Embed Size (px)
Citation preview

of January 30, 2014.This information is current as
Receptor LociLong-Distance Relationships at Antigen
ManagingFactor, and V(D)J Recombination: Chromatin Architecture, CCCTC-Binding
Han-Yu Shih and Michael S. Krangel
http://www.jimmunol.org/content/190/10/4915doi: 10.4049/jimmunol.1300218
2013; 190:4915-4921; ;J Immunol
Referenceshttp://www.jimmunol.org/content/190/10/4915.full#ref-list-1
, 29 of which you can access for free at: cites 71 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2013 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
at U
niv of New
cstle Auchm
uty Lib/Serials Sec on January 30, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Chromatin Architecture, CCCTC-Binding Factor, and V(D)JRecombination: Managing Long-Distance Relationships atAntigen Receptor LociHan-Yu Shih and Michael S. Krangel
The rearrangement of T and B lymphocyte Ag recep-tor loci occurs within a highly complex chromosomalenvironment and is orchestrated through complexmechanisms. During the past decade, a large body ofliterature has highlighted the significance of chromatinarchitecture at Ag receptor loci in supporting the geno-mic assembly process: in preparation for recombina-tion, these loci tend to contract and form multipleloops that shorten the distances between gene segmentsand facilitate recombination events. CCCTC-bindingfactor, CTCF, has received much attention in this regardsince it has emerged as an important regulator of chro-matin organization and transcription. In this review, wesummarize recent work outlining conformational dy-namics at Ag receptor loci during lymphocyte develop-ment and we discuss the role of CTCF in Ag receptorlocus conformation and repertoire development. TheJournal of Immunology, 2013, 190: 4915–4921.
Adaptive immunity in jawed vertebrates is mediatedby T and B lymphocytes that express highly diverseand clonally distributed Ag receptors. The diversity
of lymphocyte Ag receptors is generated primarily by the as-sembly of V, D, and J region gene segments at TCR and Igloci (1). This process, known as V(D)J recombination, is cat-alyzed by the RAG-1 and RAG-2 proteins. The RAG proteinsrecognize recombination signal sequences (RSSs) that flank allV, D, and J gene segments, and with two RSSs held in a syn-aptic complex, create double-strand breaks that can be rejoinedto assemble V, D, and J gene segments with tremendouscombinatorial diversity. Ag receptor loci undergo recombina-tion in a manner that is regulated according to cell lineage anddevelopmental stage in T and B lymphocyte precursors (2, 3).During T lymphocyte development in the thymus, Tcrb, Tcrd,and Tcrg genes recombine at the CD42CD82 double-negative(DN) stage, whereas Tcra genes recombine at the CD4+CD8+
double-positive (DP) stage. B lymphocyte development in the
bone marrow is similarly characterized by developmentallystaged recombination of Ig H (Igh) and L (Igk and Igl) chaingenes, with Igh recombination in pro-B cells and Igk and Iglrecombination in pre-B cells. Recombination events are alsoregulated within individual loci; for example, D-to-J recom-bination precedes V-to-DJ recombination at both Igh andTcrb, and proximal V-to-J recombination precedes distal V-to-J recombination at Tcra (2, 3).The V(D)J recombination programs at TCR and Ig loci are
controlled at multiple levels (3–5). Most fundamental is therestriction imposed by RAG protein expression, which, exceptin special circumstances (6), is limited to developing pro- andpre-B cells and DN and DP thymocytes (7). Beyond that,the individual Ag receptor loci carry complex arrays of cis-regulatory elements, including enhancers and promoters, thatdictate regional changes to transcription and chromatin struc-ture that, in turn, allow RAG proteins to access particular RSSsat the appropriate developmental stage (3–5). In many instan-ces, functional communication between enhancers and pro-moters must occur over long distances across a landscape thatincludes many such elements, raising questions as to how thesefunctional interactions are targeted and regulated (8). Mostrecently, the spatial dynamics of Ag receptor loci withinimmature lymphocyte nuclei have also emerged as a criticalaspect of their regulation (4, 9, 10). The seminal observationrelied on three-dimensional fluorescence in situ hybridization(3D-FISH) to visualize Ig loci in cell nuclei: Igh and Igkmovedaway from the nuclear periphery, and the two ends of the Ighlocus were less separated in nuclei of pro-B cells as comparedwith other cells (11). Subsequent studies revealed “contraction”to be a general property of Ag receptor loci that occurs duringthe developmental stage in which V segments undergo recom-bination, with “decontraction” occurring subsequently (12–16). Because contraction and decontraction can be detectedeven in recombinase-deficient nuclei, they are thought to setthe stage for and to terminate long-distance recombinationevents, respectively, during lymphocyte development. BecauseRAG proteins bind preferentially to J or D-J clusters to form
Department of Immunology, Duke University Medical Center, Durham NC 27710
Received for publication January 23, 2013. Accepted for publication March 7, 2013.
This work was supported by National Institutes of Health Grant R37 GM41052 (toM.S.K.).
Address correspondence and reprint requests to Dr. Michael S. Krangel, Department ofImmunology, Duke University Medical Center, Campus Box 3010, Durham, NC27710. E-mail address: [email protected]
Abbreviations used in this article: 3C, chromosome conformation capture; 39CBE, 39CTCF-binding element; Cer, contracting element for recombination; CTCF, CCCTC-
binding factor; 3D-FISH, three-dimensional fluorescence in situ hybridization; DN,double negative; DP, double positive; Ea, Tcra enhancer; Em, intronic Igh enhancer;iEk, intronic Igk enhancer; IGCR1, intergenic control region 1; PAIR, Pax5-activatedintergenic repeat; 39RR, 39 regulatory region; RSS, recombination signal sequence; Sis,silencer in the intervening sequence; TEA, T early a; YY1, Yin-Yang 1.
Copyright� 2013 by TheAmerican Association of Immunologists, Inc. 0022-1767/13/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1300218
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

“recombination centers,” contraction and decontraction wouldmove distant V gene segments into or out of these recombina-tion centers to allow regulated assembly of a V gene repertoire (4).
Chromatin architecture, CCCTC-binding factor, and cohesin
Eukaryotic genomes are packaged at multiple levels to solvea critical “space” problem in the nucleus while simultaneouslyfacilitating transcription and replication of the DNA template(17). Recent technological advances in the analysis of genomicspatial relationships have allowed insights into the principlesof higher order chromatin organization (18). These technolo-gies are all derivative of the chromosome conformation capturetechnique (3C), in which interactingDNA fragments are initiallytrapped by chemical crosslinking; following a restriction enzymedigest and addition of DNA ligase under conditions favoringintramolecular ligation, any two interacting DNA fragments canthen be identified and quantified by PCR using oligonucleotideprimers specific for the two fragments (19). In a version of thistechnology known as 4C, circular DNA resulting from ligationof both ends of two interacting fragments is amplified by in-verse PCR using two primers in a “bait” or “viewpoint” sequence,and the entire universe of sequences interacting with the baitis determined by microarray or deep sequencing (20, 21). Ina further adaptation of this technology known as Hi-C, there isno defined bait; instead, deep sequencing is used to identify theentire genomic universe of interacting sequences (22). Duringthe last several years, 4C and Hi-C analysis has revealed segre-gation of the genome into discrete spatial compartments of upto a fewmegabases in length that correspond to domains of activeor inactive chromatin. Long-distance interactions are more fre-quent within a domain but also occur between domains, withactive chromatin domains interacting more frequently with otheractive domains, and inactive chromatin domains interactingmorefrequently with other inactive domains (20, 22–24). The datasupport a “fractal globule” model of the genome in which lo-cally looped and packaged globular units are then further as-sembled to package the entire genome.CCCTC-binding factor, CTCF, is a highly conserved, ubiq-
uitously expressed transcription factor that binds a GC-richconsensus sequence (25). CTCF has been attributed multiplefunctions, including transcriptional activation, transcriptionalrepression, and transcriptional insulation (the ability to blockan enhancer from activating a promoter when CTCF is situatedbetween the two elements). Additionally, CTCF has beenshown to function as a chromatin organizer that mediates long-distance looping interactions in the genome (25). The ability ofCTCF to mediate such loops may underlie some or all of theabove activities; CTCF-mediated loops could activate tran-scription by bringing together enhancer and promoter elementsthat are associated with nearby CTCF sites, or they could in-sulate or suppress transcription by segregating enhancers andpromoters that have intervening CTCF sites. Indeed, there aretens of thousands of CTCF-binding sites distributed in a rangeof contexts across mammalian genomes (26–30). As might beexpected for a role in insulation, the boundaries between to-pologically and functionally defined chromatin domains arehighly enriched for CTCF sites. However, only 15% of allCTCF sites are located at these boundaries (24); other CTCFsites are distributed at enhancers, promoters, and other genicand intergenic sites and mediate interactions between thesesites (24, 30–32). Recent analysis of the pro-B cell genome
indicated that long-distance interactions within chromatinspatial compartments were associated with CTCF binding,whereas those between spatial compartments were associatedwith the binding of lineage-specific transcription factors (23).Cohesin is a multisubunit protein complex that is well
known for its role in mediating sister chromatid cohesion dur-ing cell division; it is thought to functionby forming a closed ringaround the two newly replicated DNA double strands (33).Cohesin is now also appreciated to play important roles in geneexpression and chromatin organization in interphase cells:notably, it binds in a CTCF-dependent fashion to ∼70% ofCTCF-binding sites genome-wide (34–36) and is necessary forCTCF-dependent insulation and looping (37–39). Althoughin this manner CTCF and cohesin function together at manysites in the genome, they can also interact with other tran-scription factors and can function independently of one an-other (40, 41).Chromatin immunoprecipitation microarray and chroma-
tin immunoprecipitation sequencing analyses have revealedAg receptor loci to be particularly enriched in binding sites forCTCF and cohesin (26, 42–47). As a result, both proteinshave drawn attention as potential regulators of locus confor-mation, transcriptional activity, and V(D)J recombination.We focus below on recent studies of the Igh, Igk, and Tcra/Tcrd loci. Much less is known about Tcrb locus architectureand there have been no studies of the Tcrg and Igl loci.The studies of locus architecture and transcription dis-
cussed below have generally been conducted using immature cellpopulations isolated from recombinase-deficient mice, becausethis eliminates the confounding effects of changes in spatialrelationships that are a consequence of V(D)J recombination.That said, it is an assumption that the RAG proteins themselveswould not substantially impact the parameters being measured;this could be addressed by analyzing cells isolated from miceexpressing a catalytically inactive RAG protein complex (48).
Igh locus
The murine Igh locus on chromosome 12 contains nearly200 VH gene segments spanning 2.7 Mb of DNA, followedby 10–13 DH, 4 JH, and 8 CH gene segments (2) (Fig. 1A). Ighlocus V(D)J recombination occurs in a strictly ordered fash-ion, with DH-to-JH recombination preceding VH-to-DHJHrecombination in pro-B cells. The VH-to-DHJH step is furtherregulated to support allelic exclusion, as it is terminated as aconsequence of feedback inhibition in pre-B cells. Notably, eachIgh allele generally undergoes VH-to-DHJH recombination onlyonce, because this rearrangement eliminates all unrearrangedDH gene segments. Therefore, although several DH-proximalVH gene segments do rearrange at elevated frequencies, DH-distal VH gene segments must be able to compete effectivelywith these proximal VH gene segments to allow the assemblyof a diverse Igh repertoire.3D-FISH data have shown that the Igh locus contracts
specifically in pro-B cells (11, 12, 14, 49) and that this con-formation brings all VH gene segments into proximity of theDHJH cluster (13). Spatial distance measurements and com-puter modeling suggested that in its extended conformation inprepro-B cells, the Igh locus is organized into at least threerosette-like compartments, each composed of multiple DNAloops, and that these compartments merge in pro-B cells (13).Unrearranged Igh loci then appear to decontract in pre-B cells,
4916 BRIEF REVIEWS: REGULATION OF V(D)J RECOMBINATION BY LOCUS CONFORMATION
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
correlating with the suppression of VH-to-DH recombinationassociated with allelic exclusion (14). Molecular mechanismsof Igh contraction and decontraction are only partly under-stood. Transcription factors Pax5, Yin-Yang 1 (YY1), andIkaros have been shown to be essential for complete Igh locuscontraction as well as normal frequencies of distal VH rear-rangement (12, 50, 51). Because distal VH segments appear tobe “accessible” in Pax5- or YY1-deficient pro-B cells based ontheir germline transcription and histone modifications, it hasbeen inferred that distal VH-to-DHJH rearrangement relieson Igh locus contraction (12, 50). Elimination of the Ighintronic enhancer (Em), located between JH and Cm segments,also causes a loss of Igh contraction (52). YY1 may directlyregulate Igh contraction by binding to Em (50, 52) and to othersites in the VH array (53). However, it may also influence Ighcontraction indirectly by regulating the expression of Pax5(50, 53). Igh locus contraction was also reduced by CTCFknockdown, but the effect was rather modest (54). The datasuggest that multiple factors cooperate to shape the contractedconfiguration of the Igh locus in pro-B cells.The Igh locus contains 85 or more CTCF sites, most of
which are distributed across the VH region (43, 54) (Fig. 1A).CTCF sites in the proximal portion of the VH array are locatedimmediately downstream of VH RSSs, whereas those in thedistal portion are intergenic (42, 44). Included among the latterare CTCF sites at a series of 14 homologous Pax5-activated
intergenic repeat (PAIR) elements, some of which express Pax5-dependent and pro-B cell–specific antisense transcripts (43,53, 54). Only a few CTCF binding sites are found outside ofthe VH region. Two are located ∼3.2 and 5.6 kb upstream ofDFL16.1, the most 59 DH gene segment, in a region called59DFL16 or intergenic control region 1 (IGCR1); nine othersare densely arrayed in an area called the 39 CTCF-bindingelement (39CBE) that flanks the 39 regulatory region (39RR)downstream of the CH gene segments (42, 54–56). Most Ighlocus CTCF sites also bind cohesin (54). Notably, neitherCTCF nor cohesin bind to Em (52, 54).Long-distance DNA interactions across the Igh locus have
recently been probed at high resolution by 3C and 4C (52–54, 56) (Fig. 1A). These studies have described a 300-kb,multiple-loop structural domain at the 39 end of the Ighlocus, defined by interactions among IGCR1, Em, 39RR, and39CBE, that are present in both prepro-B cells and pro-B cells.The interaction between IGCR1 and 39CBE is CTCF- andcohesin-dependent (54), whereas Em interactions with IGCR1and 39CBE are CTCF- and cohesin-independent (52, 54). Eminteractions with these sites may be mediated by YY1, as thisfactor binds not only to Em, but in one study was found to bindto IGCR1 and 39CBE as well (52). Further 4C analyses con-ducted using either a proximal VH site or a distal VH site as baitdescribed two additional multiple-loop regions, each spanningseveral hundred kb. Most, but not all, interacting sequences
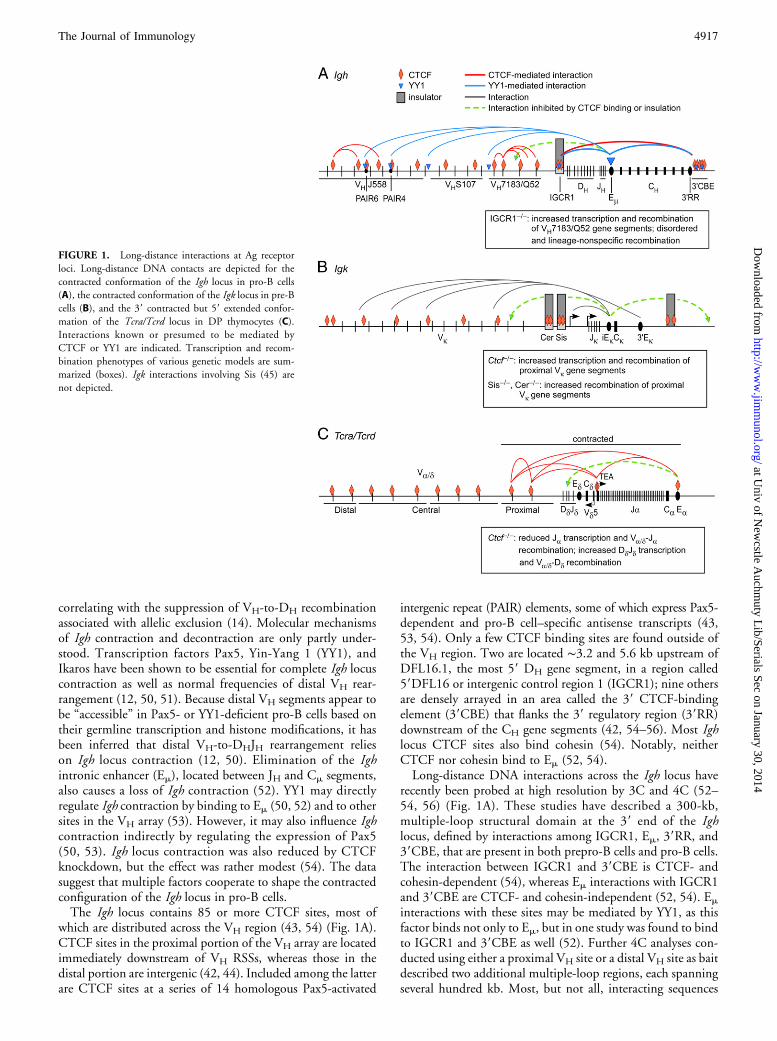
FIGURE 1. Long-distance interactions at Ag receptor
loci. Long-distance DNA contacts are depicted for the
contracted conformation of the Igh locus in pro-B cells
(A), the contracted conformation of the Igk locus in pre-B
cells (B), and the 39 contracted but 59 extended confor-
mation of the Tcra/Tcrd locus in DP thymocytes (C).
Interactions known or presumed to be mediated by
CTCF or YY1 are indicated. Transcription and recom-
bination phenotypes of various genetic models are sum-
marized (boxes). Igk interactions involving Sis (45) are
not depicted.
The Journal of Immunology 4917
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

bound CTCF; however, the CTCF dependency of theseinteractions requires further study (52). Because the twomultiple-loop regions do not interact with each other, theywere hypothesized to reside in distinct structural domains (52).Nevertheless, more comprehensive studies of long-distanceinteractions will be needed for a better picture of Igh domainstructure in prepro- and pro-B cells.In addition to the relatively local interactions described
above, Em was shown to interact with two distant sites, one inthe proximal portion of the VH array (near the 59 end of theVH7183 gene segments) and one in the distal portion of the VH
array (near the 39 end of the VHJ558 gene segments) (Fig. 1A).These long-distance interactions are Em-dependent and mayalso involve YY1, as YY1 binding was detected at both sites(52). It was also recently shown that Em interacts across 1.8–2.0Mb with two PAIR elements (PAIR4 and PAIR6) that aremajor sites of intergenic antisense transcription in the distalportion of the VH array (53). These long-distance interactionsare pro-B cell–specific and YY1-dependent; hence, they corre-late with, and may reflect, Igh locus contraction. Notably, long-distance Em–PAIR interactions do not reflect a functional rolefor Em in PAIR activation, because PAIR antisense transcriptionis Em-independent. Nevertheless, PAIR transcription, similarto Igh contraction, is YY1-dependent (53). Therefore, it wassuggested that PAIR and Em elements might interact becausethey are independently recruited into the same transcriptionfactory and that this corecruitmentmay be the basis for Igh locuscontraction (44, 53, 57). Consistent with this, knockdown ofCTCF in pro-B cells caused increased PAIR transcription andincreased PAIR–Em interactions (53, 54). However, this obser-vation is inconsistent with the notion that CTCF promotes Ighlocus contraction; the modest Igh conformational change de-tected by 3D-FISH in CTCF knockdown pro-B cells (54)might reflect more local changes in loop organization withinIgh domains, rather than decontraction per se.How do changes in CTCF-mediated loop structures impact
Igh recombination? Deletion of the 4-kb IGCR1 region ormutation of its two CTCF binding sites led to dramatic dys-regulation of the VH repertoire, with substantially elevatedtranscription and recombination of the most DH-proximalVH gene segments (56) (Fig. 1A). Remarkably, proximal VH genesegments were also found to rearrange to DH gene segmentsprior to DH-JH joining and to DH or DHJH segments even inthymocytes, and they were not subject to feedback inhibitionfrom productively assembled Igh alleles (56). Deletion of theIGCR1 region also disrupted the IGCR1–39CBE loop, as wellas Em interactions with both elements. However, because thefunctional dysregulation requires only mutation of the IGCR1CTCF binding sites, it may specifically reflect the loss of theCTCF-dependent IGCR1–39CBE loop (54, 56). The datasuggest that CTCF plays a critical role in insulating the proxi-mal VH gene segments from the influence of Em, perhaps a di-rect result of CTCF-mediated looping between the IGCR1 and39CBE regions. Consistent with this interpretation, deletion ofhs5-7, containing seven of nine CTCF sites in the 39CBE re-gion, partially reduced the IGCR1-39RR/39CBE interactionand caused a mild increase in proximal VH usage (58). More-over, a dysregulation similar to that documented for IGCR1deletion was observed for a distal VH gene segment that wasrepositioned between IGCR1 and the DH cluster on an oth-erwise wild-type Igh allele (59).
If IGCR1 functions as an insulator, it might be expectedto suppress physical interactions between Em and upstreamsites in the Igh locus. However, it is apparent that Em caninteract with numerous distant sites in the locus in pro-B cells,even in the presence of IGCR1 and an IGCR1–39CBE loop(52, 53). Therefore, the dynamics of IGCR1-mediated loopingand the basis for its impacts on long-distance DNA contacts,transcriptional activation, and V(D)J recombination will beimportant areas for future study.A note of caution that applies to the interpretation of all
studies involving knockout or knockdown of pleiotropictranscription or architectural proteins is that it is difficult toknow whether observed effects are direct (as is often assumed)or indirect. Studies involving binding site mutations, as in theexample of IGCR1, can be particularly revealing in this regard.
Igk locus
Studies of the murine Igk locus are yielding a picture rathersimilar to that of the Igh locus. The murine Igk locus onchromosome 6 contains 96 functional Vk gene segments thatare distributed across 3.2 Mb and are situated upstream of4 Jk gene segments and Ck (Fig. 1B). Although Igk alleles canundergo secondary Vk-to-Jk recombination to replace an initialVkJk rearrangement that is out-of-frame or that provokesautoreactivity, opportunities for secondary rearrangements arelimited by the small number of Jk segments. Therefore, Igkconformational features should be important to foster the de-velopment of a broad Vk repertoire by providing Jk-proximaland -distal Vk gene segments similar opportunities for rear-rangement. Based on 3D-FISH, the Igk locus was reportedto undergo contraction in pre-B cells to facilitate Vk-to-Jk re-combination at this stage (14). However, a recent Hi-C studydocumented an extensive, locus-wide network of physical inter-actions involvingVk segments and the intronic k enhancer (iEk,situated in the Jk-Ck intron) that, although absent in prepro-B cells, is apparent in pro-B cells well before the initiation ofIgk recombination (23). The presence of this interaction net-work in pro-B cells suggests that Igk contraction may occurearlier during B cell development than previously thought.The Igk locus contains ∼60 sites to which CTCF binds in
pre-B cells; binding was found to be lower in pro-B cells (42,45). Strong CTCF binding was detected at the silencer in theintervening sequence (Sis), a recombination silencer in theintergenic region between Vk and Jk segments (60, 61), atthe newly described contracting element for recombination(Cer) immediately upstream of Sis (62) and at the 59 and 39boundaries of the locus (42, 45) (Fig. 1B). Other CTCF sitesare distributed at intergenic locations across the 3.2-Mb Vk
array. A functional role for CTCF in Igk locus recombinationwas evaluated in mb1-cre Ctcff/f mice supplied with a pre-rearranged Igm transgene (45). The pre-B cell Vk repertoire inthese mice was strongly biased toward usage of proximal Vk
gene segments. Consistent with this, germline transcription ofproximal Vk gene segments was substantially increased, aswere interactions of iEk and the 39 k enhancer (39Ek) withsites distributed across the proximal 1 Mb of the Vk array(45). The enhancers also displayed increased interactions withsites outside of the Igk locus. The data suggest that CTCFplays a role in insulation, and that dysregulation of the Vk
repertoire is a consequence of hyper-activation of proximal Vk
segments by the Igk enhancers in the absence of CTCF.
4918 BRIEF REVIEWS: REGULATION OF V(D)J RECOMBINATION BY LOCUS CONFORMATION
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

Sis-deficient mice also displayed a proximally biased Vk
repertoire, indicating that the Sis CTCF site may contributeto insulation (61) (Fig. 1B). Cer deletion imparted an evenstronger proximal bias to Vk rearrangement and also allowedVk-Jk rearrangement in thymocytes (62). Thus, Cer may con-tribute to insulation as well. However, Cer (unlike Sis) deletionmodestly reduced Igk locus contraction, suggesting locusconformation as a potential explanation for the proximal Vk
bias (62). Additional experiments will be required to determinewhether the conformational change and proximal Vk bias re-flect lost binding of CTCF or of another factor to Cer, andanalysis on a recombinase-deficient background may be neededto fully evaluate effects of Sis and Cer on transcription andlong-distance Ek interactions. Although Sis and Cer have bothbeen attributed other functions (60–62), their potential rolesas insulators separating Vk segments from the Ek-regulatedJk domain appear similar to that of IGCR1 in the Igh locus.CTCF likely contributes to the network of long-distance
interactions at the Igk locus detected by Hi-C in pro-B cells(23). However, many of the described iEk interactions mappedto sites of E2A occupancy. This suggests the possibility that E2Amay drive the clustering of widely distributed Vk gene segmentswith iEk and may in this way facilitate contraction and Vk-to-Jkrearrangement (23). This possibility warrants further study.
Tcra/Tcrd locus
As compared with either Igh or Igk, the Tcra/Tcrd locus onmurine chromosome 14 (Fig. 1C) boasts a more nuancedprogram of conformational states and distinct regulatoryfunctions for CTCF. These features are thought to facilitatea complex developmental program that transitions from Tcrdgene assembly in DN thymocytes to Tcra gene assembly in DPthymocytes (3). The locus contains∼100 V gene segments thatare distributed across 1.5 Mb. A small subset of these V seg-ments rearrange to two Dd and Jd gene segments to assembleaTcrd repertoire. In contrast, most V segments can rearrange toa large array of 61 Ja gene segments to assemble a Tcra rep-ertoire.Notably, similar to Igh recombination, complete V-to-Dd-
to-Jd recombination can only occur once per allele due toelimination of Dd gene segments. In contrast, the large arraysof Va and Ja gene segments can support multiple rounds ofVa-to-Ja recombination on each allele, allowing thymocytesmultiple chances to assemble a Tcra gene that can promotepositive selection (3). It is well established that initial (primary)rearrangements in early DP thymocytes are targeted to Jasegments at the extreme 59 end of the Ja array by the activity ofthe T early a (TEA) and Ja49 promoters, and that as a func-tion of DP thymocyte lifespan, subsequent (secondary) rear-rangements are targeted to progressively more 39 Ja segmentsby the activities of the introduced Va gene segment promoters(63–66). This 59-to-39 Ja progression is coupled with an in-evitable, reciprocal 39-to-59 progression V gene segment uti-lization, but there is controversy over the extent to which thisprogression is regulated and coordinated with the Ja progres-sion. PCR analysis of thymic Va-to-Ja rearrangements hasshown that the most Ja-proximal Va gene segments are biasedto rearrange to the most 59 Ja gene segments, that the mostJa-distal Va gene segments are biased to rearrange to substan-tially more 39 Ja gene segments, and that broadly distributedmultimember Va families tend to rearrange to the entire set of
Ja gene segments (67–69). This has been interpreted as a reg-ulated and coordinated progression of Va gene segment avail-ability. Recent deep-sequencing analysis of Tcra transcripts inperipheral CD8+ T cells painted a picture that was generallyconsistent with the observations outlined above (70). However,features of the observed combinatorial diversity conflictedwith the notion of coordinated Va and Ja progressions, leadingthe authors to propose that following primary rearrangementsbetween 39 Va and 59 Ja gene segments, the remaining Va
segments simultaneously become available for recombinationto Ja gene segments (70). Further analysis may be required toconclusively resolve this issue. For example, the study discussedabove analyzed a peripheral CD8 T cell repertoire that hadbeen shaped by thymic selection (70), rather than the completespectrum of recombination events occurring in DP thymo-cytes. Moreover, additional factors, including duplicated ortriplicated portions of the V array featuring nearly identical Vgene segments, make analysis of the Tcra/Tcrd locus repertoireparticularly challenging.Analysis of Tcra/Tcrd locus conformation by 3D-FISH
revealed that, as compared with control B cells, the locus isfully contracted in DN thymocytes but undergoes 59 end ex-tension to adopt a unique 39 contracted and 59 decontractedconfiguration in DP thymocytes (16) (Fig. 1C) (although anearlier study had reached different conclusions for unknownreasons) (15). What would be the rationale for this behavior? InDN thymocytes the locus is limited to a single round of Tcrdgene recombination per allele and both proximal and distalVd gene segments serve as recombination substrates (reviewedin Ref. 71); this Igh-like behavior is best facilitated by a fullycontracted configuration of the locus. In contrast, in DP thy-mocytes the locus undergoes multiple rounds of recombinationwith primary recombination biased toward use of 39 Va and59 Ja gene segments. A 39 contracted and 59 decontracted con-figuration would favor the initial usage of 39 Va gene segmentsand thereby save more 59 Va gene segments for subsequentrounds of recombination (16). Note that the conformationsdescribed above are adopted by the unrearranged Tcra/Tcrdlocus in recombinase-deficient thymocytes. Whether and howlocus conformation in DP thymocytes adjusts to prior Tcrdgene or primary Tcra gene recombination are not known.Recent 3C studies have documented a network of interac-
tions within the 39 contracted domain of the Tcra/Tcrd locusthat forms in DP thymocytes (Fig. 1C). The Tcra enhancer(Ea) becomes active in DP thymocytes and is known to activatepromoters distributed across 500 kb (71), a region that maycorrespond to the contracted 39 domain. Included among thesepromoters are the TEA promoter associated with 59 Ja genesegments and the promoters of proximal V gene segments. Eawas shown to contact individual V and J promoters and tobring these promoters in contact with each other to forma “chromatin hub” (47). Because RAG proteins preferentiallybind to 59 Ja gene segments to form a recombination center inDP thymocytes (48), hub formation would bring proximal Vgene segments into this recombination center and would fa-cilitate the synapsis of V and J RSSs to support primary Tcrarecombination. Notably, although Ea is required to establishthis network of DNA contacts within the contracted portionof the Tcra/Tcrd locus, it is not required for 39 end contractionper se (16). This differs from the reported role for Em in Ighlocus contraction (52). CTCF also plays no role in 39 end
The Journal of Immunology 4919
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

contraction in DP thymocytes (47). Hence, the molecular basisfor Tcra/Tcrd locus contraction is unknown.As is the case for Igh and Igk, there are many binding sites
for CTCF and cohesin in the Tcra/Tcrd locus (46, 47) (Fig.1C). However, the distribution of binding sites is strikinglydifferent from Igh and Igk, because in the Tcra/Tcrd locus,CTCF sites generally mark cis-regulatory elements, includingmost V gene segment promoters, the TEA promoter, and Ea(47). Conditional knockout of Ctcf or Rad21 (which encodesa cohesin subunit) caused reduced interactions between Eaand the TEA promoter, reduced TEA transcription, and re-duced 59 Ja accessiblity (46, 47). Loss of CTCF also partiallydisrupted interactions between Ea and proximal Va gene seg-ments and between Va and Ja gene segments (47). As a result,CTCF and cohesin deficiency were associated with reducedVa-to-Ja rearrangement (46, 47). Surprisingly, CTCF defi-ciency, or deletion of the TEA promoter and its associatedCTCF site, caused increased interaction between Ea and theDdJdCd cluster and increased transcription and recombinationof Dd and Jd gene segments. Because the DdJdCd cluster lacksCTCF sites, it was suggested that CTCF normally synergizeswith Ea-bound factors to specify CTCF-marked promoters(e.g., TEA) as Ea targets, and that this suppresses Ea inter-actions with suboptimal, CTCF-free targets (e.g., Dd and Jdpromoters). With CTCF eliminated, this specificity is lost,leading to reduced Tcra gene rearrangement and increasedTcrd gene rearrangement in DP thymocytes (47). CTCF-dependent long-distance interactions are thought to serve pri-marily a targeting function at the Tcra/Tcrd locus, rather than aninsulating function as at Igh and Igk, because Tcra/Tcrd locusCTCF sites are positioned at, rather than between, critical cis-regulatory elements.
ConclusionsAg receptor locus conformation is manipulated at multiplelevels to support gene assembly by V(D)J recombination. Theevidence suggests at least two layers of organization: multiple-loop rosette-like structures span hundreds of kb, and inter-actions between such structures mediate global spatial rela-tionships. Long-distance interaction networks appear to beorchestrated by ubiquitous and lineage-specific transcriptionfactors (YY1, Pax5, and E2A) as well as chromatin architecturalproteins (CTCF and cohesin). However, there appears not tobe a single paradigm that fits all Ag receptor loci. CTCF-mediated looping can suppress or stimulate V(D)J recombi-nation through effects on RSS accessibility, depending onwhether those loops insulate enhancers from promoters (e.g.,Igh and Igk) or target enhancers to promoters (e.g., Tcra).CTCF-mediated looping may also regulate V(D)J recombi-nation by influencing RSS synapsis. Synapsis may be facili-tated when RSSs are positioned near cis-regulatory elementsbrought into contact by CTCF (e.g., Tcra). Synapsis was alsoshown to be inhibited when accessible RSSs are segregatedinto different chromatin loops by CTCF (72), although thisresult is difficult to reconcile with the obvious need for RSSsynapsis between loops at Ag receptor loci. It seems clear thatthe development of Ag receptor repertoires must occur as anexceedingly complex function of conformational states andspatial relationships dictated by the distribution of bindingsites for CTCF and other factors. It will take additional timeand effort to make sense of it all.
DisclosuresThe authors have no financial conflicts of interest.
References1. Schatz, D. G., and P. C. Swanson. 2011. V(D)J recombination: mechanisms of
initiation. Annu. Rev. Genet. 45: 167–202.2. Jung, D., C. Giallourakis, R. Mostoslavsky, and F. W. Alt. 2006. Mechanism and
control of V(D)J recombination at the immunoglobulin heavy chain locus. Annu.Rev. Immunol. 24: 541–570.
3. Krangel, M. S. 2009. Mechanics of T cell receptor gene rearrangement. Curr. Opin.Immunol. 21: 133–139.
4. Schatz, D. G., and Y. Ji. 2011. Recombination centres and the orchestration of V(D)J recombination. Nat. Rev. Immunol. 11: 251–263.
5. Osipovich, O., and E. M. Oltz. 2010. Regulation of antigen receptor gene assemblyby genetic-epigenetic crosstalk. Semin. Immunol. 22: 313–322.
6. Hale, J. S., K. T. Ames, T. E. Boursalian, and P. J. Fink. 2010. Cutting edge: Ragdeletion in peripheral T cells blocks TCR revision. J. Immunol. 184: 5964–5968.
7. Kuo, T. C., and M. S. Schlissel. 2009. Mechanisms controlling expression of theRAG locus during lymphocyte development. Curr. Opin. Immunol. 21: 173–178.
8. Krivega, I., and A. Dean. 2012. Enhancer and promoter interactions-long distancecalls. Curr. Opin. Genet. Dev. 22: 79–85.
9. Jhunjhunwala, S., M. C. van Zelm, M. M. Peak, and C. Murre. 2009. Chromatinarchitecture and the generation of antigen receptor diversity. Cell 138: 435–448.
10. Bossen, C., R. Mansson, and C. Murre. 2012. Chromatin topology and the regu-lation of antigen receptor assembly. Annu. Rev. Immunol. 30: 337–356.
11. Kosak, S. T., J. A. Skok, K. L. Medina, R. Riblet, M. M. Le Beau, A. G. Fisher, andH. Singh. 2002. Subnuclear compartmentalization of immunoglobulin lociduring lymphocyte development. Science 296: 158–162.
12. Fuxa, M., J. Skok, A. Souabni, G. Salvagiotto, E. Roldan, and M. Busslinger. 2004.Pax5 induces V-to-DJ rearrangements and locus contraction of the immunoglobulinheavy-chain gene. Genes Dev. 18: 411–422.
13. Jhunjhunwala, S., M. C. van Zelm, M. M. Peak, S. Cutchin, R. Riblet, J. J. vanDongen, F. G. Grosveld, T. A. Knoch, and C. Murre. 2008. The 3D structure ofthe immunoglobulin heavy-chain locus: implications for long-range genomic inter-actions. Cell 133: 265–279.
14. Roldan, E., M. Fuxa, W. Chong, D. Martinez, M. Novatchkova, M. Busslinger, andJ. A. Skok. 2005. Locus ‘decontraction’ and centromeric recruitment contribute toallelic exclusion of the immunoglobulin heavy-chain gene. Nat. Immunol. 6: 31–41.
15. Skok, J. A., R. Gisler, M. Novatchkova, D. Farmer, W. de Laat, and M. Busslinger.2007. Reversible contraction by looping of the Tcra and Tcrb loci in rearrangingthymocytes. Nat. Immunol. 8: 378–387.
16. Shih, H. Y., andM. S. Krangel. 2010. Distinct contracted conformations of the Tcra/Tcrd locus during Tcra and Tcrd recombination. J. Exp. Med. 207: 1835–1841.
17. Hubner, M. R., M. A. Eckersley-Maslin, and D. L. Spector. 2012. Chromatinorganization and transcriptional regulation. Curr. Opin. Genet. Dev. DOI: 10.1016/j.gde.2012.11.006.
18. de Wit, E., and W. de Laat. 2012. A decade of 3C technologies: insights intonuclear organization. Genes Dev. 26: 11–24.
19. Dekker, J., K. Rippe, M. Dekker, and N. Kleckner. 2002. Capturing chromosomeconformation. Science 295: 1306–1311.
20. Simonis, M., P. Klous, E. Splinter, Y. Moshkin, R. Willemsen, E. de Wit, B. vanSteensel, and W. de Laat. 2006. Nuclear organization of active and inactive chro-matin domains uncovered by chromosome conformation capture-on-chip (4C).Nat. Genet. 38: 1348–1354.
21. Zhao, Z., G. Tavoosidana, M. Sjolinder, A. Gondor, P. Mariano, S. Wang,C. Kanduri, M. Lezcano, K. S. Sandhu, U. Singh, et al. 2006. Circular chromosomeconformation capture (4C) uncovers extensive networks of epigenetically regulatedintra- and interchromosomal interactions. Nat. Genet. 38: 1341–1347.
22. Lieberman-Aiden, E., N. L. van Berkum, L. Williams, M. Imakaev, T. Ragoczy,A. Telling, I. Amit, B. R. Lajoie, P. J. Sabo, M. O. Dorschner, et al. 2009.Comprehensive mapping of long-range interactions reveals folding principles of thehuman genome. Science 326: 289–293.
23. Lin, Y. C., C. Benner, R. Mansson, S. Heinz, K. Miyazaki, M. Miyazaki,V. Chandra, C. Bossen, C. K. Glass, and C. Murre. 2012. Global changes in thenuclear positioning of genes and intra- and interdomain genomic interactions thatorchestrate B cell fate. Nat. Immunol. 13: 1196–1204.
24. Dixon, J. R., S. Selvaraj, F. Yue, A. Kim, Y. Li, Y. Shen, M. Hu, J. S. Liu, andB. Ren. 2012. Topological domains in mammalian genomes identified by analysisof chromatin interactions. Nature 485: 376–380.
25. Phillips, J. E., and V. G. Corces. 2009. CTCF: master weaver of the genome. Cell137: 1194–1211.
26. Kim, T. H., Z. K. Abdullaev, A. D. Smith, K. A. Ching, D. I. Loukinov,R. D. Green, M. Q. Zhang, V. V. Lobanenkov, and B. Ren. 2007. Analysis of thevertebrate insulator protein CTCF-binding sites in the human genome. Cell 128:1231–1245.
27. Cuddapah, S., R. Jothi, D. E. Schones, T. Y. Roh, K. Cui, and K. Zhao. 2009.Global analysis of the insulator binding protein CTCF in chromatin barrier regionsreveals demarcation of active and repressive domains. Genome Res. 19: 24–32.
28. Jothi, R., S. Cuddapah, A. Barski, K. Cui, and K. Zhao. 2008. Genome-wideidentification of in vivo protein-DNA binding sites from ChIP-Seq data. NucleicAcids Res. 36: 5221–5231.
29. Xie, X., T. S. Mikkelsen, A. Gnirke, K. Lindblad-Toh, M. Kellis, and E. S. Lander.2007. Systematic discovery of regulatory motifs in conserved regions of the humangenome, including thousands of CTCF insulator sites. Proc. Natl. Acad. Sci. USA104: 7145–7150.
4920 BRIEF REVIEWS: REGULATION OF V(D)J RECOMBINATION BY LOCUS CONFORMATION
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

30. Handoko, L., H. Xu, G. Li, C. Y. Ngan, E. Chew, M. Schnapp, C. W. Lee, C. Ye,J. L. Ping, F. Mulawadi, et al. 2011. CTCF-mediated functional chromatin inter-actome in pluripotent cells. Nat. Genet. 43: 630–638.
31. Sanyal, A., B. R. Lajoie, G. Jain, and J. Dekker. 2012. The long-range interactionlandscape of gene promoters. Nature 489: 109–113.
32. Shen, Y., F. Yue, D. F. McCleary, Z. Ye, L. Edsall, S. Kuan, U. Wagner, J. Dixon,L. Lee, V. V. Lobanenkov, and B. Ren. 2012. A map of the cis-regulatory sequencesin the mouse genome. Nature 488: 116–120.
33. Wood, A. J., A. F. Severson, and B. J. Meyer. 2010. Condensin and cohesincomplexity: the expanding repertoire of functions. Nat. Rev. Genet. 11: 391–404.
34. Parelho, V., S. Hadjur, M. Spivakov, M. Leleu, S. Sauer, H. C. Gregson, A. Jarmuz,C. Canzonetta, Z. Webster, T. Nesterova, et al. 2008. Cohesins functionally asso-ciate with CTCF on mammalian chromosome arms. Cell 132: 422–433.
35. Wendt, K. S., K. Yoshida, T. Itoh, M. Bando, B. Koch, E. Schirghuber,S. Tsutsumi, G. Nagae, K. Ishihara, T. Mishiro, et al. 2008. Cohesin mediatestranscriptional insulation by CCCTC-binding factor. Nature 451: 796–801.
36. Rubio, E. D., D. J. Reiss, P. L. Welcsh, C. M. Disteche, G. N. Filippova,N. S. Baliga, R. Aebersold, J. A. Ranish, and A. Krumm. 2008. CTCF physicallylinks cohesin to chromatin. Proc. Natl. Acad. Sci. USA 105: 8309–8314.
37. Hou, C., R. Dale, and A. Dean. 2010. Cell type specificity of chromatin organi-zation mediated by CTCF and cohesin. Proc. Natl. Acad. Sci. USA 107: 3651–3656.
38. Nativio, R., K. S. Wendt, Y. Ito, J. E. Huddleston, S. Uribe-Lewis, K. Woodfine,C. Krueger, W. Reik, J. M. Peters, and A. Murrell. 2009. Cohesin is required forhigher-order chromatin conformation at the imprinted IGF2-H19 locus. PLoSGenet. 5: e1000739.
39. Chien, R., W. Zeng, S. Kawauchi, M. A. Bender, R. Santos, H. C. Gregson,J. A. Schmiesing, D. A. Newkirk, X. Kong, A. R. Ball, Jr., et al. 2011. Cohesinmediates chromatin interactions that regulate mammalian b-globin expression. J.Biol. Chem. 286: 17870–17878.
40. Schmidt, D., P. C. Schwalie, C. S. Ross-Innes, A. Hurtado, G. D. Brown,J. S. Carroll, P. Flicek, and D. T. Odom. 2010. A CTCF-independent role forcohesin in tissue-specific transcription. Genome Res. 20: 578–588.
41. Kagey,M. H., J. J. Newman, S. Bilodeau, Y. Zhan, D. A. Orlando, N. L. van Berkum,C. C. Ebmeier, J. Goossens, P. B. Rahl, S. S. Levine, et al. 2010. Mediator and cohesinconnect gene expression and chromatin architecture. Nature 467: 430–435.
42. Degner, S. C., T. P. Wong, G. Jankevicius, and A. J. Feeney. 2009. Cutting edge:developmental stage-specific recruitment of cohesin to CTCF sites throughoutimmunoglobulin loci during B lymphocyte development. J. Immunol. 182: 44–48.
43. Ebert, A., S. McManus, H. Tagoh, J. Medvedovic, G. Salvagiotto, M. Novatchkova,I. Tamir, A. Sommer, M. Jaritz, and M. Busslinger. 2011. The distal V(H) genecluster of the Igh locus contains distinct regulatory elements with Pax5 transcriptionfactor-dependent activity in pro-B cells. Immunity 34: 175–187.
44. Lucas, J. S., C. Bossen, and C. Murre. 2011. Transcription and recombinationfactories: common features? Curr. Opin. Cell Biol. 23: 318–324.
45. Ribeiro de Almeida, C., R. Stadhouders, M. J. de Bruijn, I. M. Bergen,S. Thongjuea, B. Lenhard, W. van Ijcken, F. Grosveld, N. Galjart, E. Soler, andR. W. Hendriks. 2011. The DNA-binding protein CTCF limits proximal Vk re-combination and restricts k enhancer interactions to the immunoglobulin k lightchain locus. Immunity 35: 501–513.
46. Seitan, V. C., B. Hao, K. Tachibana-Konwalski, T. Lavagnolli, H. Mira-Bontenbal,K. E. Brown, G. Teng, T. Carroll, A. Terry, K. Horan, et al. 2011. A role forcohesin in T-cell-receptor rearrangement and thymocyte differentiation. Nature476: 467–471.
47. Shih, H. Y., J. Verma-Gaur, A. Torkamani, A. J. Feeney, N. Galjart, andM. S. Krangel. 2012. Tcra gene recombination is supported by a Tcra enhancer- andCTCF-dependent chromatin hub. Proc. Natl. Acad. Sci. USA 109: E3493–E3502.
48. Ji, Y., W. Resch, E. Corbett, A. Yamane, R. Casellas, and D. G. Schatz. 2010. The invivo pattern of binding of RAG1 andRAG2 to antigen receptor loci.Cell 141: 419–431.
49. Sayegh, C. E., S. Jhunjhunwala, R. Riblet, and C. Murre. 2005. Visualization oflooping involving the immunoglobulin heavy-chain locus in developing B cells.Genes Dev. 19: 322–327.
50. Liu, H., M. Schmidt-Supprian, Y. Shi, E. Hobeika, N. Barteneva, H. Jumaa,R. Pelanda, M. Reth, J. Skok, K. Rajewsky, and Y. Shi. 2007. Yin Yang 1 is a criticalregulator of B-cell development. Genes Dev. 21: 1179–1189.
51. Reynaud, D., I. A. Demarco, K. L. Reddy, H. Schjerven, E. Bertolino, Z. Chen,S. T. Smale, S. Winandy, and H. Singh. 2008. Regulation of B cell fate commit-ment and immunoglobulin heavy-chain gene rearrangements by Ikaros. Nat.Immunol. 9: 927–936.
52. Guo, C., T. Gerasimova, H. Hao, I. Ivanova, T. Chakraborty, R. Selimyan,E. M. Oltz, and R. Sen. 2011. Two forms of loops generate the chromatin con-formation of the immunoglobulin heavy-chain gene locus. Cell 147: 332–343.
53. Verma-Gaur, J., A. Torkamani, L. Schaffer, S. R. Head, N. J. Schork, andA. J. Feeney. 2012. Noncoding transcription within the Igh distal VH region atPAIR elements affects the 3D structure of the Igh locus in pro-B cells. Proc. Natl.Acad. Sci. USA 109: 17004–17009.
54. Degner, S. C., J. Verma-Gaur, T. P. Wong, C. Bossen, G. M. Iverson,A. Torkamani, C. Vettermann, Y. C. Lin, Z. Ju, D. Schulz, et al. 2011. CCCTC-binding factor (CTCF) and cohesin influence the genomic architecture of the Ighlocus and antisense transcription in pro-B cells. Proc. Natl. Acad. Sci. USA 108:9566–9571.
55. Garrett, F. E., A. V. Emelyanov, M. A. Sepulveda, P. Flanagan, S. Volpi, F. Li,D. Loukinov, L. A. Eckhardt, V. V. Lobanenkov, and B. K. Birshtein. 2005.Chromatin architecture near a potential 39 end of the igh locus involves modularregulation of histone modifications during B-cell development and in vivo occu-pancy at CTCF sites. Mol. Cell. Biol. 25: 1511–1525.
56. Guo, C., H. S. Yoon, A. Franklin, S. Jain, A. Ebert, H. L. Cheng, E. Hansen,O. Despo, C. Bossen, C. Vettermann, et al. 2011. CTCF-binding elements mediatecontrol of V(D)J recombination. Nature 477: 424–430.
57. Marenduzzo, D., I. Faro-Trindade, and P. R. Cook. 2007. What are the molecularties that maintain genomic loops? Trends Genet. 23: 126–133.
58. Volpi, S. A., J. Verma-Gaur, R. Hassan, Z. Ju, S. Roa, S. Chatterjee, U. Werling,H. Hou, Jr., B. Will, U. Steidl, et al. 2012. Germline deletion of Igh 39 regulatoryregion elements hs 5, 6, 7 (hs5–7) affects B cell-specific regulation, rearrangement,and insulation of the Igh locus. J. Immunol. 188: 2556–2566.
59. Bates, J. G., D. Cado, H. Nolla, and M. S. Schlissel. 2007. Chromosomal positionof a VH gene segment determines its activation and inactivation as a substrate for V(D)J recombination. J. Exp. Med. 204: 3247–3256.
60. Liu, Z., P. Widlak, Y. Zou, F. Xiao, M. Oh, S. Li, M. Y. Chang, J. W. Shay, andW. T. Garrard. 2006. A recombination silencer that specifies heterochromatinpositioning and ikaros association in the immunoglobulin k locus. Immunity 24:405–415.
61. Xiang, Y., X. Zhou, S. L. Hewitt, J. A. Skok, and W. T. Garrard. 2011. A multi-functional element in the mouse Igk locus that specifies repertoire and Ig locisubnuclear location. J. Immunol. 186: 5356–5366.
62. Xiang, Y., S. K. Park, and W. T. Garrard. 2013. Vk gene repertoire and locuscontraction are specified by critical DNase I hypersensitive sites within the Vk-Jkintervening region. J. Immunol. 190: 1819–1826.
63. Guo, J., A. Hawwari, H. Li, Z. Sun, S. K. Mahanta, D. R. Littman, M. S. Krangel,and Y. W. He. 2002. Regulation of the TCRa repertoire by the survival window ofCD4+CD8+ thymocytes. Nat. Immunol. 3: 469–476.
64. Hawwari, A., and M. S. Krangel. 2007. Role for rearranged variable gene segmentsin directing secondary T cell receptor a recombination. Proc. Natl. Acad. Sci. USA104: 903–907.
65. Mauvieux, L., I. Villey, and J. P. de Villartay. 2001. T early a (TEA) regulatesinitial TCRVAJA rearrangements and leads to TCRJA coincidence. Eur. J. Immu-nol. 31: 2080–2086.
66. Villey, I., D. Caillol, F. Selz, P. Ferrier, and J. P. de Villartay. 1996. Defectin rearrangement of the most 59 TCR-J a following targeted deletion of Tearly a (TEA): implications for TCR a locus accessibility. Immunity 5: 331–342.
67. Pasqual, N., M. Gallagher, C. Aude-Garcia, M. Loiodice, F. Thuderoz,J. Demongeot, R. Ceredig, P. N. Marche, and E. Jouvin-Marche. 2002. Quanti-tative and qualitative changes in V-J a rearrangements during mouse thymocytesdifferentiation: implication for a limited T cell receptor a chain repertoire. J. Exp.Med. 196: 1163–1173.
68. Thuderoz, F., M. A. Simonet, O. Hansen, N. Pasqual, A. Dariz, T. P. Baum,V. Hierle, J. Demongeot, P. N. Marche, and E. Jouvin-Marche. 2010. Numericalmodelling of the V-J combinations of the T cell receptor TRA/TRD locus. PLOSComput. Biol. 6: e1000682.
69. Jouvin-Marche, E., P. Fuschiotti, and P. N. Marche. 2009. Dynamic aspects ofTCRa gene recombination: qualitative and quantitative assessments of the TCRachain repertoire in man and mouse. Adv. Exp. Med. Biol. 650: 82–92.
70. Genolet, R., B. J. Stevenson, L. Farinelli, M. Osteras, and I. F. Luescher. 2012.Highly diverse TCRa chain repertoire of pre-immune CD8+ T cells reveals newinsights in gene recombination. EMBO J. 31: 1666–1678.
71. Hawwari, A., and M. S. Krangel. 2005. Regulation of TCR d and a repertoires bylocal and long-distance control of variable gene segment chromatin structure. J. Exp.Med. 202: 467–472.
72. Shrimali, S., S. Srivastava, G. Varma, A. Grinberg, K. Pfeifer, and M. Srivastava.2012. An ectopic CTCF-dependent transcriptional insulator influences the choice ofVb gene segments for VDJ recombination at TCRb locus. Nucleic Acids Res. 40:7753–7765.
The Journal of Immunology 4921
at Univ of N
ewcstle A
uchmuty L
ib/Serials Sec on January 30, 2014http://w
ww
.jimm
unol.org/D
ownloaded from





























