Embed Size (px)
Citation preview

Molecular & Biochemical Parasitology 133 (2004) 27–35
Chloroquine resistance inPlasmodium chabaudi: arechloroquine-resistance transporter (crt) and multi-drug
resistance (mdr1) orthologues involved?�
Paul Hunta,∗, Pedro V.L. Cravob, Paul Donleavya, Jane M.-R. Carltonc, David Wallikeraa Institute of Cell, Animal and Population Biology, University of Edinburgh, West Mains Road, Edinburgh EH9 3JT, Scotland, UK
b Centro de Malária e Outras Doenças Tropicais/IHMT, Rua da Junqueira, 96, 1349-008 Lisbon, Portugalc The Institute for Genomics Research, 9712 Medical Center Drive, Rockville, MD 20850, USA
Received 19 June 2003; received in revised form 25 August 2003; accepted 25 August 2003
Abstract
We have identified in the rodent malaria parasitePlasmodium chabaudiorthologues of twoPlasmodium falciparumgenes,pfcrt andpfmdr1which have been implicated as determinants of chloroquine resistance in the latter species. The sequences of theP. chabaudigenes,denoted, respectively,pccg10andpcmdr1, were first determined in the chloroquine-sensitive clone AS, and found to be highly similar tothose ofP. falciparum. Forpccg10, there was a nucleotide sequence identity of 68.6% and amino acid sequence identity of 75.1% withinthe predicted coding region. Forpcmdr1, the sequence identities were 75.0% (nucleotide) and 78.1% (amino acid). The sequences of thegenes were then determined in threeP. chabaudiclones selected from clone AS which possessed three different levels of resistance tochloroquine. The sequences of both genes in all mutants were found to be identical to those of the sensitive AS from which they had beenderived. Polymorphic sites were found in both genes between the AS clones and a genetically unrelated sensitive clone AJ. Analysis ofgenetic crosses between AJ and resistant AS clones showed no linkage between inherited parental alleles ofpccrt andpcmdr1and drugresponses of the cloned progeny. This showed that neither of these genes, nor genes closely linked to them, were determinants of thechloroquine resistance in theP. chabaudimutants.© 2003 Elsevier B.V. All rights reserved.
Keywords: Plasmodium chabaudi; Plasmodium falciparum; Drug resistance; Chloroquine; Chloroquine-resistance transporter (crt); Multi-drug resistancegene-1 (mdr1)
1. Introduction
The development and spread of resistance to anti-malarialdrugs continues to obstruct efforts to control the disease.Chloroquine is a highly effective drug which is cheap, safeand easy to distribute. The genetic basis of resistance ofPlasmodium falciparumto chloroquine has, therefore, beena highly active research area for many years.
Chloroquine interferes with the detoxification of haematinin the parasite’s food vacuole. Resistance is associated with
Abbreviations: ATP, adenosine triphosphate; DNA, deoxyribonucleicacid; EDTA, ethylenediamine tetra acetic acid; PCR, polymerase chainreaction; RFLP, restriction fragment length polymorphism; SDS, sodiumdodecyl sulphate
� Note: Nucleotide sequence data reported in this paper forpccg10have been deposited in GenBankTM, submission number: AY304549.
∗ Corresponding author. Tel.:+44-131-650-8661;fax: +44-131-650-6564.
E-mail address:[email protected] (P. Hunt).
reduced accumulation of chloroquine in the vacuole, whichresults from reduced uptake of the drug, increased efflux,or a combination of the two processes[1]. A number ofcandidate genes inP. falciparumhave been proposed to beinvolved in chloroquine resistance, each concerned in mem-brane transport. Most notably, an allele of theP. falciparumchloroquine-resistance transporter gene (pfcrt) containingthreonine at position 76 (pfcrt-76T) has been demonstratedto be strongly linked to chloroquine resistance in the progenyof a cross between resistant and sensitive clones[2]. Trans-fection work has recently confirmed the role of this gene inthis type of resistance inP. falciparum[3]. Field observa-tions have generally confirmed thatpfcrt-76T correlates withboth clinical responses to the drug in malaria patients andin vitro chloroquine responses of cultured isolates[4–8].
Other studies have examined whether there is an associa-tion between chloroquine resistance and polymorphisms inpfmdr1, a gene which encodes Pgh-1, an orthologue of one ofthe P-glycoproteins expressed in multi-drug resistant human
0166-6851/$ – see front matter © 2003 Elsevier B.V. All rights reserved.doi:10.1016/j.molbiopara.2003.08.010

28 P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35
cancer cells and belonging to a superfamily of ATP-bindingcassette (ABC) transporters. Foote et al.[9] claimed thattwo alleles ofpfmdr1were associated with chloroquine re-sistance in isolates ofP. falciparumfrom several countries.However, no association was found between chloroquineresponse andpfmdr1 alleles in a genetic cross[10] or inseveral field studies (e.g.[11,12]). However, more recentwork has suggested that one allele of this gene,pfmdr1-86Y,may confer higher levels of resistance to parasites bearingthe pfcrt-76T allele[4,6]. A second allele,pfmdr1-1034S,-1042N, -1246D, has been shown in transfection studies tomodulate the chloroquine response of a resistant parasite[13].
Recently, analysis of high density polymorphic markersaround thepfcrt gene ofP. falciparumhas suggested thatchloroquine-resistant mutants arose from four foci in SouthAmerica and Southeast Asia[14]. This mode of emergenceis in marked contrast to the presumed multifocal originsof pyrimethamine resistance which arises from single pointmutations, especially 108N, in the gene encoding dihydro-folate reductase (pfdhfr) [15,16].
In view of the growing evidence of a role for bothpfcrtandpfmdr1 in determining chloroquine responses inP. fal-ciparum, we wished to investigate whether orthologues ofthese genes were involved in chloroquine-resistant mutantsof the rodent malaria parasitePlasmodium chabaudi. A lin-eage of genetically related clones resistant to different lev-els of chloroquine derived from a common sensitive cloneis available in this species[17]. The pfmdr1orthologue inP. chabaudi, pcmdr1, has previously been identified and se-quenced[18]. In the current work, we report full sequencesof pcmdr1and ofpccg10, the orthologue of theP. falciparumpfcrt gene, in the related chloroquine-resistant and sensitiveclones ofP. chabaudi. We also examine the relationship ofboth genes to chloroquine resistance in crosses between re-sistant clones and a genetically unrelated sensitive clone.
2. Materials and methods
2.1. Parasites
Plasmodium chabaudiclones were maintained in labo-ratory mice and passaged through mosquitoes (Anophelesstephensi), as described by Walliker et al.[19].
The lineage of theP. chabaudiclones used in this workand their drug sensitivities are shown inTable 1. Briefly, allwere derived from the drug-sensitive AS isolate which wasfirst cloned by limiting dilution, then selected for resistanceto pyrimethamine and re-cloned[19]. This clone, designatedAS(0CQ), was then progressively selected for resistance tochloroquine, yielding AS(3CQ), AS(15CQ) and AS(30CQ)[20,21]. AS(15CQ) was also subjected to further selectionwith mefloquine, yielding AS(15MF/3)[18]. The resistanceof each of these clones was stable after passage through miceand mosquitoes in the absence of the drugs.
Table 1P. chabaudiclones used in this study
Clone Selectedfrom
Chloroquineresponse
Cross withAJ
Reference
AS Sensitive [19]AS(0CQ) AS Sensitive [19]AS(3CQ) AS(0CQ) Resistant (R) Yes [20,22]AS(15CQ) AS(3CQ) Resistant (HR) [21]AS(15MF/3) AS(15CQ) Resistant (HR) Yes [18]AS(30CQ) AS(15CQ) Resistant (VHR) [21]
AJ Sensitive [18–22]
R, resistant; HR, high resistance; VHR, very high resistance.
Clones AS(3CQ) and AS(15MF/3) have been crossed witha genetically distinct clone AJ[18,20,22]. A selection ofcloned recombinant progeny of two of these crosses[18,22]were used in the current work.
2.2. Tests for chloroquine sensitivity
Parent and cross progeny clones were tested for chloro-quine sensitivity in CBA mice, using daily oral doses of3 mg kg−1 chloroquine for 8 days, as described by Carltonet al. [22]. Resistance levels were classified as LR (low re-sistance), R (resistance similar to AS(3CQ)) and HR (resis-tance similar to AS(15CQ) and AS(15MF/3)).
2.3. DNA extraction
Parasitised red cells were harvested from mice into cit-rate saline and passed through a column of fibrous cellulosepowder (CF11, Whatman) to remove mouse leucocytes[23].The parasite pellet was washed twice in PBS, and parasitesreleased from their host cells by saponin lysis[24]. The par-asite pellet was resuspended in lysis solution (10 mM TrispH 8.0, 50 mM EDTA, 0.1% SDS, Proteinase K 1 mg ml−1)and incubated overnight at 42◦C. After phenol extraction,DNA was first precipitated by propan-2-ol and 4 M ammo-nium acetate, and then treated using standard procedures[25].
2.4. PCR amplification of pccg10 and pcmdr1, and genesequencing
Orthologues of theP. falciparum crtgene in other speciesare denotedcg10 [26]. We adopt this nomenclature here,and therefore call theP. chabaudigenepccg10.
Two sets of degenerate primers, pcrt-01 and pcrt-05(5′-CKATTTCYCTKGTCATGTTKGARAARC-3′ and 5′-GCRATKGCTATKGCTGGWCCYTGTATRCAAC-3′),and pcrt-04 and pcrt-06 (5′-GAYGARCGTTAYMGRGAR-TTRGATA-3′ and 5′-TGYGAYGATTGYGAHGGWGCWTGG-3′), were designed to amplify two regions of theP.chabaudi pccg10gene using sequences highly conservedbetweenP. falciparum, P. vivaxandP. knowlesi(data kindlyprovided by T. Nomura and T. Wellems). The products were

P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35 29
cloned and sequenced, and the sequence obtained used todesign primers to amplify the intervening sequence. Se-quences 5′ and 3′ to this sequence were generated by nestedPCR using inverse libraries ofP. chabaudigenerated usingcombinations of the restriction enzymes,VspI, NdeI, BspIand Csp43I. Briefly, genomic DNA was digested with therestriction enzymes, diluted and ligated. This material wasused as template in PCR reactions using primers designed todirect DNA synthesis away from known sequence. Productswere cloned, sequenced and the ligation point identified.The data obtained were confirmed by conventional PCR. Se-quences ofpccg10were then assembled using sequence datain both directions from repeated overlapping PCR products.
A similar strategy, but also including the identificationof clones from a size-selected sonicated DNA library inpUC18, and the use of VectoretteTM libraries, was used toamplify and sequencepcmdr1, starting with a PCR productcontaining the 3′ ATP binding cassette[18,22].
PCR products were either sequenced directly using ap-propriate primers or cloned using TOPO TA Cloning®
(InvitrogenTM) prior to sequencing of plasmid DNA. Af-ter cloning, a number of isolated transformants were usedto generate plasmid DNA using QIAprep Spin MiniprepKits (Qiagen®). Sequencing used ABI Prism® BigDyeTM
Terminator (Applied Biosystems) cycle sequencing readyreaction kits. All sequences were repeated using at leastthree independent PCR reactions and clonings, and anyambiguities successfully resolved. Gene sequences wereanalysed and aligned using GCG (University of WisconsinGenetics Computer Group).
BLAST searches to theP. chabaudigenome sequencedata were performed athttp://www.sanger.ac.uk/Projects/P chabaudi/blastserver.shtml. Searches ofP. yoelii, P.bergheiand P. vivax cg10genomic and cDNA sequences,for intron position predictions, were made atwww.ncbi.nlm.nih.gov/projects/Malaria/plasmodiumbl.html.
2.5. Inheritance of pccg10 and pcmdr1 alleles in crossprogeny clones
To test whether the chloroquine responses of crossprogeny clones were associated withpccg10, two PCR sizepolymorphisms inpccg10which distinguished AS and AJalleles (seeSection 3) were used to characterise each clone.Primers pccrt-19 (5′-CTCTGGTCATATTTGAAAAGC-3′)and pccrt-20 (5′-GCAAACTAATTGGAAATGAAATG-3 ′)were used to amplify a gene fragment which spanned bothpolymorphisms. Products were digested with bothEcoRVandHindIII and analysed by gel electrophoresis using 3%Metaphor agarose (Flowgen).
For pcmdr1, a single nucleotide substitution (seeSection 3) was used to distinguish the AS and AJ al-leles. DNA was amplified using primers pcmdr1–48(5′-TAAGAGATGAAGGTGGTATAAG-3′) and pcmdr1–43(5′-GCTTTTTCCATAAGGTCCTAC-3′). Products were di-gested withPagI and analysed by gel electrophoresis on
1.5% agarose. For clones from the AS(15MF/3)× AJ cross,some of which carry two copies of thepcmdr1gene[18],amplified DNA was sequenced using primer pcmdr1–43.Clones bearing apcmdr1gene from both parents could beidentified by two nucleotides appearing at the polymorphicsite on the fluorescent electropherogram.
2.6. Linkage analysis
Linkage between bothpcmdr1 and pccg10 to chloro-quine susceptibility was calculated as a linkage ratio[18].The linkage ratio is the number of progeny clones showinglinkage divided by the total number of progeny. This ra-tio was calculated in two ways, low-resistance clones beinggrouped either with resistant clones or with sensitive clones.Also, in the AS(15MF) × AJ cross, two copies ofpcmdr1were present in seven of the progeny clones; in four clonesboth copies were inherited from the AS(15MF/3) parent,while in the other three clones one copy was inherited fromAS(15MF/3) and one from AJ[18]. For this cross, link-age ratios were calculated twice, once where all the latterthree clones were included in the AS(15MF/3) category forpcmdr1allele, and once where they were included in the AJcategory. The statistical significance of linkage was evalu-ated using a chi-squared test.
3. Results
3.1. Plasmodium chabaudi cg10 gene (pccg10)
Sequences of DNA amplified using primer pairs pcrt-01and pcrt-05, and pcrt-04 and pcrt-06, fromP. chabaudiAS(sens) DNA as template were similar to the correspond-ing sequences ofP. falciparum pfcrt. DNA between thesefragments was then amplified using primers pccg10–11(5′-GTTGAAAAAGGAATATGTTATAAAGG-3 ′) and pccg10–12 (5′-CAAATTTATATGCGCCCCTTTTCC-3′). Fur-ther sequence both 5′ and 3′ was obtained by a number ofrounds of inverse PCR. 4009 bp of nucleotide sequence wassubmitted to DDBJ/EMBL/GenBank databases (accessionnumber AY304549). BLAST searches of theP. chabaudishotgun sequencing project identified identical sequences,giving independent confirmation of a total of 1512 bp of thesequence data reported here.
Thepccg10sequence was aligned with orthologousP. fal-ciparum(pfcrt), P. berghei(pbcg10), P. yoelii (pycg10), andP. vivax (pvcg10) genomic and cDNA sequences in orderto predict a possible exon/intron structure forpccg10. Thegene structure ofpfcrt (13 exons and 12 introns) could besuperimposed easily on thepccg10sequence data, yieldinga gene structure with predicted 5′ and 3′ intron splice sitesfor pccg10which were highly similar topfcrt (Fig. 1). ForP. chabaudi, the consensus sequences were GTAwr(5′) and–wwyAG (3′) (R, purines A or G; W, A or T; Y, pyrimidinesC or T), compared to GTAAr and WWYAG, respectively,

30 P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35
Fig. 1. Gene structureP. chabaudi cg10andP. falciparum crt. The 13 exons ofP. falciparum crt(numbered 1–13) and the predicted exons ofP. chabaudicg10 are indicated (black shading). Introns and 5′- and 3′-UT regions are also shown (grey shading). Similarity betweenP. falciparumand P. chabaudiexons is indicated by hatched areas.
for P. falciparum(upper case denotes invariant nucleotides,lower case denotes one or two exceptions in 12 introns).However, there was no intron to correspond withpfcrt in-tron 2. Instead, another intron 3′ to that in thepfcrt genewas found inpccg10. This structure was also found in theorthologous genes of the rodent malaria speciesP. berghei,P. yoelii, as well as in that ofP. vivax(data not shown).
Table 2shows the lengths of the predictedpccg10andpfcrt exons and introns, which, are small and remarkablysimilar in size in both species. Forpccg10, with the exceptionof the first (480 bp) and fifth (151 bp) intron, the sizes of theintrons are within the range 92–121 bp.
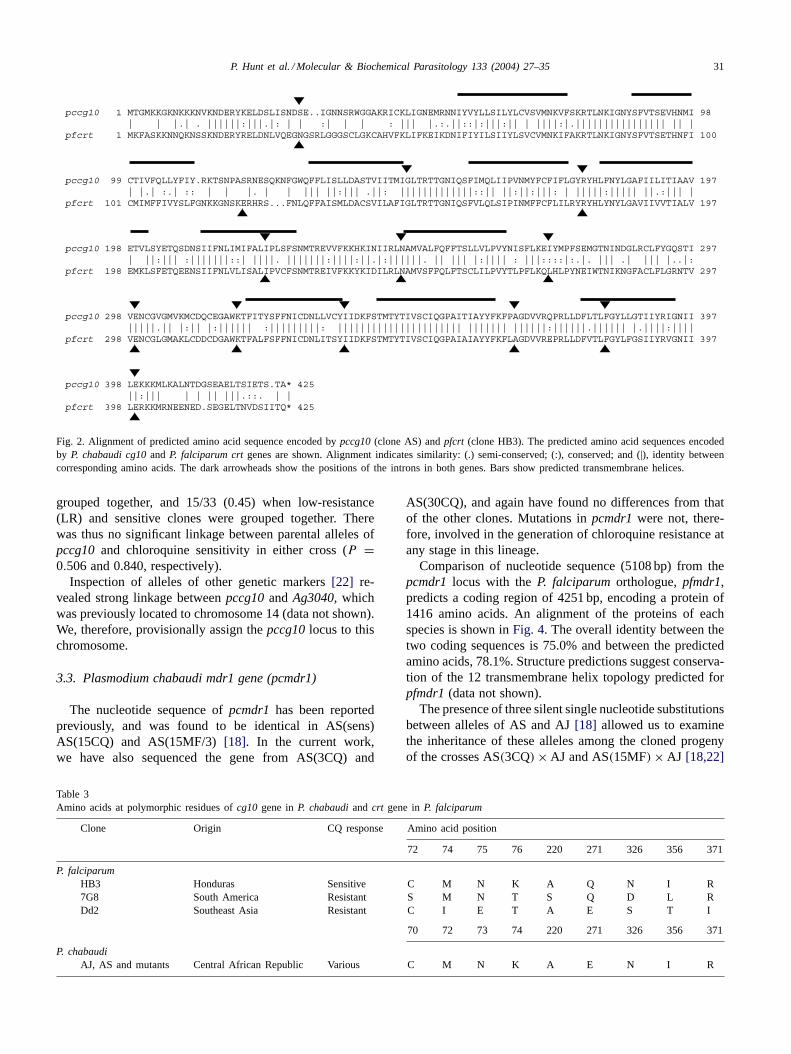
The presumed coding sequence forpccg10(1275 bp) wasused to generate a predicted amino acid sequence of 424amino acids. An alignment of this sequence with that en-coded bypfcrt is shown inFig. 2. The predicted nucleotidecoding sequences share 68.6% identity, and the predictedamino acid sequences 75.1% identity. Structure predictionsfor pccg10defined 10 transmembrane helices, as is the case
Table 2Exon and intron lengths inP. chabaudi cg10and P. falciparum crt
Exon length (bp) Intron length (bp)
P. chabaudi P. falciparum P. chabaudi P. falciparum
5′-UT – – 767 –1 91 91 479 1692 348 269 115 1633 94 173 113 1694 133 133 121 1615 72 72 151 1236 76 76 109 1057 83 83 118 1268 51 51 108 1379 57 57 94 142
10 93 93 92 19411 45 45 93 13712 55 55 110 16913 77 77 – –3′-UT – – 264 –
Total codingregion
1275 1275
Total introns 1703 1795
of pfcrt [2] (data not shown). The positions of these helicesare indicated in relation to the intron positions inFig. 2.Most of the predicted helices are contained within single ex-ons, even though these exons are often only slightly longerthan the predicted helix.
The predicted amino acid residues at positions homolo-gous to the polymorphic sites inP. falciparumare shown inTable 3. All residues are identical to those found in one orother of a number of clones and isolates ofP. falciparum[2].Codon 74 inP. chabaudi(homologous to aa 76 inP. falci-parum) encodes lysine, which is the same as that associatedwith chloroquine sensitivity inP. falciparum.
Overlapping fragments ofpccg10from all theP. chabaudiclones were amplified and sequenced. No differenceswere found between any of the clones of the AS lin-eage, i.e. AS(sens), AS(3CQ), AS(15CQ), AS(30CQ) orAS(15MF/3). However, there were differences between thealleles of the gene in the AS clones and in AJ. 23 poly-morphic sites were identified, of which 20 were single basesubstitutions (Table 4). Two of these occurred in coding se-quences, and both were silent (synonymous) substitutions.There were three insertion/deletion polymorphisms; a sin-gle base deletion in AS in the 5′-untranslated region, a fourbase deletion in intron 2 and a 14 bp insertion in intron 4with respect to AJ.
3.2. Analysis of cross progeny clones for pccg10 alleles
The 4 bp difference in intron 2 and 14 bp difference inintron 4 between AS and AJ allowed us to test the inheritanceof parentalpccg10alleles in the progeny clones derived fromthe two crosses AS(3CQ)×AJ [22] and AS(15MF/3)× AJ[18].
DNA from progeny clones was amplified using primerspccrt-19 and pccrt-20, digested withHindIII and EcoRVand resolved on 3% metaphor gels. As well as a visiblenon-polymorphic band, AS alleles gave polymorphic bandsof 291 and 258 bp while AJ alleles gave corresponding bandsof 295 and 244 bp.
Progeny clone phenotypes and theirpccg10alleles areshown inFig. 3. The overall linkage ratios were 19/33 (0.58)when resistant (R) and low-resistance (LR) clones were
P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35 31
Fig. 2. Alignment of predicted amino acid sequence encoded bypccg10(clone AS) andpfcrt (clone HB3). The predicted amino acid sequences encodedby P. chabaudi cg10and P. falciparum crtgenes are shown. Alignment indicates similarity: (.) semi-conserved; (:), conserved; and (|), identity betweencorresponding amino acids. The dark arrowheads show the positions of the introns in both genes. Bars show predicted transmembrane helices.
grouped together, and 15/33 (0.45) when low-resistance(LR) and sensitive clones were grouped together. Therewas thus no significant linkage between parental alleles ofpccg10and chloroquine sensitivity in either cross (P =0.506 and 0.840, respectively).
Inspection of alleles of other genetic markers[22] re-vealed strong linkage betweenpccg10andAg3040, whichwas previously located to chromosome 14 (data not shown).We, therefore, provisionally assign thepccg10locus to thischromosome.
3.3. Plasmodium chabaudi mdr1 gene (pcmdr1)
The nucleotide sequence ofpcmdr1 has been reportedpreviously, and was found to be identical in AS(sens)AS(15CQ) and AS(15MF/3)[18]. In the current work,we have also sequenced the gene from AS(3CQ) and
Table 3Amino acids at polymorphic residues ofcg10 gene inP. chabaudiand crt gene inP. falciparum
Clone Origin CQ response Amino acid position
72 74 75 76 220 271 326 356 371
P. falciparumHB3 Honduras Sensitive C M N K A Q N I R7G8 South America Resistant S M N T S Q D L RDd2 Southeast Asia Resistant C I E T A E S T I
70 72 73 74 220 271 326 356 371
P. chabaudiAJ, AS and mutants Central African Republic Various C M N K A E N I R
AS(30CQ), and again have found no differences from thatof the other clones. Mutations inpcmdr1were not, there-fore, involved in the generation of chloroquine resistance atany stage in this lineage.
Comparison of nucleotide sequence (5108 bp) from thepcmdr1 locus with theP. falciparumorthologue,pfmdr1,predicts a coding region of 4251 bp, encoding a protein of1416 amino acids. An alignment of the proteins of eachspecies is shown inFig. 4. The overall identity between thetwo coding sequences is 75.0% and between the predictedamino acids, 78.1%. Structure predictions suggest conserva-tion of the 12 transmembrane helix topology predicted forpfmdr1(data not shown).
The presence of three silent single nucleotide substitutionsbetween alleles of AS and AJ[18] allowed us to examinethe inheritance of these alleles among the cloned progenyof the crosses AS(3CQ) × AJ and AS(15MF) × AJ [18,22]

32 P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35
Table 4Polymorphisms betweenP. chabaudiclones AS and AJ inpccg10gene
Fig. 3. Characteristics of progeny clones from crosses between AS(3CQ) × AJ and AS(15MF/3)× AJ. Chloroquine responses: lr, low resistance; r,resistant. Characteristics of the clones have been re-tested since the earlier papers[18,22], and the following differences found: (i) clone 253/5 ischloroquine sensitive, not resistant as previously reported; (ii) clones 39/2 and 388/12 contain thepcmdr1 allele of parent AS, not AJ as previouslyreported, while clone 441/11 contains the parent AJpcmdr1 allele, not AS as previously reported. These discrepancies could be due to the fact thatsequencing was used to typepcmdr1alleles in the current study, while RFLP-typing was used in the earlier study; it is possible that in these three clonesrecombination could have occurred between the gene and the restriction site used in the earlier typing.
(Fig. 3). Progeny clones which showed low resistance tochloroquine were grouped with either sensitive clones or re-sistant clones. The linkage ratios (LR+ R v S) are 19/31(0.61) and 18/31 (0.58) where clones containing two parentalpcmdr1alleles were assigned to AS(15MF/3) or AJ allelegroups, respectively. When low-resistance clones are as-signed as sensitive (LR+S v R), the linkage ratios are 16/31(0.52) and 17/31 (0.55), respectively. None of these fourtests show significant linkage (chi-squared test,P = 0.25,0.35, 0.64, 0.61, respectively), and there was, therefore, noevidence of linkage betweenpcmdr1alleles and chloroquineresponse.
4. Discussion
In this work, we have characterised theP. chabaudiortho-logues of theP. falciparumgenespfcrt andpfmdr1, both ofwhich have been implicated as determinants of chloroquineresistance inP. falciparum. We find that no mutations hadoccurred inpccg10or pcmdr1 in the chloroquine-resistantclones AS(3CQ), AS(15CQ), AS(30CQ) or AS(15MF/3),
all of which had been derived from the chloroquine-sensitiveAS(sens) clone. In addition, AS alleles ofpccg10or pcmdr1did not co-segregate with resistance in progeny of crossesbetween resistant AS and sensitive AJ clones. We conclude,therefore, that neither of these genes, nor genes closelylinked to them, determine this type of resistance inP.chabaudi, at least in the AS clones studied here.
The gene structure ofpccg10is very similar topbcg10,pycg10andpvcg10, and differs frompfcrt only at the posi-tion of the second intron. These results are consistent witha model of intron evolution where introns may be progres-sively lost from an ancestral gene. According to this model,the ancestral gene would have contained at least 13 introns(equivalent to the 11 introns common to bothP. falciparumandP. chabaudi, plus 2 introns corresponding to intron 2 ofP. falciparumand intron 2 ofP. chabaudi, respectively). Sub-sequently, ancestral intron 3 (P. chabaudiintron 2) wouldhave been lost in theP. falciparumlineage, while ancestralintron 2 (P. falciparumintron 2) would have been lost in thealternative branch prior to the divergence ofP. chabaudi, P.berghei, P. yoelii and P. vivax. This is consistent with thephylogeny of these species inferred fromSSUrRNA, the cir-

P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35 33
Fig. 4. Alignment of predicted amino acid sequence encoded bypcmdr1 (AS) and pfmdr1 (D10). The predicted amino acid sequences encoded byP. chabaudi mdr1and P. falciparum mdr1are shown. Alignment indicates similarity: (.) semi-conserved; (:), conserved; and (|), identity betweencorresponding amino acids.
cumsporozoite protein genecspand mitochondrialcytb se-quences[27].
It is possible to speculate whether genes such as the an-cestralcg10/crtevolved by the sequential addition of smalldomain-like segments, each encoding one transmembranehelix. A transmembrane topology prediction suggests 10transmembrane helices in the protein. The positions and ex-tent of these fit within individual exons of the genes fromboth P. falciparumand P. chabaudiremarkably well (Fig.1). If this is the case then the ancestral gene may have in-cluded further introns between transmembrane helices 1, 2and 3 (i.e. aa 76–88 and 110–130, usingpccg10numbering).
In this regard, it would be of obvious interest to examineorthologues of other apicomplexans which diverged fromPlasmodiumprior to the branching of theP. falciparumandP. vivax/rodent malaria lineages.
Our finding thatpccg10is not a determinant of chloro-quine resistance in theseP. chabaudimutants is of con-siderable interest in view of the clear involvement of itsorthologuepfcrt in P. falciparumresistance. The fact thatpcmdr1was not involved in resistance either was also of in-terest, although less surprising since its role inP. falciparumresistance is less clear-cut than that ofpfcrt. There is goodfield evidence that certain alleles ofpfmdr1, notably one

34 P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35
containing 86Y and a second containing 1034C, 1042D and1246Y, modify the level of chloroquine resistance inP. fal-ciparumonce it has been established by apfcrt mutant[4,6].
In a previous study on AS(3CQ), it was found that anunidentified gene on chromosome 11 ofP. chabaudiwaslinked to chloroquine resistance among cross progeny[22].It also seems clear that chloroquine resistance in bothP. fal-ciparum and in P. chabaudiis, at least at higher levels, amultigenic trait. ForP. chabaudi, the original selection ofparasites resistant to low levels of drug required repeatedpassage of parasites over several weeks using gradual in-creases in dosage[20,21]. This contrasts with the single-stepselection of parasites resistant to pyrimethamine which com-monly occurs following just a single course of treatmentwith a high dose of drug[19]. These different dynamics ofselection mirror the situation inP. falciparumwhere the ap-pearance of parasites resistant to antifolate drugs such aspyrimethamine occurred quickly and frequently, whilst thegeneration of chloroquine resistance took many years of ex-posure to the drug[28] and has probably occurred only alimited number of times[14].
When the cross between the AJ× AS(3CQ) clones ofP.chabaudiwas analysed for drug response, progeny cloneswith at least two resistant phenotypes, LR and R, were ob-served. It seems quite likely, therefore, that at least two genesinfluence the level of chloroquine resistance in AS(3CQ).In the cross, these genes would have been inherited inde-pendently among the progeny, giving new haplotypes withdifferent levels of drug response. There was good linkage ofthe resistance phenotypes with the locus on chromosome 11,and quantitive trait loci (QTL) analysis indicated that cer-tain loci on other chromosomes could be involved[22]. Wehave also demonstrated here that progeny exhibiting inter-mediate chloroquine-resistance phenotypes were generatedfrom the cross between AJ and AS(15MF/3), again suggest-ing that two or more unlinked loci determining resistancehad undergone recombination. We were unable to identifyany progeny with a level of resistance similar to that of theAS(15MF/3) parent. However, the number of progeny ex-amined from this cross was small (Fig. 3), and so this resultis explicable if several loci were involved in the generationof high level chloroquine resistance of AS(15MF/3). Therewas also no apparent relationship between chloroquine andmefloquine responses among the progeny.
The possibility that over-expression ofpccg10 andpcmdr1 may contribute to chloroquine resistance has notbeen addressed here. In an earlier study[18], no amplifica-tion or over-expression ofpcmdr1was noted in the chloro-quine resistant clone AS(15CQ) which was derived fromAS(3CQ) and was the progenitor of AS(30CQ); it is, there-fore, unlikely thatpcmdr1 is over-expressed in AS(3CQ).With regard topccg10, the linkage between markers onchromosome 11 and chloroquine resistance[22] suggeststhat if a change in expression ofpccg10were to be a causeof the resistance, it would be dependent upon a mutation ina regulatory gene on this chromosome.
An explanation for the contrasting role of thecrt genein P. falciparumand P. chabaudiis that the basic mecha-nism for chloroquine resistance may be different in thesespecies. In this regard, ourP. chabaudifinding is compara-ble to that ofP. vivax, where there is no association betweenchloroquine resistance and coding mutations in theP. vivaxorthologue ofpfcrt [26]. This situation contrasts with thatfor pyrimethamine-resistance where mutations in dihydro-folate reductase (dhfr), the target enzyme for this antifolatedrug, are at similar sites in the gene in differentPlasmod-ium species[29–31]. Amplification of bothdhfr andmdr1has also been found to confer resistance, respectively, topyrimethamine and mefloquine inP. falciparumand rodentmalaria models[18,32], although this mechanism appears tobe less important than mutation in generating resistance infield conditions[17]. This could, therefore, be a reflectionof differing ways in which the selection of resistant para-sites occurs in the field as opposed to within an experimentallaboratory model.
In the context of a multigenic origin of chloroquine re-sistance, it may be that thepfcrt polymorphism (K76T) isa necessary final mutation which depends upon other pre-vious mutations before selection in the field is observed forP. falciparum. In this scenario, the rodent model may turnout to be a sensitive one for identifying mutations in othergenes which lead to lower levels of resistance and whichare a prerequisite for higher level resistance developing inP. falciparum. In any case, the identification of the geneswhich are involved in chloroquine resistance inP. chabaudiis likely to help us to understand the resistance phenotypeand its genetic basis in bothP. vivaxandP. falciparum.
Acknowledgements
We thank Richard Fawcett and Ronnie Mooney for ex-cellent technical assistance. The work was supported by theUK Medical Research Council. P.V.L.C. was supported bythe Fundaçao para a Ciencia e Tecnologia and the Centro deMalaria e Outras Doenças Tropicais, Lisbon, Portugal.
References
[1] Wellems TE, Plowe CV. Chloroquine-resistant malaria. J Infect Dis2001;184:770–6.
[2] Fidock DA, Nomura T, Talley AT, et al. Mutations in the P. falciparumdigestive vacuole transmembrane protein Pfcrt and evidence for theirrole in chloroquine-resistance. Mol Cell 2000;6:861–71.
[3] Sidhu AB, Verdier-Pinard D, Fidock D. Chloroquine resistance inPlasmodium falciparummalaria conferred bypfcrt mutations. Sci-ence 2002;298:210–3.
[4] Babiker HA, Pringle SJ, Abdel-Mushin A, Mackinnon M, HuntP, Walliker D. High-level chloroquine-resistance in Sudanese iso-lates of Plasmodium falciparumis associated with mutations inthe chloroquine-resistance transporter genepfcrt and the multidrug-resistance genepfmdr1. J Infect Dis 2001;183:1535–8.

P. Hunt et al. / Molecular & Biochemical Parasitology 133 (2004) 27–35 35
[5] Chen N, Russell B, Staley J, Kotecka B, Nasveld P, Cheng Q.Sequence polymorphisms inpfcrt are strongly associated withchloroquine resistance inPlasmodium falciparum. J Infect Dis2001;183:1543–5.
[6] Djimde A, Doumbo OK, Steketee RW, Plowe CV. Application of amolecular marker for surveillance of chloroquine-resistant falciparummalaria. Lancet 2001;358:890–1.
[7] Dorsey G, Kamya MR, Singh A, Rosenthal PJ. Polymorphisms in thePlasmodium falciparum pfcrtandpfmdr-1genes and clinical responseto chloroquine in Kampala, Uganda. J Infect Dis 2001;183:1417–20.
[8] Mayor AG, Gomez-Olive X, Aponte JJ, et al. Prevalence of theK76T mutation in the putativePlasmodium falciparumchloroquineresistance transporter (pfcrt) gene and its relation to chloroquineresistance in Mozambique. J Infect Dis 2001;183:1413–6.
[9] Foote SJ, Kyle DE, Martin RK, et al. Several alleles of the multidrug-resistance gene are closely linked to chloroquine-resistance inPlas-modium falciparum. Nature 1990;345:255–8.
[10] Wellems TE, Panton LJ, Gluzman IY, et al. Chloroquine resistanceis not linked to mdr-like genes in aPlasmodium falciparumcross.Nature 1990;345:253–5.
[11] Awad-El-Kariem FM, Miles MA, Warhurst DC. Chloroquine resis-tant Plasmodium falciparumisolates from the Sudan lack two muta-tions in the pfmdr1 gene thought to be associated with chloroquineresistance. Trans R Soc Trop Med Hyg 1992;86:587–9.
[12] Haruki K, Bray PG, Ward SA, Hommel M, Ritchie GY. Chloroquineresistance ofPlasmodium falciparum: further evidence for a lack ofassociation with mutations of thepfmdr1 gene. Trans R Soc TropMed Hyg 1994;88:694.
[13] Reed MB, Saliba KJ, Caruana SR, Kirk K, Cowman AF. Pgh1 mod-ulates sensitivity and resistance to multiple antimalarials inPlas-modium falciparum. Nature 2000;403:906–9.
[14] Wootton JC, Feng X, Ferdig MT, et al. Genetic diversity andchloroquine selective sweeps inPlasmodium falciparum. Nature2002;418:320–3.
[15] Peterson DS, Walliker D, Wellems TE. Evidence that a point mutationin dihydrofolate reductase-thymidylate synthase confers resistanceto pyrimethamine in falciparum malaria. Proc Natl Acad Sci USA1988;85:9114–8.
[16] Sibley CH, Hyde JH, Sims PFG, et al. Pyrimethamine-sulfadoxineresistance inPlasmodium falciparum: what next? Trends Parasitol2001;17:582–8.
[17] Carlton JM, Hayton K, Cravo PV, Walliker D. Of mice and malariamutants: unravelling the genetics of drug-resistance using rodentmalaria models. Trends Parasitol 2001;17:236–42.
[18] Cravo PVL, Carlton JM-R, Hunt P, Bisoni L, Padua RA, WallikerD. Genetics of mefloquine resistance in the rodent malaria parasitePlasmodium chabaudi. Antimicrob Agents Chemother 2003;47:709–18.
[19] Walliker D, Carter R, Sanderson A. Genetic studies onPlasmod-ium chabaudi: recombination between enzyme markers. Parasitology1975;66:309–20.
[20] Rosario VE. Genetics of chloroquine-resistance in malaria parasites.Nature 1976;261:585–6.
[21] Padua RA.Plasmodium chabaudi: genetics of resistance to chloro-quine. Exp Parasitol 1981;52:419–26.
[22] Carlton J, Mackinnon M, Walliker D. A chloroquine-resistance locusin the rodent malaria parasitePlasmodium chabaudi. Mol BiochemParasitol 1998;93:57–72.
[23] Homewood CA, Neame KD. A comparison of methods used for theremoval of white cells from malaria-infected blood. Ann Trop MedParasitol 1976;70:249–51.
[24] Carter R. Studies on enzyme variation in the murine malaria parasitesPlasmodium berghei, P. yoelii, P. vinckeiand P. chabaudiby starchgel electrophoresis. Parasitology 1978;76:241–67.
[25] Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratorymanual, 2nd ed. New York: Cold Spring Harbor Laboratory; 1989.
[26] Nomura T, Carlton JM-R, Baird JK, et al. Evidence for differentmechanisms of chloroquine resistance in 2 Plasmodium species thatcause human malaria. J Infect Dis 2001;183:1653–61.
[27] Ayala FJ, Escalante AA, Lal AA, Rich SM. Evolutionary relation-ships of human malaria parasites. In: Sherman IW, editor. Malaria:parasite biology, pathogenesis and protection. Washington: ASMPress; 1998. p. 285–300.
[28] Peters W. Chemotherapy and drug resistance in malaria, 2nd ed.New York: Academic Press; 1987.
[29] Cowman AF, Lew M. Antifolate drug selection results in duplicationand rearrangement of chromosome 7 inPlasmodium chabaudi. MolCell Biol 1989;9:5182–8.
[30] Cowman AF, Lew AM. Chromosomal rearrangements and pointmutations in the DHFR-TS gene ofPlasmodium chabaudiunderantifolate selection. Mol Biochem Parasitol 1990;42:21–9.
[31] Cheng Q, Saul A. The dihydrofolate reductase domain of ro-dent malarias: point mutations and pyrimethamine resistance. MolBiochem Parasitol 1994;65:361–3.
[32] Thaithong S, Ranford-Cartwright LC, Siripoon N, et al.Plasmod-ium falciparum: gene mutations and amplification of dihydrofolategenes in parasites grown in vitro in presence of pyrimethamine. ExpParasitol 2001;98:59–70.