Embed Size (px)
Citation preview

ARTICLE IN PRESS
Int. J. Hyg. Environ.-Health 208 (2005) 211–218
1438-4639/$ - se
doi:10.1016/j.ijh
�Correspond
E-mail addr
www.elsevier.de/ijheh
Chemopreventive properties of trans - resveratrol against the cytotoxicityof chloroacetanilide herbicides in vitro
Zsuzsanna Kocsis, Zoltan L. Marcsek�, Matyas G. Jakab, Bela Szende, Anna Tompa
National Institute of Chemical Safety, ‘‘Jozsef Fodor’’ National Center for Public Health, P.O. Box 36,
Nagyvarad ter 2, H-1450 Budapest, Hungary
Received 18 May 2004; received in revised form 27 December 2004; accepted 12 January 2005
Abstract
The beneficial effect of trans-resveratrol (RESV) on health is well documented. Our aim was to study the putativepreventive effect of RESV on the cytotoxicity of frequently used herbicides (alachlor, acetochlor). Estrogen receptorpositive (ER+) MCF-7 human mammary carcinoma, HepG2 (ER+) human hepatocellular carcinoma and VEROestrogen receptor negative (ER�) non-transformed monkey fibroblast cell lines were treated with alachlor andacetochlor (2–500 mg/ml) as toxic agents, and RESV (10 mM) as preventive agent. The MTT dye reduction assay wasperformed to test cytotoxicity, and flow cytometry to test cell proliferation and apoptosis.
RESV is not cytotoxic in the concentration range of 1–100 mM on neither cell lines examined after 24 h, but cytotoxicon Vero and MCF-7 cells at 100 mM after 48 h, and on all three cell lines after 72 h. On both ER+ cell lines astimulation of viability occurs in the low concentration range (0.5–12.5 mM) as detected by the MTT assay. Cell cycleanalysis of the culture shows a significant increase of S-phase cells at low concentrations of RESV (10–50 mM) and adecrease in the 100–200 mM concentration range. The ratio of apoptotic cells significantly increases after theadministration of 50 mM RESV, depending on the incubation time.
The cytotoxicity of 20–65 mg/ml alachlor and 10–65 mg/ml acetochlor was significantly decreased by the addition of10 mM RESV in Vero ER� cells whereas no significant change was detected on ER+ cell lines MCF-7 and HepG2.
These results show that RESV protects non-transformed ER� cells, but has no such effect on ER+ tumor cells.r 2005 Elsevier GmbH. All rights reserved.
Keywords: Trans-resveratrol; Cytotoxicity; Vero; HepG2; MCF-7; Alachlor; Acetochlor
Introduction
The chloroacetanilide herbicides acetochlor (2-methyl-ethyl-N-(etoxymethyl)-chloroacetanilide) andalachlor (2,6-diethyl-N-(metoxymethyl)-chloroacetani-lide) have similar toxicological properties (Osano etal., 2002), and one of their most significant is the
e front matter r 2005 Elsevier GmbH. All rights reserved.
eh.2005.01.021
ing author. Tel.: +361 476 1397; fax: +36 1 476 1227.
ess: [email protected] (Z.L. Marcsek).
induction of nasal adenomas in rats in 2-year feedingstudies (Wetmore et al., 1999). The proposed mode ofaction of alachlor and acetochlor is their metabolicconversion to quinone-imine through a sulfoxide pro-duct (Green et al., 2000), the formation of proteinadducts (Lambert et al., 1999), generation of reactiveoxygen radicals and modification of cell death (Bagchi etal., 1995), induction of sister chromatide exchange (Hillet al., 1997) and compensatory hyperplasia leading toadenomas (Genter et al., 2000). Referring to the

ARTICLE IN PRESSZ. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218212
apoptosis-inducing activity of alachlor (Burow et al.,1999), there are three reliable parameters to evaluate theprotecting effect of any possible antioxidants: theapoptotic index, the ratio of cells in the S-phase of thecell cycle, and the viability (MTT assay) of culturedcells.
Trans-resveratrol (3,40,5-trihydroxy-trans-stilbene,RESV) is a naturally occurring phytoalexin in the fruitsand leaves of several plants (grapes, peanuts, etc.). It is astilbene derivative and has a similar chemical structureas the synthetic estrogen diethylstilbestrol. One of itsbiological activities is a phytoestrogen-like effect (Lang-cake and Pryce, 1976; Gehm et al., 1997; Bowers et al.,2000).
RESV was identified in 1963 from dried roots ofPolygonum cuspidatum, a Japanese remedy called Ko-jo-kon (Nonomura et al., 1963). In the traditionalmedication in the ancient Chinese and Japanese culturesKo-jo-kon was used in the therapy of dermatitis,gonorrhea, hyperlipidemy. RESV was detected in grapes(Vitis vinifera) in 1976 and in wine in 1992. RESV issynthesized in the epidermal layer of the leaves andgrapes where its concentration is 50–100 mg/kg (Siemannand Creasy, 1992). The amount of RESV is lower inwhite wine than in red wine, where the grape skin is notseparated from the must. The RESV content of winesvaries – depending on the breed – in the range of1–35 mM (Goldberg et al., 1995). Its synthesis isenhanced after mold infections and by UV irradiation(Langcake and Pryce, 1976; Hain et al., 1993).
The in vitro and in vivo biological activity of RESVhas been examined in a great variety of test systems. Itsbeneficial properties on humans is the positive effect onthe circulatory system, realized in the blood vessels aswell as preventing the development of plaques andsclerotic areas (Nakagawa et al., 2001; Gusman et al.,2001).
RESV also inhibits cell proliferation in a dose-dependent manner. In connection with decreasedDNA synthesis, Fontecave et al. could show theinhibition of ribonucleotide reductase (1998). It alsohas inhibitory activities on the synthesis of prostaglan-din as well as the enzyme 5-lipoxygenase. The anti-proliferative properties of RESV corresponds well to itsability to inhibit the general cyclin-dependent kinasescausing a p21WAF1-mediated cell cycle delay (Ahmadet al., 2001; Kim et al., 2003). RESV is an estrogen-agonist, a phytoestrogen which inhibits the growth ofER+ human mammary carcinoma cells (Ashby et al.,1999; Pozo-Guisado et al., 2002).
The aim of this work was to study the putativeprotecting effect of RESV against the cytotoxicity ofalachlor and acetochlor in ER� and ER+ cell cultures,using the MTT assay to determine cytotoxic effects ongrowth and viability, and flow cytometry to calculate theratio of apoptotic cells and S-phase cells.
Materials and methods
Chemicals
Trans-resveratrol, and the MTT reagent were pur-chased from Sigma (St. Louis, MS, USA), alachlor andacetochlor from Fluka (Fluka Holding AG, Buchs,Switzerland).
Cell cultures
Two estrogen receptor positive (ER+) cell lines(MCF-7 (ECACC 86012803) human breast adenocarci-noma and HepG2 (ATCC HB-8065) hepatocellularcarcinoma) as well as an estrogen receptor negative(ER�) cell line (Vero (ATCC CCL-81) normal kidneyepithelial cells from African green monkey) were used.ER+ cell lines were chosen because of the possiblebiological effect of the phytoestrogen RESV. ER� Verocells, which are recommended for cytotoxicity studies,may react in a different way to RESV. Cells were storedin liquid nitrogen and cultured in DMEM withoutphenol red (GIBCO, Meckenheim, Germany), usingplastic culture dishes or microwell plates and thermanoxcoverslips (Nunc A/S, Roskilde, Denmark). DMEMmedium was supplemented with 10% (v/v) heat-inactivated fetal calf serum (FCS) (GIBCO InvitrogenLife Technologies, Paisley, Scotland), 2mM L-glutamin,and antibiotics: 100 units/ml penicillin and 100 mg/mlstreptomycin (Sigma, St. Louis, USA). Cells growing asa monolayer were kept at 37 1C in a humidifiedthermostat in air containing 5% CO2. For the study ofapoptosis, necrosis, and cell cycle the cells were seededinto Petri dishes with a density of 5� 105 cells, for theMTT assay 96-well microplates were used with 3� 103
cells per well.
Treatment
Cytotoxicity testing of RESV (at 24, 48 and 72 h) wasperformed in the concentration range of 0.1 and 100 mMusing all cell lines. Control cultures were maintained inDMEM similar to the treated cultures.
Four parallel samples of cells were plated and after24 h the cultures treated with 100 ml of RESV in varyingconcentrations in 96-well tissue culture plates. After 24,48 and 72 h apoptosis and cell cycle studies and theMTT assay were done. Since 10 mM of RESV proved tobe the highest non-toxic dose, this concentration wasused in our experiments. According to our pilotcytotoxicity study the dose range of alachlor was 1.95and 500 mg/ml (HepG2 and MCF-7), or 7.8–500 mg/ml(Vero). The dose range of acetochlor was between 1.95and 500 mg/ml for all three cell lines. Single treatment byvarious doses of alachlor or acetochlor (Figs. 4 and 5)

ARTICLE IN PRESSZ. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218 213
together with 10 mM RESV was performed 24 h afterplating on four parallel samples. The MTT assay wasdone 24 h after treatment, referring to results of ourprevious study.
MTT assay
Cytotoxic effects on the growth and viability of cellswere determined using the tetrazolium dye (MTT; 3[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)assay as described by Horiuchi et al. (1988). A total of3� 103 cells/ml were plated in 96-microwell plates. MTTsolution was prepared at 5mg/ml in PBS, filter-sterilizedand stored in the dark at 4 1C for a maximum of 1month. To 100 ml of culture 20 ml of MTT reagent wasadded. After incubation for 3 h at 37 1C the water-insoluble formazan dye formed was solubilized byaddition of 100 ml propanol to the culture wells. Theplates were further incubated for 20min at roomtemperature, and optical density (OD) of the wells weredetermined using an Anthos 2020 (Salzburg, Austria)ELISA microplate reader at a test wavelength of 570 nmand a reference wavelength of 690 nm. Each set includedblank background control wells containing an appro-priate volume of media but no cells. All experimentswere performed at least three times, with four wells foreach concentration of RESV. The control cells weregrown under the same conditions without the additionof RESVl. Cell survival (percent of control) wascalculated relative to untreated control cells.
Flow cytometry
MCF-7 cells were cultured for 24 h in 10 cm Petridishes at an initial plating density of 5� 105 cells/dish inDMEM containing 10% FCS and antibiotics. Then thecells were treated as described above for 24 and 48 h.Pulse labeling of cells with 5 mg/ml 5-bromo-20-deoxyur-idine (BrdU, Sigma) was performed 2 h prior to thetermination of the cultures. After trypsinization cellswere washed twice with PBS, fixed in 1ml of ice-cold70% ethanol and stored at �20 1C until furtherprocessing. For the determination of the cell cyclephases, DNA was stained with propidium iodide (PI)and the incorporated BrdU was detected immunocyto-chemically with FITC-labeled monoclonal antibodies(Becton Dickinson). DNA denaturation prior to PI andFITC-anti-BrdU staining was performed at roomtemperature using a modification of the methoddescribed by van Erp et al. (1988). Briefly, ethanol-fixedcells were washed twice in PBS and the resulting pelletwas resuspended in 700 ml of 2M HCl containing0.2mg/ml pepsin (Sigma), incubated for 30min andneutralized by 1ml of 0.1M Na2B4O7 (Reanal, Hun-gary). Cells were washed twice in PBS and resuspended
in 200 ml PBS containing 0.5% Tween 20 (Sigma), then20 ml FITC-conjugated anti-BrdU was added. Sampleswere incubated in the dark for 30min, diluted with500 ml PBS, centrifuged and the cellular pellet wasresuspended in 600 ml PBS, RNase A (Sigma) was addedto a final concentration of 0.1mg/ml and incubated for20min. Finally, 10 ml of 1mg/ml PI (Sigma) was added.Flow cytometry analysis was performed in a FACSCa-libur (Becton Dickinson) flow cytometer at a flow rate ofapproximately 400 cells/s. CellQuest software (BectonDickinson) was used for data analysis. The percentagesof S-phase and apoptotic fractions were determined onthe basis of PI and anti-BrdU-FITC/PI fluorescence.
Statistical analysis
Statistical analyses were performed with the Student’spaired t-test using GraphPad Prism (GraphPad Soft-ware, Inc., San Diego, CA, USA) statistical analysisprogram and po0:05 was considered to be significant.
Results
In vitro cytotoxicity of RESV
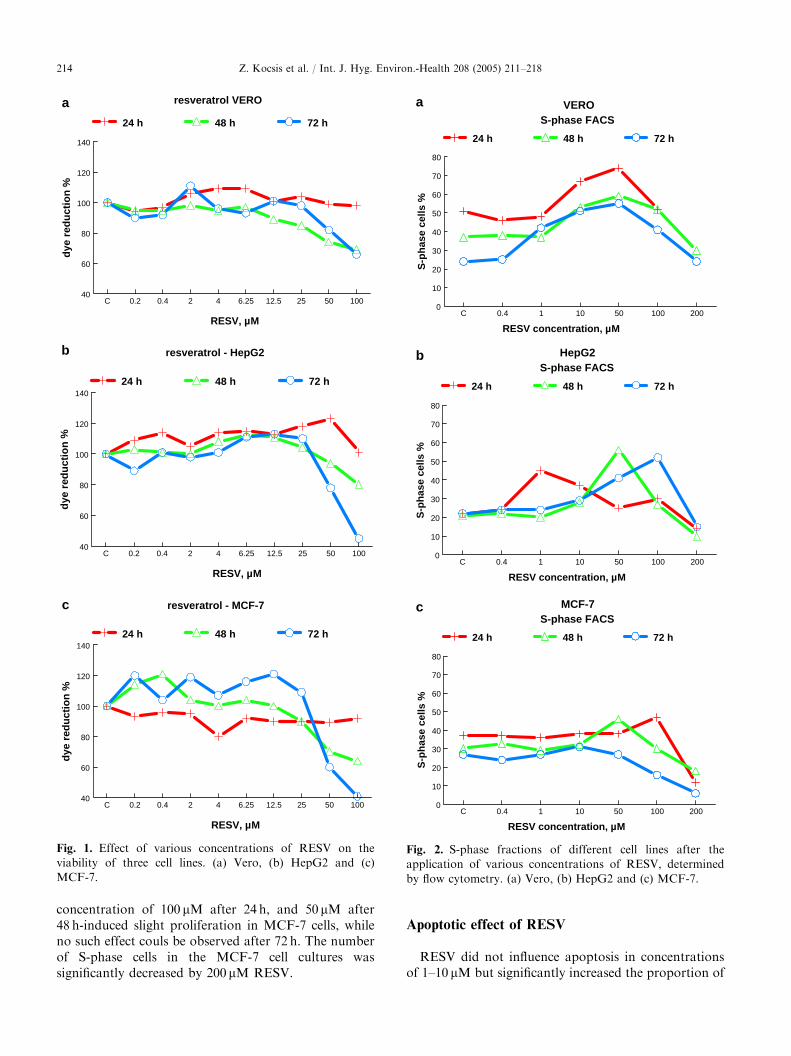
In the MTT assay, RESV did not significantlyinfluence the viability of Vero cells in the dose rangebetween 0 and 50 mM. RESV concentrations higher than50 mM proved to be cytotoxic (corresponding to 50%reduction of viability measured by the MTT assay) at 48and 72 h after RESV administration, respectively (Fig.1a). A slight but significant increase of viability wasobserved for HepG2 and MCF-7 cells, depending on thedose and the treatment period (Fig. 1b and c).Cytotoxicity became evident at RESV concentrationsof 50 mM and above at treatments longer than 48 h inboth ER+ tumor cell lines.
Effect of RESV on cell proliferation activity
S-phase analysis by flow cytometry showed that Verocells were significantly accumulated in the S-phase afterRESV administration in the dose range between 10 and100 mM, while RESV concentrations below 10 mM hadno effect on the S-phase proportion except after 72 h oftreatment, when 1 mM of RESV showed stimulation(Fig. 2a). For HepG2 cells, a proliferation enhancingeffect was observed at RESV concentrations of 1 and10 mM after 24 h (Fig. 2b). This effect could also beobserved at 50 mM after 48 h, and at 50 and 100 mM after72 h. The proportion of S-phases in HepG2 cellsdecreased significantly at 200 mM in all treatmentperiods. MCF-7 cells did not react with proliferationat RESV concentrations of 1–10 mM (Fig. 2c). RESV
ARTICLE IN PRESS
a
C 0.2 0.4 2 4 6.25 12.5 25 50 100
RESV, µM
40
60
80
100
120
140
dye
red
uct
ion
%
resveratrol VERO
24 h 48 h 72 h
b
C 0.2 0.4 2 4 6.25 12.5 25 50 100
RESV, µM
40
60
80
100
120
140
dye
red
uct
ion
%
resveratrol - HepG2
24 h 48 h 72 h
c
C 0.2 0.4 2 4 6.25 12.5 25 50 100
RESV, µM
40
60
80
100
120
140
dye
red
uct
ion
%
resveratrol - MCF-7
24 h 48 h 72 h
Fig. 1. Effect of various concentrations of RESV on the
viability of three cell lines. (a) Vero, (b) HepG2 and (c)
MCF-7.
a
C 0.4 1 10 50 100 200
RESV concentration, µM
0
10
20
30
40
50
60
70
80
S-p
has
e ce
lls %
VEROS-phase FACS
24 h 48 h 72 h
b
C 0.4 1 10 50 100 200
RESV concentration, µM
0
10
20
30
40
50
60
70
80
S-p
has
e ce
lls %
HepG2S-phase FACS
24 h 48 h 72 h
c
C 0.4 1 10 50 100 200
RESV concentration, µM
0
10
20
30
40
50
60
70
80
S-p
has
e ce
lls %
MCF-7S-phase FACS
24 h 48 h 72 h
Fig. 2. S-phase fractions of different cell lines after the
application of various concentrations of RESV, determined
by flow cytometry. (a) Vero, (b) HepG2 and (c) MCF-7.
Z. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218214
concentration of 100 mM after 24 h, and 50 mM after48 h-induced slight proliferation in MCF-7 cells, whileno such effect couls be observed after 72 h. The numberof S-phase cells in the MCF-7 cell cultures wassignificantly decreased by 200 mM RESV.
Apoptotic effect of RESV
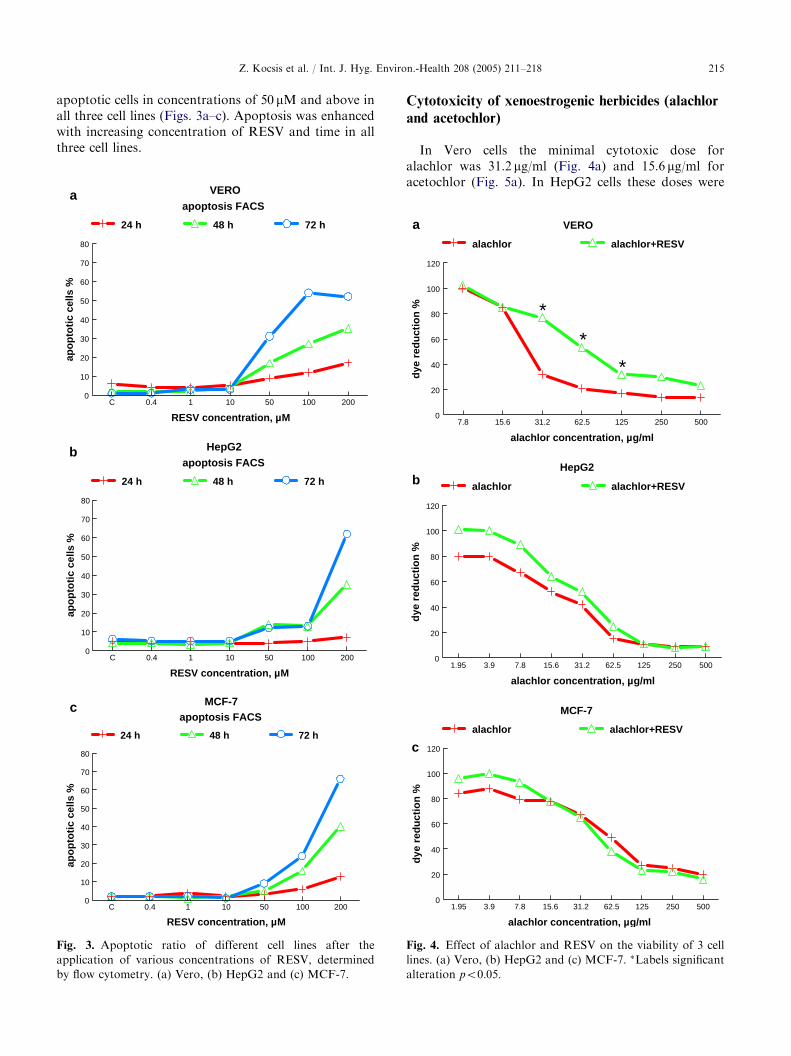
RESV did not influence apoptosis in concentrationsof 1–10 mM but significantly increased the proportion of
ARTICLE IN PRESSZ. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218 215
apoptotic cells in concentrations of 50 mM and above inall three cell lines (Figs. 3a–c). Apoptosis was enhancedwith increasing concentration of RESV and time in allthree cell lines.
a
C 0.4 1 10 50 100 200
RESV concentration, µM
0
10
20
30
40
50
60
70
80
apo
pto
tic
cells
%
VEROapoptosis FACS
24 h 48 h 72 h
b
C 0.4 1 10 50 100 200
RESV concentration, µM
0
10
20
30
40
50
60
70
80
apo
pto
tic
cells
%
HepG2apoptosis FACS
24 h 48 h 72 h
c
C 0.4 1 10 50 100 200
RESV concentration, µM
0
10
20
30
40
50
60
70
80
apo
pto
tic
cells
%
MCF-7apoptosis FACS
24 h 48 h 72 h
Fig. 3. Apoptotic ratio of different cell lines after the
application of various concentrations of RESV, determined
by flow cytometry. (a) Vero, (b) HepG2 and (c) MCF-7.
Cytotoxicity of xenoestrogenic herbicides (alachlor
and acetochlor)
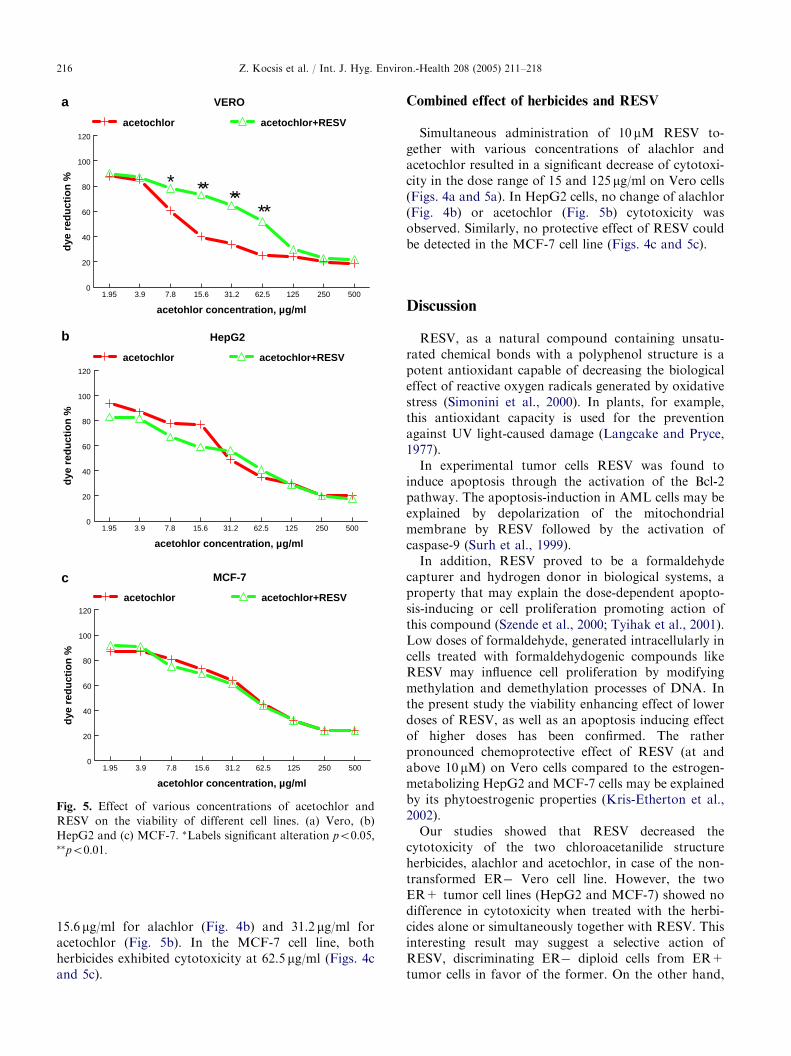
In Vero cells the minimal cytotoxic dose foralachlor was 31.2 mg/ml (Fig. 4a) and 15.6 mg/ml foracetochlor (Fig. 5a). In HepG2 cells these doses were
a
7.8 15.6 31.2 62.5 125 250 500
alachlor concentration, µg/ml
0
20
40
60
80
100
120
dye
red
uct
ion
%
VERO
alachlor alachlor+RESV
*
**
b
1.95 3.9 7.8 15.6 31.2 62.5 125 250 500
alachlor concentration, µg/ml
0
20
40
60
80
100
120
dye
red
uct
ion
%
HepG2
alachlor alachlor+RESV
c
1.95 3.9 7.8 15.6 31.2 62.5 125 250 500
alachlor concentration, µg/ml
0
20
40
60
80
100
120
dye
red
uct
ion
%
MCF-7
alachlor alachlor+RESV
Fig. 4. Effect of alachlor and RESV on the viability of 3 cell
lines. (a) Vero, (b) HepG2 and (c) MCF-7. �Labels significant
alteration po0:05:
ARTICLE IN PRESS
a
1.95 3.9 7.8 15.6 31.2 62.5 125 250 500
acetohlor concentration, µg/ml
0
20
40
60
80
100
120
dye
red
uct
ion
%
VERO
acetochlor acetochlor+RESV
* * **
* **
b
1.95 3.9 7.8 15.6 31.2 62.5 125 250 500
acetohlor concentration, µg/ml
0
20
40
60
80
100
120
dye
red
uct
ion
%
HepG2
acetochlor acetochlor+RESV
c
1.95 3.9 7.8 15.6 31.2 62.5 125 250 500
acetohlor concentration, µg/ml
0
20
40
60
80
100
120
dye
red
uct
ion
%
MCF-7
acetochlor acetochlor+RESV
Fig. 5. Effect of various concentrations of acetochlor and
RESV on the viability of different cell lines. (a) Vero, (b)
HepG2 and (c) MCF-7. �Labels significant alteration po0:05;��po0:01:
Z. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218216
15.6 mg/ml for alachlor (Fig. 4b) and 31.2 mg/ml foracetochlor (Fig. 5b). In the MCF-7 cell line, bothherbicides exhibited cytotoxicity at 62.5 mg/ml (Figs. 4cand 5c).
Combined effect of herbicides and RESV
Simultaneous administration of 10 mM RESV to-gether with various concentrations of alachlor andacetochlor resulted in a significant decrease of cytotoxi-city in the dose range of 15 and 125 mg/ml on Vero cells(Figs. 4a and 5a). In HepG2 cells, no change of alachlor(Fig. 4b) or acetochlor (Fig. 5b) cytotoxicity wasobserved. Similarly, no protective effect of RESV couldbe detected in the MCF-7 cell line (Figs. 4c and 5c).
Discussion
RESV, as a natural compound containing unsatu-rated chemical bonds with a polyphenol structure is apotent antioxidant capable of decreasing the biologicaleffect of reactive oxygen radicals generated by oxidativestress (Simonini et al., 2000). In plants, for example,this antioxidant capacity is used for the preventionagainst UV light-caused damage (Langcake and Pryce,1977).
In experimental tumor cells RESV was found toinduce apoptosis through the activation of the Bcl-2pathway. The apoptosis-induction in AML cells may beexplained by depolarization of the mitochondrialmembrane by RESV followed by the activation ofcaspase-9 (Surh et al., 1999).
In addition, RESV proved to be a formaldehydecapturer and hydrogen donor in biological systems, aproperty that may explain the dose-dependent apopto-sis-inducing or cell proliferation promoting action ofthis compound (Szende et al., 2000; Tyihak et al., 2001).Low doses of formaldehyde, generated intracellularly incells treated with formaldehydogenic compounds likeRESV may influence cell proliferation by modifyingmethylation and demethylation processes of DNA. Inthe present study the viability enhancing effect of lowerdoses of RESV, as well as an apoptosis inducing effectof higher doses has been confirmed. The ratherpronounced chemoprotective effect of RESV (at andabove 10 mM) on Vero cells compared to the estrogen-metabolizing HepG2 and MCF-7 cells may be explainedby its phytoestrogenic properties (Kris-Etherton et al.,2002).
Our studies showed that RESV decreased thecytotoxicity of the two chloroacetanilide structureherbicides, alachlor and acetochlor, in case of the non-transformed ER� Vero cell line. However, the twoER+ tumor cell lines (HepG2 and MCF-7) showed nodifference in cytotoxicity when treated with the herbi-cides alone or simultaneously together with RESV. Thisinteresting result may suggest a selective action ofRESV, discriminating ER� diploid cells from ER+tumor cells in favor of the former. On the other hand,

ARTICLE IN PRESSZ. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218 217
the selective mode of action may be based on theestrogen receptor positivity or negativity of the targetcells. RESV, as a phytoestrogen, may interfere with theestrogen receptors of the tumor cells resulting in the lackof preventing the cytotoxicity of alachlor and aceto-chlor. The mode of action of alachlor, a xenoestrogen,and acetochlor, the structure of which is similar to thatof alachlor, might be explained by the cellular metabo-lizing capability present in HepG2 and MCF-7 butmissing in Vero cells. Chloroacetamide herbicides aremetabolized by the cytochrom P450-related system(Hanioka et al., 2002).
From the practical point of view, RESV may beeffectively used as a protective or preventiveagent against alachlor or acetochlor intoxicationsin humans. Such intoxication may occur amongworkers of factories producing herbicides, among thosewho distribute these compounds in field as well assuicides in several hundreds of cases yearly (Teixeiraet al., 2004).
Acknowledgements
This work was supported in part by Grants SzechenyiNKFP-1/016/2001 and OTKA T30799. The authorswould like to thank Katalin Molnar for her excellenttechnical help.
References
Ahmad, N., Adhami, V.M., Afaq, F., Feyes, D.K., Mukhtar,
H., 2001. Resveratrol caused WAF-1/p21-mediated G1-
phase arrest of cell cycle and induction of apoptosis in
human epidermoid carcinoma A431 cells. Clin. Cancer Res.
7, 1466–1473.
Ashby, J., Tinwell, H., Haseman, J., 1999. Lack of effects for
low dose levels of bisphenol A and diethylstilbestrol on the
prostate gland of CF1 mice exposed in utero. Regul.
Toxicol. Pharmacol. 30, 156–166.
Bagchi, D., Bagchi, M., Hassoun, E.A., Stohs, S.J., 1995. In
vitro and in vivo generation of reactive oxygen species,
DNA damage and lactate dehydrogenase leakage by
selected pesticides. Toxicology 104, 129–140.
Bowers, J.L., Tyulmenkov, V.V., Jernigan, S.C., Klinge, C.M.,
2000. Resveratrol acts as a mixed agonist/antagonist for
estrogen receptors alpha and beta. Endocrinology 141,
3657–3667.
Burow, M.E., Tang, Y., Collins-Burow, B.M., Krajewski, S.,
Reed, J.C., McLachlan, J.A., Beckman, B.S., 1999. Effects
of environmental estrogens on tumor necrosis factor alpha-
mediated apoptosis in MCF-7 cells. Carcinogenesis 20,
2057–2061.
Fontecave, M., Lepoivre, M., Elleingand, E., Gerez, C.,
Guittet, O., 1998. Resveratrol, a remarkable inhibitor of
ribonucleotide reductase. FEBS Lett. 421, 277–279.
Gehm, B.D., McAndrews, J.M., Chien, P.Y., Jameson, J.L.,
1997. Resveratrol, a polyphenolic compound found in
grapes and wine, is an agonist for the estrogen receptor.
Proc. Natl. Acad. Sci. USA 94, 14138–14143.
Genter, M.B., Burman, D.M., Dingeldein, M.W., Clough, I.,
Bolon, B., 2000. Evolution of alachlor-induced nasal
neoplasms in the Long-Evans rat. Toxicol. Pathol. 28,
770–781.
Goldberg, D.M., Ng, E., Karumanchiri, A., Yan, J., Diamandis,
E.P., Soleas, G.J., 1995. Assay of resveratrol glucosides and
isomers in wine by direct-injection high-performance liquid
chromatography. J. Chromatogr. A 708, 89–98.
Green, T., Lee, R., Moore, R.B., Ashby, J., Willis, G.A.,
Lund, V.J., Clapp, M.J.L., 2000. Acetochlor-induced rat
nasal tumors: further studies on the mode of action and
relevance to humans. Regul. Toxicol. Pharmacol. 32,
127–133.
Gusman, J., Malonne, H., Atassi, G., 2001. A reappraisal of
the potential chemopreventive and chemotherapeutic prop-
erties of resveratrol. Carcinogenesis 22, 1111–1117.
Hain, R., Reif, H.J., Krause, E., Langebartels, R., Kindl, H.,
Vornam, B., Wiese, W., Schmelzer, E., Schreier, P.H.,
Stocker, R.H., Stenzel, K., 1993. Disease resistance results
from foreign phytoalexin expression in a novel plant.
Nature 361, 153–156.
Hanioka, N., Watanabe, K., Yoda, R., Ando, M., 2002. Effect
of alachlor on hepatic cytochrome P450 enzymes in rats.
Drug Chem. Toxicol. 25, 25–37.
Hill, A.B., Jefferies, P.R., Quistad, G.B., Casida, J.E., 1997.
Dialkylquinoneimine metabolites of chloroacetanilide her-
bicides induce sister chromatid exchanges in cultured
human lymphocytes. Mutat. Res./Genet. Toxicol. Environ.
Mutagen. 395, 159–171.
Horiuchi, N., Nakagava, K., Sasaki, Y., Minato, K., Fujiwara,
Y., Nezu, K., Ohe, Y., Sajio, N., 1988. In vitro antitumor
activity of mitomicyn C derivative (RM-49) and a new
anticancer antibiotic (FK 973) against lung cancer cell lines
determined by tetrazolium dye (MTT) assay. Cancer
Chemother. Pharmacol. 22, 246–250.
Kim, Y.A., Lee, W.H., Choi, T.H., Rhee, S.H., Park, K.Y.,
Choi, Y.H., 2003. Involvement of p21WAF1/CIP1, pRB,
Bax and NF-kappaB in induction of growth arrest and
apoptosis by resveratrol in human lung carcinoma A549
cells. Int. J. Oncol. 23, 1143–1149.
Kris-Etherton, P.M., Hecker, K.D., Bonanome, A., Coval,
S.M., Binkoski, A.E., Hilpert, K.F., Griel, A.E., Etherton,
T.D., 2002. Bioactive compounds in foods: their role in the
prevention of cardiovascular disease and cancer. Am. J.
Med. 113 (Suppl 9B), 71S–88S.
Lambert, G.R., Padgett, W.T., George, M.H., Kitchin, K.T.,
Nesnow, S., 1999. Quantitative analysis of alachlor protein
adducts by gas chromatography–mass spectrometry. Anal.
Biochem. 268, 289–296.
Langcake, P., Pryce, R.J., 1976. The production of resveratrol
by Vitis vinifera and other members of the Vitaceae as a
response to infection or injury. Physiol. Plant Pathol. 9,
77–86.
Langcake, P., Pryce, R.J., 1977. The production of resveratrol
and the viniferins by grapevines in response to ultraviolet
irradiation. Phytochemistry 16, 1193–1196.

ARTICLE IN PRESSZ. Kocsis et al. / Int. J. Hyg. Environ.-Health 208 (2005) 211–218218
Nakagawa, H., Kiyozuka, K., Uemura, Y., Senzaki, H.,
Shikata, N., Hioki, K., Tsubura, A., 2001. Resveratrol
inhibitis human breast cancer cell growth and may mitigate
the effect of linoleic acid, a potent breast cancer cell
stimulator. J. Cancer Res. Clin. Oncol. 127, 258–264.
Nonomura, S., Kanagawa, H., Makimoto, A., 1963. Chemical
constituents of polygonaceous plants. I. Studies on the
components of Ko-jo-kon (Polygonum cuspidatum Sieb et
Zucc). Yakugaku Zasshi 83, 983–988.
Osano, O., Admiraal, W., Klamer, H.J., Pastor, D., Bleeker,
E.A., 2002. Comparative toxic and genotoxic effects of
cloroacetanilide formamidines and their degradation pro-
ducts on Vibrio fischeri and Chironomus riparius. Environ.
Pollut. 119, 195–202.
Pozo-Guisado, E., Alvarez-Barrientos, A., Mulero-Navarro,
S., Santiago-Josefat, B., Fernandez-Salguero, P.M., 2002.
The antiproliferative activity of resveratrol results in
apoptosis in MCF-7 but not in MDA-MB-231 human
breast cancer cells: cell-specific alteration of the cell cycle.
Biochem. Pharmacol. 64, 1375–1386.
Siemann, E.H., Creasy, L.L., 1992. Concentration of
phytoalexin resveratrol in wine. Am. J. Enol. Vitic. 43,
49–52.
Simonini, G., Pignone, A., Generini, S., Falcini, F., Matucci,
C.M., 2000. Emerging potentials for an antioxidant therapy
as a new approach to the treatment of systemic sclerosis.
Toxicology 155, 1–15.
Surh, Y.-J., Hurh, Y.-J., Kang, J.-Y., Lee, E., Kong, G., Lee,
S.J., 1999. Resveratrol, an antioxidant present in red wine,
induces apoptosis in human promyelocytic leukemia (HL-
60) cells. Cancer Lett. 140, 1–10.
Szende, B., Tyihak, E., Kiraly Veghely, Z., 2000. Dose-dependent
effect of resveratrol on proliferation and apoptosis in
endothelial and tumor cell cultures. Exp. Mol. Med. 32, 88–92.
Teixeira, H., Proenca, P., Alvarenga, M., Oliveira, M.,
Marques, E.P., Vieira, D.N., 2004. Pesticide intoxications
in the Centre of Portugal: three years analysis. Forensic Sci.
Int. 143, 199–204.
Tyihak, E., Bocsi, J., Timar, F., Racz, G., Szende, B., 2001.
Formaldehyde promotes and inhibits the proliferation of
cultured tumour and endothelial cells. Cell Prolif. 34, 135–141.
van Erp, P.E., Brons, P.P., Boezeman, J.B., de Jongh, G.J.,
Bauer, F.W., 1988. A rapid flow cytometric method for
bivariate bromodeoxyuridine/DNA analysis using simulta-
neous proteolytic enzyme digestion and acid denaturation.
Cytometry 9, 627–630.
Wetmore, B.A., Mitchell, A.D., Meyer, S.A., Genter, M.B.,
1999. Evidence for site-specific bioactivation of alachlor in
the olfactory mucosa of the Long-Evans rat. Toxicol. Sci.
49, 202–212.





























