Embed Size (px)
Citation preview

Neuroscienee Letters, 140 (1992) 5 8 5 c) 1992 Elsevier Scientific Publishers Ireland Ltd. All rights reserved 0304-3940/92/$ 05.00
NSL 08649
Chronic estradiol treatment increases anterior pituitary but not striatal D 2 dopamine receptor mRNA levels in rats
Daniel Ldvesque a, Bernard Gagn6 b, Nicholas Barden b and Thdrbse Di Paolo"
"Department q/'Molecular Endocrinology, CHUL Researeh Center and Sehool ql'Pharma<v, Laval University, Ste-Foy, Que. (Canada) and eDepartment of Molecular Psychogeneties, CH UL Research Centre and La val University, Ste-Foy. Que. ~ (~mada )
(Received 5 August 1991: Revised version received 2 March 1992: Accepted 3 March 1992)
Key words': Dopamine D 2 receptor mRNA: 17fl-Estradiol; Rat anterior pituitary gland: Rat striatum: Northern blot analysis
The effect of chronic 17fl-estradiol treatment (10 ,ug, twice each day, for 2 weeks) of ovariectomized rats on D e dopamine (DA) receptor mRNA levels was investigated in striatum and anterior pituitary gland tissues. We used 32P-labeled probes specific for D~ receptor and fl-actin mRNAs in Northern blot analysis. The ratio of D e DA receptor mRNA/fl-actin mRNA level was significantly increased in the anterior pituitary of estradiol- treated rats compared to vehicle-treated animals. The D2 DA receptor mRNA/fl-actin mRNA ratio in the striatum was not affected by estradiol treatment. However, the medial portion of the striatum showed a significantly lower ratio compared to the lateral portion of the striatum in both vehicle- and estradiol-treated rats. Thus, the estradiol effect on anterior pituitary D~ receptors may implicate transcriptional regulation, whereas our results do not support this hypothesis for the estradiol action on striatal D 2 receptors.
It is now well documented that estradiol affects D, dopamine (DA) receptors in the central nervous system of mammals [19]. In rat, chronic estradiol treatment in- creases the density of striatal D 2 DA receptors [7 9, 12, 19]. In the anterior pituitary, the modulation by estrogen of D 2 DA receptors is more complex and depends on the doses and duration of exposure to this hormone [7]. Lac- totroph cells of the adenohypophysis bearing DA recep- tors contain high estradiol receptor levels [17] as meas- ured with [3H]estradiol binding while, using similar tech- niques, no binding is observed in the striatum [18]. Hence, a classical genomic steroid receptor for estrogen is documented in the anterior pituitary while in the stria- tum no receptor for estrogen or alternatively the pres- ence of a different receptor is likely. Therefore, in spite of extensive investigation, the mechanism underlying the interaction between DAergic systems and steroid hormones is still poorly understood. In this report we studied the effect of chronic treatment of rats with estra- diol o n D 2 receptor mRNA levels in both anterior pitui- tary gland and striatum.
Adult Sprague-Dawley female rats (250 350 g) were
Corre.spondence: T. Di Paolo, Centre de Recherches du CHUL, Centre Hospitalier de l'Universit6 Laval, Ddpartement d'Endocrinologie Moldculaire, 2705, Boulevard Laurier, Ste-Foy, Que. GIV 4G2 Can- ada. Fax: (I) (418) 654-2761.
purchased from Charles River Canada Inc. (St-Constant, Que.). They were bilaterally ovariectomized (OVX) under ether anesthesia, housed two per cage and main- tained at 22 23°C on a 14/10 h light/dark cycle (lights on from 05.00 h to 19.00 h). They had free access to rat chow and water. The day after ovariectomy, ten animals received subcutaneous injections of 10 #g of 17fl-estra- diol (Sigma) in 0.2 ml, twice each day for 2 weeks, while a second control group (10 animals) received injections of the vehicle only (0.3% gelatin in saline solution). All animals were sacrificed by decapitation in the morning (between 09.00 h and 11.00 h) the day after their last injection.
The brain was removed from the skull and the stria- turn was immediately dissected into its lateral and medial portions, frozen in liquid nitrogen and kept at -70°C until assay. Anterior pituitary tissue was separated from the intermediate and posterior lobes, rapidly frozen in liquid nitrogen and kept at -70°C until assayed.
Total RNA was isolated by a modification of the method described by Chirgwin et al. [4]. Frozen tissue was homogenized in 0.66 ml of 5 M guanidium isothiocy- anate, 50 mM Tris-HCl pH 7.6, 10 mM EDTA, 5% fl- mercaptoethanol, 3% sodium lauryl sarcosine and a trace of Antifoam-A (Sigma) with 10 strokes of a 2.0 ml Potter-Elvejeim apparatus. After addition of 0.33 ml of 80% (w/v) cesium chloride (CsCI) in homogenization

~ - - 28S rRNA
D 2 mRNA
13- Actin mRNA
iii! ili! ! ii i:iii ! i!ii!
Fig. I. Northern blot analysis of D2 mRNA relative to fl-actin mRNA levels in anterior pituitary. Total RNA (t0/,tg) from pituitary was sepa- rated on 1.0% agarose-formaldehyde denaturing gel, blotted on a nylon membrane, and successively hybridized at 65°C with D~ and ,8-actin cRNA probes. Under these stringent conditions, single strong bands, corresponding to the ~2.5 kb Dz mRNA and the ~2.0 kb,8-actin mRNA
are identified.
buffer to the homogenate, the mixture was carefully lay- ered on 0.3 ml of a 5.7 M CsCI, 100 mM EDTA cushion in a 1.5 ml polycarbonate tube and centrifuged in a Beck- man TL-100 ultracentrifuge at 70,000 rpm for 4 h at 20°C. Pellets were resuspended in TE buffer ( 10 mM Tris pH 8.0, 1 mM EDTA) 1% SDS. RNA was twice precipi- tated from 0.3 M sodium acetate by addition of 2 vols, of ethanol at -80°C. RNA was collected by centrifugation at 15,000 × g at 4°C for 20 min. Final pellets were dried under vacuum and resuspended in 150 ~1 1 x TE. Ali- quots were taken for estimation of purity and RNA con- tent was determined by spectrophotometric absorbance at 260/280 nm.
Northern blot filters were prepared by electrophoresis of 10 ~tg of RNA on a 1% agarose denaturing gel con- taining formaldehyde at 30 mA overnight with subse- quent transfer of the separated RNA to a nylon filter
(Hybond-N, Amersham) by passive diffusion under high salt concentrations.
D~ mRNA affixed to nylon filters was hybridized to a 495 bp D 2 cRNA [32p]-UTP labelled probe. Probe copies were generated from a rat D2 cDNA cloned into p64T7 [14]. Control of the total amount of RNA on Northern blots was achieved by hybridization with afl-actin cRNA probe produced from a 1,500 bp fl-actin Pstl fragment [10] inserted into the pGEM-I riboprobe system vector (Promega Biotech, Madison, WI). Nylon filters were pre- hybridized for 4 h at 42°C in 50% formamide 5 × SSC, 0.06% Ficoll, 0.06% polyvinytpyrrolidone, 0.06% BSA, 50 mM sodium phosphate pH 6.5, 250/,tg/ml denatured salmon sperm DNA (ssDNA), 250 /,tg/ml tRNA and 0.1% SDS. The filters were first hybridized with the D~ probe at 65°C for 18-24 h in the same buffer containing 1.5 x 10 ~' cpm [s2P]-labelled D2 cRNA per ml of buffer. At the end of the incubation, filters were washed in 1 × SSC, 0.1% SDS twice for 30 min at room temperature and then in 0.1 × SSC, 0.1% SDS twice for I h at 68°C. Under these stringent conditions a single radioactive band was detected with the D 2 cRNA probe. Filters were air-dried and exposed to Kodak X-Omat film (Eastman Kodak, Rochester, NY) with a Cronex intensifying screen (Du- pont, Wilmington, DE) at -80°C. The hybridization procedure was subsequently repeated on the same filter using the,8-actin cRNA probe.
The resulting autoradiograms were analyzed by meas- urements of mean gray tone with a Ras-image analysis system (Amersham). All comparisons of mRNA levels on autoradiograms were made from RNA samples hy- bridized on the same filter. The statistical significance of any difference between groups was analyzed with the Duncan-Kramer multiple range test [11] after one-way analysis of variance (ANOVA).
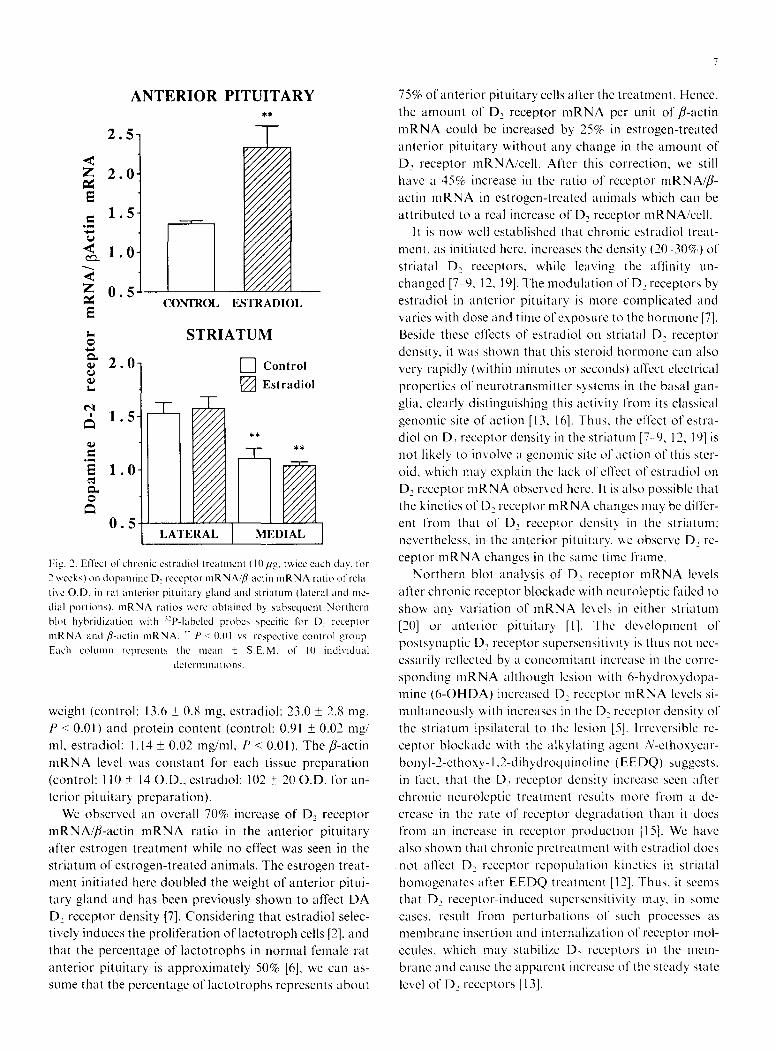
Fig. 1 shows a representative example of an autoradio- gram from anterior pituitary RNA preparation showing the D~ receptor mRNA and fl-actin mRNA bands. Chronic treatment with estradiol (10/Jg, twice each day for 2 weeks) increased the ratio of D2 receptor mRNA/fl- actin mRNA level in the rat anterior pituitary gland (70% increase, P < 0.01) (Fig. 2). Neither lateral nor me- dial striatal D2 receptor mRNA/fl-actin mRNA ratios were affected by the estradiol treatment (Fig. 2). How- ever, in both vehicle- and estradiol-treated animals, a sig- nificantly lower D2 receptor mRNA/fl-actin mRNA ratio was seen in the medial striatum compared to the lateral striatum. This observation confirmed previous data ob- tained by in situ hybridization [21], and indicates that D~ receptor mRNA distribution follows the distribution of D~ receptor density, which also has a lateral-to-medial gradient of distribution in the rat striatum [3]. Estradiol treatment also increased the rat anterior pituitary gland
,< Z
E
.< Z
E t _
° ~
E
2 , 5 -
2 . 0 .
1 .5 -
1 . 0
0 . 5
ANTERIOR PITUITARY
CONTROL ESTRADIOL
STRIATUM
2 .0 - [ ] Control [ ] Estradiol
1.5.
1.0 ~ 0 . 5 LATERAL [ MEDIAL ]
Fig. 2. Effect of chronic estradiol treatment (10 ,ug, twice each clay, for 2 weeks) on dopamine D, receptor mRNA,,fl-actin mRNA ratio of rela- tive O.D. in rat anterior pituitary gland and striatum (lateral and me- dial portions), mRNA ratios were obtained by subsequent Northern bh,t hybridization with '2P-labeled probes specific liar D e receptor mRNA and ,&actin mRNA. *" P < 0.01 vs. respective control group. Each cohunn represents the mean + S.E.M. of 10 individual
determinations.
weight (control: 13.6 + 0.8 mg, estradiol: 23.0 _+ 2.8 mg,
P < 0.01) and protein content (control: 0.91 + 0.02 mg/ ml, estradiol: 1.14 + 0.02 mg/ml, P < 0.01). The fl-actin
m R N A level was constant for each tissue preparation
(control: 110 k 14 O.D., estradiol: 102 _+ 20 O.D. for an- terior pituitary preparation).
We observed an overall 70% increase of De receptor
mRNA//3-actin mRNA ratio in the anterior pituitary after estrogen treatment while no effect was seen in the
striatum of estrogen-treated animals. The estrogen treat- metat initiated here doubled the weight of anterior pitui- tary gland and has been previously shown to affect DA
De receptor density [7]. Considering that estradiol selec- tively induces the proliferation oflactotroph cells [2], and that the percentage of lactotrophs in normal female rat
anterior pituitary is approximately 50% [6], we can as-
sume thai the percentage of lactotrophs represents about
75% of anterior pituitary cells after the treatment. Hence,
the amount of De receptor m R N A per unit of fl-actin m R N A could be increased by 25% in estrogen-treated anterior pituitary without any change in the amount of
D e receptor mRNA/cell. After this correction, we still have a 45% increase in the ratio of receptor mRNA/fl-
actin m R N A in estrogen-treated animals which can be
attributed to a real increase of D e receptor mRNA/cell. It is now well established that chronic estradiol treat-
ment, as initiated here, increases the density' (20 30%) of
striatal D, receptors, while leaving the affinity un- changed [7 9, 12, 19]. The modulation of D: receptors by
estradiol in anterior pituitary is more complicated and varies with dose and time of exposure to the hormone [7].
Beside these effects of estradiol on striatal D z receptor density, it was shown that this steroid hormone can also
very rapidly (within minutes or seconds) afI'ect electrical
properties of neurotransmitter systems in the basal gan-
glia, clearly distinguishing this activity from its classical
genomic site of action [13, 16]. Thus. the effect of estra- diol on D 2 receptor density in the striatum [7 9, 12, 19] is
not likely to involve a genomic site of action of this ster- oid, which may explain the lack of effect ot" estradiol on
De receptor mRNA observed here. It is a]so possible lhat
the kinetics of D3 receptor mRNA changes may be differ- ent from that of De receptor density in the striatum:
nevertheless, in the anterior pituitary, we observe D, re- ceptor mRNA changes in the same time frame.
Northern blot analysis of D, receptor m R N A levels after chronic receptor blockade with neuroleptic titiled to
show any variation of mRNA levels in either striatum
[20] or anterior pituitary [1]. The development of
postsynaptic D3 receptor supersensitivity is thus not nec- essarily reflected by a concomitant increase in the corre- sponding mRNA although lesion with 6-hydroxydopa-
mine (6-OHDA) increased De receptor mRNA levels si- multaneously with increases in the D~ receptor density of
the striatum ipsilateral to the lesion [5]. Irreversible re- ceptor blockade with the alkylatmg agent N-ethox,xcar-
bonyl-2-ethoxy- 1,2-dihydroq uinoline (EEDQ) suggests,
in fact, that the D~ receptor density increase seen after
chronic neuroleptic treatment results more from a de- crease in the rate of receptor degradation than it does
from an increase in receptor production [15]. We have also shown that chronic pretreatment with estradiol does
not affect De receptor repopulation kinetics in striatal
homogenates after EEDQ treatment [12]. Thus. it seems that D e receptor-induced supersensitivity may, in some cases, result from perturbations of such processes as membrane insertion and internalization of receptor mol- ecules, which may stabilize D, receptors in the mem- brane and cause the apparent increase of the steady state
level of De receptors [13].

In s u m m a r y , o u r f ind ings sugges t tha t the effects o f
e s t r ad io l on a n t e r i o r p i t u i t a r y D2 r ecep to r s m a y i nvo lve
t r a n s c r i p t i o n a l r egu l a t i on , w h e r e a s o u r resul ts s u p p o r t
the h y p o t h e s i s o f an a l t e r n a t i v e m e c h a n i s m o f ac t i on fo r
e s t r ad io l on s t r ia ta l r ecep to r s .
We t h a n k Dr . O l i v e r Civel l i fo r the gif t o f D2 r e c e p t o r
c D N A . T h i s w o r k was s u p p o r t e d by a M R C o f C a n a d a
g r a n t to T . D . P . D . L . is h o l d e r o f a s t u d e n t s h i p f r o m the
M R C o f C a n a d a .
1 Autelitano, D.J., Snyder, L., Sealfon, S.C. and Roberts, J.L., Dopamine D2-receptor messenger RNA is differentially regulated by dopaminergic agents in rat anterior and neurointermediate pit ui- tary, Mol. Cell. Endoerinol., 67 (1989) 101-105.
2 Borgundvaag, B. and George, S.R., Estrogen regulation of rat ante- rior pituitary adenylate cyclase, Mol. Cell. Endocrinol., 59 (1988) 35-45.
3 Boyson, S.J., McGonigle, P.M. and Molinofl, P.M., Quantitative autoradiographic localization of the D~ and D 2 subtypes of dopa- mine receptors in rat brain, J. Neurosci., 6 (1986) 3177- 3188.
4 Chirgwin, J.M., Pryzbyla, A.E., MacDonald, R.J. and Rutter, W.J.. Isolation of biologically active ribonucleic acid from sources en- riched in ribonuclease, Biochemistry, 18 (1979) 5294-5299.
5 Coirini, H., Schumacher, M., Angulo, J.A, and McEwen, B.S., In- crease in striatal dopamine D2 receptor mRNA after lesions and haloperidol treatment, Eur. J. Pharmacol., 186 (1990) 369-371.
6 Dada, M.O., Cambell, G.T. and Blake, C.A., Pars distalis cell quan- tification in normal adult male and female rats, J. Endocrinol., 101 (1984) 87 94.
7 Di Paolo, T. and Falardeau, P., Modulation of brain and pituitary dopamine receptors by estrogens and prolactin, Prog. Neuro-Psy- chopharmacol. Biol. Psychiatry, 9 (1985) 473-480.
8 Di Paolo, T., Poyet, P. and Labrie, F., Effect ofprolactin and estra- diol on rat striatal dopamine receptors, Life Sci., 31 (1982) 2921~ 2929.
9 Falardeau, P. and Di Paolo, T., Regional effect of estradiol on rat caudate putamen dopamine receptors: lateral-medial differences, Neurosci. Lett., 74 (1987) 43 48.
10 Farmer, S.R., Ben-Ze'ev, A. and Penmann, S., Regulation of actin mRNA levels and translation respond to changes in cell configura- tion, Mol. Cell. Biol., 3 (1983) 182-189.
I 1 Kramer, C.Y., Extension of multiple range tests for group means with unequal number of replications, Biometrics, 12 (1956) 307 310.
12 L6vesque, D. and Di Paolo, T., Dopamine receptor reappearance after irreversible receptor blockade: effect of chronic estradiol treat- ment of ovariectomized rats, Mol. Pharmacol:, 39 (t991) 659-665.
13 McEwen, B.S., Non-genomic and genomic effects of steroids on neural activity, Trends Pharmacol. Sci., 12 (1991) 141-147.
14 Meador-Woodruff, J.H., Manson, A., Bunzow, J.R., Van Tol. H.H.M., Stanley, S.L and Civelli, O., Distribution of D~ dopamine receptor in rat brain, Proc. Natl. Acad. Sci. U.S.A., 86 (1989) 7625. 7628.
15 Pich, E.M., Benfenati, F., Farabegoli, C., Fuxe, K., Meller, E., Arosson, M., Goldstein, M. and Agnati, L.F., Chronic haloperidot affects striatal D 2 dopamine receptor reappearance after irreversible receptor blockade, Brain Res., 435 (1987) 147 t52.
16 Schumacher, M., Rapid membrane effects of steroid hormones: an emerging concept in neuroendocrinology, Trends Neurosci., 13 (1990) 359-362.
17 Stumpf, W.E. and Sar, M., Autoradiographic techniques lbr local- izing steroid hormones, Methods Enzymol., 36 (1975) 135-156.
18 Stumpt, W.E., Sar, M. and Keefer, D.A., Atlas of estrogen target cells in the rat brain. In Anatomical Neuroendocrinology, Interna- tional Conference on Neurobiology of CNS-Hormone Interactions, Karger, Basel, 1975, pp. 104-119.
19 Van Hartesveldt, C. and Joyce, J.N., Effects of estrogen on the basal ganglia, Neurosci. Biobehav. Rev., 10 (1986) 1-14.
20 Van Tol, H.H.M., Riva, M., Civelli, O. and Creese, 1.. Lack o1 effect of chronic dopamine receptor blockade on D 2 dopamine re- ceptor mRNA level, Neurosci. Lett., 111 (1990) 303-308.
21 Weiner, D.M. and Brann, M.R., The distribution ofa dopamine D2 mRNA in rat brain, FEBS Lett., 253 (1989) 207-213.

![Regulation of dopamine neurotransmission from serotonergic ...€¦ · DA by serotonergic terminals [45, 47, 64, 65, 77]. This overwhelming exposure of the DA-depleted striatal MSNs](https://img.pdfslide.us/doc/110x75/608c24dabdeeb6661d2d50cd/regulation-of-dopamine-neurotransmission-from-serotonergic-da-by-serotonergic.jpg)


![HBM 2014- Educational Course Stimulation...VMAT DAT PET Scanning Striatal Uptake Uptake . Dopamine D2/D3 Receptor Imaging Agent in Striatum [11C]raclopride [123I]IBZM PET agent Affinity](https://img.pdfslide.us/doc/110x75/6092d384b8277237d56acd36/hbm-2014-educational-stimulation-vmat-dat-pet-scanning-striatal-uptake-uptake.jpg)


















![STRIATAL BINDING OF 2-AMINO-6,7-[3H]DIHYDROXY-1,2,3 ...for two to five individual experiments. Binding isotherms and Scatchard analyses were done using [3H]ADTN, [3H]dopamine, and](https://img.pdfslide.us/doc/110x75/60ce3f179cc6562dfb79dada/striatal-binding-of-2-amino-67-3hdihydroxy-123-for-two-to-five-individual.jpg)





