Embed Size (px)
Citation preview

Journal of Archaeological Science 34 (2007) 48e58http://www.elsevier.com/locate/jas
Shell tool use by early members of Homo erectus in Sangiran,central Java, Indonesia: cut mark evidence
Kildo Choi a,*, Dubel Driwantoro b
a 5015 Sheboygan Avenue, Suite 204, Madison, WI 53705, USAb Pusat Penelitian Arkeologi Nasional, Jl. Raya Condet Pejaten No. 4, Jakarta 12001, Indonesia
Received 11 January 2006; received in revised form 21 March 2006; accepted 29 March 2006
Abstract
Sangiran has been known as a source of fossil Homo erectus but is better known for the absence of archaeological tools. Cut mark analysis ofPleistocene mammalian fossils documents 18 cut marks inflicted by tools of thick clamshell flakes on two bovid bones created during butchery atthe Pucangan Formation in Sangiran between 1.6 and 1.5 million years ago. These cut marks document the use of the first tools in Sangiran andthe oldest evidence of shell tool use in the world.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Thick clam shell; Cut mark; Homo erectus; Sangiran; Java
1. Introduction
Sangiran has yielded fossil Homo erectus, representingseveral dozen individuals, from the upper 15-m section ofthe Pucangan Formation and the entire Kabuh Formation[32] ranging over 1.6e1.0 million years ago (myr) [38,59].However, the absence of archaeological tools has long puzzledpalaeoanthropologists. Our research focused on tool use andthe diet of early members of H. erectus found in the PucanganFormation, known as the Black Clay deposits, datable between1.6 and 1.5 myr. To date, H. erectus fossils recovered from theformation represent approximately a dozen individuals and areclassified as Meganthropus [33] or the Robust Group [79]against gracile ones in the Kabuh Formation. Since no archae-ological tools have been found in the entire formation, weexamined evidence from hominids’ butchery activities pre-served as cut marks on the faunal fossils excavated from theformation. This research is based on the premise that cutmarks indicate hominids’ tool use during butchery even in
* Corresponding author. Tel.: þ1 608 238 4911.
E-mail address: [email protected] (K. Choi).
0305-4403/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jas.2006.03.013
the absence of the actual tools [19,20]. Recovered archaeolog-ical cut marks were examined using scanning electronicmicroscopy to identify the specific raw materials and type ofthe tools.
2. Archaeological problem
In Southeast Asia, H. erectus’s old stone tools that are con-vincingly datable to the Lower- and Middle Pleistocene ageare almost non-existent although those of similar age are abun-dantly documented outside the region [37]. In Island SoutheastAsia, there have been numerous claims of old stone tools(Fig. 1) but other researchers have discredited most of thesefindings. For instance, stone tools of Cabenge, southern Sula-wesi, were reported in association with faunal remains [70],yet, the lithic remains were argued to have originated in termi-nal Pleistocene gravel beds of the Upper Pleistocene age[4,11,36,55]. Lithic flakes and bifacial objects found with steg-odon fossils in Flores Island [25,43,44] and similar remainsfrom Sumbawa [63] were commonly criticized due to unclearstratigraphic associations. Pebble choppers of Sambungmacanwere reported to be 0.9 myr [34] but the channel-fill context

49K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
Fig. 1. Location of Sangiran and other sites.
made findings questionable. The flake tools from Ngandong[72] were repudiated by unclear geologic context [7,9,10].Chalcedony/jasper flake tools from the surfaces of the NotopuroFormation at Ngebung in Sangiran, initially believed to be0.8 myr [71,73e75], were reinterpreted to be of earlyHolocene age [8]. Lithic flakes and bifaces from Pajitan, southCentral Java, have been reevaluated as the late Upper Pleisto-cene age [3]. All of the above discoveries were ambiguousdue to their unclear geological context, occurrence in a second-ary deposit or having younger ages than the originally claimed.
During the 1990s, excavation at the Mata Menge site onFlores Island, Indonesia, produced 0.8 myr lithic flake toolsin association with megafaunal remains [45,65]. In Sangiran,Semah’s team [60] excavated chalcedony flakes and andesiteobjects from the lower section of the Kabuh Formation atNgebung (Fig. 2). Earlier, von Koenigswald and Bartstra[6,8,74,75] warned that a large quantity of chalcedony flakesand andesite choppers exist in the Notopuro Formation andyounger deposits. Analysis of the Ngebung assemblagedisclosed association of chalcedony flakes in all of the artifactbearing layers [62]. The combined ESR/Ur-series dates ofa few mammal teeth provided dates too young for the Kabuh For-mation [21]. Thus these lithic remains are thought to have beendislocated from the Notopuro Formation or younger deposits [64].
Widianto [81] reported chalcedony flakes and andesiteobjects from the Grenzbank deposit, a discrete sandstone con-glomerate deposit situated between the Kabuh and PucanganFormations, at Dayu in Sangiran. Recently, Widianto [80]made another discovery of chalcedony flakes from 3 m belowthe Pucangan Formation at the same locality of Dayu. How-ever, we note that chalcedony flakes appear repeatedly in thedeposits of different ages at the same locality. In addition,the currently earliest mammals in Sangiran appear approxi-mately 40 m above and the earliest known fossil H. erectusis nearly 70 m above these new findings. Regardless of thereliability of the above lithic remains, no archaeological stonetool has yet been discovered from the Pucangan Formation.
The scarcity of pre-Upper Pleistocene stone tools is wellreflected in three regional explanations. Bartstra [5] arguedthat the absence of in situ Lower/Middle Pleistocene lithicassemblages in Island Southeast Asia indicates that Homosapiens were the first stone tool makers in the region. Yet, thediscovery of 0.8 myr stone tools at Mata Menge contradictedthis proposition. Puech [54] proposed that dental wear of a molarof Sangiran 4 indicated vegetarian diet, thus sophisticated lithictechnology developed only at a later date in Java. The idea of H.erectus’s non-lithic tool use has achieved the greatest popularityamong scholars of Southeast Asia [16,17,30,51,52,57,76,83].The concept is best conveyed in the Bamboo-Karst Model thatemphasizes the replacement of Acheulean bifaces by bambootools [52], or lignic (wooden) tools [30], thus lacking in thechanges paralleling western Eurasian Lower and Middle Palae-olithic industries such as Acheulean hand-axes and Levalloistools. However, no archaeological evidence has yet substanti-ated or repudiated this idea owing to the perishable nature ofthe material.
3. Rationale and materials
We opened doors to all possibilities for early H. erectus’sselection of raw materials for their butchery tools. A total of12 lithic and non-lithic raw materials were selected and usedfor experimental butchery to build new definitions of cutmarks for each material and to distinguish one raw materialfrom the other. Because conventional definitions of cut markscover only those of stone tools [19,61], a set of the new defi-nitions is necessary. A complete discussion of the cut marksand butchery efficiency of the 12 materials will be publishedelsewhere.
The selected 12 raw materials include four types of lithicand eight types of non-lithic materials. The lithic materialsare: soft limestone of the Kalibeng Formation, coarse-grainedandesite of the Lower Lahar (volcanic mudflow), fine-grainedbasalt of the Notopuro Formation and rhyolite of Baksoko
50 K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
River near Pajitan area. The non-lithic raw materials are: fourvarieties of bamboos such as green and dry Bamboosa (thin-walled) and Dendrocalamus (thick-walled), thick and thinclam shells, coconut shell flakes and dry bovid bone flakes.
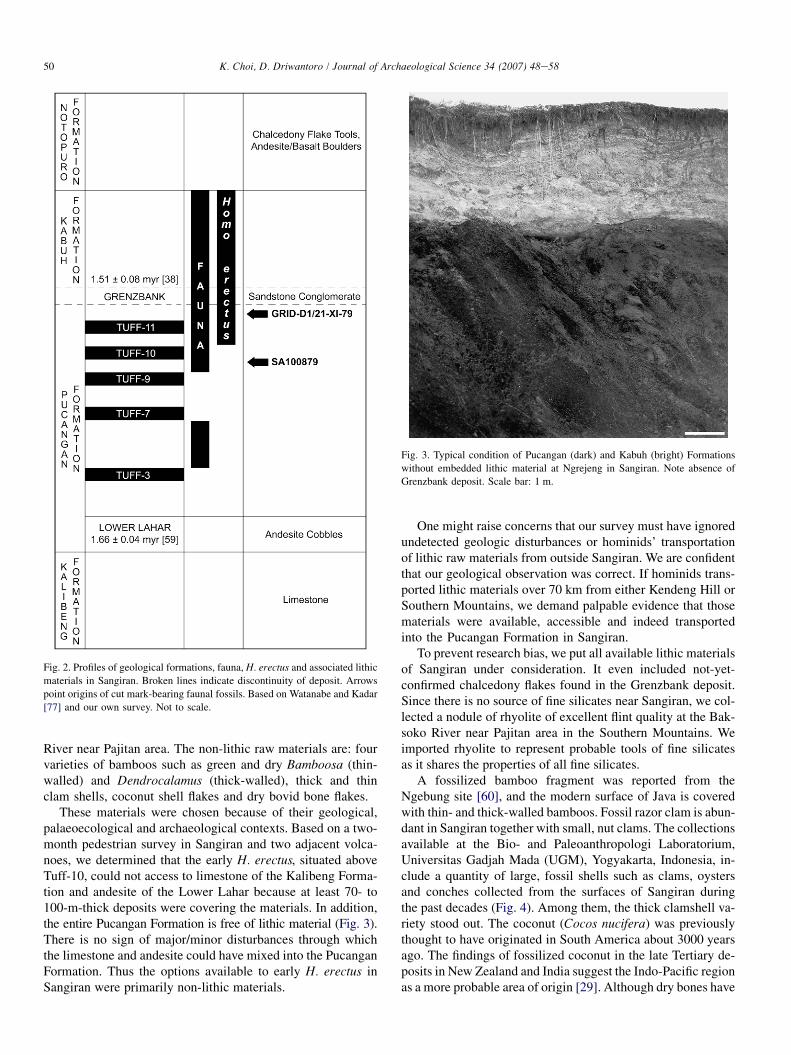
These materials were chosen because of their geological,palaeoecological and archaeological contexts. Based on a two-month pedestrian survey in Sangiran and two adjacent volca-noes, we determined that the early H. erectus, situated aboveTuff-10, could not access to limestone of the Kalibeng Forma-tion and andesite of the Lower Lahar because at least 70- to100-m-thick deposits were covering the materials. In addition,the entire Pucangan Formation is free of lithic material (Fig. 3).There is no sign of major/minor disturbances through whichthe limestone and andesite could have mixed into the PucanganFormation. Thus the options available to early H. erectus inSangiran were primarily non-lithic materials.
Fig. 2. Profiles of geological formations, fauna, H. erectus and associated lithic
materials in Sangiran. Broken lines indicate discontinuity of deposit. Arrows
point origins of cut mark-bearing faunal fossils. Based on Watanabe and Kadar
[77] and our own survey. Not to scale.
One might raise concerns that our survey must have ignoredundetected geologic disturbances or hominids’ transportationof lithic raw materials from outside Sangiran. We are confidentthat our geological observation was correct. If hominids trans-ported lithic materials over 70 km from either Kendeng Hill orSouthern Mountains, we demand palpable evidence that thosematerials were available, accessible and indeed transportedinto the Pucangan Formation in Sangiran.
To prevent research bias, we put all available lithic materialsof Sangiran under consideration. It even included not-yet-confirmed chalcedony flakes found in the Grenzbank deposit.Since there is no source of fine silicates near Sangiran, we col-lected a nodule of rhyolite of excellent flint quality at the Bak-soko River near Pajitan area in the Southern Mountains. Weimported rhyolite to represent probable tools of fine silicatesas it shares the properties of all fine silicates.
A fossilized bamboo fragment was reported from theNgebung site [60], and the modern surface of Java is coveredwith thin- and thick-walled bamboos. Fossil razor clam is abun-dant in Sangiran together with small, nut clams. The collectionsavailable at the Bio- and Paleoanthropologi Laboratorium,Universitas Gadjah Mada (UGM), Yogyakarta, Indonesia, in-clude a quantity of large, fossil shells such as clams, oystersand conches collected from the surfaces of Sangiran duringthe past decades (Fig. 4). Among them, the thick clamshell va-riety stood out. The coconut (Cocos nucifera) was previouslythought to have originated in South America about 3000 yearsago. The findings of fossilized coconut in the late Tertiary de-posits in New Zealand and India suggest the Indo-Pacific regionas a more probable area of origin [29]. Although dry bones have
Fig. 3. Typical condition of Pucangan (dark) and Kabuh (bright) Formations
without embedded lithic material at Ngrejeng in Sangiran. Note absence of
Grenzbank deposit. Scale bar: 1 m.

51K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
Fig. 4. Thick fossil shells of Sangiran among UGM Collection. A, Bullidae; B, Veneridae; C, Carditidae; D,E, Anomiidae; F, Littprinidae; G, Fasciolariidae. Scale
bar: 5 cm.
been proposed as butchery tools in Great Plains of North America[23,24,78], the faunal profile of the Pucangan Formation inSangiran shows bovids alongside cervids and large ungulates.We believe these 12 options are sufficient to cover the rangeof tool selection by the early H. erectus in Sangiran.
Due to limited availability and time constraints in Java, wesubstituted similar materials for three of the raw materials.During the survey, we found no large, thick fossil shellswith which we could conduct the butchery experiment. Asshellfish is unpopular in the local Muslim culture, large clamswere unavailable even at the local fish markets. This forced usto conduct butchery experiments in the United States usingsubstituted but physically similar thick, fossil clam shell(Veneridae) from Florida. To accommodate time constraintsin Java, butchery with flakes of coconut shell of Malaysianorigin and bone flakes of Wisconsin cattle were conductedwith bovid forelimbs available in Wisconsin.
At 18 institutions in Indonesia, Germany and the Nether-lands, the first author examined a total of 34,000 specimensof the Pleistocene faunal fossils recovered from both thePucangan and Kabuh Formations of Sangiran and CentralJava including excavated fossils, surface collections, antlersand teeth. Fossils from the lacustrine clay deposits of thePucangan Formation preserved bone surfaces in excellent con-dition with natural gloss, while those from the riverine sanddeposits of the Kabuh Formation, which constituted morethan 90% of the research collection, tended to show sandyconcretions cemented onto the bone surfaces and eroded sur-faces. Nearly all of the fossils from the Kabuh Formationwere unsuitable for cut mark analysis.
Excavated Pleistocene faunal fossil collections in Java arerare compared with the large volume of surface collections.A total of 2000 fossils of the Dubois Collection at the Natio-naal Natuurhistorisch Museum, Leiden, the Netherlands, orig-inated in the Kabuh Formation. The condition of bones in thiscollection was not good enough to observe cut marks.
Approximately 100 surviving fossils of the Selenka Collectionat the Museum fur Naturkunde, Humboldt University, Berlin,Germany, were excavated from the black clay deposits inTrinil. The collection consists of predominantly teeth togetherwith a few mandibles and limb bones. The bones are wellpreserved but no cut marks were recovered.
The IndonesianeJapanese Expedition 1976e1979 Collec-tion, at the Geological Research and Development Center(GRDC), Bandung, Indonesia, totaled approximately 1000specimens. The von Koenigswald collection of the NgandongTerrace at the Quaternary Geologi Laboratorium, Bandung,Indonesia, totaled 500 specimens. The Budisampoerno Collec-tion excavated from the Pucangan Formation in Sangiran,available at the UGM had only 50 fossils. We found cut marksamong the three latter collections. The total number of exca-vated fossils preserving bone surfaces suitable for cut markanalysis was only 400 excluding teeth and antlers. The collec-tions of Mata Menge, Ngebung and Dayu were inaccessible.
4. Archaeological evidence
4.1. Fossil: SA100879
The IndonesianeJapanese Expedition 1976e1979 exca-vated a complete left radius (SA100879) of a young adultbovid (Bos sp.) from Level 4 of Trench III in the Pucangan For-mation at Bukuran in Sangiran [1,35]. The specimen revealedtwo clusters of cut marks on a well-preserved, glossy bone sur-face. A cluster of 11 slicing grooves is arranged in a criss-crosspattern immediately above the radial tuberosity where the mus-cle biceps brachii is attached (Fig. 5). The second cluster has twogrooves arranged in sub-parallel orientation on the proximo-lateral end of the midshaft. Unlike stone tool cut marks, whichtypically form V cross-sections [19], these grooves are broadand flat. Gross morphology of the grooves include ‘double-track’ grooves with two parallel tracks with one bone ridge in

52 K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
Fig. 5. Archaeological cut marks on SA100879. (A) Location of cut marks. (B) Double-track grooves seen in E. (C) Double-track groove at second cluster. (D)
Double-track and canal groove in a single incision. (E) Cut marks (arrow) above radial tuberosity. Scale bars: B,C,D 1 mm; E 1 cm.
between (Fig. 5B,C), wide and flat ‘canal-like’ grooves (lowerhalf of Fig. 5D), grooves containing the above two features ina single incision (Fig. 5D), and wide V grooves at the upperleft corner of Fig. 5B. All of the grooves are 0.5e1.2 mmwide and less than 0.5 mm deep.
Based on cross-examination against experimental cutmarks, tools of thick clamshell flakes turned out to be themost plausible material for creating these cut marks. In EastAfrica, Toth and Woods [69] suspected a possible use of shelltools by the (Plio)-Pleistocene hominids, yet, no archaeologi-cal evidence has been recovered. Their study focused onmussel- and oyster-shells with retouched edges and concludedthat shell tool cut marks cannot be distinguished from those ofthe stone tools due to their similarity. However, we testedbroken flakes of thick clam shells without retouching sincewe assume that hominids would have first discovered the sharpand durable edges from the broken live/fossil shells.
Shell, either fresh or fossil, is composed of calcium carbon-ate in the form of calcite and aragonite or both in a layeredstructure. Bivalves like thick clam shells (Veneridae) are builtof three units: an outer prismatic layer (cortex); a midsection;and an inner prismatic layer (Fig. 6A,C). Mineral combinationin the three layers varies by the water temperature, i.e., allthree layers are aragonitic in warm water species, and oneprismatic calcite and two aragonitic layers in temperate waterspecies [40].
Sizes of our experimental shells range between 8 � 6 and10 � 8 cm with 4e10 mm in thickness, comprised of 0.7e2.0 mm for the cortex, 3.0e6.0 mm for the midsection, and0.5e1.5 mm for the inner layer. When thick clam shells frac-ture under impact, such as a hammer stone, another shell, ora human heel, they tend to produce 3e5 triangular and rectan-gular flakes. Edges along the fracture surfaces are either com-plex with concave, blunt and acute cross-sections (Fig. 6A) ordominantly blunt with angles of nearly 90 � (Fig. 6C). Thesethree types of edges coexist along any fracture surface in ran-dom combination. Based on experimental specimens, eachvalve offers 20e25 cm of fracture surface. Blunt edges thatare nearly 90 � are most common, an estimated 60% of theentire edge, while concave and acute edges take up approxi-mately 20% each. Because each layer fractures in its ownorientation, this fracture process is uncontrollable. Deliberatereplication of a specific cross-section is impossible. Edges atboth the cortex and inner layer have potential for cutting tools.
Although thick clam shells tend to fracture along thegrowth rings (annuli) yielding blunt edges, erratic fracturecauses some areas of the cortex to hold fragments of twoannuli which work as two barbs (Fig. 6A,B,G). These twobarbs constitute the edge in a concave cross-section and createcut marks with two parallel grooves (double-tracks) and onebone ridge in between (Fig. 6G,K). Unlike the sharp cuttingedges of fine silicates, the barbs are slightly rounded and
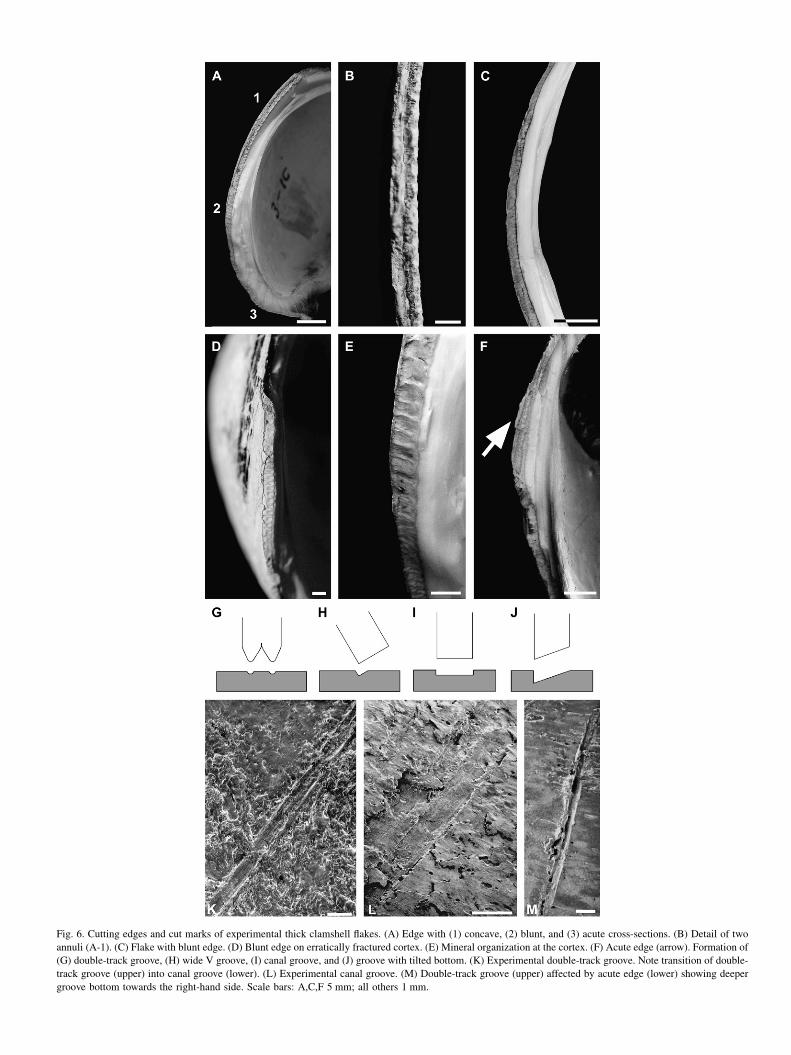
Fig. 6. Cutting edges and cut marks of experimental thick clamshell flakes. (A) Edge with (1) concave, (2) blunt, and (3) acute cross-sections. (B) Detail of two
annuli (A-1). (C) Flake with blunt edge. (D) Blunt edge on erratically fractured cortex. (E) Mineral organization at the cortex. (F) Acute edge (arrow). Formation of
(G) double-track groove, (H) wide V groove, (I) canal groove, and (J) groove with tilted bottom. (K) Experimental double-track groove. Note transition of double-
track groove (upper) into canal groove (lower). (L) Experimental canal groove. (M) Double-track groove (upper) affected by acute edge (lower) showing deeper
groove bottom towards the right-hand side. Scale bars: A,C,F 5 mm; all others 1 mm.
54 K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
dull at the tip while some portion of it is broken. Thus thegrooves exhibit shallow, round-bottomed tracks with jaggedmargins with breadth between 0.5e1.4 mm. These groovesmatch perfectly with the morphology and size of the archaeo-logical double-track cut marks on SA100879.
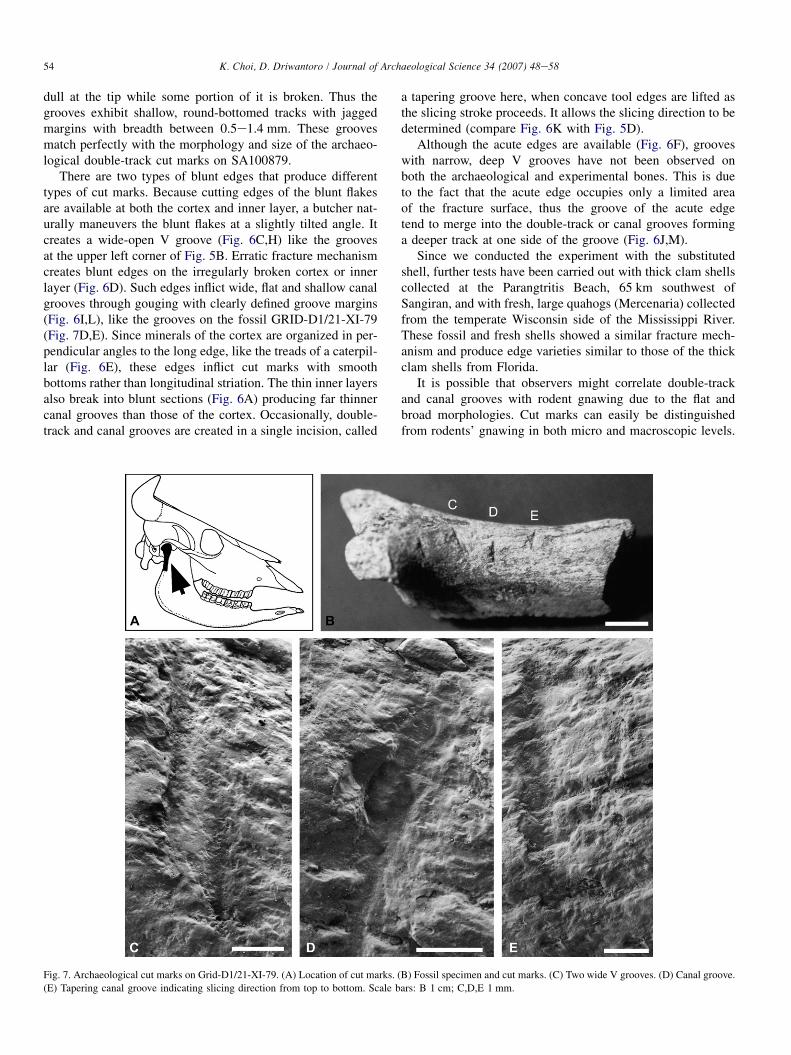
There are two types of blunt edges that produce differenttypes of cut marks. Because cutting edges of the blunt flakesare available at both the cortex and inner layer, a butcher nat-urally maneuvers the blunt flakes at a slightly tilted angle. Itcreates a wide-open V groove (Fig. 6C,H) like the groovesat the upper left corner of Fig. 5B. Erratic fracture mechanismcreates blunt edges on the irregularly broken cortex or innerlayer (Fig. 6D). Such edges inflict wide, flat and shallow canalgrooves through gouging with clearly defined groove margins(Fig. 6I,L), like the grooves on the fossil GRID-D1/21-XI-79(Fig. 7D,E). Since minerals of the cortex are organized in per-pendicular angles to the long edge, like the treads of a caterpil-lar (Fig. 6E), these edges inflict cut marks with smoothbottoms rather than longitudinal striation. The thin inner layersalso break into blunt sections (Fig. 6A) producing far thinnercanal grooves than those of the cortex. Occasionally, double-track and canal grooves are created in a single incision, called
a tapering groove here, when concave tool edges are lifted asthe slicing stroke proceeds. It allows the slicing direction to bedetermined (compare Fig. 6K with Fig. 5D).
Although the acute edges are available (Fig. 6F), grooveswith narrow, deep V grooves have not been observed onboth the archaeological and experimental bones. This is dueto the fact that the acute edge occupies only a limited areaof the fracture surface, thus the groove of the acute edgetend to merge into the double-track or canal grooves forminga deeper track at one side of the groove (Fig. 6J,M).
Since we conducted the experiment with the substitutedshell, further tests have been carried out with thick clam shellscollected at the Parangtritis Beach, 65 km southwest ofSangiran, and with fresh, large quahogs (Mercenaria) collectedfrom the temperate Wisconsin side of the Mississippi River.These fossil and fresh shells showed a similar fracture mech-anism and produce edge varieties similar to those of the thickclam shells from Florida.
It is possible that observers might correlate double-trackand canal grooves with rodent gnawing due to the flat andbroad morphologies. Cut marks can easily be distinguishedfrom rodents’ gnawing in both micro and macroscopic levels.
Fig. 7. Archaeological cut marks on Grid-D1/21-XI-79. (A) Location of cut marks. (B) Fossil specimen and cut marks. (C) Two wide V grooves. (D) Canal groove.
(E) Tapering canal groove indicating slicing direction from top to bottom. Scale bars: B 1 cm; C,D,E 1 mm.

55K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
A rodent starts gnawing at a focal area and then expands toother areas forming focally aggregated, redundant concentra-tion of the grooves. Rodent gnawing is slow, repeated grindingand grooves occur in pairs of round-bottomed or scooped-outcondition, i.e., monotonous or lacking variations (see [41]:Fig. a,b: 196; [42]: Fig. 21; [53]: Fig. 1e; [56]: Fig. 35;[61]: Figs. 13,18). On the contrary, butchery is an energeticslicing with various cutting edges inflicting different grooves(double-track, canal, and tapering groove) in varying angle(upright or low angled; parallel or sub-parallel), thus creatinggrooves of dissimilar sizes. In particular, the tapering groove isthe best indicator of energetic slicing by hominids. In addition,grooves in SA100879 are concentrated only on flat areaswhere gnawing would have been quite difficult for a rodentwith a small jaw while easily gnawable areas such as theepiphyses, high ridges and bone edges are untouched.
At the microscopic level, the tracks of SA100879, approx-imately 0.2 mm width each, are too narrow to reflect rodent in-cisors (Fig. 5BeD). Likewise, the bone ridge between the twotracks may suggest rodents’ work, but the 0.5e1.2 mmbreadth, including both the tracks and a ridge, is too narrowto indicate a pair of incisors of rodents of any size or taxon.A single incisor of a rodent cannot create the tracks. ‘‘Sincethe upper and lower incisors must match up, the loss of onewill result in the corresponding incisor in the other jaw grow-ing in an abnormally spiral fashion, seriously compromisingthe rodent’s ability to eat’’ [2].
In SA100879, no groove exhibits a U cross-section, the fin-gerprint of a carnivore tooth. Although damaged carnivoreteeth are known to create flat-bottomed grooves [27,53], theabsence of U groove among the 13 grooves should suggest un-likely situation where all the teeth of a carnivore were dam-aged into flat profiles. Moreover, an animal jaw works likea denticulate vice meaning that a carnivore’s ravaging shouldcreate corresponding tooth pitting on the opposite surfaces ofthe bone [14]. Carnivores tend to gnaw heavily on the epiph-yses or high ridges of the bone [26]. It is also known that bluntor large cusps of adult carnivores create irregular groove mar-gins owing to incomplete slicing of the bone lamellae [27].None of these types of damage are seen in SA100879.
Sedimentary or fluvial abrasions are known to smooth thegrooves rather than creating double-track grooves [61].Behrensmeyer et al. [12,13] proposed the most useful criteriato distinguish cut marks from trample damage. Her definitionsstress that trampling in sandy grains inflicts V and U grooveswith associated diffuse scratches, where as the same cause innon-sandy soil creates patches of localized polish on the bonesurface. Also emphasized was that the sub-parallel formationof the grooves at the muscle/tendon attachment areas indicaterepeated involvement of butchery tools.
Haynes [28] demonstrated that elephant trampling couldcreate a single or a cluster of parallel, V grooves like thoseof stone tools. However, his illustrations ([28]: Fig. 4.37,38,40: 160e161) are self-evident that trample marks occurwhere cut marks are rarely expected: e.g., on the head of thefemur, not around the neck of a femur; on the centrum(intervertebral) plate, not around the proximal/distal end of
the vertebral body or surfaces of the lamina, spinous/trans-verse processes; and on the surface of the wide, flat occipitalbone, not around the occipital fossae. Extremely long groovessurrounded by fine, parallel striations on the occipital surfacecover the area as large as a human palm. No experienced eyeswould interpret these as cut marks. There is no report of eitherdouble-track or canal groove. Even other scholars’ observa-tions of trampling [22,48,49] do not find double-track or canalgrooves but commonly emphasize occurrence of V and Ugrooves in association with unidirectional striations or ran-domly oriented superficial scratches dominantly on the shaft.
The upper section of the Pucangan Formation, as the blackcolor indicates, is organic-matter based lacustrine clay whereBehrensmeyer’s localized polish is expected under trampling.The excavators fine-combed the remains to recover fragmentsas small as broken shark teeth. Neither single complete/brokenshells nor shell beds were detected in Level 4 [1]. There aretwo natural beds of fossil gastropods and bivalves, smallerthan a thumbnail, in Levels 3 and 5 [35]. Yet, cutting edgesof those small shells are too fragile to mimic the grooves onSA100879. Pucangan Formation is free of lithic materials.We found no water channel(s) that might have transportedjumbles of lithic materials and broken shells into the PucanganFormation in Sangiran.
The source and taxonomy of the thick clam shell that in-flicted these cut marks are currently unclear. On occasions dur-ing survey, local peasants solicited us to purchase fossil shellsof all sizes for a small cash return. It appears that local inhab-itants seem to have the first access to the large shells becauserice paddy extensively occupies the deposits of the PucanganFormation in Sangiran.
The micro and macroscopic details of the grooves indicatehominids’ intentional defleshing of the biceps muscle off theradius using tools of thick clamshell flakes. The age of thespecimen is estimated to be between 1.6 and 1.5 myr by brack-eting two 40Ar/39Ar dates between the Lower Lahar, 1.66 �0.04 myr [58], and the basal unit of the Kabuh Formation,1.51 � 0.08 myr [38]. This fossil is currently curated at theGRDC, Bandung, Indonesia.
4.2. Fossil: GRID-D1/21-XI-79
During a survey at the Bukuran in Sangiran in 1979,Mr. Budisampoerno [18] of the UGM excavated approxi-mately 50 faunal fossils from 50 cm below the contact surfacebetween the Pucangan and Kabuh Formations. One of thesefossils had cut marks. The specimen (GRID-D1/21-XI-79)shows a total of five canal and wide V grooves arranged insub-parallel formation in approximately 1.0-cm intervals atthe posterio-medial border of the right mandibular condylarneck of a bovid (Bos sp.) (Fig. 7A,B).
As with SA100879, absence of U groove makes carnivoregnawing unlikely. Given the anatomical location, a carnivoreshould have created conspicuous pittings and scorings on thelateral and medial surfaces of the mandibular condyle [14]rather than on the ridge only. Grooves are arranged as a seriesof isolated, single grooves without focally aggregated,

56 K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
redundant grooves, typical of rodent damage. Two grooves inFig. 7C, each 0.3 mm wide, are arranged within 1 mm whileone groove is at a higher location than the other. This wouldrequire two separate gnawings by a single rodent incisor atbest rather than two sets of incisors. If the 2.0-mm wide groovein Fig. 7E were created by two sets of incisors, then all othergrooves, less than 1 mm wide, should be the by-products ofa single incisor. Again, rodents’ gnawing with a single set of in-cisors is infeasible and can be ruled out in this case.
Absence of smoothened grooves eliminates the possibilityof sedimentary or fluvial abrasion [61]. As with SA100879,trampling on this specimen in clay should have createdpatches of polish [12,13] rather than broad canal grooves atthe muscle attachment in sub-parallel orientation. Neither uni-directional striation nor polish is detected along any grooves,eliminating the possibility of trampling. Neither a shell flakenor natural shell bed is known near this specimen. It isextremely unlikely that thick shell flakes could have acciden-tally scratched several times only at the muscle attachment incriss-cross angle at two different stratigraphic positions. Wesuspected mimicking of double-track or canal grooves bynatural agents. However, there is no publication of bonedamage that reports such grooves.
A better explanation is that the grooves in Fig. 7C are twowide V grooves created during tilted slicing by a hominid toutilize the blunt cutting edges available at the cortex or innerlayer of a clamshell flake (Fig. 6C,H). Other adjacent groovesare flat and wide canal grooves with clear groove marginsgouged by a blunt edge. Judging from the smooth bottomsand clearly defined groove margins, these grooves matchexactly with the irregularly fractured blunt edges describedearlier (Fig. 6D,I,L). In particular, the presence of a taperingcanal groove (Fig. 7E) at the narrow ridge of the condylarneck is a clear indication of hominids’ vigorous defleshingof the pterygoyd muscle using thick clamshell flakes ratherthan rodents’ repeated gnawing, trampling or accidentalscratching. The age of GRID-D1/21-XI-79 is similar toSA100879, between 1.6 and 1.5 myr, but relatively youngerthan SA100879 (Fig. 2). GRID-D1/21-XI-79 is currentlystored at the UGM, Yogyakarta, Indonesia.
5. Implications
Among the research collections, we only find cut marks ofthick clamshell flakes. None of the other raw materials amongthe 12 we tested created similar grooves. Our experimentalbutchery revealed the performance of thick clamshell flakesto be nearly as efficient as rhyolite of high-quality flint. Onthe contrary, limestone of the Kalibeng and andesite of theLower Lahar did not make tools superior to those of thenon-lithic materials we tested. It supports feasibility of thickclamshell flakes for primary butchery tool.
It was above Tuff-9, after a hiatus caused by marine impact,that the mammals of estuarine, marsh and terrestrial compo-nents returned to Sangiran, probably together with early mem-bers of H. erectus [1,67]. Fossil SA100879 was excavatedfrom the deposit between Tuff-9 and -10, while GRID-D1/
21-XI-79 originated from the terminal section of the forma-tion. Both specimens are solidly associated with the hominidsand mammals. Stratigraphic positions of the allegedly earlierfossil H. erectus such as Sangiran 27 and 31, dated at1.6 myr [66], are ambiguous due to insufficient geologicaldetails.
We assign early H. erectus in Sangiran as the responsibleagent for those cut marks. A recent study has suggested prob-able use of lithic and non-lithic tools by hominins before earlyHomo in Africa [50]. However, neither Orangutan nor otherprimates in Southeast Asia are believed to be capable of butch-ery. There is no hominid species that predates H. erectus in theregion. The thick clamshell tools that imprinted on two bovidfossils appear to be the first tools in Sangiran and substantiateH. erectus’s use of non-lithic tools.
Our new evidence permits addressing H. erectus’s mode ofdiet. We expect that hominids’ meat extraction would createcut marks exclusively at the muscle attachments and highmeat areas. Dismembering a carcass solely for the technolog-ical use of bones would inflict cut marks predominantly at thearticular surfaces. Skinning would leave cut marks dependingon the areas and size of the removed, e.g., on the non-meatymetapodials [53,82] or phalanges [14]. Occurrence of cutmarks only at the muscle attachments on SA100879 andGrid-D1/21-XI-79 indicates hominids’ butchery for meatextraction in contradiction to Puech’s vegetarian perspective.It reinforces the perspective that meat eating was a widespreadmode of subsistence among the Pleistocene hominids andplayed a significant role in the development of early humancultures [19,31].
Anthropologists tend to agree that stone tool use is a significantsign of humanness [39,46,58,84]. It is also the norm to equatetools with biological species of hominids, e.g., early Homo withthe Oldowan tools, H. erectus with the Acheulean hand-axe andlithic chopper-chopping tools in East Asia, European Neander-thals with the Mousterian tools, and the anatomically modern hu-mans with the elongated blades [15,37,39,58]. However, scholars[16,17,30,51,52,57,76,83] in favor of non-lithic adaptation havelong been pointing out that such convention may work outsideSoutheast Asia where stone tools dominate the (Plio)-Pleistocenearchaeological assemblages. Non-lithic artifacts appear as late as0.5 myr sincewooden spears have emerged at the sites of Clacton-on-Sea in England [47] and Shoningen in Germany [68]. The1.6e1.5 myr age of our evidence indicates the oldest archaeolog-ical evidence of using shell tools in the world. It is the firstevidence of H. erectus’s use of non-lithic tool as opposed to thestone tool-centered convention. Based on the limited and indirectevidence of cut marks, however, we neither argue for pan-Javanuse of shell tools by early H. erectus nor preclude use of stonetools alongside non-lithic tools.
6. Conclusion
New evidence of shell tool use suggests that there was a non-lithic tool adaptation among the early members of H. erectus inSangiran during 1.6e1.5 myr. This suggests that shell tools ofSangiran are a significantdbut often misseddcharacteristic

57K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
of early human adaptations parallel to the stone tool traditions.This may explain why stone tools are lacking at the PucanganFormation in Sangiran. This research demonstrates that archae-ologists must continue searching for evidence of non-lithic tooluse in the region and around the world.
Acknowledgments
We thank the following individuals in random order:Dr. Hasan Muarif Ambary and Dr. Truman Simanjuntak ofthe Pusat Penelitian Arkeologi Nasional-Jakarta; Ibu Wati ofthe LIPI (Indonesian Institute of Sciences); Mr. I Gusti NgurahAnom of the Protectoria and Development of Historical andArchaeological Heritage; Mr. Surotmo and Mr. Gunadi ofthe Sangiran Field Museum; Dr. Fachroel Aziz of the GRDCand Quaternary Geologi Laboratorium; Dr. Tony Djubiantonoof the Balai Arkeologi-Bandung; Mr. Tobing of the MusiumGeologi; Dr. Harry Widianto, Dr. Goenadi and Dr. Nurhadiof the Balai Arkeologi-Yogyakarta; Professor Teuku Jacob,Dr. Agu Suprijo, Mr. Koeshardjono and Ms. Neni Rahmawatiof the Bio- and Paleoanthropologi Laboratotium, UGM;Dr. John de Vos and Mr. Reinier van Zelst of the Nationaal Na-tuurhistorisch Museum; Dr. Jens L. Franzen and Dr. ChristineHertler of the Naturmuseum und Forschungsinstitut Sencken-berg; Dr. Hans-Peter Schultz and Dr. Wolf-Dieter Heinrich ofthe Museum fur Naturkunde, Humboldt University; ProfessorJ. Mark Kenoyer, Professor Henry T. Bunn, Professor T. Doug-las Price, Professor Richard J. Sherwood, Dr. James H. Burton,Heidi Barnhill and Gwendolyn Kelly of the University of Wis-consineMadison; Professor Vance T. Holliday of the ArizonaState University. The NSF (US) and the American BambooSociety funded this research.
References
[1] M. Aimi, F. Aziz, Vertebrate fossils from the Sangiran dome, Mojokerto,
Trinil and Sambungmacan areas, in: N. Watanabe, D. Kadar (Eds.),
Quaternary Geology of the Hominid Fossil Bearing Formations in
Java, GRDC Special Publication No. 4, Bandung, 1985, pp. 155e198.
[2] D. Alderton, Rodents of the World, Facts On File, New York, 1996.
[3] G.J. Bartstra, Contributions to the Study of the Paleolithic Pajitan
Culture, Java, Indonesia. Part 1, E.J. Brill, Leiden, 1976.
[4] G.J. Bartstra, Note on new data concerning the fossil vertebrates and
stone tools in the Walanae Valley in South Sulawesi (Celebes), Modern
Quaternary Research in Southeast Asia 4 (1978) 71e72.
[5] G.J. Bartstra, Homo erectus erectus: the search for his artifacts, Current
Anthropology 23 (1982) 318e320.
[6] G.J. Bartstra, Sangiran, the stone implements of Ngebung, and the Paleo-
lithic Java, Modern Quaternary Research in Southeast Asia 9 (1985)
99e113.
[7] G.J. Bartstra, Late Homo erectus or Ngandong man of Java, Palaeohisto-
ria 29 (1987) 1e7.
[8] G.J. Bartstra, Basoeki, Recent work on the Pleistocene and the Paleo-
lithic of Java, Current Anthropology 30 (1989) 241e244.
[9] G.J. Bartstra, Basoeki, B. Santosa Azis, Solo Valley Research 1975: Java,
Indonesia, Modern Quaternary Research in Southeast Asia 2 (1976)
23e36.
[10] G.J. Bartstra, S. Soegondho, A. van der Wijk, Ngandong man: age and
artifacts, Journal of Human Evolution 17 (1988) 325e337.
[11] G.J. Bartstra, D.A. Hooijer, B. Kallupa, M.A. Akib, Notes on fossil
vertebrates and stone tools from Sulawesi, Indonesia, and the
stratigraphy of the Northern Walanae Depression, Palaeohistoria
33e34 (1994) 1e18.
[12] A.K. Behrensmeyer, K. Gordon, G. Yanagi, Trampling as a cause of bone
surface damage and pseudo-cutmarks, Nature 319 (1986) 768e771.
[13] A.K. Behrensmeyer, K. Gordon, G. Yanagi, Nonhuman bone modifica-
tion in Miocene fossils from Pakistan, in: R. Bonnichsen, M.H. Sorg
(Eds.), Bone Modification, Center for the Study of the First Americans.
Institute for Quaternary Studies, University of Maine, Orono, Maine,
1989, pp. 99e120.
[14] L.R. Binford, Bones: Ancient Men and Modern Myths, Academic Press,
New York, 1981.
[15] F. Bordes, Physical evolution and technological evolution in man: a par-
allelism, World Archaeology 3 (1971) 1e5.
[16] P.I. Boriskovskii, Vietnam in Primeval Times: Part I, Soviet Anthropol-
ogy and Archaeology 7 (1968) 14e32.
[17] P.I. Boriskovskii, Vietnam in Primeval Times: Part II, Soviet Anthropol-
ogy and Archaeology 7 (1968e69) 3e19.
[18] Budisampoerno, Analisasi Temuan Ekskavasi Sangiran 1979e1986 (in
Indonesian) Manuscript on File, Laboratory of Bioanthropologi and
Paleoanthropologi, UGM, Yogyakarta, 1986.
[19] H.T. Bunn, Archaeological evidence for meat-eating by Plio-Pleistocene
hominids from Koobi Fora and Olduvai Gorge, Nature 291 (1981) 574e577.
[20] H.T. Bunn, et al., FxJj 50: an early Pleistocene site in Northern Kenya,
World Archaeology 12 (1980) 109e136.
[21] C. Falgueres, dating layers and fossils in Sangiran dome: method and
results, in: T. Simanjuntak, B. Prasetyo, R. Handini (Eds.), Sangiran:
Man, Culture, and Environment in Pleistocene Times, Yayasan Obor
Indonesia, Jakarta, 2001, pp. 309e319.
[22] A.R. Fiorillo, An experimental study of trampling: implications for the
fossil record, in: R. Bonnichsen, M.H. Sorg (Eds.), Bone Modification,
Center for the Study of the First Americans. Institute for Quaternary
Studies, University of Maine, Orono, Maine, 1989, pp. 61e71.
[23] G.C. Frison, The Glenrock Buffalo Jump, 48CO304, Plains Anthropolo-
gist 15, Part 2, Memoir 7 (1970) 1e66.
[24] G.C. Frison, Prehistoric Hunters of the High Plains, second ed.,
Academic Press, New York, 1991.
[25] I.C. Glover, E.A. Glover, Pleistocene flakes stone tools from Timor and
Flores, Mankind 7 (1970) 88e90.
[26] G. Haynes, Evidence of carnivore gnawing on Pleistocene and recent
mammalian bones, Paleobiology 6 (1980) 341e351.
[27] G. Haynes, A guide for differentiating mammalian carnivore taxa respon-
sible for gnawing to herbivore limb bones, Paleobiology 9 (1983) 164e172.
[28] G. Haynes, Mammoths, Mastodonts, and Elephants: Biology, Behavior,
and the Fossil Record, Cambridge University Press, Cambridge, 1991.
[29] C.B. Heiser, Seed to Civilization: The Story of Food, Harvard University
Press, Cambridge, 1990.
[30] K.L. Hutterer, Reinterpreting the Southeast Asian Paleolithic, in:
J. Allen, J. Golson, R. Jones (Eds.), Sunda and Sahul: Prehistoric Studies
in Southeast Asia, Melanesia and Australia, Academic Press, New York,
1977, pp. 31e71.
[31] G.L. Isaac, D.C. Crader, To what extent were early hominids carnivorous?:
an archaeological perspective, in: R.S. Harding, G. Teleki (Eds.),
Omnivorous Primates, Columbia University Press, New York, 1981,
pp. 37e103.
[32] M. Itihara, et al., Geology and stratigraphy of the Sangiran Area. Quater-
nary geology of the hominid fossil bearing formations in Java, in:
N. Watanabe, D. Kadar (Eds.), Quaternary Geology of the Hominid
Fossil Bearing Formations in Java, GRDC Special Publication No. 4,
Bandung, 1985, pp. 11e43.
[33] T. Jacob, The Pithecanthropus of Indonesia: phenotype, genetics and
ecology, in: L. Konigsson (Ed.), Current Argument on Early Man, Perga-
mon Press, New York, 1980, pp. 170e179.
[34] T. Jacob, R.P. Soejono, L.G. Freeman, F.H. Brown, Stone tools
from Mid-Pleistocene sediments in Java, Science 202 (1978)
885e887.
[35] A.P. Kadar, T. Shibasaki, F. Aziz, Bukuran site, in: N. Watanabe,
D. Kadar (Eds.), Quaternary Geology of the Hominid Fossil Bearing

58 K. Choi, D. Driwantoro / Journal of Archaeological Science 34 (2007) 48e58
Formations in Java, GRDC Special Publication No. 4, Bandung, 1985,
pp. 75e77.
[36] S.G. Keates, G.J. Bartstra, Island migration of early modern Homo sapi-
ens in Southeast Asia: the artifacts from the Walanae Depression, Sula-
wesi, Indonesia, Palaeohistoria 33e34 (1994) 19e30.
[37] R.G. Klein, The Human Career: Human Biological and Cultural Origins,
second ed. University of Chicago Press, Chicago, 1999.
[38] R. Larick, et al., Early Pleistocene 40Ar/39Ar ages for Bapang Formation
hominins, Central Jawa, Indonesia, Proceedings of the National Acad-
emy of Science, USA 98 (2001) 4866e4871.
[39] L.S.B. Leakey, Adam’s Ancestors: the Evolution of Man and His Culture,
Harper and Row Publishers, New York, 1960.
[40] H.A. Lowenstam, S. Weiner, On Biomineralization, Oxford University
Press, New York, 1989.
[41] L.R. Lyman, Vertebrate Taphonomy, Cambridge University Press, London,
1994.
[42] J.M. Maguire, D. Pemberton, M.H. Collette, The Makapansgat
limeworks grey breccia: hominids, hyenas, hytricids or hillwash? Paleon-
tologia Africana 23 (1980) 75e98.
[43] J. Maringer, T. Verhoeven, Die steinartefakte aus der Stegodon-
Fossilschicht von mengeruda auf Flores (in German), Anthropos 65
(1970) 229e247.
[44] J. Maringer, T. Verhoeven, Die oberflachenfunde aus dem fossilgebiet
von Mengeruda und Olabula auf Flores (in German), Indonesien, Anthro-
pos 65 (1970) 530e546.
[45] M. Morwood, P. O’Sullivan, F. Aziz, A. Raza, Fission-track ages of stone
tools and fossils on the east Indonesian islands of Flores, Nature 392
(1998) 173e176.
[46] K.P. Oakley, Man the tool-maker, University of Chicago Press, Chicago, 1961.
[47] K.P. Oakley, P. Andrews, L. Keeley, A Reappraisal of the Clacton Spear-
point, Proceedings of the Prehistoric Society 43 (1977) 13e30.
[48] J.S. Oliver, Analogues and site context: bone damages from Shield Trap
Cave, in: R. Bonnichsen, M.H. Sorg (Eds.), Bone Modification, Center
for the Study of the First Americans. Institute for Quaternary Studies,
University of Maine, Orono, Maine, 1989, pp. 73e98.
[49] S.L. Olsen, P. Shipman, Surface modification on bone: trampling versus
butchery, Journal of Archaeological Science 15 (1988) 535e553.
[50] M.A. Panger, A.S. Brooks, B.G. Richmond, B. Wood, Older than the
Oldowan? Rethinking the emergence of hominin tool use, Evolutionary
Anthropology 11 (2002) 235e245.
[51] Pham Huy Thong, Our stone age: from the Mount Do industry to the
Hoabinh industry, Vietnamese Studies 12 (1978) 9e49.
[52] G. Pope, The Antiquity and Paleoenvironment of the Asian Hominidae,
in: R. Whyte (Ed.), The Evolution of the East Asian Environment,
University of Hong Kong, Hong Kong, 1984, pp. 827e847.
[53] R. Potts, P. Shipman, Cutmarks made by stone tools on bones from
Olduvai Gorge, Tanzania, Nature 29 (1981) 577e580.
[54] P.F. Puech, Tooth wear, diet, and the artifacts of Java man, Current
Anthropology 24 (1983) 381e382.
[55] S. Sartono, The age of thevertebrate fossil and artefacts from Cabenge in South
Sulawesi, Modern Quaternary Research in Southeast Asia 5 (1979) 65e82.
[56] J.J. Saunders, Late Pleistocene Vertebrates of the Western Ozark High-
land, Missouri, Illinois State Museum Reports of Investigations 33
(1977) 1e118.
[57] K. Schick, The Movius line reconsidered: Perspectives on the Earlier
Paleolithic of Eastern Asia, in: R. Corruccini, R. Ciochon (Eds.), Integra-
tive Paths to the Past: Paleoanthropological Advances in Honor of
F. Clark Howell, Prentice Hall, Englewood Cliffs, 1994, pp. 569e596.
[58] K. Schick, N. Toth, Making Silent Stones Speak, Simon and Schuster,
New York, 1993.
[59] F. Semah, H. Saleki, G. Falgueres, G. Feraud, T. Djubiantono, Did Early
Man reach in Java during the Late Pliocene? Journal of Archaeological
Science 27 (2000) 763e769.
[60] F. Semah, A.-M. Semah, T. Djubiantono, H. Simanjuntak, Did they also
make stone tools? Journal of Human Evolution 23 (1992) 439e446.
[61] P. Shipman, J. Rose, Early hominid hunting, butchering, and carcass-
processing behaviors: approaches to the fossil record, Journal of Anthro-
pology and Archaeology 2 (1983) 57e98.
[62] T. Simanjuntak, F. Semah, A new insight into the Sangiran Flake Indus-
try, IPPA Bulletin 14 (1996) 22e26.
[63] R.P. Soejono, New data on the Paleolithic industry in Indonesia, 1st Con-
gress International de Paleontologie Humaine, Nice, 1982.
[64] R.P. Soejono, Remarks on the development and problems of the Paleo-
lithic in Indonesia, in: T. Simanjuntak, B. Prasetyo, R. Handini (Eds.),
Sangiran: Man, Culture, and Environment in Pleistocene Times, Yayasan
Obor Indonesia, Jakarta, 2001, pp. 143e153.
[65] Y. Sondaar, G.D. Van den Bergh, B. Mubroto, F. Aziz, J. de Vos,
U.L. Batu, Middle Pleistocene faunal turnover and colonization of Flores
(Indonesia) by Homo erectus, 319, Centre Recherche Academie Sciences,
Paris, 1994, pp. 1255e1262.
[66] C. Swisher, G. Curtis, T. Jacob, A. Getty, A. Suprijo, Widiasmoro, Age of
the Earliest Known Hominids in Java, Indonesia, Science 263 (1994)
1118e1121.
[67] B. Theunissen, J. de Vos, P.Y. Sondaar, F. Aziz, The establishment of
a chronological framework for the hominid-bearing deposits of Java:
a historical survey, Geological Society of America Special Paper 242
(1990) 39e54.
[68] H. Thieme, Lower Paleolithic hunting spears from Germany, Nature 385
(1997) 807e810.
[69] N. Toth, M. Woods, Molluscan shell knives and experimental cut-marks
on bones, Journal Field Archaeology 16 (1989) 250e255.
[70] H.R. van Heekeren, The Stone Age of Indonesia, second ed., Verhande-
lingen van Het Koninklijk Instituut voor Taal-, Land-, en Volkenkunde,
vol. 61, Martinus Nijhoff, The Hague, 1972, pp. 1e247.
[71] G.H.R. von Koenigswald, Early Paleolithic stone implements from Java,
Bulletin of the Raffles Museum B-1 (1936) 52e60.
[72] G.H.R. von Koenigswald, Introduction in morphology of solo man,
Anthropological Papers of the American Museum of Natural History
43 (Part 3) (1951) 211e221.
[73] G.H.R. von Koenigswald, Lithic industries of Pithecanthropus erectus of
Java, in: F. Ikawa-Smith (Ed.), Early Paleolithic in South and East Asia,
Mouton Publishers, The Hague, 1978, pp. 23e27.
[74] G.H.R. von Koenigswald, A.K. Ghosh, Stone implements from the Trinil
beds of Sangiran, Central Java I, Koninklijke Nederlandse Akademie van
Wetenschappen Series B-76 (1973) 1e17.
[75] G.H.R. von Koenigswald, A.K. Ghosh, Stone implements from the Trinil
beds of Sangiran, Central Java II, Koninklijke Nederlandse Akademie
van Wetenschappen Series B-76 (1973) 18e34.
[76] H. Watanabe, The chopper-chopping tool complex of Eastern Asia: an
ethnoarchaeological-ecological reexamination, Journal of Anthropology
and Archaeology 4 (1985) 1e18.
[77] N. Watanabe, D. Kadar, Quaternary Geology of the Hominid Fossil Bear-
ing Formations in Java, GRDC Special Publication No. 4, Bandung,
1985.
[78] J.B. Wheat, The Jurgens Site, Plains Anthropologist 24, Part 2, Memoir
15 (1979) 1e153.
[79] H. Widianto, The perspective on the evolution of Javanese Homo erectusbased on morphological and stratigraphic characteristics, in: T. Simanjuntak,
B. Prasetyo, R. Handini (Eds.), Sangiran: Man, Culture, and Environ-
ment in Pleistocene Times, Yayasan Obor Indonesia, Jakarta, 2001,
pp. 24e45.
[80] H. Widianto, The oldest Homo erectus stone tools in Java: from the
Lower Pleistocene Pucangan Formation in Sangiran, IPPA Bulletin
<http://arts.anu.edu.au/arcworld/ippa/>; 2006 (Abstract accessed on
September 30, 2005).
[81] H. Widianto, B. Toha, T. Simanjuntak, Penelitian situs Sangiran:
Eksistensi artefak pada awal kala Pleistosen Tengah dan stratigrafi enda-
pan teras di atas seri Kabuh dan seri Notopuro (in Indonesian), in: Berita
Penelitian Arkeologi 3, Balai Arkeologi Yogyakarta, Yogyakarta, 1998.
[82] M.C. Wilson, Cut marks and early hominids: evidence for skinning,
Nature 298 (1982) 303.
[83] M. Wolpoff, Human evolution at the peripheries: the pattern at the east-
ern edge, in: P.V. Tobias (Ed.), Human Evolution: Past, Present and
Future, Alan R. Liss, New York, 1985, pp. 355e365.
[84] T. Wynn, W.C. McGrew, An ape’s view of the Oldowan, Man 24 (1989)
383e398.





























