Embed Size (px)
Citation preview

Cholesterol metabolism in the rat adrenal cortex: acute temporal changes following stress
JOSEPHINE ALFANO, ROBERT C. BEDERSEN, ROBERT E. KRAMER, ' AND ALEXANDER C. BROWNIE Bepartmerzts of Biochemistry and Puthology, State University ofNew York at Buflalo, Buflalo, NY, U.S.A. 14214
Received September 2, 1982
Alfano, J., Pedersen, R. C., Kramer, R. E. & Brownie, A. C. (1983) Cholesterol metabolism in the rat adrenal cortex: acute temporal changes following stress. Carz. J. Biochem. Cell Biol. 61, 708-713
Adrenal and serum corticosterone concentrations, cholesterol association with adrenal cortical cytochrome P-450,,, (the cytochrome P-450 catalyzing the conversion of cholesterol to pregnenolone), and adrenal cortical activities of ch~lesteryl ester hydrolase, acyl-CoA:cholesterol acyltransferase, and cholesterol side-chain cleavage have been determined at various times following stress in female rats. The paramount importance of cholesterol side-chain cleavage activation in the response to stress at the low point of the circadian rhythm is confirmed. At the high point of the circadian rhythm, there is evidence that the provision of free cholesterol for the cytochrome P-450,,, enzyme system may also be controlled. The data support a coordinate action of ACTH and pro-y-melanotropin in controlling cholesterol metabolism in the adrenal cortex following stress.
Alfano, J., Pedersen, R. C., Kramer, R. E. & Brownie, A. C. (1983) Cholesterol metabolism in the rat adrenal cortex: acute temporal changes following stess. Can. J. Biochenz. Cell Biol. 61, 708-713
Nous avons soumis des rates ii un stress et nous avons dktermink les paramktres suivants ii divers intervalles de temps: concentrations de la corticostCrone dans les surrknales et le drum, association du cholestkrol au cytochrome P-450,,, (cytochrome P-450 catalysant la conversion du cholestkrol en prkgn&nolone) de la corticosurr6rale et les activitks corticosurrknales de la cholestkryl ester hydrolase, de l'acyl-CoA:cholestkrol acyltransfkrase et du clivage de la chaine latkrale du cholestkrol. Nous avons confirm6 l'importance extrlime du clivage de la chaine latkrale du cholestkrol en rkponse au stress au point infkrieur du rythme circadien. Au point supCrieur du rythme circadien, il y a des preuves que la rkserve de cholestkrol libre pour le systbme enzymatique cytochrome P-450,,, peut aussi litre contrdlke. Les rksultats montrent une action coordonnke de 1'ACTH et de la pro-y-mklanotropine dans le contrdle du mktabolisme du cholestkrol dans le cortex surrknalien suite ii un stress.
[Traduit par la revue]
Introduction It is well recognized that ACTH is of primary
importance in the control of adrenal cortical hormone biosynthesis and that an increase in circulating ACTH levels following stress precedes the rise in blood glucocorticoids (1). Established actions of ACTFH in- clude the activation of cholesterol side-chain cleavage (2-4) and an increase in the uptake of cholesterol-rich serum lipoproteins (5). Studies on the effect of stress at the high point in the circadian rhythm (6) have suggested that a rate-limiting event is the provision of free choles- terol to the mitochondria1 cholesterol side-chain cleav- age system. For that reason the effect of stress on the activity of CEH and ACAT is of interest.
ABBREVIATIONS: cytochmme P-450,,, , cytochrome P-450 side-chain cleavage (the cytochrome P-450 catalyzing the con- version of cholesterol to pregnenolone); ACTPH, adrenocor- ticotropin; CEH, cholesteryl ester hydrolase (EC 3.1.1.13); ACAT, acyl-CoA:cholesterol acyltransferase (EC 2.3.1.26); Trilostane, (2a, 4a,5a, 17a)-4,5-epoxy- 17-hydroxy-3-oxoan- drostane-2-carbonitrile; HGI, heat-generated type I absorb- ance change; PII, pregnenolone-induced type I1 absorbance change; AUFS, absorbance units full scale; pro-y-MSH, pro- y-melanotropin; POMC, pro-opiomelanocortin.
'present address: Department of Obstetrics and Gynecol- ogy, The University of Texas Health Science Center at Dallas, Dallas, TX, U.S.A. 75235.
During the circadian rhythm in the rat adrenal-pitui- tary axis the fluctuation in plasma corticosterone is much greater than that of ACTH (7), and Dallman et al. (8) have reported that there is a greater adrenal response to exogenous ACTH in the evening (high point) than in the morning (low point). Moreover, since blood glucocorti- coid levels increase within a few minutes of imposing stress, data concerning the phenomena intrinsic to ACTH-stimulated steroidogenesis should be developed within that time period (9). Thus, the present study was designed to examine acute changes in adrenal cortical function after stress at both the low and high points of the circadian rhythm.
Materials and methods Materials
Pregnenolone was a product of Steraloids, Wilton, NH. Cholesterol, cholesteryl oleate, [ 9 ,10 -~~(~) ]o l ea t e (88 mCi/ m o l , 1 Ci = 37 GBq) and cholesteryl [9,10- oleate ate (1 0 mCi /mol ) were from Applied Science, State College, PA. Cholesteryl [l- '4~]oleate (55 mCi/mmol), [oleoyl-l- 14~]oleoyl coenzyme A (50 mCi/mmol), and ~ 7 - ~ ~ ] ~ r e g n e n - olone (25 Ci/mmol) were purchased from New England Nu- clear, Boston, MA. Trilostane was a gift of Sterling-Winthrop, New York, NY. Other materials were reagent grade or better.
Animal experimentation and tissue preparation Female Sprague-Dawley rats (Holtzman Co., Madison,
WI) weight 200-225 g were individually caged in three rooms
Can
. J. B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f L
aval
on
09/1
0/13
For
pers
onal
use
onl
y.

ALFANO ET AL.
TABLE 1. Adrenal cortical activity in quiescent rats at the low and high points of the circadian rhythm
Plasma corticosterone (pg/dL) 3.8k0.9 44.4k7.0 Adrenal corticosterone (pg / 100 mg protein) 5 .05 1.1 133.4k24.8 HGI (AA3w20 X lo3 /min per milligram protein) 2.1 k0.5 11.621.5 PII x lo3 /min per milligram protein) 19.5 k 4.9 30.8k4.4 Side-chain cleavage
( m o l product/min per milligram protein) 0.850.1 2.050.3 CEH (nmol product/min per milligram protein) 1.221.2 2.5k 1.2 ACAT (nmol product/min per milligram protein) 2.52~0.1 2.550.1
NOTE: Values are expressed as mean -+ SEM ( n = 8).
with controlled temperature (22 + 1°C) and fluorescent illumination (lights on 0600- 1808). Rat chow and water were allowed ad libitum. All rats were handled daily for at least 7 days prior to each experiment to acclimate them to animal quarter entry and removal from cages.
Four experiments were carried out at 0800 and four at 1800. In each experiment, 12 animals were killed by decapitation, 2 under quiescent conditions (6) and 2 each at 2, 5, 7, 10, and 15 min after stress. Stress was accomplished by the brief intraperitoneal insertion of a 20-gauge needle.
Rats were decapitated and trunk blood was collected into chilled beakers containing sodium heparin (50 U) in 50 uL of physiological saline. The inner zones of the adrenal cortex were then removed by rapid adrenal enucleation in situ. Tissue from each adrenal pair was homogenized in 5 mL of ice-cold Tris-sucrose and subcellular fractions were prepared by differential centrifugation, as previously described (10). Assays were carried out using cell fractions and plasma obtained from individual animals.
Enzyme assays Cholesterol side-chain cleavage activity was measured as
previously described (1 I), except that Trilostane was substi- tuted for cyanoketone as an inhibitor of 3P-01-dehydrogenase- isomerase. Mitochondria1 suspensions containing approxi- mately 0.1 mg/mL protein were incubated for 2 min at 37°C. The pregnenolone formed from endogenous cholesterol was extracted into methylene chloride and quantitated by radioim- munoassay (1 2).
The extent of endogenous cholesterol association with the ~nitochondrial cytochrome P-450,,, complex was determined by techniques which measure the absorbance change asso- ciated with substrate (cholesterol) binding (type I) and dis- sociation (type 11) from the cytochrome P-45OsCc complex. The HGI (1 3) is observed when mitochondria1 samples are warmed from 1 to 37'C. The type 11 (reverse type I) absorbance change is observed when cholesterol is displaced from the complex by pregnenolone (3). All measurements were carried out using an Arninco DW-2 recording spectrophotometer with mono- chromators set at 398 and 420 nm. The mitochondrial protein concentration in these assays was approximately 0.1 mg/mL. Cytochrome P-458 concentrations were measured according to Omura and Sato (14).
Neutral CEH activity in the 105 000 X g supernatant fraction was determined radioisotopically, as previously de- scribed (15). Microsomal ACAT activity was assayed accord-
ing to Balasubramaniam et a / . (lQ), except that incubations were carried out for 15 min and contained 25-50 pg protein per 0.20 mL of reaction buffer (pH 6.8).
Miscellaneous Corticosterone in plasma and adrenal cytosol was deter-
mined fluorometrically (17) using an ethanolic H2S04 reagent. To eliminate nonspecific background fluorescence, the plasma assays were blanked against rat serum which had been stripped of steroids.
Protein was determined by the Bradford technique (18) using standards of bovine serum albumin.
All organic solvents were redistilled before use.
Results Circadian rhythm studies
The concentrations of corticosterone in the plasma and adrenal cytosol of quiescent rats were 15-fold and 26-fold higher, respectively, at 1800 as compared with 0800 (Table 1). This is consistent with a previous report from this laboratory (6) and with studies by others (7, 19). The in vitro cholesterol side-chain cleavage activity in mitochondria isolated from quiescent rats was also elevated in the evening, although the magnitude of the increase (2.5-fold) was not as great as that seen in corticosterone concentrations. The HGI and BII absor- bance changes, each of which reflects the association of cholesterol with cytochrome B-450,,,, were increased to about the same extent as cholesterol side-chain cleav- age. CEH activity was increased approximately twofold at 1800 as compared with 0800, whereas there was no change in ACAT activity.
Efiects of stress There were no significant changes in any of the
monitored variables at 2 min after an 0800 stress. At 5 min, the concentrations of corticosterone in plasma and the adrenal cytosol were significantly increased and continued to rise throughout the 15-min period (Fig. 1). An increase in cholesterol side-chain cleavage activity was maximal at 5 min after stress and declined thereafter to a level that was twofold greater at 15 min than the quiescent control level (Fig. 2). A similar temporal
Can
. J. B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f L
aval
on
09/1
0/13
For
pers
onal
use
onl
y.
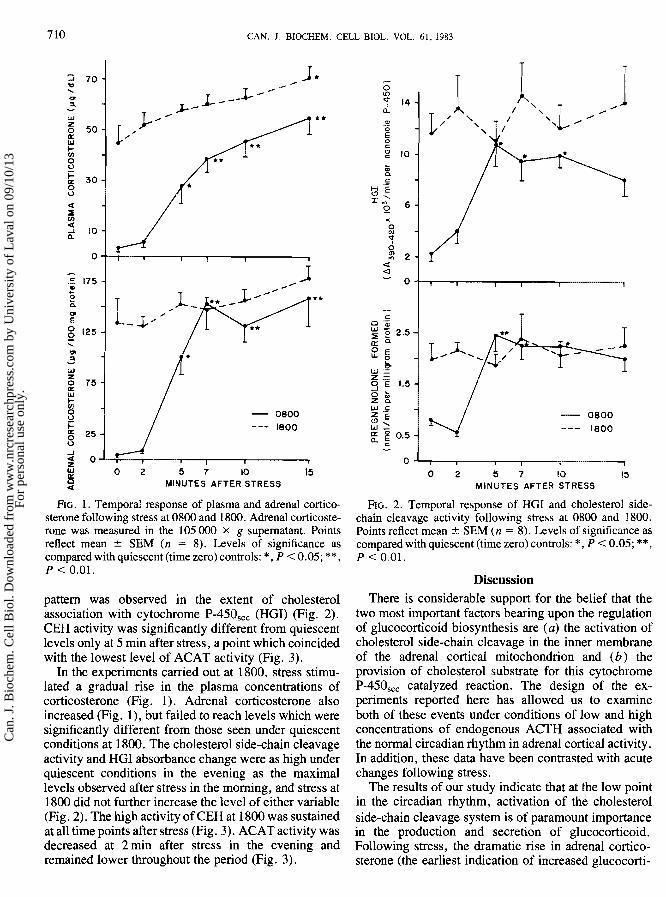
7 10 CAN. J . BIOCHEM. CELL BIOL. VOL. 61, 1983
1 O w 0 2 5 7 10 15 B Q MINUTES AFTER STRESS
FIG. 1. Temporal response of plasma and adrenal cortico- sterone following stress at 0800 and 1800. Adrenal corticoste- rone was measured in the 105 000 X g supernatant. Points reflect mean k SEM (n = 8). Levels of significance as compared with quiescent (time zero) controls: * , P < 0.05; ** , P < 0.01.
pattern was observed in the extent of cholesterol association with cytochrome P-450,,, (HGI) (Fig. 2). CEH activity was significantly different from quiescent levels only at 5 min after stress, a point which coincided with the lowest level of ACAT activity (Fig. 3).
In the experiments carried out at 1800. stress stimu- lated a gradual rise in the plasma concentrations of corticosterone (Fig. I). Adrenal corticosterone also increased (Fig. 1), but failed to reach levels which were significantly different from those seen under quiescent conditions at 1800. The cholesterol side-chain cleavage activity and PIG1 absorbance change were as high under quiescent conditions in the evening as the maximal levels observed after stress in the morning, and stress at 1800 did not further increase the level of either variable (Fig. 2). The high activity of CEH at 1800 was sustained at all time points after stress (Fig. 3). ACAT activity was decreased at 2 min after stress in the evening and - remained lower throughout the period (Fig. 3).
0 2 5 7 I 0 15 MINUTES AFTER STRESS
FIG. 2. Temporal response of HGI and cholesterol side- chain cleavage activity following stress at 0800 and 1800. Points reflect mean iz SEM (n = 8). Levels of significance as compared with quiescent (time zero) controls: *, P < 0.05; **, P < 0.01.
Discussion There is considerable support for the belief that the
two most important factors bearing upon the regulation of glucocorticoid biosynthesis are (a) the activation of cholesterol side-chain cleavage in the inner membrane of the adrenal cortical mitochondrion and ( b ) the provision of cholesterol substrate for this cytochrome P-450,,, catalyzed reaction. The design of the ex- periments reported here has allowed us to examine both of these events under conditions of low and high concentrations of endogenous ACTH associated with the normal circadian rhythm in adrenal cortical activity. In addition, these data have been contrasted with acute changes following stress.
The results of our study indicate that at the low point in the circadian rhythm, activation of the cholesterol side-chain cleavage system is of paramount importance in the production and secretion of glucocorticoid. Following stress, the dramatic rise in adrenal cortico- sterone (the earliest indication of increased glucocorti-
Can
. J. B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f L
aval
on
09/1
0/13
For
pers
onal
use
onl
y.
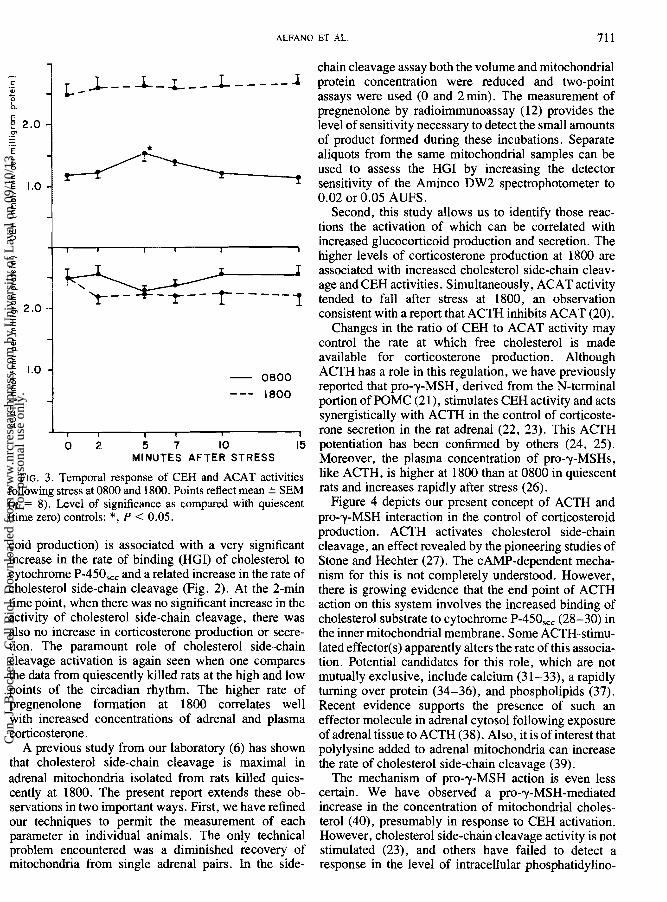
0 2 5 7 10 15 MINUTES AFTER STRESS
FIG. 3. Temporal response of CEH and ACAT activities following stress at 0800 and 1800. Points reflect mean + SEM (n = 8). Lave1 of significance as compared with quiescent (time zero) controls: *, P < 0.05.
coid production) is associated with a very significant increase in the rate of binding (HGI) of cholesterol to cytochrome B-450,,, and a related increase in the rate of cholesterol side-chain cleavage (Fig. 2). At the 2-min time point, when there was no significant increase in the activity of cholesterol side-chain cleavage, there was also no increase in corticosterone production or secre- tion. The paramount role of cholesterol side-chain cleavage activation is again seen when one compares the data from quiescently killed rats at the high and low points of the circadian rhythm. The higher rate of pregnenolone formation at 1800 correlates well with increased concentrations of adrenal and plasma corticosterone .
A previous study from our laboratory (6) has shown that cholesterol side-chain cleavage is maximal in adrenal mitochondria isolated from rats killed quies- cently at 1800. The present report extends these ob- servations in two important ways. First, we have refined our techniques to permit the measurement of each parameter in individual animals. The only technical problem encountered was a diminished recovery of mitochondria from single adrenal pairs. In the side-
chain cleavage assay both the volume and mitochondrial protein concentration were reduced and two-point assays were used (0 and 2 min). The measurement of pregnenolone by radioimmunoassay (12) provides the level of sensitivity necessary to detect the small amounts of product formed during these incubations. Separate aliquots from the same mitochondrial samples can be used to assess the HGI by increasing the detector sensitivity of the Aminco DW2 spectrophotometer to 0.02 or 0.05 AUFS.
Second, this study allows us to identify those reac- tions the activation of which can be correlated with increased glucocorticoid production and secretion. The higher levels of corticosterone production at 1800 are associated with increased cholesterol side-chain cleav- age and CEH activities. Simultaneously, ACAT activity tended to fall after stress at 1800, an observation consistent with a report that ACTH inhibits ACAT (20).
Changes in the ratio of CEH to ACAT activity may control the rate at which free cholesterol is made available for corticosterone production. Although ACTH has a role in this regulation, we have previously reported that pro- y -MSH , derived from the N-terminal portion of BOMC (21), stimulates CEH activity and acts synergistically with ACTH in the control of corticoste- rone secretion in the rat adrenal (22, 23). This ACTH potentiation has been confirmed by others (24, 25). Moreover, the plasma concentration of pro-y-MSHs, like ACTH, is higher at 1800 than at 0800 in quiescent rats and increases rapidly after stress (26).
Figure 4 depicts our present concept of ACTH and pro-y-MSH interaction in the control of corticosteroid production. ACTH activates cholesterol side-chain cleavage, an effect revealed by the pioneering studies of Stone and Hechter (27). The CAMP-dependent mecha- nism for this is not completely understood. However, there is growing evidence that the end point of ACTH action on this system involves the increased binding of cholesterol substrate to cytochrome P-450,,, (28-30) in the inner mitochondrial membrane. Some ACTH-stimu- lated effector(s) apparently alters the rate of this associa- tion. Potential candidates for this role, which are not mutually exclusive, include calcium (3 1-33), a rapidly turning over protein (34-36), and phospholipids (37). Recent evidence supports the presence of such an effector molecule in adrenal cytosol following exposure of adrenal tissue to ACTH (38). Also, it is of interest that polylysine added to adrenal mitochondria can increase the rate of cholesterol side-chain cleavage (39).
The mechanism of pro-y-MSH action is even less certain. We have observed a pro-y-MSH-mediated increase in the concentration of mitochondrial choles- terol (40), presumably in response to CEH activation. However, cholesterol side-chain cleavage activity is not stimulated (23), and others have failed to detect a response in the level of intracellular phosphatidylino-
Can
. J. B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f L
aval
on
09/1
0/13
For
pers
onal
use
onl
y.
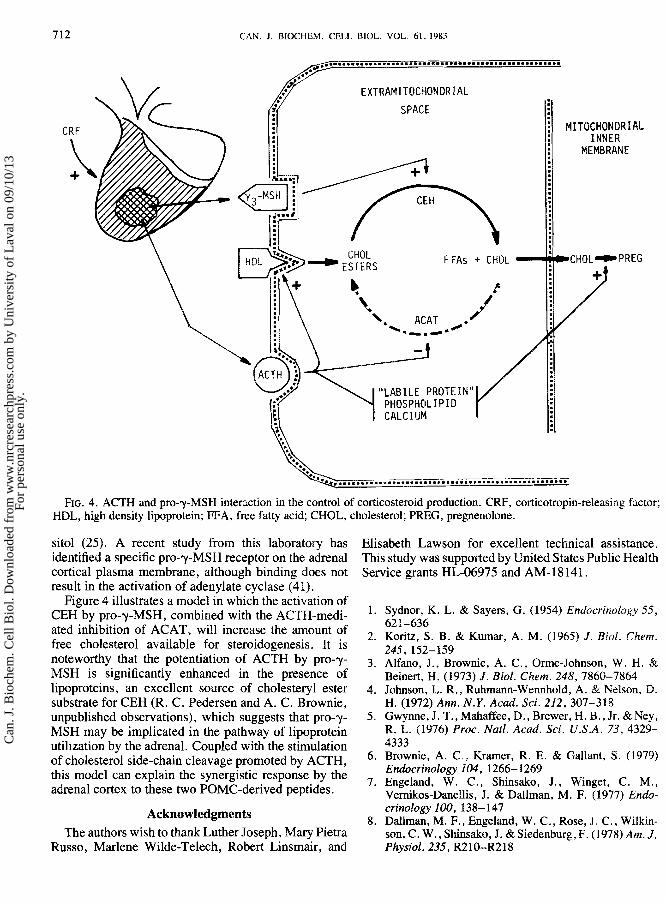
CAN. J . BIBCHEM. CELL BIOL. VOL. 61, 1983
EXTRAMITOCHONDRIAL
MITOCHONDRIAL
"LABILE PROTEIN"
FIG. 4. ACTH and pro-y-MSH interaction in the control of corticosteroid production. CRF, corticotropin-releasing factor; HDL, high density lipoprotein; FFA, free fatty acid; CHOL, cholesterol; PREG, pregnenolone.
sitol (25). A recent study from this laboratory has identified a specific pro-y-MSH receptor on the adrenal cortical plasma membrane, although binding does not result in the activation of adenylate cyclase (411).
Figure 4 illustrates a model in which the activation of CEH by pro-y-MSH, combined with the AGTH-medi- ated inhibition of ACAT, will increase the amount of free cholesterol available for steroidogenesis. It is noteworthy that the potentiation of ACTH by pro-y- MSH is significantly enhanced in the presence of lipoproteins, an excellent source of cholesteryl ester substrate for CEH (R. C. Pedersen and A. C. Brownie, unpublished observations), which suggests that pro-y- MSH may be implicated in the pathway of lipoprotein utilization by the adrenal. Coupled with the stimulation of cholesterol side-chain cleavage promoted by ACTH, this model can explain the synergistic response by the adrenal cortex to these two POMC-derived peptides.
Elisabeth Lawson for excellent technical assistance. This study was supported by United States Public Health Service grants HL-06975 and AM- 18 11411.
1. Sydnor, K. L. & Sayers, G. (1 954) Endocrinology 55, 621 -636
2. Koritz, S. B. & Kuma, A. M. (1965) J. Bkol. Chem. 245, 152-159
3. Alfano, J., Brownie, A. C., Orm-Johnson, W. H. & Beinert, H. (1973) J. Biol. Chem. 248,7860-7864
4. Johnson, L. R., Ruhmann-Wennhold, A. & Nelson, D. H. (1972) Ann. N.Y. Acad. Sci. 212, 307-318
5. Gwynne, J. T., Mahaffee, D., Brewer, H. B., Jr. & Ney, R. L. (1976) Proc. Natl. Acad. Sci. U.S.A. 73, 4329- 4333
6. Brownie, A. C., Krmer, R. E. & Gallant, S. (1979) Endocrinology 104, 1266- 1269
7. Engeland, W. C., Shinsako, J., Winget, C. M., Vernikos-Danellis, J. & Dallman, Me F. (1977) Endo-
Acknowledgments crinology 100, 138-147 8. Dallman, M. F., Engeland, W. C., Rose, J. C., Wilkin-
The authors wish to thank Luther Joseph, Mary Pietra son, C. W., Shinsaks, J. & Siedenburg, F. (1978) Am. 9. - Russo, Marlene Wilde-Telech, Robert Linsmair, and Bhysiol. 235, R210-R218
Can
. J. B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f L
aval
on
09/1
0/13
For
pers
onal
use
onl
y.

ALFAWO ET AL. 713
9. Haynes, R. C., Jr. (1968) in Functions of the Adrenal Cortex (McKerns, K. W., ed.), vol. 1 , pp. 583-6430, Appleton-Century-Crofts , New York
10. Kramer, R. E., Gallant, S. & Brownie, A. C. (1979) J. Biol. Chem. 254, 3953-3958
1 1 . Fink, C. S., Gallant, S. & Brownie, A. C. (1980) Hypertension (Dallas) 2, 6 17-622
12. Bergon, L., Gallant, S. & Brownie, A. C. (1974) Endocrinology 94, 336-345
13. Paul, D. P., Gallant, S., Onne-Johnson, N. R., Orme- Johnson, W. H. & Brownie, A. C. (1976) J. Biol. Chem. 251, 7120-7126
14. Omura, T. & Sato, R. (1964) J. Biol. Chem. 193, 2370-2378
15. Pedersen, R. C. & Brownie, A. C. (1979) J. Steroid Biochem. 11, 1393-1400
16. Balasubramaniam, S., Goldstein, J. L., Faust, J. R., Brunschede, G. Y. & Brown, M. S. (1977) J . Biol. Chem. 252, 1771-1779
17. Silber, R. H. , Busch, R. D. & Oslapas, R. (1958) Clin. Chem. (Winston-Salem, N.C.) 4 , 278-285
18. Bradford, M. M. (1976) Anal. Biochem. 72, 248-254 19. Ixm, G., Szafarczyk, A., Belugou, J. L. & Assen-
macher, I. (1977) J. Endocrinol. 72, 113- 120 20. Shima, S., Mitsunaga, M. & Nakao, T. (1972) Endocri-
nology 90, 808-814 21. ~ i ~ p e r , B. A. & Mains, R. E. (1980) Endocrine Rev. 1 ,
1-27 22. Pedersen, R. C. & Brownie, A. C. (1980) Proc. Natl.
Acad. Sci. U.S.A. 77, 2239-2243 23. Pedersen, R. C., Brownie, A. C. & Ling, N. (1980)
Science (Washington, D . C . ) 208, 1044- 1046 24. Al-Dujaili, E. A. S., Hope, J., Estivariz, F. E., Lowry,
P. J. & Edwards, C. R. W. (1981) Nature (London) 291, 156-159
25. Farese, R. V., Sabir, M. A. & Larson, R. E. (1982)
Program of the 64th Annual Meeting of the Endocrine Society, San Francisco, CA, June 17, 1982, p. 221 (abstr.)
26. Pedersen, R. C., Ling, N. & Brownie, A. C. (1982) Endocrinology 11 0, 825-834
27. Stone, D. & Hechter, 0 . (1954) Arch. Biochem. Biophys. 51, 457-469
28. Brownie, A. C., Alfano, J. A., Jefcoate, J. R., Orme- Johnson, W. H., Beinert, H. & Simpson, E. R. (1973) Ann. N.Y. Acad. Sci. 212,344-360
29. Jefcoate, C. R. & Orme-Johnson, W. H. (1975) J . Biol. Chem. 250,467 1-4677
30. Simpson, E. R. (1979) Mol. Cell. Endocrinol. 13, 21 3-227
31. Birmingham, M. K., Elliot, F. H. & Valere, P. H. L. (1953) Endocrinology 53, 687-689
32. Farese, R. V. (1971) Endocrinology 89, 1057- 1063 33. Simpson, E. R., Jefcoate, C. R., McCarthy, J. L. &
Boyd, G . S. (1974) Eur. J . Biochem. 45, 181-188 34. Garren, L. B. , Ney, R. L. &Davis, W. W. (1965) Proc.
Natl. Acad. Sci. U.S.A. 53, 1443-1450 35. Ferguson, J. J., Jr. (1963) J. Biol. Chem. 238, 2754-
2759 36. Schulster, B. & Jenner, C. (1975) J . Steroid Biochem. 6,
389-394 37. Farese, R. V. & Sabir, A. M. (1980) Endocrinology 106,
1869- 1879 38. Neher, R., Milani, A., Solano, A. R. & Podesta, E. J.
(1982) Proc. Natl. Acad. Sci. U.S.A. 79, 1727-1731 39. Kido, T. & Kimura, T. (1981) J . Biol. Chem. 256,
8561 -8568 40. Pedersen, R. C. & Brownie, A. C. (1980) Program of the
62nd Annuul Meeting of the Endocrine Society, Washing- ton, D.C. , June 19, 1980, p. 220 (abstr.)
41. Pedersen, R. C. & Brownie, A. C. (1982) Endocrinol- ogy 112, 1279-1287
Can
. J. B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f L
aval
on
09/1
0/13
For
pers
onal
use
onl
y.