Embed Size (px)
Citation preview

Intern. J. Neuroscience, 117:47–62, 2007Copyright C© 2007 Informa HealthcareISSN: 0020-7454 / 1543-5245 onlineDOI: 10.1080/00207450500535719
CHLORPYRIFOS INCREASES THE LEVELS OFHIPPOCAMPAL NMDA RECEPTOR SUBUNITSNR2A AND NR2B IN JUVENILE AND ADULT RATS
FATIH GULTEKININANC KARAKOYUNRECEP SUTCUEMIN SAVIK
Suleyman Demirel UniversitySchool of MedicineDepartment of BiochemistryIsparta, Turkey
GOKHAN CESUR
Suleyman Demirel UniversitySchool of MedicineDepartment of PhysiologyIsparta, Turkey
HIKMET ORHAN
Suleyman Demirel UniversitySchool of Agriculture, Department of Animal ScienceUnit of BiometryIsparta, Turkey
Received 31 October 2005.This study was supported by Suleyman Demirel University Research Fund.Address correspondence to Fatih Gultekin, Suleyman Demirel Universitesi, Tip Fakultesi,
Biyokimya Anabilim Dali, 32260 Isparta, Turkey. E-mail: [email protected] or [email protected]
47
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

48 F. GULTEKIN ET AL.
NAMIK DELIBAS
Suleyman Demirel UniversitySchool of MedicineDepartment of BiochemistryIsparta, Turkey
The present study investigated the effect of chlorpyrifos on NMDA receptor subunitsNR2A and NR2B in juvenile and adult rats. Chlorpyrifos was administered withthe dose of 40 and 70 mg/kg to juvenile and adult rats, respectively. Chlorpyrifossignificantly inhibited the AChE activity in juvenile and adult rats (p < .05). NR2Aand NR2B levels significantly increased in juvenile and adult rats by chlorpyrifosapplication (p < .05). Increased NR2A and NR2B levels may reflect increasedglutaminergic activity, consequently neuronal damage. In the case of neuronaldamage, learning and memory could be affected negatively even though NR2Aand NR2B increased.
Keywords chlorpyrifos, learning, memory, NR2A, NR2B, rat
INTRODUCTION
Chlorpyrifos (0,0′-diethyl 0-[3,5,6-trichloro-2-pyridyl] phosphorothionate) isan organophosphates insecticide that is used in order to combat insects andarthropods in agricultural industries (Hayes & Laws, 1990; Schardein &Scialli, 1999; Steenland et al., 2000). Children and adults may be exposed toorganophosphate compounds by getting the residues on foods or by contactingthe contaminated surfaces in houses and gardens (Schuh et al., 2002; Zhenget al., 2000). The chlorpyrifos may accumulate in lipid tissues and cause toxicproblems (Bagchi et al., 1995).
The inhibition of acetyl cholinesterase (AChE; EC 3.1.1.7.) bychlorpyrifos-oxon in nerve synapses is one of the main mechanisms in toxicitiesof chlorpyrifos (Icenogle et al., 2004; Kousba et al., 2003). The changes incholinergic neurotransmission are crucial, for the cholinergic pathways havecritical importance in cognitive functions (Dam et al., 1999; Slotkin, 1999).
Acute and chronic organophosphate applications may play a role in thepathogenesis of hepatotoxicity, neurotoxicity, genotoxicity, embryotoxicity,and immunotoxicity by causing oxidative stress in in vivo and in vitro conditions(Abou-Donia et al., 1996; Crumpton et al., 2000; Gultekin et al., 2001;Richardson, 1995). Studies showed that different classes of pesticides includingchlorpyrifos increase the production of reactive oxygen species and causeoxidative tissue damage (Altuntas et al., 2003, 2004; Gultekin et al., 2000,2001).
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 49
The glutamate receptors of mammalians arrange most of the stimulativenerve transmission in central nervous system and contribute to the formationof the link between neurons during nerve system development. They also havea role in synaptic transmission that involve memory and learning functions.
NMDA receptors are parts of the glutamate receptor family. In experi-mental work it was shown that NMDA receptors are widespread in the brain,especially in the CA1 area of the hippocampus (Ozawa et al., 1998).
Learning and memory are complex functions involving many areas ofcentral nerve system including the limbic system. The hippocampus exists inthe limbic system and that the hippocampal formation has an important role instoring the newly obtained information. It was observed that working memorycan not be transformed to long term memory in patients with lesions affectingthe hippocampus (Tamer, 2002). In earlier studies, it was shown that NMDAreceptor system had an important role on memory function (Levin et al., 2003;Tang et al., 1999).
There are studies on the effects of chlorpyrifos on behavior, learning,and memory. It was shown that subcutaneous (sc) application of chlorpyrifosaffects the working memory of rats in acute period and causes impairments oncognitive and motor functions (Pope et al., 1992; Stanton et al., 1994). Thereare many findings on impairment of cognitive and motor functions caused byapplying chlorpyrifos acutely and repeatedly (Bushnell et al., 1994; Carr et al.,2001; Jett et al., 2001; Levin et al., 2001). It was observed that all of theseeffects of chlorpyrifos on cognitive functions differed according to the agesof the rats (Dam et al., 2000; Karanth & Pope, 2003; Mortensen et al., 1996;Slotkin et al., 2001; Zheng et al., 2000).
The question how chlorpyrifos exerts its effects on learning and memoryis still unclear. There are few studies trying to find out the mechanism ofchlorpyrifos on cognitive functions. As far as the present authors investigated,this seems to be the first study assessing the effect of chlorpyrifos on NMDAreceptors. So, the study tried to approach the understanding of a possiblemechanism of chlorpyrifos on learning and memory by assessing NMDAreceptor subunits NR2A and NR2B in juvenile and adult rats.
MATERIAL AND METHODS
Animals
In this study twenty-four male Sprague Dawley rats were used. They werejuvenile (one month) and adult (three months) rats. The rats were fed for two
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

50 F. GULTEKIN ET AL.
weeks under standard light (12 h light:12 h dark), heat (25◦C), ad libitum waterand feed.
The four groups were organized as follows. (1) Juvenile chlorpyrifos group(n = 8): Chlorpyrifos administered subcutaneously with the dose of 40 mg/kg.(2) Juvenile control group (n = 4): One dose of isotonic sodium chloridewas administered subcutaneously with the same volume as chlorpyrifosgroup. (3) Adult chlorpyrifos group (n = 8): Chlorpyrifos was administeredsubcutaneously with 70 mg/kg. (4) Adult control group (n = 4): One doseof isotonic sodium chloride was administered subcutaneously with the samevolume as chlorpyrifos group.
The weights of rats (mean ± standard deviation) in the groups of Juvenilechlorpyrifos, Juvenile control, Adult chlorpyrifos and Adult control were 54 ±9, 48 ± 16, 157 ± 16, and 156 ± 13 g, respectively.
Half the rats in all groups were decapitated on the 2nd day and the other halfon the 14th day of chlorpyrifos application and the experiment was terminated.Rats were decapitated under the anesthetization of 10% kethamine (Alfamine,Alfasan IBV) and 2% xylazin (Alfazyne, Alfasan IBV), which were appliedintramuscularly.
After decapitation, the brain was removed and both hippocampi were dis-sected, washed in ice-cold phosphate buffered saline (PBS) (50 mM), and frozenimmediately in a deepfreeze until later use. Plasma samples were obtained bygetting whole blood. All samples were protected under −80◦C till analyses.
Determination of Chlorpyrifos Dose
In the literature, maximum tolerable dose of chlorpyrifos for acute adminis-tration were established as 279 mg/kg and 127 mg/kg for adult and juvenilerats, respectively (Levin et al., 2002; Liu & Pope 1998; Pope et al., 1992). Foradult rats, chlorpyrifos was administered as 250, 200, 150, and 100 mg/kg,respectively. After the death of nearly all rats or badly affected due tochlorpyrifos intoxication, 70 mg/kg dose was determined to apply to adultrats as the tolerable dose. Similarly 125, 100, and 75 mg/kg doses were testedto juvenile rats and finally 40 mg/kg dose was determined to apply to juvenilerats as tolerable dose.
Western Blot Analyses
Antibodies against NR2A and NR2B were purchased from Sigma ChemicalCo. (St. Louis, MO, USA). All other reagents were of analytical grade or thehighest grade available.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 51
After thawing, the hippocampi were classified in pairs; each belonged to adifferent rat in the same group (2 animals/preparation). The paired hippocampiwere homogenized in ice-cold buffer [50 mM Tris-HCl (pH 7.5), 0.15 MNaCl, 1% Triton X-100, 1 mM EDTA, 2 mM EGTA, 25 µg/ml leupeptin,25 µg/ml aprotinin, and 10 µM benzamidine] to obtain 1/5 homogenate usinga glass-teflon homogenizer by 18–20 strokes. Then, the samples sonicated30 s (Bandelin Sonoplus, Germany), and aliquots were taken for proteindetermination. Method of Laemmli was used for SDS-PAGE studies (Laemmli,1970). Equal amounts of protein for each sample (56.75 µg of protein per lane)were separated by SDS/PAGE on 7.5% minigels, blotted electrophoretically toimmobilon membrane, and incubated in Tween-Tris Buffered Saline (TTBS)[50 mm Tris-HCl (pH 7.5–8.0), 150 mM NaCl, and 0.1% Tween 20], containing3% bovine serum albumin (BSA) for 30 min. Blots were incubated overnightwith anti-NR2A (1:3000) or anti-NR2B (1:5000) in 1% BSA. Blots were thensubjected to three additional 10-min washings in TTBS. Blots were incubatedwith alkaline phosphatase conjugated monoclonal antirabbit IgG (1:10000) in1% BSA for one hour at room temperature, and three additional washingswere performed with TTBS for 10 min. The membrane was incubated in20 ml of fresh reagent solution (BCIP/NBT), until color development. Imagesof immunoblots were analyzed with a computerized image analysis system(KODAK Image Station 2000 MM Multimodal Imaging System, USA). SDS-PAGE and Western Blot analyses were done on 3 independent hippocampuspreparations (2 animals/group).
Measurement of Acetylcholine Esterase
The levels of plasma AChE activation was determined in an auto analyzer(Roche/Hitachi Modular P800, Japan) by using a commercial kit (Roche,Japan).
Statistical Analyses
Statistical evaluations were performed by using “SPSS 11.0 for Windows”packet program. In general any significant differences between these groupswere evaluated by using Kruskal-Wallis test. Mann Whitney-U test was usedin order to compare the groups with each other. The results of Western Blotanalyses were given as mean ± standard deviation. Value of the “p” < .05 wasaccepted as significant.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

52 F. GULTEKIN ET AL.
Figure 1. AChE activities of adult rats. Results were expressed as mean ± standard devi-ation. ∗Significantly different groups compared to controls, p < .05, Mann Whitney-U Test.űSignificantly different groups within days, p < .05, Mann Whitney-U Test.
RESULTS
During the experiment period, behavior and general conditions of animalswere observed. No cholinergic toxicity symptoms such as diarrhea, tiredness,involuntary muscle contractions, respiratory muscle paralyze were observed inchlorpyrifos administered to the juvenile and adult rats. None of the rats diedduring the experiment period.
The results of the plasma AChE levels were shown in Figures 1 and 2.Chlorpyrifos significantly inhibited the AChE activity in juvenile and adultrats in 2nd day of application (p < .05). In juvenile chlorpyrifos group, AChEactivity was still lower than control group in day 14, but the difference was notsignificant (p > .05). However, in adult rats, AChE activity was still significantlylower in chlorpyrifos group compared with control group in day 14 (p < .05).
To evaluate protein concentrations of NMDA receptors Western Blotanalyses were done on hippocampal homogenates. Western Blot analysis ofNR2A and NR2B were shown in Figures 3–6.
The density of the protein band in the control group was accepted as 100%,and data from other groups were calculated as percentages of the control value.In juvenile rats the NR2A and NR2B levels did not significantly change in day2 (p > .05) whereas these levels significantly increased in day 14 (p < .05). Inadults, both of these two receptor subunits levels significantly increased in day
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.
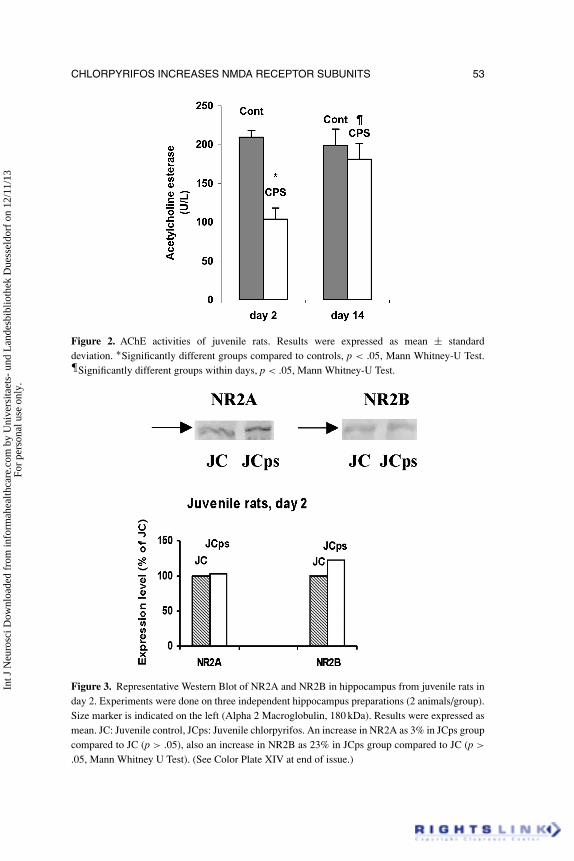
CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 53
Figure 2. AChE activities of juvenile rats. Results were expressed as mean ± standarddeviation. ∗Significantly different groups compared to controls, p < .05, Mann Whitney-U Test.űSignificantly different groups within days, p < .05, Mann Whitney-U Test.
Figure 3. Representative Western Blot of NR2A and NR2B in hippocampus from juvenile rats inday 2. Experiments were done on three independent hippocampus preparations (2 animals/group).Size marker is indicated on the left (Alpha 2 Macroglobulin, 180 kDa). Results were expressed asmean. JC: Juvenile control, JCps: Juvenile chlorpyrifos. An increase in NR2A as 3% in JCps groupcompared to JC (p > .05), also an increase in NR2B as 23% in JCps group compared to JC (p >
.05, Mann Whitney U Test). (See Color Plate XIV at end of issue.)
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

54 F. GULTEKIN ET AL.
Figure 4. Representative Western Blot of NR2A and NR2B in hippocampus from juvenile rats inday 14. Experiments were done on three independent hippocampus preparations (2 animals/group).Size marker is indicated on the left (Alpha 2 Macroglobulin, 180 kDa). Results were expressed asmean. JC: Juvenile control, JCps: Juvenile chlorpyrifos. The asterisks indicate significant changescompared to the control group, p < .05, Mann Whitney U Test. An increase in NR2A as 36% inJCps group compared to JC (p < .05), also an increase in NR2B as 31% in JCps group comparedto JC (p < .05, Mann Whitney U Test). (See Color Plate XV at end of issue.)
2 (p < .05). In day 14, whereas NR2B levels were still significantly high (p <
.05), NR2A levels decreased to control levels (p > .05) in adults.
DISCUSSION
Acetylcholine Esterase
The basic mechanism of chlorpyrifos toxicity is the inhibition of AChE bychlorpyrifos-oxon in nerve synapses (Schuh et al., 2002). In this study it wasdetermined that chlorpyrifos significantly reduced AChE activity in juvenilerats in day 2, whereas in adult rats in day 2 and day 14. The reduced AChEactivities showed that the dose of chlorpyrifos applied during the experimentwas effective. ACh accumulates in nerve synapses due to decreased AChEactivity; consequently chlorpyrifos toxicity occurs.
It was observed that the AChE activity that was reduced in juvenileand adult rats by chlorpyrifos application significantly increased in day 14comparing with day 2. This increase may indicate a reduction of AChEinhibition day by day or an increase of AChE synthesis. As a result of this,
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.
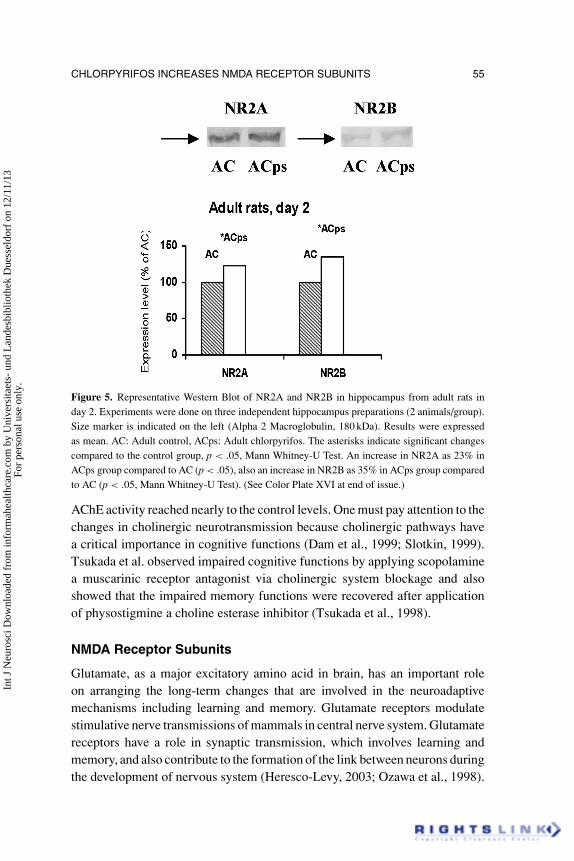
CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 55
Figure 5. Representative Western Blot of NR2A and NR2B in hippocampus from adult rats inday 2. Experiments were done on three independent hippocampus preparations (2 animals/group).Size marker is indicated on the left (Alpha 2 Macroglobulin, 180 kDa). Results were expressedas mean. AC: Adult control, ACps: Adult chlorpyrifos. The asterisks indicate significant changescompared to the control group, p < .05, Mann Whitney-U Test. An increase in NR2A as 23% inACps group compared to AC (p < .05), also an increase in NR2B as 35% in ACps group comparedto AC (p < .05, Mann Whitney-U Test). (See Color Plate XVI at end of issue.)
AChE activity reached nearly to the control levels. One must pay attention to thechanges in cholinergic neurotransmission because cholinergic pathways havea critical importance in cognitive functions (Dam et al., 1999; Slotkin, 1999).Tsukada et al. observed impaired cognitive functions by applying scopolaminea muscarinic receptor antagonist via cholinergic system blockage and alsoshowed that the impaired memory functions were recovered after applicationof physostigmine a choline esterase inhibitor (Tsukada et al., 1998).
NMDA Receptor Subunits
Glutamate, as a major excitatory amino acid in brain, has an important roleon arranging the long-term changes that are involved in the neuroadaptivemechanisms including learning and memory. Glutamate receptors modulatestimulative nerve transmissions of mammals in central nerve system. Glutamatereceptors have a role in synaptic transmission, which involves learning andmemory, and also contribute to the formation of the link between neurons duringthe development of nervous system (Heresco-Levy, 2003; Ozawa et al., 1998).
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

56 F. GULTEKIN ET AL.
Figure 6. Representative Western Blot of NR2A and NR2B in hippocampus from adult rats in day14. Experiments were done on three independent hippocampus preparations (2 animals/group).Size marker is indicated on the left (Alpha 2 Macroglobulin, 180 kDa). Results were expressedas mean. AC: Adult control, ACps: Adult chlorpyrifos. The asterisks indicates significant changescompared to the control group, p < .05, Mann Whitney-U Test. A decrease in NR2A as 1% inACps group compared to AC (p > .05), also an increase in NR2B as 14% in ACps group comparedto AC (p < .05, Mann Whitney U Test). (See Color Plate XVII at end of issue.)
NMDA receptors, as members of ionotropic glutamate receptor family,include cation-specific ion channels. It was shown in previous studies thatNMDA receptor system has an important role on memory functions. Itwas determined that dizocilpine an NMDAR antagonist makes impairmenton memory functions like hippocampectomy (Levin et al., 2003). It wasdetermined that excessive expression of NR2B receptors, localized in frontof the brain, caused increase in activation of NMDA receptor activity andcognitive performance (Tang et al., 1999).
There are many studies about the effects of chlorpyrifos on behavior andmemory. It was shown that chlorpyrifos treatment resulted in impairment inshort-term memory in a dose dependent manner and this effect returned tothe normal in a long time period in adult rats (Stanton et al., 1994). Popeet al. determined behavioral changes lasting a long time in adult rats followingchlorpyrifos application even though neurochemical parameters were regainedin a short time (Pope et al., 1992). Stanton et al. showed that chlorpyrifos caused
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 57
selective memory impairment. They conclude that the neurochemical effects ofchlorpyrifos application are transient and that the behavioral effects in juvenilerats, compared to adult rats, are mild and lasting a short time period (Stantonet al., 1994). Levin et al. determined using T-maze and 16 armed radial armmaze that chlorpyrifos application forms sex related and long time changes incognitive function (Levin et al., 2002).
Studies on the underlying mechanism of the effects of chlorpyrifos onlearning and memory are limited. Nostrandt et al. comparatively evaluated theeffects of chlorpyrifos on cholinesterase inhibition and behavioral effects. Theyreported that after the application of different doses of chlorpyrifos, behavioralchanges did not accompany cholinesterase inhibition till a trigger point. Afterthat dose, behavioral effects were observed by increasing inhibition level ofthe enzyme activity (Nostrandt et al., 1997). The present authors were unableto find any relevant study that shows whether the effects of chlorpyrifos onlearning and memory occur via NMDA receptors. This study tried to makean approach to a possible involvement of NMDA receptors in alterations ofcognitive functions caused by chlorpyrifos.
As a result of this study, it has been found that in juvenile rats the levels ofNR2A and NR2B receptor subtypes increased by day 14. It has been observedthat in adult rats the levels NR2B receptor subtypes increased on day 2 and day14 whereas NR2A increased only on day 2.
This might lead to the conclusion that the increased NR2A and NR2Breceptors levels can be related to the stimulated cholinergic activity. ACh andglutamate have many interactions in many nerve system regions includinghippocampal afferent and efferent fibers. When this relationship is consideredin terms of receptors, it is seen that NMDA receptors can localize withmuscarinic M1 receptors. ACh arranges the glutamatergic nerve transmissionpresinaptically and postsinaptically, whereas glutamate mediated NMDAreceptor activation causes ACh release (Tsukada et al., 1998). Pavlovsky et al.showed that pyridostigmine an AChE inhibitor increased the ACh levels innerve synapses as a result of enzyme inhibition and this increased ACh levelssubsequently stimulated the spontaneous glutamate release (Pavlovsky et al.,2003).
Chlorpyrifos, applied to rats in this experimental study, seems to increasethe cholinergic activity by AChE enzyme inhibition both in juvenile and adultrats. So it seems that increased ACh interacts with the glutaminergic system.Hsich et al. observed that NR2A mRNA expression would increase by chronicnicotine application. They reported that nicotine excessively would stimulatethe presinaptic α7 ACh receptor, and selectively would increase the NMDA
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

58 F. GULTEKIN ET AL.
mediated excitatory post-synaptic potential. The increased excitatory post-synaptic potential might cause to increase in NR2A mRNA expression (Hsiehet al., 2002).
The increases in receptor levels in juvenile and adult rats in day 2 and day14 were significantly different. Dissimilar reaction of the rats may be attributedto the different doses of chlorpyrifos applied to the juvenile and adult rats.
In the present study, reactions of NR2A and NR2B receptors to chlorpyrifosapplication were not the same in day 14 in adult rats. This might be due to thedifferent sensitivity of receptor subtypes to chlorpyrifos. Wirkner et al. seem tosupport this conclusion when they determined different sensitivities of receptorsubtypes against ethanol (Wirkner et al., 1999).
The increases in NR2A and NR2B level may positively affect learningand memory. It has been shown that extreme expression of NR2B receptorslocalized in front of the brain increased the activation of NMDA receptors andcognitive performance (Tang et al., 1999). In a study performed on autopsyshowed that the quantity and the expressions of NR2A and NR2B were lessin Alzheimer patients (Hynd et al., 2004). In the present study, NR2A andNR2B levels increased by chlorpyrifos treatment. Consequently, it looks likechlorpyrifos contributes positively to learning and memory. However, thereare studies that show the negative effect of chlorpyrifos on learning andmemory (Levin et al., 2002; Pope et al., 1992; Stanton et al., 1994). So, duringthe experiment, the decrease of receptor subunits were expected by contrast.One has to take into consideration the neurotoxic effects of NMDA receptorshyperfunction evaluating the effects of chlorpyrifos on learning and memorythrough changes in NR2A and NR2B levels.
Glutamate causes excitotoxic neuronal damage in the case of extremestimulation of NMDA receptors. Increased NR2A and NR2B levels, found inthe present study, may reflect the increased glutaminergic activity, consequentlyneuronal damage. In the case of neuronal damage, learning and memory couldbe affected negatively, in spite of increase in NR2A and NR2B levels.
CONCLUSION
In juvenile and adult rats, chlorpyrifos caused a decrease in AChE activityand increases in NR2A and NR2B levels. Increased ACh due to AChEinhibition may cause glutamate release. Thus, elevated glutamate releasemay stimulate NR2A and NR2B synthesis by increasing selectively NMDAmediated excitatory postsynaptic potential (EPSP). Due to only NR2A andNR2B levels were evaluated in the present study, determination of the changes
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 59
in other NMDA receptor subunits may help to understand the effects ofchlorpyrifos on NMDA receptors and to interpret the results obtained. However,advanced studies are needed, as mechanisms like the interactions betweencholinergic and neuronal system have not been identified yet in detail.
REFERENCES
Abou-Donia, M. B., Wilmarth, K. R., Abdel-Rahman, A. A., Jensen, K. F., Oehme, F.W., & Kurt, T. L. (1996). Increased neurotoxicity following concurrent exposureto pyridostigmine bromide, DEET, and chlorpyrifos. Fundamental and AppliedToxicology, 34(2), 201–222.
Altuntas, I., Delibas, N., Doguc, D. K., Ozmen, S., & Gultekin, F. (2003). Role of reactiveoxygen species in organophosphate insecticide phosalone toxicity in erythrocytesin vitro. Toxicology in Vitro, 17(2), 153–157.
Altuntas, I., Kilinc, I., Orhan, H., Demirel, R., Koylu, H., & Delibas, N. (2004). Theeffects of diazinon on lipid peroxidation and antioxidant enzymes in erythrocytesin vitro. Human and Experiemental Toxicology, 23(1), 9–13.
Bagchi, D., Bagchi, M., Hassoun, E. A., & Stohs, S. J. (1995). In vitro and in vivogeneration of reactive oxygen species, DNA damage and lactate dehydrogenaseleakage by selected pesticides. Toxicology, 104, 129–140.
Bushnell, P. J., Kelly, K. L., & Ward, T. R. (1994). Repeated inhibition of cholinesteraseby chlorpyrifos in rats: Behavioral, neurochemical and pharmacological indicesof tolerance. Journal of Pharmacology and Experimental Therapeutics, 270(1),15–25.
Carr, R. L., Chambers, H. W., Guarisco, J. A., Richardson, J. R., Tang, J., & Chambers, J.E. (2001). Effects of repeated oral postnatal exposure to chlorpyrifos on open-fieldbehavior in juvenile rats. Toxicological Sciences, 59(2), 260–267.
Crumpton, T. L., Seidler, F. J., & Slotkin, T. A. (2000). Is oxidative stress involved inthe developmental neurotoxicity of chlorpyrifos? Brain Research. DevelopmentalBrain Research, 121(2), 189–195.
Dam, K., Garcia, S. J., Seidler, F. J., & Slotkin, T. A. (1999). Neonatal chlorpyrifosexposure alters synaptic development and neuronal activity in cholinergic andcatecholaminergic pathways. Brain Research. Developmental Brain Research,116(1), 9–20.
Dam, K., Seidler, F. J., & Slotkin, T. A. (2000). Chlorpyrifos exposure during a criticalneonatal period elicits gender-selective deficits in the development of coordinationskills and locomotor activity. Brain Research. Developmental Brain Research,121(2), 179–187.
Gultekin, F., Ozturk, M., & Akdogan, M. (2000). The effect of organophosphateinsecticide chlorpyrifos-ethyl on lipid peroxidation and antioxidant enzymes(in vitro). Archives of Toxicology, 74, 533–538.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

60 F. GULTEKIN ET AL.
Gultekin, F., Delibas, N., Yasar, S., & Kilinc, I. (2001). In vivo changes in antioxidantsystems and protective role of melatonin and a combination of vitamin C andvitamin E on oxidative damage in erythrocytes induced by chlorpyrifos-ethyl inrats. Archives of Toxicology, 75, 88–96.
Gultekin, F., Delibas, N., Kutluhan, S., Akdogan, M., Kilinc, I., & Sutcu, R. (2001).The chances in antioxidant system caused by chlorpyrifos-ethyl in rat brain andprotective effect of melatonin and vitamin C + vitamin E. Selcuk Universitesi TipFakultesi Dergisi, 17, 79–86.
Hayes, W. J., & Laws, E. R. (1990). Handbook of Pesticide Toxicology, Vol. 3: Classesof Pesticides. New York: Academic Press, Inc.
Heresco-Levy, U. (2003). Glutamatergic neurotransmission modulation and themechanisms of antipsychotic atypicality. Progress in Neuropsychopharmacologyand Biological Psychiatry, 27(7), 1113–1123.
Hsieh, C. Y., Leslie, F. M., & Metherate, R. (2002). Nicotine exposure during a postnatalcritical period alters NR2A and NR2B mRNA expression in rat auditory forebrain.Brain Research. Developmental Brain Research, 133(1), 19–25.
Hynd, M. R., Scott, H. L., & Dodd, P. R. (2004). Differential expression of N-methyl-D-aspartate receptor NR2 isoforms in Alzheimer’s disease. Journal ofNeurochemistry, 90(4), 913–919.
Icenogle, L. M., Christopher, N. C., Blackwelder, W. P., Caldwell, D. P., Qiao,D., Seidler, F. J., Slotkin, T. A., & Levin, E. D. (2004). Behavioral alter-ations in adolescent and adult rats caused by a brief subtoxic exposure tochlorpyrifos during neurulation. Neurotoxicology and Teratology, 26(1), 95–101.
Jett, D. A., Navoa, R. V., Beckles, R. A., & McLemore, G. L. (2001). Cognitivefunction and cholinergic neurochemistry in weanling rats exposed to chlorpyrifos.Toxicology and Applied Pharmacology, 174(2), 89–98.
Karanth, S., & Pope, C. (2003). Age-related effects of chlorpyrifos and parathion onacetylcholine synthesis in rat striatum. Neurotoxicology and Teratology, 25(5),599–606.
Kousba, A. A., Poet, T. S., & Timchalk, C. (2003). Characterization of the in vitrokinetic interaction of chlorpyrifos-oxon with rat salivary cholinesterase: A potentialbiomonitoring matrix. Toxicology, 188, 219–232.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature, 227(5259), 680–685.
Levin, E. D., Addy, N., Nakajima, A., Christopher, N. C., Seidler, F. J., & Slotkin,T. A. (2001). Persistent behavioral consequences of neonatal chlorpyrifosexposure in rats. Brain Research. Developmental Brain Research, 130(1),83–89.
Levin, E. D., Addy, N., Baruah, A., Elias, A., Christopher, N. C., Seidler, F. J., & Slotkin,T. A. (2002). Prenatal chlorpyrifos exposure in rats causes persistent behavioralalterations. Neurotoxicology and Teratology, 24(6), 733–741.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

CHLORPYRIFOS INCREASES NMDA RECEPTOR SUBUNITS 61
Levin, E. D., Sledge, D., Baruah, A., & Addy, N. A. (2003). Ventral hippocampal NMDAblockade and nicotinic effects on memory function. Brain Research Bulletin, 61(5),489–495.
Liu, J., & Pope, C. N. (1998). Comparative presynaptic neurochemical changes in ratstriatum following exposure to chlorpyrifos or parathion. Journal of Toxicologyand Environmental Health, 53(7), 531–544.
Mortensen, S. R., Chanda, S. M., Hooper, M. J., & Padilla, S. (1996). Maturational dif-ferences in chlorpyrifos-oxonase activity may contribute to age-related sensitivityto chlorpyrifos. Journal of Biochemical Toxicology, 11(6), 279–287.
Nostrandt, A. C., Padilla, S., & Moser, V. C. (1997). The relationship of oral chlorpyrifoseffects on behavior, cholinesterase inhibition, and muscarinic receptor density inrat. Pharmacology, Biochemistry and Behavior, 58(1), 15–23.
Ozawa, S., Kamiya, H., & Tsuzuki, K. (1998). Glutamate receptors in the mammaliancentral nervous system. Progress in Neurobiology, 54(5), 581–618.
Pavlovsky, L., Browne, R. O., & Friedman, A. (2003). Pyridostigmine enhances glu-tamatergic transmission in hippocampal CA1 neurons. Experimental Neurology,179(2), 181–187.
Pope, C. N., Chakraborti, T. K., Chapman, M. L., & Farrar, J. D. (1992). Long-termneurochemical and behavioral effects induced by acute chlorpyrifos treatment.Pharmacology, Biochemistry and Behavior, 42(2), 251–256.
Richardson, R. J. (1995). Assessment of the neurotoxic potential of chlorpyrifos relativeto other organophosphorus compounds: A critical review of the literature. Journalof Toxicology and Environmental Health, 44(2), 135–165.
Schardein, J. L., & Scialli, A. R. (1999). The legislation of toxicologic safety factors:The Food Quality Protection Act with chlorpyrifos as a test case. ReproductiveToxicology, 13(1), 1–14.
Schuh, R. A., Lein, P. J., Beckles, R. A., & Jett, D. A. (2002). Noncholinesterase mech-anisms of chlorpyrifos neurotoxicity: Altered phosphorylation of Ca2+/cAMPresponse element binding protein in cultured neurons. Toxicology and AppliedPharmacology, 182(2), 176–185.
Slotkin, T. A. (1999). Developmental cholinotoxicants: Nicotine and chlorpyrifos.Environmental Health Perspectives, 107(1), 71–80.
Slotkin, T. A., Cousins, M. M., Tate, C. A., & Seidler, F. J. (2001). Persistent cholinergicpresynaptic deficits after neonatal chlorpyrifos exposure. Brain Research, 902(2),229–243.
Stanton, M. E., Mundy, W. R., Ward, T., Dulchinos, V., & Barry, C. C. (1994).Time-dependent effects of acute chlorpyrifos administration on spatial delayedalternation and cholinergic neurochemistry in weanling rats. Neurotoxicology,15(1), 201–208.
Steenland, K., Dick, R. B., Howell, R. J., et al. (2000). Neurologic functionamong termiticide applicators exposed to chlorpyrifos. Environmental HealthPerspectives, 108(4), 293–300.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.

62 F. GULTEKIN ET AL.
Tamer, D. (2002). Functinal Neuroanatomy. 3rd edn. (pp. 226–231). Ankara: ODTUImprovement Foundation Publishing and Communication Company-Metu Press.
Tang, Y. P., Shimizu, E., Dube, G. R., Rampon, C., Kerchner, G. A., Zhuo, M., Liu,G., & Tsien, J. Z. (1999). Genetic enhancement of learning and memory in mice.Nature, 401(6748), 63–69.
Tsukada, H., Kakiuchi, T., Shizuno, H., & Nishiyama, S. (1998). Interactions ofcholinergic and glutamatergic neuronal systems in the functional activation ofcerebral blood flow response: A PET study in unanesthetized monkeys. BrainResearch, 796(1–), 82–90.
Wirkner, K., Poelchen, W., Koles, L., Muhlberg, K., Scheibler, P., Allgaier, C., & Illes, P.(1999). Ethanol-induced inhibition of NMDA receptor channels. NeurochemistryInternational, 35(2), 153–162.
Zheng, Q., Olivier, K., Won, Y. K., & Pope, C. N. (2000). Comparative cholinergicneurotoxicity of oral chlorpyrifos exposures in preweanling and adult rats.Toxicological Sciences, 55(1), 124–132.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itaet
s- u
nd L
ande
sbib
lioth
ek D
uess
eldo
rf o
n 12
/11/
13Fo
r pe
rson
al u
se o
nly.





























