Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Mar. 1986, p. 849-855 Vol. 165, No. 30021-9193/86/030849-07$02.00/0Copyright X) 1986, American Society for Microbiology
Choline-Glycine Betaine Pathway Confers a High Level of OsmoticTolerance in Escherichia coliBJARNE LANDFALD AND ARNE R. STR0M*
Institute of Fisheries, University of Tromso, N-9001 Troms0, Norway
Received 6 August 1985/Accepted 14 November 1985
Glycine betaine and its precursors choline and glycine betaine aldehyde have been found to confer a highlevel of osmotic tolerance when added exogenously to cultures of Escherichia -coli at an inhibitory osmoticstrength. In this paper, the following findings are described. Choline works as an osmoprotectant only underaerobic conditions, whereas glycine betaine aldehyde and glycine betaine function both aerobically andanaerobically. No endogenous glycine betaine accumulation was detectable in osmotically stressed cells grownin the absence of the osmoprotectant itself or the precursors. A membrane-bound, 02-dependent, and electrontransfer-linked dehydrogenase *as found which oxidized choline to glycine betaine aldehyde and aldehyde toglycine betaine at nearly the same rate. It displayed Michaelis-Menten kinetics; the apparent Km values forcholine and glycine betaine aldehyde were 1.5 and 1.6 mM, respectively. Also, a soluble, NAD-dependentdehydrogenase oxidized glycine betaine aldehyde. It displayed Michaelis-Menten kinetics; the apparent Kmvalues for the aldehyde, NAD, and NADP were 0.13, 0.06, and 0.5 mM, respectively. The ch4iline-glycinebetaine pathway was osmotically regulated, i.e., full enzymic activities were found only in cells grownaerobically in choline-containing medium at an elevated osmotic strength. Chloramnphenicol inhibited theformation of the pathway in osmotically stressed cells.
Under osmotic stress, bacteria common'ly accumulatepotassium ions and organic solutes in their cytoplasnm tobuild up internal osmotic strength and prevent diffusion ofwater out of the cells. Stewart (33) pointed out that theorganic solutes used by bacteria comprise only a few typesof compounds, namely, amino acids, sugars, polyols, andbetaines (i.e., fully N-methylated amino acid derivatives).The term osmoprotectant has been coined for moleculeswhich can be accumulated in large amounts in the cell andprotect against osmotic stress and salt inactivation (18, 34).
Escherichia coli displays high versatility in synthesis anduptake of osmoprotectants, and, depending on the composi-tion of the growth medium, it accumulates most classes ofthe compoun-ds listed -above (2, 17, 21, 27, 30). However,betaines, such as glyc'ine betaine (and its precursors), prolinebetaine (stachydrine'), and y-butyrobetaine, and, to a lesserextent, proline are the only compounds found which stimu-late the growth -of E. coli and other enteric bacteria atinhibitory osmolalities when added in low concentrations(e.g., ca. 1 mM) to the growth medium (7, 16, 18, 19, 34).Glycine betaine also stimulates the respiration of a moder-ately halophilic, heterotrophic bacterium, Ba1, at a highNaCl concentration (32), and it accumulates in thehalophific, photosynthetic bacterium Ectothiorhodospirahalochloris (8) and in several hypersaline strains ofcyanobacteria (20, 29). Thus, betaines appear to play animportant role in the osmoregulation of various bacteria withdifferent ranges of osmotic tolerance, as they do in nmanymarine animals and halophytic plants (3, 38, 39F).
Choline is a precursor of glycine betaine in osmoticallystressed cells of E. coli (34) and strain Ba1 (28). The catabolicoxidation of choline has been described in several bacteria(11, 14, 24, 26, 31), but no details are known about Thisoxidation process in re-lation to bacterial osmoregulation.Here we report that the choline-glycine betaine pathway ofE. coli is osmotically regulated, i.e., only cells grown in
* Corresponding author.
choline-containing medium with an increased osmoticstrength display full enzymic activity.
MATERIALS AND METHODS
Bacterial strain and growth conditions. E. coli K10 was-obtained from R. C. Valentine ofthe University of California,Davis. The organism was grown in medium 63 (22) containing100 mM KH2PO4, 75 mM KOH, 15 mM (NH4)2S04, 1 mMMgSO4, 3.9 puM FeSO4, and 22 mM D-glucose. Half-strengthmedium 63 contained half the concentration of all theinorganic ingredients pluls 22 mM D-glucose. The osmoticstrength of the medium was increased by the addition of 0.1to 0.8 M NaCl, KCl, or sucrose, and 1 mM choline, glycinebetaine aldehyde, or glycine betaine was added as statedbelow. In experiments with chloramphenicol, 200 p.g per mlwas used. The pH was adjusted to 7.2 with NaOH whennecessary. The cells were grown at 37°C on a rotary shaker.Aerobic growth took place in flasks with baffles, -andanaerobic growth took place in rubber-stoppered flasks underan N2 atmosphere. Cells for inoculum were grown in medium63 with no additions. Cells used in enzyme studies weregrown until the optical density at 420 nm reached 4 to 6 inaerobic cultures and about 2 in anaerobic cultures. They werethen harvested by centrifugation at 8,000 x g for 15 min (4C),washed once in half of the original volume of 0.1 M sodiumphosphate (pH 7.5) in the-centrifuge, and used immediatelyor stored at -80°C in the same buffer.
Synthesis of '4C-labeled and unlabeled glycine *betainealdehyde. Unlabeled compound was prepared by a reactionbetween anhydrous trimethylamine and bromoacetaldehydediethyl acetal (Merck-Schuchardt) as -described by Bergel etal. (1), except for the following modifications. The reactionwas performed in toluene, and trimethylamine (Merck-Schuchardt) was extracted into the toluene phase from a30% (wt/vol) aqueous solution by the slow -addition of KOH{about 10 M final concentration) and careful shaking. Theorganic phase was dried by -the addition of excess solidsodium sulfate. The glycine betaine-aldehyde product was
-849
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.

850 LANDFALD AND STR0M
recrystallized as described previously (13), and its identitywas confirmed by proton nuclear magnetic resonance spec-troscopy (1H-NMR; see below). Labeled compound wasprepared by the same method starting with 2.2 g of['4C]trimethylamine (specific activity, 5.6 nCi/,umol), and theradiochemical purity of the product was confirmed by paperelectrophoresis by using 0.75 M formic acid and Whatman3MM filter paper and applying 20 V/cm.Enzymatic synthesis of ['4C]glycine betaine. Enzymatic
synthesis of [14C]glycine betaine was performed essentiallyas described previously (27). The reaction mixture con-tained, in a 0.2-ml volume, 2 ,uCi [methyl-14C]choline (spe-cific activity, 52 RCi/pimol), 10 ,umol of sodium phosphate(pH 8.0), and 6 U of choline oxidase (from Alcaligenes sp.;Sigma Chemical Co., St. Louis, Mo.). The mixture wasincubated overnight at 370C, and the glycine betaine formedwas separated from choline and glycine betaine aldehyde ona Dowex microcolumn as described below for the NAD-dependent glycine betaine-aldehyde dehydrogenase assay.The eluate was evaporated to dryness in a vacuum to removeNH3, and the product was dissolved in distilled water anddiluted to a specific activity of 15 nCi/,umol with unlabeledglycine betaine (Sigma). The radiochemical purity of theproduct was controlled by paper electrophoresis as de-scribed above.
Accumulation of glycine betaine in cells. The cells weregrown in medium 63 until the optical density at 420 nmreached 2.5, and then a 20% inoculum was transferred to thesame medium with a final concentration of 0.4 or 0.65 MNaCl and 1 mM [14C]choline (15 nCi/,umol), [14C]glycinebetaine aldehyde (5.6 nCi/4mol), or [14C]glycine betaine (15nCi/,Lmol). The cultures were incubated for at least twogenerations under aerobic or anaerobic conditions as de-scribed above. Then 1 ml was transferred to an Eppendorftube and centrifuged for 1 min at 8,000 x g, after which0.1-ml samples were rapidly withdrawn from the top fractionof the supernatant. Uptake of radioactive material by thecells was determined by the decrease in the radioactivity ofthe medium during bacterial growth. The identity of labeledmaterial accumulated in the cells was determined by extract-ing the cells with 0.35 M perchloric acid and subjecting theextract to chromatography as described for the cholinedehydrogenase assay (see below) and to paper electropho-resis as described for [14C]glycine betaine-aldehyde synthe-sis. The possibility of incorporation of labeled compoundsinto insoluble material was checked by repeatedly washingthe pellet from the perchloric acid extraction in the centri-fuge, dissolving it in Lumasolve (Lumac Systems, Basel,Switzerland), and subjecting it to liquid scintillation count-ing. Identification of intracellular glycine betaine by 'H-NMR spectroscopy (see below) was done on cells grownaerobically in medium 63 containing 0.65 M NaCl in thepresence of 1 mM unlabeled choline, glycine betainealdehyde, or glycine betaine, or in medium 63 with 0.5 MNaCI in the absence of osmoprotectant. The cells (25 to 90mg of protein) were collected by centrifugation at 8,000 x gfor 15 min and then extracted twice in 2 ml of 5% trichloro-acetic acid. The combined extracts were twice evaporated todryness and redissolved in ca. 1 ml of deuterium oxide.
Toluene treatment and freezing of cells. Toluene (0.2 ml)was added to a suspension of cells (20 to 40 mg of protein) in2 ml of ice-cold 0.1 M sodium phosphate buffer (pH 7.5). Themixture was vortexed for 1 min, 8 ml of buffer was added,and then it was centrifuged at 6,000 x g for 5 min. The cellswere suspended and washed twice in 10 ml of buffer in thecentrifuge, and finally they were suspended in 1 to 2 ml ofbuffer with 30 ,ug each of DNase and RNase added per ml.
For practical reasons, the cells were freeze stored beforetoluene treatment unless otherwise stated. This resulted inan approximately 20% reduction in both enzyme activitiescompared with those of freshly harvested cells. The cholinedehydrogenase activity was always assayed immediatelyafter the toluene treatment, whereas glycine betaine-aldehyde dehydrogenase was in most cases assayed after thetoluene-treated cells had been stored for at least 1 day at-80°C to inactivate the former dehydrogenase. This proce-dure had essentially no influence on the glycine betaine-aldehyde dehydrogenase activity.
Disruption and centrifugal fractionation of cells. Suspen-sions of freshly harvested and washed cells (about 50 mg ofprotein in 3.5 ml) in disruption buffer (0.1 M sodium phos-phate [pH 7.5], 1 mM dithiothreitol) containing 30 ,ug each ofDNase and RNase per ml were forced twice through aminiature French pressure cell (American Instrument Co.,Silver Spring, Md.) operated at 1,000 bars. Unbroken cellsand large fragments were removed by centrifugation at12,000 x g for 15 min. The supernatant (crude extract) wasdiluted threefold with disruption buffer and then centrifugedat 180,000 x g for 3 h. The high-speed supernatant wascollected, and the membrane fraction was suspended in thesame volume of disruption buffer and collected by centrifu-gation at 180,000 x g for 0.5 h. The pellet was suspended in2 ml of the buffer. The washed membranes were dissolved bythe addition of 0.3% (vol/vol) Triton X-100 (BDH Chemnicals,Poole, England) and stirring for 20 min. Undissolved mate-rial was removed by centrifugation at 40,000 x g for 20 min.All operations were performed at 0 to 4°C.
Choline dehydrogenase assay. The standard reaction mix-ture for the determination of the oxidation of choline toglycine betaine aldehyde contained, in a 1-ml volume, 10,umol of [14C]choline (specific activity, 15 nCi/,lmol), 40,umol of sodium bisulfite (grade I; Sigma; the pH of the stocksolution was adjusted to 7.5 with NaGH), 80 ,umol of sodiumphosphate (pH 7.5), and 1 to 10 mg of cell protein. Whenenzyme solubilized with Triton X-100 was tested, 0.5 ,umolof phenazine methosulfate (Sigma) was included, and so-dium bisulfite was omitted. The reaction mixture was rou-tinely incubated aerobically with shaking at 37°C. Anaerobicassays were performed in rubber-stoppered vessels repeat-edly evacuated and filled with N2, after which 50 ,umol ofglucose, 1.6 mg of glucose oxidase (G-2133; Sigma), and 10nmol of the redox indicator methylene blue were added tothe reaction mixture.At intervals up to 9 min, 200-,I samples were withdrawn,
and the reaction was stopped by the addition of 80 ,ul of 0.7M perchloric acid containing 1 mM each unlabeled glycinebetaine aldehyde and glycine betaine. Then 100 lil of 1 Msodium bisulfite was added, and the mixture was centri-fuged. The sodium bisulfite of the reaction mixture combinedwith [14C]glycine betaine aldehyde formed in the reaction(13) and prevented a further oxidation -of this intermediatemetabolite. (A concentration of 40 mM sodium bisulfite hadno effect on the choline-oxidizing activity.) The resultingaddition product was separated from choline by ion-exchange chromatography. A deproteinized sample (100 ,ul)was applied to a column (0.5 by 4 cm) packed with Dowex-50x4-200 on H' form (Sigma). The radioactive glycinebetaine aldehyde-bisulfite addition product was eluted with 2ml of 0.2 M sodium bisulfite (pH 7.0) and determined byliquid scintillation counting. Under these conditions, cholineremained on the column. In experiments in which sodiumbisulfite was omitted from the reaction mixture, this chemi-cal was added after the incubation period, and the deprotein-ized sample was applied to the column as described above.
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.
OSMOREGULATION IN E. COLI 851
44 8 2 1 8 1 6 2
0
~~~~0
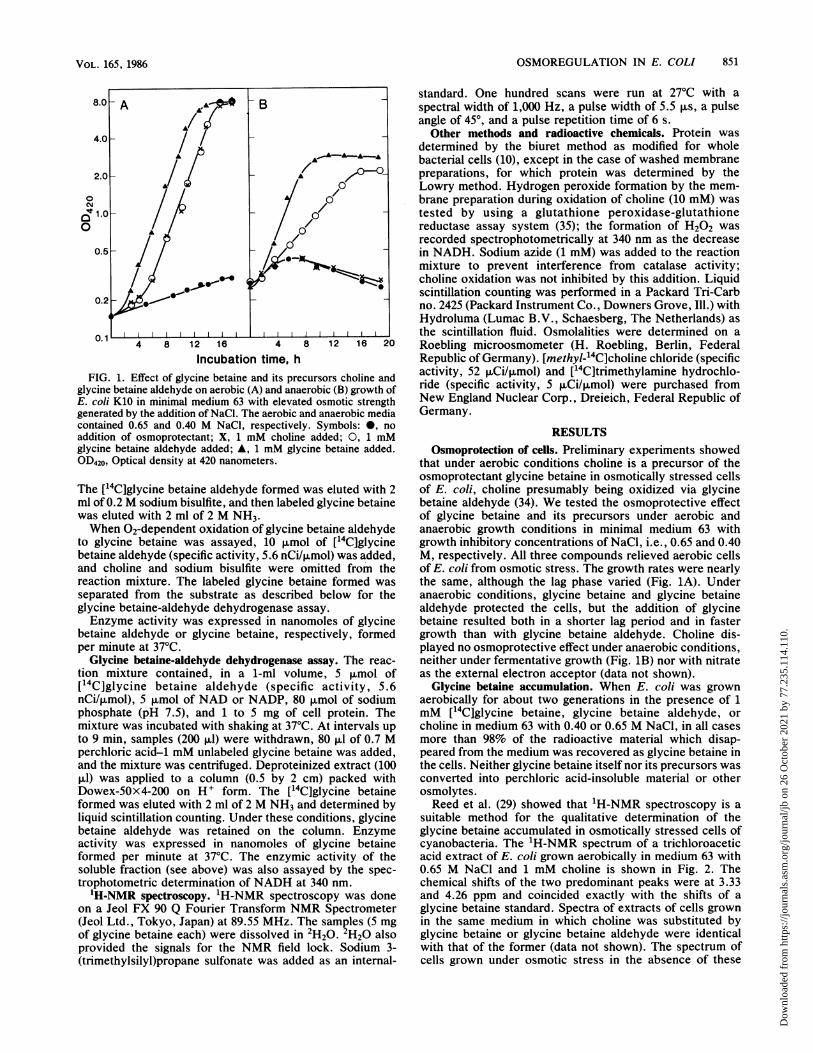
Incubation time, hFIG. 1. Effect of glycine betaine and its precursors choline and
glycine betaine aldehyde on aerobic (A) and anaerobic (B) growth ofE. coli K10 in minimal medium 63 with elevated osmotic strengthgenerated by the addition of NaCl. The aerobic and anaerobic mediacontained 0.65 and 0.40 M NaCl, respectively. Symbols:0, noaddition of osmoprotectant; X, 1 mM choline added; O, 1 mMglycine betaine aldehyde added; n,1 mM glycine betaine added.OD420, Optical density at 420 nanometers.
The [4C]glycine betaine aldehyde formed was eluted with 2ml of 0.2 M sodium bisulfite, and then labeled glycine betainewas eluted with 2 ml of 2 M NH3eWhen o2-dependent oxidation of glycine betaine aldehyde
to glycine betaine was assayed, 10mMugl of [ a4C]glycinebetaine aldehyde (specific activity, 5.6nCi/tmol)was added2and choline and sodium bisulfite were omitted from thereaction mixture. The labeled glycine betaine formed wasseparated from the substrate as described below for theglycine betaine-aldehyde dehydrogenase assay.Enzyme activity was expressed in nanomoles of glycine
betaine aldehyde or glycine betaine, respectively, formedper minute at 37sC.
Glycine betaine-aldehyde dehydrogenase assay. The reac-tion mixture contained, in a 1-ml volume, 5 ,umol of[14C]glycine betaine aldehyde (specific activity, 5.6nCi/4mol), 5 ,umol of NAD or NADP, 80 ,umol of sodiumphosphate (pH 7.5), and 1 to 5 mg of cell protein. Themixture was incubated with shaking at 37°C. At intervals up
to 9 min, samples (200 RI) were withdrawn, 80 ptl of 0.7 Mperchloric acid-1 mM unlabeled glycine betaine was added,and the mixture was centrifuged. Deproteinized extract (100RI) was applied to a column (0.5 by 2 cm) packed withDowex-50x4-200 on H+ form. The [14C]glycine betaineformed was eluted with 2 ml of 2 M NH3 and determined byliquid scintillation counting. Under these conditions, glycinebetaine aldehyde was retained on the column. Enzymeactivity was expressed in nanomoles of glycine betaineformed per minute at 37°C. The enzymic activity of thesoluble fraction (see above) was also assayed by the spec-
trophotometric determination of NADH at 340 nm.
'H-NMR spectroscopy. 1H-NMR spectroscopy was doneon a Jeol FX 90 Q Fourier Transform NMR Spectrometer(Jeol Ltd., Tokyo, Japan) at 89.55 MHz. The samples (5 mgof glycine betaine each) were dissolved in 2H20. 2H20 alsoprovided the signals for the NMR field lock. Sodium 3-(trimethylsilyl)propane sulfonate was added as an internal-
standard. One hundred scans were run at 27°C with aspectral width of 1,000 Hz, a pulse width of 5.5 ,us, a pulseangle of 450, and a pulse repetition time of 6 s.
Other methods and radioactive chemicals. Protein wasdetermined by the biuret method as modified for wholebacterial cells (10), except in the case of washed membranepreparations, for which protein was determined by theLowry method. Hydrogen peroxide formation by the mem-brane preparation during oxidation of choline (10 mM) wastested by using a glutathione peroxidase-glutathionereductase assay system (35); the formation of H202 wasrecorded spectrophotometrically at 340 nm as the decreasein NADH. Sodium azide (1 mM) was added to the reactionmixture to prevent interference from catalase activity;choline oxidation was not inhibited by this addition. Liquidscintillation counting was performed in a Packard Tri-Carbno. 2425 (Packard Instrument Co., Downers Grove, Ill.) withHydroluma (Lumac B.V., Schaesberg, The Netherlands) asthe scintillation fluid. Osmolalities were determined on aRoebling microosmometer (H. Roebling, Berlin, FederalRepublic of Germany). [methyl-"'C]choline chloride (specificactivity, 52 ,uCi/,Lmol) and ["4C]trimethylamine hydrochlo-ride (specific activity, 5 pRCi/l4mol) were purchased fromNew England Nuclear Corp., Dreieich, Federal Republic ofGermany.
RESULTSOsmoprotection of cells. Preliminary experiments showed
that under aerobic conditions choline is a precursor of theosmoprotectant glycine betaine in osmotically stressed cellsof E. coli, choline presumably being oxidized via glycinebetaine aldehyde (34). We tested the osmoprotective effectof glycine betaine and its precursors under aerobic andanaerobic growth conditions in minimal medium 63 withgrowth inhibitory concentrations of NaCl, i.e., 0.65 and 0.40M, respectively. All three compounds relieved aerobic cellsof E. coli from osmotic stress. The growth rates were nearlythe same, although the lag phase varied (Fig. 1A). Underanaerobic conditions, glycine betaine and glycine betainealdehyde protected the cells, but the addition of glycinebetaine resulted both in a shorter lag period and in fastergrowth than with glycine betaine aldehyde. Choline dis-played no osmoprotective effect under anaerobic conditions,neither under fermentative growth (Fig. 1B) nor with nitrateas the external electron acceptor (data not shown).
Glycine betaine accumulation. When E. coli was grownaerobically for about two generations in the presence of 1mM [14C]glycine betaine, glycine betaine aldehyde, orcholine in medium 63 with 0.40 or 0.65 M NaCl, in all casesmore than 98% of the radioactive material which disap-peared from the medium was recovered as glycine betaine inthe cells. Neither glycine betaine itself nor its precursors wasconverted into perchloric acid-insoluble material or otherosmolytes.Reed et al. (29) showed that 1H-NMR spectroscopy is a
suitable method for the qualitative determination of theglycine betaine accumulated in osmotically stressed cells ofcyanobacteria. The 'H-NMR spectrum of a trichloroaceticacid extract of E. coli grown aerobically in medium 63 with0.65 M NaCl and 1 mM choline is shown in Fig. 2. Thechemical shifts of the two predominant peaks were at 3.33and 4.26 ppm and coincided exactly with the shifts of aglycine betaine standard. Spectra of extracts of cells grownin the same medium in which choline was substituted byglycine betaine or glycine betaine aldehyde were identicalwith that of the former (data not shown). The spectrum ofcells grown under osmotic stress in the absence of these
VOL. 165, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.

852 LANDFALD AND STR0M
osmoprotectants did not give any detectable signals ofglycine betaine (Fig. 2). By adding glycine betaine as an
internal standard to the latter extract, we showed thatamounts as low as 1 ,g/mg of cell protein could be detectedwith 1H-NMR spectroscopy; the latter amount correspondsto an intracellular concentration of about 4 mM (27). Thus,there was no endogenous glycine betaine synthesis andaccumulation in E. coli deprived of glycine betaine or itsprecursors.As shown before (18, 27), the present radiochemical assay
revealed that the glycine betaine accumulation of cellsincreased with the NaCl concentration of the medium (Table1). This accumulation was independent of whether the cellswere supplied with the ready-made osmoticum or one of theprecursors. For cells supplied with glycine betaine or glycinebetaine aldehyde, the glycine betaine accumulation was thesame under aerobic and anaerobic growth conditions.Choline was not accumulated or converted by anaerobicallygrown cells. The present figures for glycine betaine accumu-lation (Table 1) are considerably higher than those reportedpreviously for stressed cells of E. coli incubated for a shorttime (i.e., 30 min) with glycine betaine, but they are compa-rable to data reported previously for cells actively growingwith glycine betaine for several generations (27).
calization and properties of enzymes. Radiochemicalassays were developed which allowed independent measure-ments of the oxidation of choline to glycine betaine aldehydeand of the aldehyde to glycine betaine, catalyzed by cellfractions and toluene-treated cells. Typical enzyme activitiesfound in freshly harvested cells induced by growth in me-dium 63 with 0.40 M NaCl and 1 mM choline are shown inTable 2. The washed membrane fraction prepared by high-speed centrifugation ofE. coli disrupted in a French pressurecell catalyzed cofactor-independent oxidation of both sub-strates at about the same rate. The oxidation processes were02 dependent, i.e., they were completely abolished byanaerobic incubation conditions. The same enzyme (called
A
B.. ..I.... I....I ....I.... I....I.....I ... L.. .. I....!
4 3 2 1 0
PPMFIG. 2. 1H-NMR spectra of trichloroacetic acid extracts of E.
coli K10 grown aerobically in medium 63 with elevated osmoticstrength generated with 0.65 M NaCl and containing 1 mM choline(trace A) and with 0.50 M NaCl but without added osmoprotectant(trace B).
TABLE 1. Glycine betaine accumulation in osmotically stressedcells of E. coli K1'a
Osmotic AccumulationGrowth strength of of glycinecondition medium Osmoprotectant/precursor betaineb
(osmolal) (,zmol mg ofprotein-')Aerobic 1.01 Choline 0.93 ± 0.02
Glycine betaine aldehyde 0.85 ± 0.07Glycine betaine 0.85 ± 0.06
Anaerobic 1.01 Choline 0Glycine betaine aldehyde 0.80 ± 0.06Glycine betaine 0.90 ± 0.12
Aerobic 1.47 Choline 1.75 ± 0.08Glycine betaine aldehyde 1.64 ± 0.09Glycine betaine 1.78 ± 0.05
a The cells were grown aerobically or anaerobically for two generations inthe presence of 1 mM '4C-labeled choline, glycine betaine aldehyde, or glycinebetaine in medium 63 containing 0.40 or 0.65 M NaCI (i.e., 1.01 or 1.47osmolal, respectively).
b Average of three independent determinations ± standard error.
choline dehydrogenase) seemed to catalyze both reactions,because the oxidation of both '4C-labeled substrates (10 mM)was inhibited nearly 50%o in the presence of an equimolarconcentration of the other unlabeled substrate. This notionwas supported by genetic evidence, in that all mutants foundwith defective choline dehydrogenase activity also lack the02-dependent glycine betaine aldehyde-oxidizing activity(34a).The reaction had a sharp pH optimum at 7.5 in sodium
phosphate and displayed Michaelis-Menten kinetics; theapparent Km values for choline and glycine betaine aldehydewere 1.5 and 1.6 mM, respectively. No detectable amount ofhydrogen peroxide was formed in the reactions. When themembrane fraction was treated with 0.3% Triton X-100, the02-dependent activity was destroyed; however, the solubil-ized enzyme catalyzed the oxidation of both substrates in thepresence of the artificial electron acceptor phenazinemethosulfate (Table 2). These findings indicated that theenzyme was an electron transfer-dependent dehydrogenase.It should also be noted that cells made penetrable to cholineby toluene treatment retained higher specific 02-dependent
TABLE 2. Specific activities of the choline and the glycinebetaine-aldehyde dehydrogenasea
Choline Glycine betaine-dehydrogenaseb aldehydePrepn (nmol min-' mg dehydrogenasecPrepn ~~(nmol min1 mg ( .omilmof protein-) (nmol min- mg
Toluene-treated cells 21 100
Cell-free prepnsCrude extract 8.4 110Soluble fraction 0 110Washed membrane fraction 57 (53) 0Solubilized membranes 49 (49) 0
a1The activities of harvested E. coli K1O cells grown in medium 63 with 1mM choline and 0.40M NaCI were assayed in toluene-treated cells and in cellsdisrupted in a French pressure cell.
b Figures in parentheses represent activity with [14CJglycine betainealdehyde as the substrate. Solubilized membranes were assayed withphenazine methosulfate (0.5 mM) as artificial electron acceptor.
c Assayed in the presence of NAD (5 mM) and corrected for NAD-independent activity in fractions containing both dehydrogenases.
J. BACTERIOL.
J-.--
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.

OSMOREGULATION IN E. COLI 853
NAD(P) NAD(P)H
+H20\ / GYNE
CHOLINE GLYCINEBETAINE-
TGLYCINE-ALDEHYDE +H2O [2H]
[,_[2H]
1/202 H20 1/202 H20
FIG. 3. The choline-glycine betaine pathway of E. coli K10.
enzyme activity than crude extracts of cells disrupted in aFrench pressure cell.The oxidation of glycine betaine aldehyde was also
catalyzed by a soluble NAD-dependent dehydrogenase (Ta-ble 2). The enzyme had a broad pH optimum from 7.5 to 9.5in sodium phosphate and Tris hydrochloride. It displayedMichaelis-Menten kinetics; the apparent Km values ofglycine betaine aldehyde, NAD, and NADP were 0.13, 0.06,and 0.5 mM, respectively. When comparing the spectropho-tometric determination of NADH formation with theradiochemical determination of glycine betaine formation,we found a ratio of 0.98 to 1; i.e., 1 to 1 within experimentalaccuracy. The enzyme did not oxidize choline. The choline-glycine betaine pathway of E. coli revealed in this work isshown in Fig. 3.A comparison of activities of the dehydrogenases in cells
and their affinities for the substrates indicated that the02-dependent, membrane-bound dehydrogenase functionedmainly in choline oxidation and that the NAD-dependentdehydrogenase prevented intracellular accumulation of thealdehyde. Choline oxidation seemed to be the rate-limitingstep of the pathway (Table 2). The accumulation of theobserved amount of 1.7 ,umol of glycine betaine per mg ofcell protein (Table 1) in E. coli grown in medium 63 with 0.65M NaCl at a rate of 0.52 generations per h (Fig. 1) wouldrequire a choline-oxidizing activity of 15 nmollmin per mg ofcell protein. This is in the same range as the observed in vitrocholine dehydrogenase activity of osmotically stressed cells(Table 2; Fig. 4).Osmotic regulation of dehydrogenase activities. E. coli
grown in half-strength (0.15 osmolal) or full-strength (0.28osmolal) medium 63 with 1 mM choline displayed lowactivities of the choline and the glycine betaine-aldehydedehydrogenases (Fig. 4). However, when the osmoticstrength of the medium was increased by the addition ofNaCl, these enzymic activities were increased in the cells. Amarked stimulation was observed even with a modest incre-ment of the osmotic strength to 0.46 osmolal (0.1 M NaCladded), and peak activities were observed in cells grown ina medium with osmotic strength above 1.0 osmolal (0.40 MNaCl added). The activity of the choline and the glycinebetaine-aldehyde dehydrogenases was stimulated up to30-and 10-fold, respectively, compared with activity inunstressed cells. Within the range of experimental errors,cells grown in choline-containing medium with KCl orsucrose had the same dehydrogenase activities as cellsgrown with NaCl added to the same osmotic strength (Table3). Thus, the pathway was modulated by the external os-motic pressure, not by a specific ion, in accordance with theglycine betaine uptake system in this organism (27).
In the absence of choline, an increase in the osmolality ofthe growth medium gave a much lower but significantincrease in the activities of both the dehydrogenases (Fig. 4).Cells grown in half-strength medium 63 with 50 mM choline
displayed an approximately fourfold increase in these activ-ities compared with background levels, whereas the pres-ence of 10 mM choline had essentially no effect (Table 4).This stimulation by a high choline content was not anosmotic effect, because addition of 50 mM sucrose in thepresence of 1 mM choline had no effect. Furthermore, theosmolalities of all media based on half-strength medium 63were well below that of full-strength medium 63, which didnot induce enzyme formation (Fig. 4).The presence of 1 mM glycine betaine aldehyde seemed to
stimulate slightly the choline dehydrogenase activity inosmotically stressed cells, compared with when noosmoprotectant was added, but it had no significant influ-
NaCI added, M
Ec
E
0
S-
OL
._
75
N
Cw,;
1V
Osmotic strength, osmolalFIG. 4. Effect of osmotic strength and the presence of choline or
glycine betaine aldehyde in the growth medium on choline dehydro-genase (A) and glycine betaine-aldehyde dehydrogenase (B) activi-ties of E. coli K10. The lower abscissa shows the osmotic strengthof the growth medium, and the upper abscissa shows the amounts ofNaCi added to medium 63; one point indicated below zero additionof NaCl (i.e., 0.15 osmolal) represents half-strength medium 63. Theenzymic activities were assayed in toluene-treated cells, and eachpoint represents the average of three independent measurements.Vertical bars show the standard errors. Symbols: 0, no addition ofosmoprotectant; *, 1 mM choline added; A, 1 mM glycine betainealdehyde added.
VOL. 165, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.

854 LANDFALD AND STR0M
ence on the glycine betaine-aldehyde dehydrogenase activity(Fig. 4).
Cells grown anaerobically in medium 63 with 0.20 or 0.40M NaCl and with choline, glycine betaine aldehyde, or thetwo osmolytes combined (1 mM each) showed no increase inthe enzymic activities. The background activity of theglycine betaine-aldehyde dehydrogenase (about 5 nmol/minper mg of protein) was, however, high enough to explain theslow growth and the glycine betaine formation observed inanaerobic culture with 0.40 M NaCl and 1 mM glycinebetaine aldehyde (Fig. 1 and Table 1).To further investigate the mechanism of osmotic modula-
tion of the enzymic activities, E. coli was grown in medium63 without salt and then harvested and suspended in medium63 with 0.40 M NaCI and 1 mM choline with or withoutchloramphenicol. In medium without the antibiotic, a sub-stantial increase in both enzymic activities could be detectedwithin 20 min, and 60 min after the upshock in osmolality,the cells displayed 50% of the enzymic activities of fullyinduced cells. During this period, the optical density of theculture increased 9%. In the presence of chloramphenicol(200 ,xg/ml), the enzymic activities remained at the back-ground level. Thus, the appearance of both dehydrogenaseactivities in osmotically stressed cells apparently dependedon protein synthesis.
DISCUSSIONOur data confirmed that glycine betaine is an osmoprotec-
tive compound in E. coli K10 and that choline and glycinebetaine aldehyde serve as its precursors in osmoticallystressed cells (18, 34). Cells which were deprived of glycinebetaine or the known precursors did not accumulate thisosmoprotectant. Thus, glycine betaine recently reported toaccumulate in E. coli during osmotic stress (12) is likely tooriginate from glycine betaine or choline components of theyeast extract or the Proteose Peptone of the growth mediumrather than from a complete de novo synthesis by a yetunknown biosynthetic pathway. The ability for catabolicoxidation of choline seems widespread in bacteria (14), butE. coli cannot utilize any of the metabolites of the choline-glycine betaine pathway as a substrate or building blocks forhigh-molecular-weight cell components. The osmotic modu-lation of this pathway in E. coli reflects that it functions onlyin osmoregulation. However, the present dehydrogenasesresemble those involved in the catabolism of choline inPseudomonas aeruginosa (24, 25). Also, in mammalian livercells the oxidation of choline to glycine betaine is catalyzedby two enzymes (37): a mitochondrial electron transfer-linked choline dehydrogenase (EC 1.1.99.1) and a soluble
TABLE 3. Specific activities of the choline and the glycinebetaine-aldehyde dehydrogenase of E. coli K10 cells grown in
choline containing medium 63 with various solutes added
Choline Glycineaddedhyrogenaseb betaine-aldehyde
Solute added Concna (M) dehydrogenaseb(nmol min- mg (nmol min-' mgof protein-') of protein-')
None 0.5 9NaCl 0.4 15 83KCI 0.4 14 83Sucrose 0.6 13 69a Solute concentrations were selected to give the same osmotic strength (1.0
osmolal) in media with additions.b The enzymic activities were assayed in toluene-treated cells.
TABLE 4. Specific activities of the choline dehydrogenase andthe glycine betaine-aldehyde dehydrogenase of E. coli K10 cellsgrown in half-strength medium 63 with various concentrations of
choline
CholineAdditio of dehCholine GlycineCholineAddition of d dro enasea betaine-aldehydeCholn
sucrose (50 .yg dehydrogenaseaconcn mm M) mm) (nmol min' mg (nmol min-' mgmM) ~~of protein- ') of protein-')
1 - 0.4 91 + 0.4 8
10 - 0.6 1350 - 1.6 35
a The enzymic activities were assayed in toluene-treated cells.
NAD-dependent (glycine) betaine-aldehyde dehydrogenase(EC 1.2.1.8). The mitochondrial choline dehydrogenasecatalyzes glycine betaine aldehyde oxidation at only 5% ofthe speed of choline oxidation (36), whereas the presentenzyme catalyzed both reactions equally well. In Ar-throbacter globiformis (11) and an Alcaligenes strain (26),there exists a soluble, hydrogen peroxide-forming oxidasewhich oxidizes both substrates. The former organism alsohas a soluble glycine betaine-aldehyde dehydrogenase.The control mechanism of the choline-glycine betaine
pathway of E. coli appears to be complex. The formation ofthe present dehydrogenase activities in osmotically stressedcells was prevented by chloramphenicol, indicating that theosmotic modulation of the pathway represents a de novoprotein synthesis rather than an activation of precursors.Growth of E. coli in a medium with low osmotic strength andhigh choline content (Table 4) and in a medium with highosmotic strength and no choline content (Fig. 4) resulted ina partial activation of the pathway. These findings indicatethat the expression of the system requires signals on both theexternal osmotic pressure and the internal choline content.The requirement for aerobic growth conditions for inductionof the pathway points to the existence of an additional redoxcontrol.High osmotic pressure is known to induce several trans-
port systems for cellular solutes in enteric bacteria, e.g., theKdp system for K+ (15), and the ProU (6) and ProP systemswhich are shown to transport glycine betaine (4, 5). Theosmotic induction of transport systems for glycine betainedoes not require the presence of the substrate (4, 27), but ifavailable it accumulates in response to external osmoticpressure (17, 27). Based on the present and previous find-ings, the following model has emerged for the sequence ofosmotic induction of the choline-glycine betaine pathway.High external osmotic pressure induces the transport sys-tems for the precursor choline (34a), and the resulting uptakeof choline further provokes the full expression of the path-way, presumably by an activation at the transcriptionallevel.
Presumably, these series of events are controlled bymembrane-bound proteins which sense changes in externalosmotic pressure. Two such osmotic sensor proteins havebeen identified in E. coli, namely, the KdpD protein whichosmoregulates the expression of the high-affinity K+ uptakesystem (15) and the EnvZ protein which osmoregulates theproportionate amounts of two major outer membrane pro-teins, OmpF and OmpC (9). The latter system is regulatedthrough a cascade of reactions which involve both transcrip-tional and translational control (9, 23). Thus, even in aprocaryotic organism the adaptation to changes in the os-
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.

OSMOREGULATION IN E. COLI 855
motic pressure of the environment involves a great numberof sensor proteins, transport proteins, and enzyme systems.
ACKNOWLEDGMENTSThis work was supported by a grant from the Norwegian Research
Council of Fisheries.We thank R. C. Valentine (University of California, Davis) and D.
Le Rudulier (Universite de Rennes) for useful discussions. We alsothank P. I. Larsen for his help in running the NMR spectra and B.Couzens for typing the manuscript.
LITERATURE CITED1. Bergel, F., A. Cohen, and N. C. Hindley. 1950. Syntheses of
neurotropic and musculotropic stimulators and inhibitors. PartIII. Derivatives of unsaturated w-betaines. J. Chem. Soc.1950:1439-1443.
2. Britten, R. J., and F. T. McClure. 1962. The amino acid pool inEscherichia coli. Bacteriol. Rev. 26:292-335.
3. Burton, R. F. 1983. The composition of animal cells: solutescontributing to osmotic pressure and charge balance. Comp.Biochem. Physiol. B Comp. Biochem. 76:663-671.
4. Cairney, J., I. R. Booth, and C. F. Higgins. 1985. Osmoregula-tion of gene expression in Salmonella typhimurium: proUencodes an osmotically induced betaine transport system. J.Bacteriol. 164:1224-1232.
5. Cairney, J., I. R. Booth, and C. F. Higgins. 1985. Salmonellatyphimurium proP gene encodes a transport system for theosmoprotectant betaine. J. Bacteriol. 164:1218-1223.
6. Csonka, L. N. 1982. A third L-proline permease in Salmonellatyphimurium which functions in media of elevated osmoticstrength. J. Bacteriol. 151:1433-1443.
7. Dhavises, G., and G. D. Anagnostopoulos. 1979. Influence ofamino acids and water activity on the growth of Escherichia coliB/r/1. Microbios Lett. 7:105-115.
8. Galinski, E. A., and H. G. Truper. 1982. Betaine, a compatiblesolute in the extremely halophilic phototrophic bacterium Ecto-thiorhodospira halochloris. FEMS Microbiol. Lett. 13:357-360.
9. Hall, M. N., and T. J. Silhavy. 1981. Genetic analysis of themajor outer membrane proteins of Escherichia coli. Annu. Rev.Genet. 15:91-142.
10. Herbert, D., P. J. Phipps, and R. E. Strange. 1971. Chemicalanalysis of microbial cells, p. 244-249. In J. R. Norris and D. W.Ribbons (ed.), Methods in microbiology, vol. SB. AcademicPress, Inc., New York.
11. Ikuta, S., K. Matuura, S. Imamura, H. Misaki, and Y. Horiuti.1977. Oxidative pathway of choline to betaine in the solublefraction prepared from Arthrobacter globiformis. J. Biochem.82:157-163.
12. Imhoff, J.F., and F. Rodriguez-Valera. 1984. Betaine is the maincompatible solute of halophilic eubacteria. J. Bacteriol.160:478-479.
13. Jellinek, M., D. R. Strength, and S. A. Thayer. 1959. Isolationand identification of the products of the oxidation of choline. J.Biol. Chem. 234:1171-1173.
14. Kortstee, G. J. J. 1970. The aerobic decomposition of choline bymicroorganisms. I. The ability of aerobic organisms, particu-larly coryneform bacteria, to utilize choline as the sole carbonand nitrogen source. Arch. Mikrobiol. 71:235-244.
15. Laimins, L. A., D. B. Rhoads, and W. Epstein. 1981. Osmoticcontrol of kdp operon expression in Escherichia coli. Proc.Natl. Acad. Sci. USA 78:464 468.
16. Le Rudulier, D., T. Bernard, G. Goas, and J. Hamelin. 1984.Osmoregulation in Klebsiella pneumoniae: enhancement ofanaerobic growth and nitrogen fixation under stress by prolinebetaine, -y-butyrobetaine, and other related compounds. Can. J.Microbiol. 30:299-305.
17. Le Rudulier, D., and L. Bouillard. 1983. Glycine betaine, an
osmotic effector in Klebsiella pneumoniae and other membersof the Enterobacteriaceae. Appl. Environ. Microbiol. 46:152-159.
18. Le Rudulier, D., A. R. Str0m, A. M. Dandekar, L. T. Smith, and
R. C. Valentine. 1984. Molecular biology of osmoregulation.Science 224:1064-1068.
19. Le Rudulier, D., and R. C. Valentine. 1982. Genetic engineeringin agriculture: osmoregulation. Trends Biochem. Sci. 7:431-433.
20. Mackay, M. A., R. S. Norton, and L. J. Borowitzka. 1984.Organic osmoregulatory solutes in cyanobacteria. J. Gen. Mi-crobiol. 130:2177-2191.
21. Measures, J. C. 1975. Role of amino acids in osmoregulation ofnon-halophilic bacteria. Nature (London) 257:398-400.
22. Miller, J. H. 1972. Experiments in molecular genetics, p. 431.Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
23. Mizuno, T., M.-Y. Chou, and M. Inouye. 1984. A uniquemechanism regulating gene expression: translational inhibitionby a complementary RNA transcript (micRNA). Proc. Natl.Acad. Sci. USA 81:1966-1970.
24. Nagasawa, T., Y. Kawabata, Y. Tani, and K. Ogata. 1975.Choline dehydrogenase of Pseudomonas aeruginosa A-16.Agric. Biol. Chem. 39:1513-1514.
25. Nagasawa, T., Y. Kawabata, Y. Tani, and K. Ogata. 1976.Purification and characterization of betaine aldehyde dehydro-genase from Pseudomonas aeruginosa A-16. Agric. Biol. Chem.40:1743-1749.
26. Ohta-Fukuyama, M., Y. Miyake, S. Emi, and T. Yamano. 1980.Identification and properties of the prosthetic group of cholineoxidase from Alcaligenes sp. J. Biochem. 88:197-203.
27. Perroud, B., and D. Le Rudulier. 1985. Glycine betaine trans-port in Escherichia coli: osmotic modulation. J. Bacteriol.161:393-401.
28. Rafaeli-Eshkol, D. 1968. Studies on halotolerance in a moder-ately halophilic bacterium. Biochem. J. 109:679-685.
29. Reed, R. H., J. A. Chudek, R. Foster, and W. D. P. Stewart.1984. Osmotic adjustment in cyanobacteria from hypersalineenvironments. Arch. Microbiol. 138:333-337.
30. Roller, S. D., and G. D. Anagnostopoulos. 1982. Accumulation ofcarbohydrate by Escherichia coli B/r/1 during growth at lowwater activity. Appl. Microbiol. 52:425-434.
31. Shieh, H. S. 1964. Aerobic degradation of choline. I. Fermen-tation of choline by a marine bacterium, Achromobactercholinophagum n. sp. Can. J. Microbiol. 10:837-842.
32. Shkedy-Vinkler, C., and Y. Avi-Dor. 1975. Betaine-inducedstimulation of respiration at high osmolarities in a halotolerantbacterium. Biochem. J. 150:219-226.
33. Stewart, W. D. P. 1983. Natural environments-challenges tomicrobial success and survival. Symp. Soc. Geh. Microbiol.34:1-35.
34. Str0m, A. R., D. Le Rudulier, M. W. Jakowec, R. C. Bunnell,and R. C. Valentine. 1983. Osmoregulatory (Osm) genes andosmoprotective compounds, p. 39-59. In T. Kosuge, C.Meredith, and A. Hollaender (ed.), Genetic engineering ofplants. Plenum Publishing Corp., New York.
34a.Styrvold, 0. B., P. Falkenberg, B. Landfald, M. W. Eshoo, T.Bj0rnsen, and A. R. Str0m. 1986. Selection, mapping, andcharacterization of osmoregulatory mutants of Escherichia coliblocked in the choline-glycine betaine pathway. J. Bacteriol.165:856-863.
35. Tappel, A. L. 1978. Glutathione peroxidase and hydroperox-ides. Methods Enzymol. 52:506-513.
36. Tsuge, H., Y. Nakano, H. Onishi, Y. Futamura, and K. Ohashi.1980. A novel purification and some properties of rat livermitochondrial choline dehydrogenase. Biochim. Biophys. Acta614:274-284.
37. Wilken, D. R., M. L. McMacken, and A. Rodriquez. 1970.Choline and betaine aldehyde oxidation by rat liver mito-chondria. Biochim. Biophys. Acta 216:305-317.
38. Wyn Jones, R. G., and R. Storey. 1981. Betaines, p. 171-204. InL. G. Paleg and D. Aspinall (ed.), The physiology and biochem-istry of drought resistance in plants. Academic Press Australia,Sidney, Australia.
39. Yancey, P. H., M. E. Clark, S. C. Hand, R. D. Bowlus, andG. N. Somero. 1982. Living with water stress: evolution ofosmolyte systems. Science 217:1214-1222.
VOL. 165, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
26
Oct
ober
202
1 by
77.
235.
114.
110.







![Osmoregulation Bacillus Synthesis Osmoprotectant …convert it enzymatically into glycine betaine. Transport assays. Uptake of choline in B. subtilis was measured by using [methyl-14C]choline](https://img.pdfslide.us/doc/110x75/5e4a7fe41d0756019f21e906/osmoregulation-bacillus-synthesis-osmoprotectant-convert-it-enzymatically-into-glycine.jpg)