Embed Size (px)
Citation preview

Plant, Cell and Environment (1990) 13, 79-83
Chlorophyll fluorescence measured using theFraunhofer line-depth principle and relationship tophotosynthetic rate in the field
G. A. CARTFR,' A. F. THEISEN^ & R. J. MITCHELL' 'Science and Technology Laboratory, NationalAeronautics and Space Adtninistration, Stennis Space Center, Mississippi 39529, U.S.A., -Branch ofGeophysics, U.S. Geological Survey, Denver Federal Center, Denver, Colorado 80225, U.S.A., and 'Schoolof Forestry, Auburn University, Auburn, Alabama 36849, U.S.A.
Received 21 February 1989; received in revised fortii 26 Jutic 1989; accepted for publication 31 August 1989
Abstract. A field study was conducted to determinethe relationship of solar-excited chlorophyll a fluor-escence to net COj assimilation rate in attachedleaves. The Fraunhofer line-depth principle was usedto tneasure fluorescence at 656.3 nm wavelengthwhile leaves remained exposed to full sunlight andnormal atmospheric pressures of CO, and O,. Fluor-escence induction kinetics were observed when leaveswere exposed to sunlight after 10 min in darkness.Subsequently, fluorescence varied inversely with assi-milation rate. In the C4 Zea mays, fluorescencedecreased from 2.5 to 0.8 mW m- ' nm ' as CO,assirnilation rate increased from 1 to 8/miol m ' s '(r̂ = 0.52). Ill the C, Liquidamhar styraeifiua andPinus taeda, fluorescence decreased frorn 6 to 2 mWm"^ nm"' as assimilation rate increased frotn 2 to 5or 0 to 2^nTiol tn - s ' (r' = 0.44 and 0.45, respect-ively). The Fraunhofer line-depth principle enablesthe simultaneous measurement of solar-excited fluor-escence and CO2 assitnilation rate in individualleaves, but also at larger scales. Thus, it may contri-bute significantly to field studies of the relationship offluorescence to photosynthesis.
Key-words: chlorophyll a fluorescenee; photosynthesis; Fraunhoferline; Zea may.s: Liquidambar styraeifiua; Pinus taeda.
Introduction
During the initial events of photosynthesis, absorp-tion of visible radiation by a leaf raises chlorophyll aelectrons to higher-energy quantum states (for reviewsee Govindjee, 1975; Govindjee, Aniesz & Fork,1986). Under optimal conditions, tnost of this exei-tation energy is transferred chemically, and ulti-mately enables CO2 fixation (Lichtenthaler &Rinderle, 1988). However, when conditions forphotosynthesis are less favourable, excitation energymay be lost as photons, i.e. fluorescence (F), whenthe excited electrons return to a lower-energyquantum state. For intact leaves under constant irra-diance, F is quenched, or decreases, as photosyn-
Correspotidence: Dr Gt'egory A. Carter, Scictice andTechnology Laboratory, National Aeronautics and SpaceAdministratioti, Stennis Space Cctitcr, Missis,sippi 39529, U.S.A.
thesis increases (Krause & Weis, 1984), and tnayrepresent 2-5% of the energy absorbed (Lichten-thaler & Rinderle, 1988). Although there are severalmechanistns by which F may be quenched (Krause &Weis, 1984; Sivak & Walker, 1985; Lichtenthaler &Rinderle, 1988), energy balance and experimentalresults indicate that F is significantly afiected by CO.assimilation rate (Sivak & Walker, 1985; Lichten-thaler & Rinderle, 1988).
Under field conditions, solar-excited F generallycannot be seen with the human eye or tneasureddirectly because of the intensity of reflected sunlight.However, the Fraunhofer line-depth principle(Kozyrev, 1956; Grainger & Ring, 1962; Janza, 1975)enables the measurement of F while a leaf, or otherfluorescing surface, remains exposed to sunlight. As aresult of radiation absorption by hydrogen in thesolar atmosphere, a dark, narrow absorption band,or Fraunhofer line, occurs at 656.3 ntn wavelength(the Ha line) in the solar spectrutn (Weast, Astle &Beyer, 1984). Although peak chlorophyll a fluores-cence occurs near 690 and 735 nm (Lichtenthaler &Rinderle, 1988), the Ha line can be etnployed totneasure the relatively low etnission of chlorophyll afluorescence at 656.3 ntn (see Methods for furtherdescription). This principle has been used to detectinereased F with geochetnical stress in Pinusponderosa (Watson, Hemphill & Hessin, 1973;Watson & Hetnphill, 1976; Hetnphill et al., 1977) andwith increased water stress and stotnatal resistance towater vapour diffusion in Citrus spp. (McFarlane etal., 1980). However, the Fraunhofer line-depth prin-ciple has not been used previously to study therelationship of solar-excited F to net COj assimila-tion rate (A). Given the elose relationship of stomatalresistance to A (for review see Nobel, 1983; Sharkey,1985a), the results of McFarlane et al, (1980)suggested that the Fraunhofer line-depth principlemight be used to determine the relationship of F to Ain the field. Thus, the objective of our study was tomeasure A and F for leaves exposed to full sunlightand normal atmospheric O, and CO, pressures, anddetermine the relationship of F to A for specieschosen to represent the C, and C4 photosyntheticpathways.
79

80 G. A. CARTER et al.
Materials and methods
To study the relationship of solar-excited F to A, Fand A were measured under clear-sky conditionsfrom 27 April to 6 May, 1988 at Auburn University,Alabama, U.S.A. (lat. 32° 40'N, long. 85° 30'W).Potted plants were used to position attached leaveswithin the field of view of the stationary fluorescencedetector. Selected for study were two C, species,Liquidambar styracifiua L. (sweetgum) and Pinustaeda L. (loblolly pine), and the C4 Zea mays L.(corn). The plants were grown in a glasshouse whichtransmitted 70% of incident sunlight during latewinter and spring, 1988, and were transferred to thefield 1-3 d prior to measurement.
Fluorescence from the upper (adaxial) surfaces ofmature leaves was measured for the two broadleavedspecies. Attached leaves were held horizontal to theground by clamps covered with non-fluorescent blackfelt. For pine, 75-100 needles attached to the sameshoot were held parallel to each other in the sameplane to simulate a broadleaf orientation. Fluores-cence was measured using an advanced prototype ofthe Fraunhofcr Line Discriminator (FLD Mark II)(Perkin-Elmer Inc., Norwalk, CT, U.S.A.)*, whichincorporated the Fraunhofer line-depth principle inits design (Stoerz, Hemphill & Markle, 1969; Plascyk,1975; Plascyk & Gabriel, 1975). The FLD wassupported 1.7 m above the ground by a steel frame.Each leaf was positioned 0.4 m beneath the apertureof the FLD and remained fully exposed to sunlight.At this distance, F was measured for a 3.5x3.8 cmarea on the broadleaf or needle plane. Black felt wasplaced beneath the leaf to block possible soil fluores-cence. Fluorescence and A were measured onlyduring 1000-1400 h (solar time) to insure saturatingphotosynthetically active radiation (PAR) and aconstant ratio of irradiance in the 656.3 nm Fraun-hofer line to irradiance in the adjacent spectral conti-nuum. Total irradiance (400-1100 nm) and PARincident on the leaf surface were measured with apyranometer and quantum sensor, respectively(models 200 SB and 190 SB, LICOR InstrumentsInc., Lincoln, NE, U.S.A.).
To summarize the Fraunhofer line-depth principle(Plascyk, 1975; Plascyk & Gabriel, 1975): Twomeasurements each are made of solar irradiance andradiance from the object suspected to fluoresce.These measurements are centred in the darkestportion of the Ha absorption line at 656.28 nm wave-length in the solar spectrum. A diffuse reflectivesurface is used to measure direct plus diffuse solarirradiance in the 656.28 +0.50 nm waveband, whichincludes the Ha Fraunhofer line plus the immediatelyadjacent continuum (a, approximately 880 mW m ^nm '). The Fraunhofer line-centre is measured in the
•Mention of commercial products is for inrormiUivc purposesonly, and does nol conslilule an endorsement by the United StatesNational Aeronautics and Space Administration or GeologicalSurvey.
30
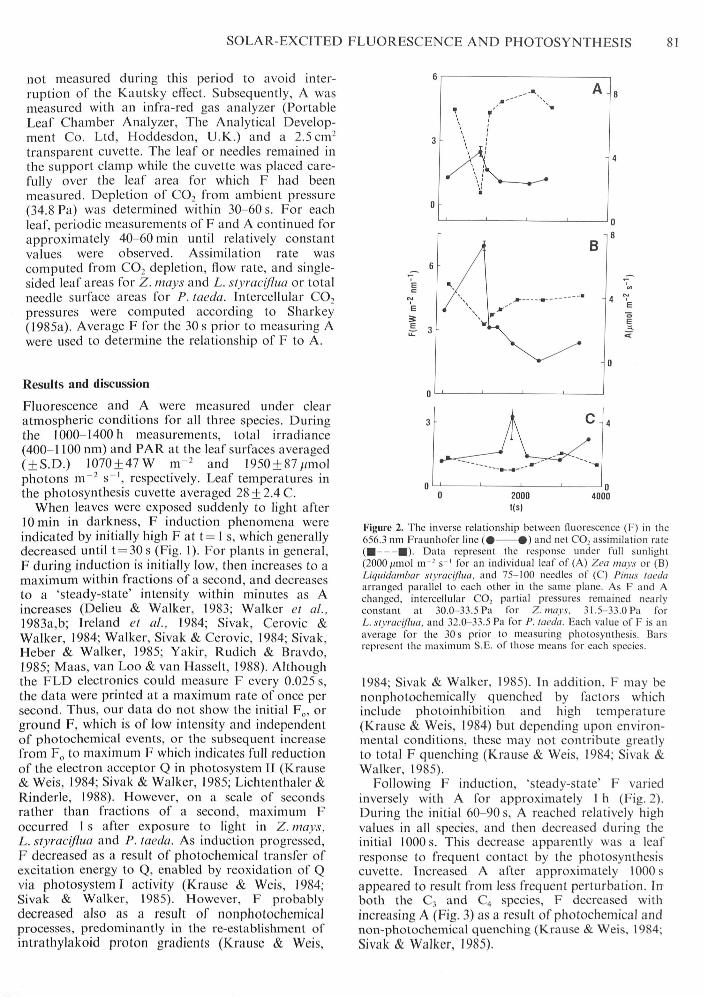
Figure. 1. Fluorescence (F) induction kinetics (the Kautsky elTect)measured under full sunlight using the iMaunhofer line-deptliprinciple. Data represent the response of an attached leaf of Zcanuiy.s ( # • ) or Lii/iiidiim/xir xlyracijlua (A A), and 75-100attached needles of Pinus taeda ( • • ) arranged parallel toeach other in the same plane. Instrumentation could not recordevents which occurred within fractions ol' a second. Thus, anyincrease to maximum F at t = l s from F,, or ground F is notshown.
more narrow band of 656.28 + 0.35 nm (b, approxi-mately 570 mW tn ' nm '). Sirnilarly, radiance of thetarget (e.g. a leaf, the landscape) continuum ismeasured in the 656.28 + 0.50 nm band (d), andradiance in the target line-centre is measured over the656.28 + 0.35 ntn band (c). If the target is non-fluor-escent, and assuming that target reflectivity at656.28+ 0.50 nm is equal to that at 656.28 + 0.35 nm,then b/a will equal c/d. When fluorescent emissionoccurs in the Fraunhofer line waveband, then c/d willbe greater than b/a. Fluorescence was computed(Plascyk, 1975; Plascyk & Gabriel, 1975) as:
= d -a(d-c)(a-b) (1)
To cause F and A to vary under field conditions, aleaf beneath the FLD, wrapped in alurninium foil toexclude light for 10 min, was exposed suddenly tosunlight. This procedure resulted in fluoiescenceinduction kinetics (the Kautsky effect). Fluorescencewas recorded initially and then every 4-5 s followingexposure to sunlight until t = 30 s. Photosynthesis was
SOLAR-EXCITED FLUORESCENCE AND PHOTOSYNTHESIS 81
not measured during this period to avoid inter-ruption of the Kautsky effect. Subsequently, A wasmeasured with an infra-red gas analyzer (PortableLeaf Chamber Analyzer, The Analytical Develop-ment Co. Ltd, Hoddesdon, U.K.) and a 2.5 cm'transparent cuvette. The leaf or needles remained inthe support clamp while the cuvette was placed care-fully over the leaf area for which F had beenmeasured. Depletion of COT from ambient pressure(34.8 Pa) was determined within 30-60 s. For eachleaf, periodic measurements of F and A continued forapproximately 40-60 min until relatively constantvalues were observed. Assimilation rate wascomputed from CO^ depletion, flow rate, and single-sided leaf areas for Z. mays and L. styraeifiiia or totalneedle surface areas for P. taeda. Intercellular COjpressures were computed according to Sharkey(1985a). Average F for the 30 s prior to measuring Awere used to determine the relationship of F to A.
Results and discussion
Fluorescence and A were measured under clearatmospheric conditions for all three species. Duringthe 1000-1400 h measurements, total irradiance(400-1100 nm) and PAR at the leaf surfaces averaged( + S.D.) 1070 + 47W m ' and 1950 + 87/imolphotons m~^ s"', respectively. Leaf temperatures inthe photosynthesis cuvette averaged 28 +2.4 C.
When leaves were exposed suddenly to light after10 min in darkness, F induction phenomena wereindicated by initially high F at t= 1 s, which generallydecreased until t = 30s (Fig. I). For plants in general,F during induction is initially low, then increases to amaximum within fractions of a second, and decreasesto a 'steady-state' intensity within minutes as Aincreases (Delieu & Walker, 1983; Walker et al.,1983a,b; Ireland et al., 1984; Sivak, Cerovic &Walker, 1984; Walker, Sivak & Cerovic, 1984; Sivak,Heber & Walker, 1985; Yakir, Rudich & Bravdo,1985; Maas, van Loo & van Hasselt, 1988). Althoughthe FLD electronics could measure F every 0.025 s,the data were printed at a maximum rate of once persecond. Thus, our data do not show the initial F̂ ,, orground F, which is of low intensity and independentof photochemical events, or the subsequent increasefrom F(, to maximum F which indicates full reductionof the electron acceptor Q in photosystem II (Krause& Weis, 1984; Sivak & Walker, 1985; Lichtcnthaler &Rinderle, 1988). However, on a scale of secondsrather than fractions of a second, maximum Foccurred 1 s after exposure to light in Z. mays,L. styraeifiua and P. taeda. As induction progtcssed,F decreased as a result of photochemical transfer ofexcitation energy to Q, enabled by reoxidation of Qvia photosystem I activity (Krause & Weis, 1984;Sivak & Walker, 1985). However, F probablydecreased also as a result of nonphotoclicmicalprocesses, predominantly in the re-establishment ofintrathylakoid proton gradients (Krause & Weis,
4000
Figure 2. The Inverse relationship between fluorescence (F) in the65(1.3 nm Fraunhofer line ( • • ) and net CO, assimilation rate( • • ) . Data represent the response under full sunlight(2000/(inol m - s ' for an individual leaf of (A) Zca iiuiy.s or (B)Liquiikinthiir slyrucifliui. and 75-100 needles of (C) Pinus lacdaarranged paraljel lo each other in the same plane. As F and Achanged, intercelhilar CO, partial pressures remained nearlyconstant at 30.0-33.5 Pa Ibr Z.mays. 31.5-33.0 Pa forL. styracijhui, and 32.0-33.5 Pa Ibr P. Uwilii. Each value of F is anaverage Ibr the 30 s prior to measuring photosynthesis. Barsrepresent the maximum S.E. of those means for each species.
1984; Sivak & Walker, 1985). In addition, F may benonphotochemically quenched by factors whichinclude photoinhibition and high temperature(Krause & Weis, 1984) but depending upon environ-tnental conditions, these may not contribute greatlyto total F quenching (Krause & Weis, 1984; Sivak &Walker, 1985).
Following F induction, 'steady-state" F variedinversely with A for approximately 1 h (Fig. 2).During the initial 60-90 s, A reached relatively highvalues in all species, and then decreased during theinitial 1000 s. This deci ease apparently was a leafresponse to frequent contact by the photosynthesiscuvette. Increased A after approximately 1000 sappeared to result from less frequent perturbation. Inboth the C, and C4 species, F decreased withincreasing A (Fig. 3) as a result of photochemical andnon-photochemical quenching (Krause & Weis, 1984;Sivak & Walker, 1985).
82 G. A. CARTER et al.
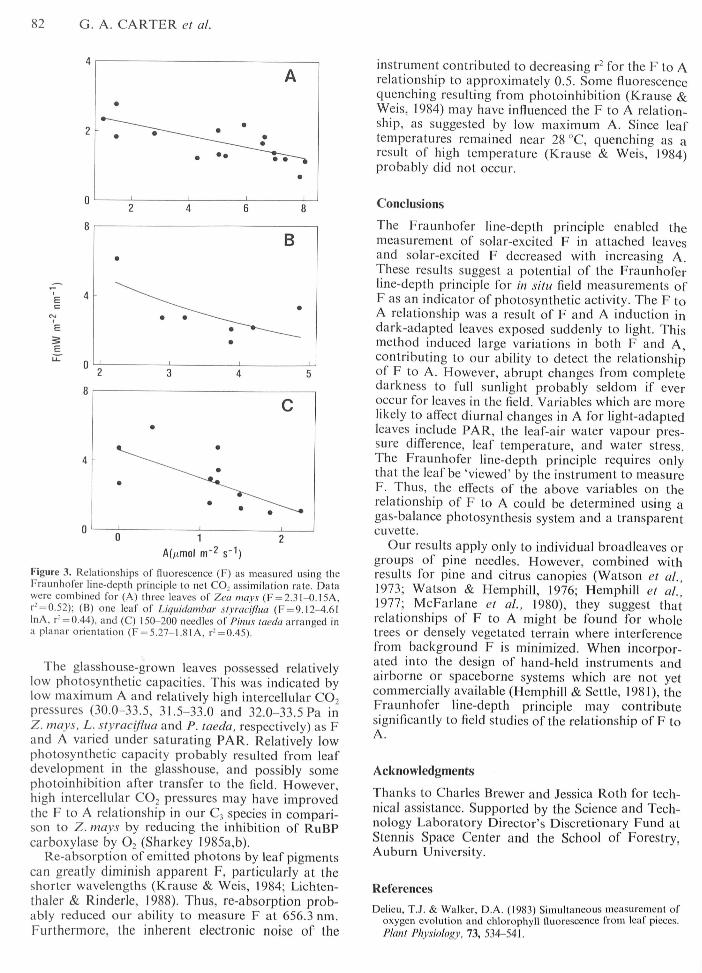
Figure 3. Relationsbips of fluorescence (F) as measured using theFraunhofer line-depth principle lo net CO, assimilation rate. Datawere combined for (A) three leaves of Z.ca mays (F = 2.3I-O.I5A,r̂ = 0.52); (B) one leaf of Liquidambar .slvracijlua (F = 9.12-4.6IInA, r- = 0.44), and (C) 150-200 needtes ol'Pinus lacda arranged ina planar orientation (F = 5.27-1.81 A. r' = 0.45).
The glasshouse-grown leaves possessed relativelylow photosynthetic capacities. This was indicated bylow maximum A and relatively high intercellular CO,pressures (30.0-33.5, 31.5-33.0 and 32.0-33.5 Pa inZ. mays, L. styraeifiua and P. taeda, respectively) as Fand A varied under saturating PAR. Relatively lowphotosynthetic capacity probably resulted from leafdevelopment in the glasshouse, and possibly somephotoinhibition after transfer to the field. However,high intercellular CO2 pressures may have improvedthe F to A relationship in our C, species in compari-son to Z. mays by reducing the inhibition of RuBPcarboxylase by O, (Sharkey I985a,b).
Re-absorption of emitted photons by leaf pigmentscan greatly diminish apparent F, particularly at theshorter wavelengths (Krause & Weis, 1984; Lichten-thaler & Rinderle, 1988). Thus, re-absorption prob-ably reduced our ability to measure F at 656.3 nm.Furthermore, the inherent electronic noise of the
instrument contributed to decreasing r" for the F to Arelationship to approximately 0.5. Some fluorescencequenching resulting from photoinhibition (Krause &Weis, 1984) may have influenced the F to A relation-ship, as suggested by low maximum A. Since leaftemperatures remained near 28 °C, quenching as aresult of high temperature (Krause & Weis, 1984)probably did not occur.
Conclusions
The Fraunhofer line-depth principle enabled themeasurement of solar-excited F in attached leavesand solar-excited F decreased with increasing A.These results suggest a potential of the Fraunhoferline-depth principle for in situ field measurements ofF as an indicator of photosynthetic activity. The F toA relationship was a result of F and A induction indark-adapted leaves exposed suddenly to light. Thismethod induced large variations in both F and A,contributing to our ability to detect the relationshipof F to A. However, abrupt changes from completedarkness to full sunlight probably seldom if everoccur for leaves in the field. Variables which are morelikely to affect diurnal changes in A for light-adaptedleaves include PAR, the leaf-air water vapour pres-sure difference, leaf temperature, and water stress.The Fraunhofer line-depth principle requires onlythat the leaf be 'viewed' by the instrument to measureF. Thus, the effeets of the above variables on therelationship of F to A could be determined using agas-balance photosynthesis system and a transparentcuvette.
Our results apply only to individual broadleaves orgroups of pine needles. However, combined withresults for pine and citrus canopies (Watson et al.,1973; Watson & Hemphill, 1976; Hemphill et al',1977; McFarlane et al., 1980), they suggest thatrelationships of F to A might be found for wholetrees or densely vegetated terrain where interferencefrom background F is minimized. When incorpor-ated into the design of hand-held instruments andairborne or spaceborne systems which are not yetcommercially available (Hemphill & Settle, 1981), theFraunhofer line-depth principle may contributesignificantly to field studies of the relationship of F toA.
Acknowledgments
Thanks to Charles Brewer and .lessica Roth for tech-nical assistance. Supported by the Science and Tech-nology Laboratory Director's Discretionary Fund atStennis Space Center and the School of Forestry,Auburn University.
References
Delieu, T..I. & Walker. D.A. (1983) Simultaneotis measurement ofoxygen evolution and ebloropliyll fluorescence from leaf pieces.Plain Plmhhfiy, 73, 5.34-.S4I.

SOLAR-EXCITED FLUORESCENCE AND PHOTOSYNTHESIS 83
Govindjee (ed.) (1975) Biocncrgctics of Pholosynthesis. AcademicPress, New York.
G.ovindjee, Amesz, .1. & Fork, D.C. (eds) (1986) Light Emission byPlains and Bacteria. Academic Press, New York.
Grainger. J.F. & Ring, J. (1962) The luminescence of the lunarsurfaee. In Physics and Aslronoiny of ilie Moon (ed. Z. Kopal).pp. 385-405. Academic Press. New York.
Hempbill. W.R.. Watson. R.D.. Bigetow. R.C. & Hes.sen, T D .(1977) Measurement of tuminescenec of geocheniically stressedand other materials. U.S. Geological Survey Professional Paper1015, 93- l t2 .
Hemphitt. W.R. & Settte, M. (1981) Work.shop on apptiealions oftuminesccnce techniques to earth resource studie.s. LPI TechnicalReport 81-03. Lunar and Planetary Institute, Houston, TX.104 pp.
Ireland, C.R.. Long, S.P. & Baker, N.R. (1984) The retationshipbetween carbon dioxide fixation and chlorophyll a fluoresceneeduring induction of photosynthesis in maize leaves at diflerenttemperatures and earbon dioxide concentrations. Planla. 160,550-558.
Janza, F.J. el al. (1975) Interaction mechanisms. In Manual ofRemote Sensin)i. Vol. 1, Atnerican Society of Remote Sensing,Falls Chureh, Virginia.
Kozyrev. N.A. (1956) The luminescence of the lunar surface andintensity of the solar corpuscular radiation. Izvestia KrymskoiAstrojizitcheskoy Otiserralorii. 16, 148-161.
Krause, G.H. & Weis, E. (1984) Chlorophyll fluorescence as a toolin plant physiology. II. Interpretation of fluoreseence signals.Photosynthe.sis Re.teareh, 5, 139-157.
Lichtenthaler, H.K. & Rinderle. U. (1988) The role of chlorophyllI1tiore.scenee in the deteetion of stress conditions In plants.C.R.C. Critical Reviews in Analytical Chemistrv. 19, suppl. 1,S29-'S85.
Maas, F.M., van Loo, E.N. & van Hasselt. P.R. (1988) Ellect oflong-term HjS fumigation on photosynthesis in spinach:correlation between CO, lixalion and chlorophyll a fluorescence.P/ivsiologia Planlarum, 72, 77-83.
McFarlane. .I.C., Watson. R.D.. Theisen, A.F., Jackson, R.D..Ehrler, W.L.. Pinter, Jr, P.J.. Idso, S.B. & Reginato, R.J. (1980)Plant stress detection by remote measuretiient of fluorescence.Applied Optics. 19, 3287-3289.
Nobel, P.S. (1983) Biophysical Plant Physiology and Ecology. W.H.Freeman & Co., San Franeiseo.
Plascyk, .I.A. (1975) The MKll Fraunhofer line di.scriminalor(FLD-Il) for airlTOine and orbital remote sensing of solar-stimulated luminescence. Optical Engineering. 14, 339-346.
Plascyk, J.A. & Gabriel, F.C. (1975) The Fraunhofer linediscriminator MKII-an airborne instrument lor precise andstandardized eeological lumineseenee measurement. IEEETraiisaetions on Inslrumenlalion and Measuremenl. llVI-24,306-313.
Sharkey. T.D. (1985a) Photosynthesis in intaet leaves of C, plants;physics, physiology, and rate limitations. The Botanical Review,51, 53-105.
Sharkey, T.D. (1985b) Steady-state room temperature fiuoreseeticeand CO, assimilation rates in intact leaves. PhotosynthesisResearch'. 7, 163-174.
Sivak. M.N., Cerovic. Z.G. & Walker. D.A. (1984) Simultaneousmeasurement of chl a fluoreseence and photosynthetic O,evolution in systems of deereasing complexity (from the leaf toIhe reconstiltiled chloroplast system. Advances in PhotosynthesisResearett. 3, (i6l-664.
Sivak. M.N. & Walker. D.A. (1985) Chlorophyll a fluorescence;can it shed light on fundamental questions in earbon dioxidefixation? Ptanl. Cell and Environment. 8, 439^48.
Sivak. M.N.. Heber, U. & Walker, D.A. (1985) Chlorophyll afluoreseenee and light-.scattering kinetics displayed by leavesduring induetion of photosynthesis. Ptanta. 163, 419^23.
Stoerz, G.E.. Hemphill, W.R. & Markle. D.A. (1969) Airbornefluorometer applieable to marine and estuarine studies, .lournalof the Marine Technical Society. 3, 11-26.
Walker. D.A., Horlon. P.. Sivak. M.N. & Quick. W.P. (1983a)Anti-parallel relationship between O, evolution and slowfluorescence induction kinetics. Pholobiochentistry andPhotobiophY.sics. 5, 35-39.
Walker. D.\.. Sivak. M.N.. Prinsley, R.T. & Cheesbrough. J.K.(1983b) Simultaneous measurement of o.scillations in oxygenevolution and ehlorophyll a fluorescenee in leaf pieces. PlantPhYsiologY. 73, 542-549.
Walker. D^A.. Sivak. M.N. & Cerovic. Z.G. (1984) Somerelationships between photosyntlietie carbon metabolism andchloropliyll a fluorescenee. .Advances in Photosynthesis Research.3, 645-652.
Watson, R.D. & Hemphill. W.R. (1976) Use of an airborneFraunhofer line discriminator for the detection of solar stimulatedluminescence. USGS Open-File Report 76-202.
Watson. R.D.. Hemphill. W.R. & Hessin. T.D. (197.3)Quantifieation of the lumine.scence intensity of naturalmaterials. Symposium for tiie management and utilization ofremote sensing data, Oet. 29-Nov. 1. Sioux Falls. SouthDakota. Proeeeding.s of the .imerican Society oJPhoiogrammelry. 364-376.
Weast. R.C. Astle. M.J. & Beyer, W.H. (eds) (1984) CRCHandbook of Chemistry and Physics. CRC Press. Boea Raton.Florida.
Yakir. D.. Rudich. J. & Bravdo. B.A. (1985) Photoaeoustie andfluorescenee measurements of the chitting response and theirretationstiip to carbon dioxide uptake in tomato plants. Planta.164, 345-353.






























