inhibition of
lysyl oxidase
expression by cigarette smoke condensate in cultured rat fetal
lungfibroblasts. Toxicol Sci 87: 197203, 2005.
54. Gaschler GJ, Bauer CM, Zavitz CC, Stampfli MR. Animal models
ofchronic obstructive pulmonary disease exacerbations. Contrib
Microbiol14: 126141, 2007.
55. Gilmore AP. Anoikis. Cell Death Differ 12, Suppl 2:
14731477, 2005.56. Girgis RE, Mozammel S, Champion HC, Li D, Peng
X, Shimoda L,
Tuder RM, Johns RA, Hassoun PM. Regression of chronic
hypoxicpulmonary hypertension by simvastatin. Am J Physiol Lung
Cell MolPhysiol 292: L1105L1110, 2007.
57. Goco RV, Kress MB, Brantigan OC. Comparison of mucus glands
inthe tracheobronchial tree of man and animals. Ann NY Acad Sci
106:555571, 1963.
58. Grootendorst DC, Gauw SA, Verhoosel RM, Sterk PJ, Hospers
JJ,Bredenbroker D, Bethke TD, Hiemstra PS, Rabe KF. The
PDE4inhibitor roflumilast reduces sputum neutrophil and eosinophil
numbersin patients with COPD. Thorax 62: 10811087, 2007.
59. Gross P, Pfitzer EA, Toker A. Experimental emphysema: its
productionwith papain in normal and silicotic rats. Arch Environ
Health 11: 5058,1965.
60. Grumelli S, Corry DB, Song LZ, Song L, Green L, Huh J,
Hacken J,Espada R, Bag R, Lewis DE, Kheradmand F. An immune basis
forlung parenchymal destruction in chronic obstructive pulmonary
diseaseand emphysema. PLoS Med 1: e8, 2004.
61. Guerassimov A, Hoshino Y, Takubo Y, Turcotte A, Yamamoto
M,Ghezzo H, Triantafillopoulos A, Whittaker K, Hoidal JR, Cosio
MG.
The development of emphysema in cigarette smoke-exposed mice
isstrain dependent. Am J Respir Crit Care Med 170: 974980,
2004.
62. Halbert RJ, Natoli JL, Gano A, Badamgarav E, Buist AS,
ManninoDM. Global burden of COPD: systematic review and
meta-analysis. Eur
Respir J 28: 523532, 2006.63. Hautamaki R, Kobayashi D, Senior
R, Shapiro S. Requirement for
macrophage elastase for cigarette smoke-induced emphysema in
mice.
Science 277: 20022004, 1997.64. Henson PM, Vandivier RW. The
matrix degrades, neutrophils invade.Nat Med 12: 280281, 2006.
65. Hogg JC, Chu F, Utokaparch S, Woods R, Elliott WM, Buzatu
L,Cherniack RM, Rogers RM, Sciurba FC, Coxson HO, Pare PD.
Thenature of small-airway obstruction in chronic obstructive
pulmonarydisease. N Engl J Med 350: 26452653, 2004.
66. Hoidal JR, Fox RB, LeMarbe PA, Perri R, Repine JE.
Alteredoxidative metabolic responses in vitro of alveolar
macrophages fromasymptomatic cigarette smokers. Am Rev Respir Dis
123: 8589, 1981.
67. Holz O, Zuhlke I, Jaksztat E, Muller KC, Welker L, Nakashima
M,Diemel KD, Branscheid D, Magnussen H, Jorres RA. Lung
fibroblastsfrom patients with emphysema show a reduced
proliferation rate inculture. Eur Respir J 24: 575579, 2004.
68. Houghton AM, Quintero PA, Perkins DL, Kobayashi DK,
KelleyDG, Marconcini LA, Mecham RP, Senior RM, Shapiro SD.
Elastin
Invited Review
L627ANIMAL MODELS OF COPD
AJP-Lung Cell Mol Physiol VOL 294 APRIL 2008 www.ajplung.org
-
8/22/2019 Cigarette Induce COPD
18/21
fragments drive disease progression in a murine model of
emphysema.J Clin Invest 116: 753759, 2006.
69. Hunninghake GW, Davidson JM, Rennard S, Szapiel S, Gadek
JE,Crystal RG. Elastin fragments attract macrophage precursors to
diseasedsites in pulmonary emphysema. Science 212: 925927,
1981.
70. Hurst JR, Perera WR, Wilkinson TMA, Donaldson GC,
WedzichaJA. Systemic and upper and lower airway inflammation at
exacerbationof chronic obstructive pulmonary disease. Am J Respir
Crit Care Med
173: 7178, 2006.71. Iizuka T, Ishii Y, Itoh K, Kiwamoto T,
Kimura T, Matsuno Y,Morishima Y, Hegab AE, Homma S, Nomura A,
Sakamoto T,
Shimura M, Yoshida A, Yamamoto M, Sekizawa K. Nrf2-deficientmice
are highly susceptible to cigarette smoke-induced emphysema.Genes
Cells 10: 11131125, 2005.
72. Imai K, Dalal SS, Chen ES, Downey R, Schulman LL, Ginsburg
M,DArmiento J. Human collagenase (matrix metalloproteinase-1)
expres-sion in the lungs of patients with emphysema. Am J Respir
Crit Care Med163: 786791, 2001.
73. Imai K, Mercer BA, Schulman LL, Sonnett R, DArmiento
JM.Correlation of lung surface area to apoptosis and proliferation
in humanemphysema. Eur Respir J 25: 250258, 2005.
74. Ito K, Ito M, Elliott WM, Cosio B, Caramori G, Kon OM,
BarczykA, Hayashi S, Adcock IM, Hogg JC, Barnes PJ. Decreased
histonedeacetylase activity in chronic obstructive pulmonary
disease. N Engl
J Med 352: 19671976, 2005.
75. Jamal K, Fleetham JA, Thurlbeck WM. Cor pulmonale:
correlationwith central airway lesions, peripheral airway lesions,
emphysema, andcontrol of breathing. Am Rev Respir Dis 141:
11721177, 1990.
76. Janoff A. Elastases and emphysema. Current assessment of the
protease-antiprotease hypothesis. Am Rev Respir Dis 132: 417433,
1985.
77. Jeffery PK. Remodeling in asthma and chronic obstructive
lung disease.Am J Respir Crit Care Med 164: S28S38, 2001.
78. Kang MJ, Homer RJ, Gallo A, Lee CG, Crothers KA, Cho
SJ,Rochester C, Cain H, Chupp G, Yoon HJ, Elias JA. IL-18 is
inducedand IL-18 receptor alpha plays a critical role in the
pathogenesis ofcigarette smoke-induced pulmonary emphysema and
inflammation. J
Immunol 178: 19481959, 2007.79. Kasahara Y, Tuder RM,
Taraseviciene-Stewart L, Le Cras TD,
Abman S, Hirth PK, Waltenberger J, Voelkel NF. Inhibition of
VEGFreceptors causes lung cell apoptosis and emphysema. J Clin
Invest 106:13111319, 2000.
80. Kharitonov SA, Sjobring U. Lipopolysaccharide challenge of
humansas a model for chronic obstructive lung disease
exacerbations. Contrib
Microbiol 14: 83100, 2007.81. Kleeberger SR, Peden D.
Gene-environment interactions in asthma and
other respiratory diseases. Annu Rev Med 56: 383400, 2005.82.
Komori M, In












![Research Paper Cigarette smoke-induced lung inflammation ......CCR1 expression are associated with increased risk of exacerbations in patients with COPD [12–16]. However, the critical](https://img.pdfslide.us/doc/110x75/5f1b82ffdb5f9216f371d576/research-paper-cigarette-smoke-induced-lung-inflammation-ccr1-expression.jpg)
![PrimedLowFrequencyTranscranialMagneticStimulation ... · A smoking-cue paradigm was used to induce cigarette cravingfollowingpreviously established methods [7]. We used E-prime software](https://img.pdfslide.us/doc/110x75/5f64033276b75b7f671435a0/primedlowfrequencytranscranialmagneticstimulation-a-smoking-cue-paradigm-was.jpg)