Embed Size (px)
Citation preview

303
Chloride channels: a molecular perspective Thomas J Jentsch
Plasma membrane Ct- channels perform a variety of
functions, including control of excitability in neurons and
muscle, cell volume regulation and transepithelial transport.
Structurally, three classes of Cl- channels have been
identified: ligand-gated, postsynaptic Cl- channels (e.g. GABA
and glycine receptors); the cystic fibrosis transmembrane
conductance regulator (CFTR) Cl- channels (which belong
to the traffic ATPase superfamily); and the CLC family of
Cl- channels. Recent developments of note include further
characterization of the expanding CLC Cl- channel family,
advances in understanding the regulation of the CFTR
Cl- channel and its emergent role as a regulator of other
channels, clarification of issues related to swelling-activated
Cl- channels, and the discovery that several co-transporter
molecules are now known to induce Cl- currents in Xenopus
oocytes.
Address Centre for Molecular Neurobiology Hamburg (ZMNH), Hamburg University, Martinistrasse 52, O-20246 Hamburg, Germany
Abbreviations CFTR cystic fibrosis transmembrane conductance regulator GABA yaminobutyric acid NBF nucleotide-binding fold
Current Opinion in Neurobiology 1996, 6:303-310
0 Current Biology Ltd ISSN 0959-4388
Introduction Chloride channels are found in the plasma membrane of probably all eukaryotic cells. In contrast to the intracellular concentration of ions such as Caz+ or Na+, the intracellular concentration of Cl- is close to its electrochemical equilibrium. Thus, Cl- has no established function as a second messenger, and is only rarely involved in electrical excitation, Rather, in a way similar to K+ channels, Cl- channels dampen electrical excitability. This is most evident in the nervous system, where GABA and glycine receptors generally generate inhibitory postsynaptic currents, and in skeletal muscle. Mutations in glycine receptor subunits result in hyperekplexia (startle disease) [l], which is attributable to a lack of inhibitory input in a subset of neurons. Furthermore, mutations in the skeletal muscle Cl- channel protein CLC-1 lead to muscle hyperexcitability, resulting in myotonia [2,3]. Interestingly, GABA receptors may have, as an exception, excitatory effects in certain neurons where co-transporters can raise the intracellular Cl- concentration above its electrochemical equilibrium [4].
Chloride is also important for the osmotic control of water flow. Although Cl- is accompanied by K+ when its efflux via swelling-activated Cl- channels counteracts osmotic swelling, Cl- is often joined by Na+ when it creates tiny transepithelial osmotic gradients necessary for fluid secretion or absorption. Being not far from its electrochemical equilibrium, transport of Cl- across both membranes (apical and basolateral) is energetically cheap. The importance of Cl- channels for transepithelial transport is apparent from disorders such as cholera or cystic fibrosis, which are attributable to an activation or mutational inactivation, respectively, of CAMP-regulated Cl- channels. Further, the recent discovery that mutations in a kidney Cl- channel (CLC-5) lead to kidney stones [S”] again suggests a defect in transepithelial transport.
Cl- channels also fulfil important roles in intracellular organelles, including endocytotic and synaptic vesicles. This topic has been reviewed recently f6] and will not be covered here. Instead, this review will focus primarily on Cl- channels whose molecular identity is known, without, however, considering postsynaptic GABA and glycine receptors. The emphasis is placed on the most recent developments in this field. Focusing exclusively on Cl- channels in the nervous system would ignore many important developments, and is thus avoided.
CLC chloride channel proteins The CLC proteins form the only known large gene family of Cl- channels. They show a high degree of evolutionary conservation and homologues are present in yeast and bacteria. CLC proteins comprise about 12 transmembrane domains (Figure lb) 17,801 and function as multimers [2,9].
In mammals, nine different CLC genes are currently known. On the basis of homology, these can be assigned to one of three branches (Figure 2). The first branch, which is most closely related to the Tof~%!o channel CLC-0 [lo], includes: the muscle Cl- channel CLC-1, which modulates muscle excitability and whose inactivation causes myotonia f&3]; CLC-2, a ubiquitously expressed channel activated by cell swelling or hyperpolarization [ll]; and the kidney-specific transcripts CLC-Ka and CLC-Kb (or CLC-Kl and CLC-K2 in the rat) [12-141. The second branch includes CLC-3 and CLC-4, two putative channels expressed in several different tissues such as brain and kidney [7,15-171. An important new addition is CLCd [l&19], the functional inactivation of which leads to kidney stones [5**,18]. Finally, the third and newest branch comprises CLC-6 and CLC-7 [ZO], which are found in most tissues but are not expressed functionally as Cl- channels. Below, I describe in more

304 Signalling mechanisms
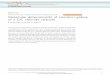
Figure 1
(a) GABA and glycine receptors (b) CLC Cl’ channels
D9.. .................. D12 ................... ...................
(c) CFTR
Q 1996 Current Opinion in Neurobiology
Topology models for GABA and glycine receptors, CLC Cl- channels, and CFTR. (a) GABA and glycine receptors are typical members of the ligand-gated ion channel superfamily, which includes the nicotinic acetylcholine receptor. Members have four transmembrane domains (Ml-M4) and probably function ss (mostly hetero-) pentamers. (b) CLC Cl- channels have about 12 tran~embrane domains (01 -Dl2), the exact topology of which is not yet known [8*,29]. A previous model I1 01 had to be modified, as the loop between D8 and 09 is now known to be glycosylated [9,13]. Analysis of new CLC genes suggests that D4 is not a conserved hydrophobic domain, eliminating it as a transmembrane span [20]. Whereas D2 is thought not to cross the membrane in an alternative model [91, several algorithms predict that it is a very good candidate for a transmembrane domain. Further, deletion of this domain destroys channel activity [5**,7] and results in kidney stones [5**]. The topology in the broad hydrophobic region OS-D1 2 is obscure, and the conserved region 013 is known to be cytoplasmic [l 11. (c) CFTR has two blocks of six putative transmembrane domains (TM1 -TM6 and TM7-TM12), which are followed by the nucleotide-binding folds NBFl and NBFP, respectively, The regulatory ‘R’ domain must be phosphorylated before regulation of channel opening by ATP (at the NBFs) can occur.
detail exciting developments regarding specific CLC Cl- mechanism by which a defect in a Cl- channel leads to channel proteins. hypercalciuria and proteinuria is still unclear.
CLC-5 CLC-1
Dent’s disease is a rare X-linked form of nephrolithiasis In contrast, the pathogenesis of Cl- channel myotonia associated with hypercalciuria and low-molecular-weight is well understood [‘2,3,21,22]. In the absence of the proteinuria. It is now clear that Dent’s disease and two high resting Cl- conductance mediated by CLC-1, the similar disorders are caused by loss-of-function mutations muscle membrane is electrically hyperexcitable. Whereas in CLC-5 [5**], which is mainly expressed in kidney mutations with a complete loss of function cause recessive [l&19]. In the rat, transcripts are present in large portions myotonia, dominant myotonia is attributable to dominant- of the nephron, including late parts of the proximal negative effects of mutant subunits in the multimeric tubule, and significant expression is also observed in brain channel [Z]. Many dominant mutations shift the voltage 1191. When expressed in Xenopus oocytes, both rat and dependence of CLC-1 to positive voltages, and even human CLC-5 elicit anion currents with a conductivity wild-type/mutant heteromeric channels can no longer sequence N03-->Cl-> Br> I-. Mutations found in kidney contribute to the repolarization of action potentials [Zl]. stone pedigrees specifically reduced or abolished these These mutations, as well as a recessive mutation that alters currents [S”]. Curiously, CLC5 elicits currents only at the voltage dependence in a different way [ZZ], are located very positive, probably unphysiological, voltages. The at very different positions in the CLC-1 channel protein.

Chloride channels: a molecular perspective Jentsch 305
Dendrogram showing the degree of similarity between the known mammalian members of the CLC superfamily. Members of the fmt branch, which includes CLC-1, CLC-2, CLC-Ka and CLCKb, are those most closely related to the Torpedo channel UC-O, the founding member of this gene family [lo]. The second branch, including CLC-3, CLC-4 and CLC-5, contains those members most closely related to the yeast homologue SCCLC 1301. (Interestingly, the recent completion of the yeast genome project indicates that there is only one CLC gene in yeast, which we suggest calling ‘scCW.) Finally, a recent add&ion to the CLC sup&amity is the CLC-6 and CLG7 branch. Tissue expression patterns and proposed functions are also indicated.
Thus, it seems unlikely that all these mutations affect gate structures directly.
CLC-0 Whereas many cation channels sense the membrane voltage using charged amino acids in transmembrane domains, ‘fast’ gating of CLC-0 can be described by a fundamentally different model in which the permeating anion itself provides the ‘gating charge’ [@I. Opening rates are increased by the presence of anions close to the internal end of the pore. This model provides a simple explanation for the observed voltage and Cl--concentration dependence of CLC-0 gating, and is supported by the analysis of mutations at the very end of the transmembrane spans. These mutations modulate gating, single-channel conductance, and ion selectivity, suggesting that this region is important for permeation. Ion selectivity is however affected by mutations in other regions as well [Z], so it has not yet been possible to convincingly identify all the structures directly lining the pore region.
CLC-2
CLC-2 can be activated by cell swelling or strong hyperpolarization in Xenopus oocytes [l I]. Superficially similar currents are present in native cells, including epithelia [23-251, neurons [26,27] and osteoblasts [28], but
the osmosensitivity has often not been checked and may even be reversed [28]. Although northern analysis suggests that CLC-2 is expressed ubiquitously, in situ hybridization reveals that CLC-2 is expressed differentiaIly in brain 1271, according to a pattern that differs from that of CLC3 [15,17]. It was proposed that CLC-2 serves to prevent neuronal Cl- accumulation above equilibrium, thereby modulating the effects of postsynaptic GABA receptors [4,26]. Consistent with this hypothesis, expression was observed to be absent or low in neurons exhibiting a ‘paradoxical’ excitation by GABA, attributable to a Cl- efflux via GABA receptors in cells with a high internal Cl- concentration [27].
CLC-4, CLC-6, CLC-7 and CLC-KS
Many CLCs cannot be expressed functionally (such as CLC4, CLC-6 and CLC-7) [7,17,20,29], and for others expression is controversial. These proteins are best classified as putative Cl- channels. Some groups were unable to find novel Cl- currents with both CLC-KS and CLC-3 [13,17,19]. By contrast, another group reported slightly outwardly rectifying currents with CLC-Kl [lZ], CLC-K2 1141 and CLC3 (151. Identical currents were obtained with a splice variant of CLC-K2 from which domain D2. (see Figure 1) has been deleted [14]. This is surprising as D2 is predicted to be a transmembrane

306 Signalling mechanisms
domain, and as a similar deletion in CLC-5 eliminates channel activity and leads to kidney stones [5"'].
The lack of functional expression could be explained by complicated regulation of channel activity or by the lack of important subunits, among others. Alternatively, these putative channels might serve intracellular roles. The iron-repressible petite phenotype (deficiency in respiration) of a yeast strain deleted for the yeast CLC homologue scCLC encoded by the GEF-1 gene ([30]; see also Figure 2 legend) may point in the direction of the latter explanation.
Multiple functions of CFTR An increasing number of reports indicate that CFTR, the gene product defective in cystic fibrosis, performs regulatory roles in addition to being a cAMP-activated CI-channel . This additional function has been signifi- cantly, albeit indirectly, bolstered by the discovery that the sulfonylurea receptor (SUR) regulates a member of the inwardly rectifying K ÷ channel family (BIR) to yield ATP-semitive K ÷ channels [31]. Both C F T R and the sulfonylurea receptor belong to the superfamily of traffic ATPases, combining two blocks of transmembrane spans with two nucleotide-binding folds (Figure lc), although their topologies differ in detail.
Data obtained previously indicated that C F T R regulates an outwardly rectifying CI- channel expressed in many cells [32]. Intriguing experiments now suggest that this regulation is mediated by ATP, which leaves the cell in response to C F T R stimulation [33°,34]. This exodus might be achieved via a pathway closely regulated by CFTR, or directly via the C F T R pore itself, as suggested previously [35]. On the other hand, no effect of C F T R on ATP efflux was found in a different system [36]. A direct ATP-channel function of C F T R may be surprising for a pore otherwise highly selective for small anions. Indeed, several groups have reported recently that C F T R does not conduct ATP [37]. Extracellular ATP may activate outwardly rectifying CI- channels by interacting with purinergic receptors [33°,38] Whether this activation is mediated by direct interaction, intracellular Ca 2÷, and/or other pathways is not yet clear, and the molecular identity of the outward rectifier is unknown. Intriguingly, some mutations associated with cystic fibrosis have been shown to have differential effects on intrinsic CI- channel and regulator activities [39].
In addition to a defective CI- conductance, airway epithelia of cystic fibrosis patients show an increase in Na÷ currents. It is now known that C F T R leads to cAMP-stimulated inhibition of co-transfected epithelial Na ÷ channels [40"]. Although this effect is small in fibroblasts, the inhibition observed in epithelial cells seems substantial. Interactions between both channels were also observed in the Xenopus oocyte expression system [41]. The mechanism of this interaction is still
unclear, but these observations suggest that a search for interactions with other channels might be fruitful. Indeed, first reports indicate that C F T R may also regulate inwardly rectifying K ÷ channels (CM McNicholas, WB Guggino, SC Hebert, ME Egan, Pediatr Pulmonol 1995, 19:77), and indirect evidence suggests that C F T R may also affect volume-activated K ÷ channels [42]. Clearly, more research is needed to elucidate these effects, as it would be surprising if C F T R were involved in all these diverse processes.
In view of these regulatory roles, the additional function of CFTR as a CI- channel is surprising, but well established. The ATP-channel model [33"], although very controversial [36,37], reconciles both functions. As C F T R belongs to the traffic ATPase family, several studies have focused on the roles of its two nucleotide-binding folds (NBFs) [43",44"°,45,46]. In contrast with 'normal' transport ATPases, in which ATP is hydrolyzed upon each active transport cycle, the passive pore of the C F T R channel is thought to be regulated by ATP hydrolysis. This regulation can only occur after C F T R has been phosphorylated by protein kinase A at the 'R domain'. Extending previous results [47], the new data suggest that ATP hydrolysis at NBF1 is important for channel opening. Binding of ATP at the second NBF (NBF2) is thought to stabilize the open state, and ATP hydrolysis at NBF2 facilitates channel closure [44"%46]. A previously undetected second open state (02) with a slightly different conductance has been exploited in an elegant series of experiments [44"]. The results suggest that ATP hydrolysis at NBF2 leads to a transition from the first open state (O1) to O 2, from which the channel can close spontaneously [44"']. Although these studies only indirectly suggest that ATP is hydrolyzed by CFTR, new biochemical data indicate that NBF1 (in a fusion protein) may indeed function as an ATPase [48].
The most frequent mutant form of CFTR, AF508, does not reach the cell surface but instead is degraded in a pre-Golgi compartment. It has now been shown that, rather like other membrane proteins, both wild-type and mutant C F T R proteins are degraded in part by the ubiquitin-proteasome pathway [49,50].
Although changes in ion selectivity have been corre- lated with mutations in several transmembrane domains ([51,52]; but, see [53]), the pore of this extensively studied molecule is not yet well understood. It is a sure bet that C F T R still has many surprises in store.
Chloride channels and regulation of cell volume The regulation of cell volume in response to hypo-osmotic swelling is of crucial importance for most cells. This is accomplished by the regulated loss of osmolytes, which is mediated in part via K ÷ and CI- channels. A first report that expression of the multidrug resistance P-glycoprotein

Chloride channels: a molecular perspective Jentsch 307
(MDR1) generates volume-activated CI - channe l s [54] has stirred considerable interest and controversy over the years [55-60]. It now seems clear that MDR1 is not itself a CI- channel. Instead, it is thought to confer regulation by protein kinase C onto endogenous swelling-activated CI- channels [61]. As MDR1 is a traffic ATPase (indeed, a typical one), this suggestion fits very well the regulatory roles of C F T R and the sulfonylurea receptor. (Interestingly, but also controver- sially, M D R l - - l i k e C F T R - - h a s been suggested to be an ATP channel [62].) T h e mechanism of interaction between MDR1 and the volume-activated channel is nonetheless not known, and the regulatory role has been questioned also [63].
Osmotic swelling activates CLC-2 in the oocyte expression system, and structures important for this regulation have been localized to its amino terminus [11]. CLC-2 does not however underlie the typical volume-activated CI- channel observed in numerous cell types. In contrast with CLC-2, the typical volume-activated current displays an I->CI- permselectivity, is outwardly rectifying, deactivates at positive potentials, and depends on intracellular ATP. A detailed study in glioma cells [64 °] has shown that this channel has a single-channel conductance of 15-50pS (depending on voltage), resolving a discrepancy between single-channel [65] and noise [66,67] analysis, which suggested a lower value. This current is blocked by extracellular nucleotides in several cell types [68-70], resembling the current seen upon overexpression of plcln, a putative C I - c h a n n e l [71]. Indeed, antisense oligonucleotides [70] or antibodies [72] directed against plc1 n inhibit the volume-activated CI- current in native cells. A convincing proof that plci n and the CI- channel are one and the same is missing, however, and it is possible that plcI n instead interacts with endogenous CI- channels [72].
In the case of regulatory volume decrease, a loss of intracellular organic osmolytes (taurine and myo-inositol, among others) may be equally important as a loss of KCI. An increasing body of evidence suggests that this loss may involve the same swelling-activated anion channel [73-75].
C h l o r i d e currents a s s o c i a t e d w i th co - t ranspor te rs Several co-transport or counter-transport molecules elicit anion currents when expressed in Xenopus oocytes. For instance, several members of an excitatory amino-acid/Na ÷ co-transporter family were shown to yield aspartate- evoked CI- currents when expressed in Xenopus oocytes [76,77]. These rather small currents were not stoichio- metrically coupled to amino acid transport, suggesting that these transporters may have an additional function of a 'ligand-gated' CI- channel. This could dampen cell excitability upon neurotransmitter re-uptake.
In addition, expression of a renal sodium-phosphate co-transporter (NaPi-1) in Xenopus oocytes yielded quite large, time-independent, slightly outwardly rectifying anion currents with an I-->CI- selectivity [78], which superficially resemble the currents reported, for example, for CLC-3 [15]. This current was not directly dependent on the substrates of the co-transporter (i.e. Na ÷ and PO42-).
Finally, expression in Xenopus oocytes of the trout erythrocyte anion exchanger (AE, or band 3), but not of mouse anion exchanger AE1, induced increased taurine fluxes and a sizeable CI-conductance that was only slightly outwardly rectifying [75]. Analysis of chimeric transporters has shown that an extracellular loop and carboxy-terminal transmembrane domains are necessary for the induction of these currents.
It has not yet been established whether these proteins pos- sess dual transporter/channel activities, or whether their expression in Xenopus oocytes activates endogenous chan- nels. Oocytes possess a variety of endogenous CI- chan- nels, w h i c h - - i n some cases, very reproducib ly- -may be activated by the overexpression of foreign proteins that are not themselves channels. Thus, expression of diverse membrane proteins elicits hyperpolarization-activated an- ion currents in Xenopus oocytes [20,79°,80]. These currents were previously thought to be specific for phospholemman [81]. Several criteria must be met before a cloned cDNA can safely be assumed to encode a channel [29]. One can, however, by no means exclude the possibility that molecules have dual transporter/channel functions. This scenario seems particularly likely for the aspartate-gated CI- currents.
C o n c l u s i o n s Though three gene families of CI- channels are known, our molecular picture of CI- channels is far from complete. Several important channels have not yet been cloned, including the seemingly ubiquitous volume-activated CI- channel (which may also conduct organic osmolytes), and Ci- channels activated by intracellular CaZ+. Homol- ogy cloning continues to uncover new members of the CLC family, but many of these proteins have yet to be assigned a function. In the absence of unambiguous functional expression, it is even possible that some are not even CI- channels. Many proteins overexpressed in Xenopus oocytes (and possibly other cells) yield CI- cur- rents, and it is often difficult to exclude the fact that they only activate endogenous channels. Surprisingly, C F T R is the only member of its gene family that has convincingly shown to be a CI- channel. Its regulatory effects on other channels, which have also been demonstrated wi th other traffic ATPases [82], are still poorly understood, and represent an exciting field for future discoveries.

308 Signalling mechanisms
In addition to the well recognized roles of postsynaptic ligand-gated CI- channels for signal transduction in the CNS, the role of voltage-activated CI -channe l s in controlling electrical excitability becomes increasingly clear. This is most obvious for skeletal muscle [2], but probably is also true for neuronal cells [26,27], and it would not be surprising if genetic defects in brain CI- channels turn out to be the basis for certain inherited neurological disorders. The emerging complex regulations of ion channels, as exemplified by the multiple functions of CFTR, is likely to add an additional level of complexity to neurobiology as well.
Acknowledgements l thank Michael Pusch, Blanche Schwappach, and Klaus Steinmeyer for critical reading of the manuscript. Work in the author's laboratory is supported by the Deutsche Forschungsgemeinschaft, the US Muscular Dystrophy Association, and the Fonds der Chemischen lndustrie.
References and recommended reading Papers of particular interest, published within the annual period of review, have been highlighted as:
• of special interest • • of outstanding interest
1. Rajendra S, Schofield PR: Molecular mechanisms of Inherited startle syndrome. Trends Neurosci 1995, 18:80-82.
2. Steinmeyer K, Lorenz C, Pusch M, Koch MC, Jentsch TJ: Multimeric structure of ClC-1 chloride channel revealed by mutations in dominant myotonla congenita. EMBO J 1994, 13:737-743.
3. Meyer-Kleine C, Steinmeyer K, Ricker K, Jentsch TJ, Koch MC: Spectrum of mutations In the major human skeletal muscle chloride channel gene (ClCN1) leading to myotonla. Am J Hum Genet 1995, 57:1325-1334.
4. Staley K, Soldo BL, Proctor WR: ionic mechanism of neuronal excitation by Inhibitory GABA A receptors. Science 1995, 269:977-981.
5. Lloyd SE, Pearce SHS, Fisher SE, Steinmeyer K, Schwappach B, • e Scheinman S J, Harding B, Bolino A, Devoto M, Goodyer Pet al.:
A common molecular basis for three Inherited kidney stone diseases. Nature 1996, 379:445-449.
Dent's disease, X-linked recessive nephrolithiasis, and X-linked recessive hy- pophosphatemic rickets (XLRH) are three kidney stone disorders that share the symptoms hypercalciuria and proteinuria. All three diseases are shown to be caused by inactivating mutations in the CLC-5 CI- channel that, in humans, are very kidney specific. These mutations include non-sense and mis-sense mutations, as well as an in-frame deletion of transmembrane do- main D2. Functional expression in Xenopus oocytes shows that these muta- tions specifically reduce or abolish CLC-5 currents. As proteins are mainly re-absorbed by the proximal tubule, it is important to note that CLC-5 is also expressed in that segment [19]; however, the mechanism by which CLC-5 inactivation leads to hyporcalciuria and proteinuria is unclear.
6. AI-Awqati Q: Chloride channels of Intracellular organelles. Curr Opin Cell Biol 1995, 7:504-508.
7. Jentsch TJ, Gi~nther W, Pusch M, Schwappach B: Properties of voltage-gated chloride channels of the ClC gene family. J Physiol (Lond) 1995, 482:19S-25S.
8. Pusch M, Ludewig U, Rehfeldt A, Jentsch TJ: Gating of the • voltage-dependent chloride channel ClC-0 by the permeant
anion. Nature 1995, 373:527-531. This paper provides evidence for a gating model that differs in a fundamental way from the intrinsic voltage sensor model for cation channels. Open proba- bility of the 'fast' gate of the Torpedo channel CLC-O is shown to depend with a slope of -50 mV to -60 mV per tenfold change of external, but not internal, e l - concentration. This effect and the nominal gating charge of 1 can be explained by a simple model in which CI- itself provides the gating charge. It senses the voltage as it crosses the membrane, and its presence at the end of the pore facilitates channel opening. Further, both conductance and gating show an anomalous mole fraction behaviour, and mutations decreasing the ion selectivity of conductance also decrease the ion selectivity of gating.
9. Middleton RE, Pheasant D J, Miller C: Purification, reconstitution, and subunlt composition of a voltage-gated chloride channel from Torpedo electroplax. Biochemistry 1994, 33:13189-13198.
10. Jentsch TJ, Steinmeyer K, Schwarz G: Primary structure of Torpedo mermorata chloride channel Isolated by expression cloning in Xenopus oocytes. Nature 1990, 348:510-514.
11. Gr0nder S, Thiemann A, Pusch M, Jentsch TJ: Regions involved in the opening of CLC-2 chloride channel by voltage and cell volume. Nature 1992, 360:759-762.
12. Uchida S, Sasaki S, Nitta K, Uchida K, Horita S, Nihei H, Marumo F: Locellzation and functional charactarlzation of rat kidney-specific chloride channel, ClC-KI. J C/in Invest 1995, 95:104-113.
13. Kieferle S, Fong P, Bans M, Vandewalle A, Jentsch TJ: Two highly homologous members of the ClC chloride channel family expressed in both rat and human kidney. Proc Nat/Acad Sci USA 1994, 91:6943-6947.
14. Adachi S, Uchida S, Ito H, Hata H, Hiroe M, Marumo F, Sasaki S: Two Isoforms of a chloride channel predominantly expressed In thick ascending limb of Henle's loop and collecting ducts of rat kidney. J Biol Chem 1994, 269:17677-17683.
15. Kawasaki M, Uchida S, Monkawa T, Miyawaki A, Mikoshiba K, Marumo F, Sasaki S: Cloning and expression of a protein kinase C-regulated chlodde channel abundantly expressed in rat brain neuronal cells. Neuron 1994, 12:597-604.
16. Van Slegtenhorst MA, Bassi MT, Borseni G, Wapenar MC, Ferrero GB, De Conciliis L, Rugarli El, Grillo A, Franco B, Zoghni HY, Ballabio A: A gene from the Xp22.3 region shares homology with voltage-gated chloride channels. Hum Mol Genet 1994, 3:547-552.
17 Borsani G, Rugarti El, Taglialatela M, Wong C, Ballabio A: Characterization of a human and murlne gene (CLCN3) sharing similarities to voltage-gated chloride channels and to a yeast Integral membrane protein. Genomics 1995, 27:131-141.
18. Fisher SE, Black GCM, Lloyd SE, Hatchwell E, Wrong O, Thakker RV, Craig IW: Isolation and partial characterization of a chloride channel gene which Is expressed In kidney and is a candidate for Dent's disease (an X-linked hereditary nephrollthlasls). Hum Mol Genet 1994, 3:2053-2059.
19. Steinmeyer K, Schwappach B, Bens M, Vandewalle A, Jentsch TJ: Cloning and functional expression of rat CLC-5, a chloride channel related to kidney disease. J Biol Chem 1995, 270:31172-31177.
20. Brandt S, Jentsch TJ: CLC-6 and CLC-7 are two novel broadly expressed members of the CLC chloride channel family. FEBS Lett 1995, 377:15-20.
21. Pusch M, Steinmeyer K, Koch MC, Jentsch TJ: Mutations in dominant human myotonia congenlta drastically alter the voltage dependence of the CIC-1 chloride channel. Neuron 1995, 15:1455-1463.
22. Fahlke C, R~idel R, Mitrovic N, Zhou M, George AL: An aspertlc acid residue Important for voltage-dependent gating of human muscle chloride channels. Neuron 1995, 15:463-472.
23. Komwatana P, Dinudom A, Young JA, Cook Dh Characterization of the el- conductance in the granular duct cells of mouse mandibular glands. Pflugers Arch 1994, 428:641-647
24. Fritsch J, Edelman A: Modulation of the hyperpolarlzation- activated el- current in human intestinal T84 cells by phosphorylation. J Physiol (L ond) 1996, 490.1:115-128.
25. Arreola J, Park K, Melvin JE, Begenisich T: Three distinct chloride channels control anion movements in rat parotld aclnar calls. J Physiol (Lond) 1996, 490:351-362.
26. Staley K: The role of an inwardly rectifying chloride conductance in postaynaptlc inhibition. J Neurophysiol 1994, 72:273-284.
27. Smith RL, Clayton GH, Wilcox CL, Esudero KVV, Staley K J: Differential expression of an Inwardly rectifying chloride conductance in rat brain neurons: a potential mechanism for cell-specific modulation of postsynaptic inhibition. J Neurosci 1995, 15:4057-4067.
28. Chesnoy-Marchais D, Fritsch J: Activation by hyperpolarlzatlon and atypical osmosensitivlty of a el - current In rat osteoblestlc cells. J Membr Bio11994, 140:173-188.
29. Jentsch TJ: Molecular physiology of anion channels. Curt Opin Cell Biol 1994, 6:600-606.

Chloride channels: a molecular perspective Jentsch 309
30. Greene JR, Brown NH, DiDomenico BJ, Kaplan J, Eide DJ: The GEF.1 gene of Saccheromyces cerevisiee encodes an integral membrane protein; mutations in which have effects on respiration and Iron-limited growth. Mo/Gen Genet 1993, 241:542-553.
31. Inagaki N, Gonoi T, Clement JP, Namba N, Inazawa N, Gonz~lez G, Aguilar-Bryan L, Seino S, Bryan J: Reconstitutlon of IKMp: an inward rectifier subunlt plus the sulfonylurea receptor. Science 1995, 270:1166-1170.
32. Egan M, Flotte T, Afione S, Solow R, Zeitlin PL, Carter B J, Guggino WB: Defective regulation of outwardly rectifying Cl- channels by protein klnase A corrected by insertion of CFTR. Nature 1992, 358:581-584.
33. Schwiabert EM, Egan ME, Hwang TH, Fulmer SB, Allen SS, • Cutting GR, Guggino WB: CFTR regulates outwardly rectifying
chloride channels through an autocrlne mechanism involving ATR Cell 1995, 81:1063-1073.
This work suggests that CFTR, in addition to being a linear CI- channel, also regulates an outwardly rectifying CI- channel. This regulation depends on the efflux of ATP from the cells after stimulation by protein kinase A. ATP-acavenging systems applied to the extracellular surface prevents this activation, which presumably is mediated via purinergic P2u receptors. The mechanism for this regulation (e.g. direct protein-protein interactions, or second messengers) is not yet clear. Under appropriate conditions, CFTR stimulation elicits currents carried by ATP anions, which either flow through a channel regulated by CFTR or through CFTR itself [35]. The latter model would reconcile the channel and regulator function of CFTR, but is highly controversial (see [36,37]).
34. Jovov B, Ismailov II, Berdlev BK, Fuller CM, Sorscher E J, Dedman JR, Kaetzel MA, Benos D J: Interaction between cystic fibrosis transmembrane conductance regulator and outwardly rectifying chlodde channels. J Biol Chem 1995, 270:29194-29200.
35. Reisin IL, Prat AG, Abraham EH, Amara JF, Gregory R J, Ausiallo DA, Cantiello HF: The cystic fibrosis trensmembrane conductance regulator Is a dual ATP and chloride channel. J Biol Chem 1994, 269:20584-20591.
36. Takahashi T, Matsushita K, Welsh M J, Stokes JB: Effect of cAMP on intracellular and extracellular ATP content of Cl- secratlng epithelia and 3T3 fibroblasts. J Biol Chem 1994, 269:17853-17857.
37. Reddy MM, Quinton PM, Haws C, Wins J J, Grygorczyk R, Tabcharani JA, Hanrahan JW, Gunderson KL, Kopito RR: Failure of the cystic fibrosis transmembrane conductance regulator to conduct ATR Science 1996, 271:1876-1879.
38. Stutts MJ, Rtz JG, Paradiso AM, Boucher RC: Multiple modes of regulation of airway epithelial chloride secretion by extracelluler ATR Am J Physiol 1994, 267:C1442-C1451.
39. Fulmer SB, Schwiebert EM, Morales MM, Guggino WB, Cutting GR: Two cystic fibrosis transmembrane conductance regulator mutations have different effects on both pulmonary phenotype and regulation of outwardly rectified chloride currents. Proc Natl Acad Sci USA 1995, 92:6832-6836.
40. Stutts M J, Canessa CM, Olsen JC, Hamrick M, Cohn JA, • Rossier BC, Boucher RC: CFTR as a cAMP-dependent regulator
of sodium channels. Science 1995, 269:847-850. This important work investigates the effects of CFTR on the epithelial Na + channel ENaC in cells transfected with all three ENaC subunits. Using epithelial cell layers of transfected MDCK cells in CI--free solutions, it is shown that forskolin (which raises cAMP) stimulates short-circuit current in the absence of CFTR. This current is mediated in large part by ENaC in the apical membrane. In cells co-transfected with CFTR, in contrast, cAMP causes a slight inhibition. Although these effects are large, whole-cell cur- rent measurements in transfected 3T3 cells yield similar, but much less im- pressive results. Broadly similar results have been obtained in the Xenopus oocyte expression system [41 ]. The mechanism of interaction between both channel molecules is still unclear. The results of this work are highly relevant for cystic fibrosis, as an increase in Na + re-absorption contributes to lung disease in the patients studied.
41. Mall M, Hipper A, Greger R, Kunzelmann K: Wild type but not ~F508 CFTR Inhibits Na + conductance when coexpressed in Xenopus oocytes. FEBS Lett 1996, 381:47-52.
42. Valverde MA, O'Brien JA, Sep01veda FV, Ratctiff RA, Evans M J, Colledge WH: Impaired cell volume regulation in intestinal crypt epithelia of cystic fibrosis mice. Proc Nat/Acad Sci USA 1995, 92:9038-9041.
43. Carson MR, Travis SM, Welsh M J: The two nucleotide-blndlng • domains of cystic fibrosis transmombrane conductance
regulator (CFTR) have distinct functions in controlling channel activity. J Bio/Chem 1995, 270:1711-1717.
Using patch-clamp analysis of transfected cells, the analysis of a number of CFTR point mutants, and of ATP and non-hydrolyzable analogs indi- rectly, suggests that ATP hydrolysis at NBF1 initiates burst activity (during which the channel fluctuates many times between open and closed states), whereas hydrolysis of ATP at NBF2 terminates the burst. Non-hydrolyzable ATP analogs prolong the duration of only some burets. These conclusions are broadly similar to those presented in [44"].
44. Gunderson KL, Kopito RR: Conformational states of CFTR • . associated with channel gating: the role of ATP binding and
hydrolysis. Cell 1995, 82:231-239. Heavy electrical filtering (10 Hz) allows the detection of a previously unknown open state (O 2) of CFTR reconstituted into lipid bilayere. 02 differs by only 1 pS from the other open state O1. Statistical analysis shows that channel gating predominantly occurs in an asymmetric cycle C->O!--~O2--)C, where C is the closed state. This indicates an input of energy, which is most likely supplied by ATP hydrolysis. Non-hydrolyzable compounds lock CFTR in the O1 state. The predominant irreversible step is the O1--)O 2 transition, which is coupled to ATP hydrolysis at NBF2. These conclusions are supported by the analysis of point mutants. Thus, this elegant study suggests that each cycle of CFTR opening is coupled to the hydmlysia of one ATP at NBF2 (and possibly one ATP at NBF1, the role of which is less clear). The appar- ent contrast with a study on intact cells [43"] - which suggests that ATP hydrolysis at NBF2 terminates a burst of activity - may be attributable, in part, to the heavy filtering employed here, which may not allow the detection of fast closures.
45. Schultz BD, Venglarik C J, Bridges R J, Frizzell RA: Regulation of CFTR Cl- channel gating by ADP and ATP analogues. J Gen Physiol 1995, 105:329-361.
46. Wilkinson D J, Mansoure MK, Watson PY, Smit LS, Collins FS, Dawson DC: CFTR: the nucleotide binding folds regulate the accessibility of the activated stste. J Gen Physiol 1996, 107:103-119.
47. Hwang TC, Nagel G, Naim AC, Gadsby DC: Regulation of the gating of cystic fibrosis transmembrane conductance regulator Cl channels by phosphorylation and ATP hydrolysis. Proc Natl Acad Sci USA 1994, 91:4698-4702.
48. Ko YH, Pedersen PL: The first nucleoflde binding fold of the cystic fibrosis tmnsmembrane conductance regulator can function as an active ATPase. J Bio/Chem 1995, 270:22093-22096.
49. Jensen TJ, Loo MA, Pind S, Williams DB, Goldberg AL, Riordan JR: Multiple proteolytic systems, Including the proteasome, contribute to CFTR processing. Ceil 1995, 83:129-135.
50. Ward LW, Omura S, Kopito RR: Degradation of CFTR by the ubiqultln-proteasome pathway. Ceil 1995, 83:121-127.
51. Anderson MP, Gregory R J, Thompson S, Souza DW, Paul S, Mulligan RC, Smith AE, Welsh M J: Demonstration that CFTR Is a chlodde channel by alteration of its anion selectivity. Science 1991, 253:202-205.
52. McDonough S, Davidson N, Lester HA, McCarty NA: Novel pore- lining residues In CFTR that govem permeation and open- channel block. Neuron 1994, 13:623-634.
53. Carroll TP, Morales MM, Fulmer SB, Allen SS, Rotte TR, Cutting GR, Guggino WB: Alternate translation initiation codons can create functional forms of cystic fibrosis transmembrane conductance regulator. J Biol Chem 1995, 270:11941-11946.
54. Valverde MA, Diaz M, Sep01veda FV, Gill DR, Hyde SC, Higgins CF: Volume-regulated chloride channels associated with the human multldrug-reslstance P-glycoprotein. Nature 1992, 355:830-833.
55. Mori XK, Bond TD, Loo TW, Clarke DM, Bear CE: Failure of P-glycoprotein (MDR1) expressed in Xenopus oocytes to produce swelling-activated chlodde channel activity. J Physiol (Lond) 1995, 468.3:707-714.
56. Rasola A, Galietta I_IV, Gruanert DC, Romeo G: Volume-sensitive chloride currents in four epithelial cell lines are not directly correlated to the expression of the MDR-1 gene. J Biol Chem 1995, 269:1432-1436.
57. De Greef C, Sehrer J, Viana F, Van Acker K, Eggermont J, Martens L, Raeymaekere L Droogmans G, Nilius B: Volume-activated chloride currents are not correlated with P-glycoprotein expression. Biochem J 1995, 307:713-718.
58. Luckie DB, Krouse ME, Harper KL, Law TC, Wine J J: Selection for MDR1/P-glycoprotein enhances swelling-activated

310 Signalling mechanisms
K + and Cl- currents in NIH/3T3 ceils. Am J Physiol 1994, 267 :C650-C658.
59. Ehring GR, Osipchuk YV, Cahalan MD: Swelling-activated chlodde channels In multidrug-sensitive and -resistant cells. J Gen Physio11994, 104:1129-1161.
60. Kunzelmann K, Slotki IN, Koslowski T, Ausiello DA, Greger R, Cabantchik Zh Effects of P-glycoproteln expression on cyclic AMP and volume-activated ion fluxes and conductances in HT-29 colon edenocsrcinoma cells. J Cell Physio/1994, 161:393-406.
61. Hardy SP, Goodfellow HR, Valverde MA, Gill DR, Sept~lveda IV, Higgins CF: Protein klnase C-mediated phosphorylation of the human multidrug resistance P-glycoproteln regulates cell volume-activated chloride channels. EMBO J 1995, 14:68-75.
62. Abraham EH, Pret AG, Gerweck L, Seneveretne T, Arceci RJ, Kramer R, Guidotti G, Cantiello HF: The muItidrug resistance (mdrl) gene product functions as an ATP channel. Proc Nat/ Aced Sci USA 1993, 90:312-316.
63. Tominaga M, Torninaga T, Miwa A, Okada Y: Volume-sensitive chloride channel activity does not depend on endogenous P-glycoprotein. J Bio/Chem 1995, 270:27887-27893.
64. Jackson PS, Strange K: Single-channel properties of a volume- • sensitive anion conductance. Current activation occurs by
abrupt switching of closed channels to an open state. J Gen Physio/1995, 105:643-660.
This detailed study resolves a controversy regarding the single-channel con- ductance of volume-sensitive anion channels. Because open probability does not increase in a graded fashion, stationary noise analysis incorrectly sug- gests a small single-channel conductance, whereas single-channel data and non-stationary noise analysis consistently reveal a 15 pS to 50 i0S channel (at 0 mV and +120 mV, respectively). Once 'activated', channel open probability is 1. It is speculated that this 'activation' may be attributable to insertion of porin-like molecules into the plasma membrane, in a model similar to that for plci n [70]. Together with its companion paper [68], this is a thorough biophysical analysis of swelling-activated CI- channels, which probably also allow the flow of organic osmolytes [73].
65. Solc CK, Wine J J: Swelling-induced and depolarization-induced Cl- channels In normal and cystic fibrosis epithelial cell. Am J Physiol 1991,261 :C658-C674.
66. Lewis RS, Ross PE, Cahalan MD: Chloride channels activated by osmotic stress in T lymphocytes. J Gen Physiol 1993, 101:801-803.
67. Nilius B, Oike M, Zahradnik I, Droogmans G: Activation of a CI- current by hypotonic volume Increase in human endothelial cells. J Gen Physiol 1994, 103:787-805.
68. Jackson PS, Strange K: Characterization of the voltage- dependent properties of a volume-sensitive anion conductance. J Gen Physiol 1995, 105:661-677.
69. Ackerman M J, Wickman KD, Clapham DE: Hypotonlcity activates a native chloride currents in Xenopus oocytes. J Gen Physiol 1994, 103:153-179.
70. Gschwentner M, Nagl UO, W~II E, Schmarda A, Ritter M, Paulmichl M: Antlsense ollgonucleotldes suppress cell-volume-
induced activation of chloride channels. Pflugers Arch 1995, 430:464-470.
71. Paulmichl M, Li Y, Wickman K, Ackerman M, Peralta E, Clapham DE: New mammalian chloride channel identified by expression cloning. Nature 1992, 356:238-241.
72. Krapivinsky GB, Ackerman M J, Gordon F_A, Krapivinsky LD, Clapham DE: Molecular characterization of a swelling-induced chloride conductance regulatory protein, plcl n. Cell 1 g94, 76:439-448.
73. Strange K, Jackson PS: Swelling-activated organic osmolyte efflux: a new role for anion channels. Kidney Int 1995, 48:994-1003.
74. Roy G: Amino acid current through anion channels in cultured human glial cells. J Membr Bio11995, 147:35-44.
75. Fi6vet B, Gabillat N, Borgese F, Motels R: Expression of band 3 anion exchanger induces chloride current and taurine transport: structure-function analysis. EMBO J 1995, 14:5158-5169.
76. Fairman WA, Vandenberg RJ, Arriza JL, Kavanaugh MP, Amara SG: An excitatory amino-acid transporter with properties of e ligand-gated chloride channel. Nature 1995, 375:599-603.
77. Wadiche JI, Amara SG, Kavanaugh MP: Ion fluxes associated with excitatory amino acid transport. Neuron 1 g95, 15:721-728.
78. Busch AE, Schuster A, Waldegger S, Wagner CA, Zempel G, Broer S, Biber J, Murer H, Lang F: Expression of s renal type I sodium/phosphate-transporter (NaPI-1) induces a conductance in Xenopus oocytes permeable for organic and inorganic anions. Proc Natl Acad Sci USA 1 g96, in press.
79. Tzounopoulos T, Maylie J, Adeiman JP: Induction of endogenous • channels by high levels of heterologous membrane proteins in
Xenopus oocytes. Biophys J 1995, 69:904-908. This paper shows that overexpression of many membrane proteins, including those that do not function as channels, induces slowly hyperpolarization- activated currents in Xenopus oocytes. Thus, these currents, which were thought to be specific for phospholemman [81], are attributable to an acti- vation of an endogenous Xenopua oocyte channel. Similar observations have been made for non-functional CLC proteins [2,20]. The experiments strongly suggests that hyperpolarization opens a non-specific cation channel, and that the inflow of Ca 2+ activates a Ca2+-dependent CI- channel. Thus, this paper investigates one of the pitfalls encountered when studying functional expression of CI- channels.
80. Shimbo K, Brassard DL, Lamb RA, Pinto LH: Viral and cellular small integral membrane proteins can modify ion channel endogenous to Xenopus oocytes. Biophys J 1995, 69:1819-1829.
81. Moorman JR, Palmer C J, John JE, Durieux ME, Jones LR: Phospholemman expression induces a hyperpolsrlzatlon- activated chloride current in Xenopus oocytes. J Biol Chem 1992, 267:14551-14554.
82. Higgins CF: The ABC of channel regulation. Cell 1995, 82:693-696.