Embed Size (px)
Citation preview

Prevention and Epidemiology
Chemopreventive Activity of Plant Flavonoid Isorhamnetin inColorectal Cancer Is Mediated by Oncogenic Src andb-Catenin
Shakir M. Saud1,2, Matthew R. Young2, Yava L. Jones-Hall6, Lilia Ileva3, Moses O. Evbuomwan5,Jennifer Wise4, Nancy H. Colburn2, Young S. Kim1, and Gerd Bobe7,8
AbstractAnalysis of the Polyp Prevention Trial showed an association between an isorhamnetin-rich diet and a reduced
risk of advanced adenoma recurrence; however, the mechanism behind the chemoprotective effects ofisorhamnetin remains unclear. Here, we show that isorhamnetin prevents colorectal tumorigenesis of FVB/Nmice treated with the chemical carcinogen azoxymethane and subsequently exposed to colonic irritant dextransodium sulfate (DSS). Dietary isorhamnetin decreased mortality, tumor number, and tumor burden by 62%,35%, and 59%, respectively. MRI, histopathology, and immunohistochemical analysis revealed that dietaryisorhamnetin resolved the DSS-induced inflammatory response faster than the control diet. Isorhamnetininhibited AOM/DSS–induced oncogenic c-Src activation and b-catenin nuclear translocation, while promotingthe expression of C-terminal Src kinase (CSK), a negative regulator of Src family of tyrosine kinases. Similarly, inHT-29 colon cancer cells, isorhamnetin inhibited oncogenic Src activity and b-catenin nuclear translocation byinducing expression of csk, as verified by RNA interference knockdown of csk. Our observations suggest thechemoprotective effects of isorhamnetin in colon cancer are linked to its anti-inflammatory activities and itsinhibition of oncogenic Src activity and consequential loss of nuclear b-catenin, activities that are dependent onCSK expression. Cancer Res; 73(17); 5473–84. �2013 AACR.
IntroductionColorectal cancer is the fourth most commonly diagnosed
type of cancer in the United States, and it is the second leadingcause of cancer-related deaths (1). Chronic inflammation, suchas ulcerative colitis and Crohn disease, are associated withincreased risk of colorectal cancer (2–5).The Src family of tyrosine kinases (SFK) are nonreceptor
protein tyrosine kinases that are activated in multiple can-cers, including colorectal cancer (6). Increased Src activity inprimary colorectal cancer is an indicator of poor prognosis
(7). C-terminal Src kinase, also known as c-Src kinase (CSK),negatively regulates SFK by phosphorylation of a C-terminaltyrosine (Y530 in c-Src; ref. 8). C-terminal phosphorylation ofSFK stabilizes the protein in an inhibitory confirmation thatprevents autophosphorylation of the activation loop tyrosine(Y419 in c-Src). The receptor-like tyrosine phosphatase CD45and similar phosphatases impose a reciprocal regulation ofSrc by removing the C-terminal tyrosine phosphate (9).
Activated Src can regulate many downstream pathwaysincluding phosphoinositide 3-kinase (PI3K), Ras–Mek–extra-cellular signal–regulated kinase (ERK), STAT3, and p130 toincrease survival, proliferation, angiogenesis, motility, andinvasion (6). Activated Src can also phosphorylate b-catenin,causing its release from E-cadherin sequestration at the plas-ma membrane and enhancing its nuclear localization (10).Colorectal cancer seems to be sensitive to loss of Src regulation.Both the loss of expression of the negative regulator CSK andthe overexpression of Src have been implicated in colorectalcarcinogenesis (8).
The cost of treating colorectal cancer is estimated to be $6.5billion per year (11). Dietary change and use of dietary supple-ments, both feasible and safe, represent a viable and importantstrategy for preventing colorectal cancer (12). Recent analysisof the Polyp Prevention Trial (PPT), a clinical trial that inves-tigated the role of diet modulation in the prevention ofcolorectal adenoma recurrence, suggested that consuming anisorhamnetin-rich diet was associated with a decreased risk ofadvanced adenoma recurrence (13).
Authors' Affiliations: 1Nutritional Science Research Group, Division ofCancer Prevention, National Cancer Institute, Rockville; 2Laboratory ofCancer Prevention, Center for Cancer Research, National Cancer Institute;3Small Animal Imaging Program, 4Laboratory of Animal Science, SAIC-Frederick, Frederick; 5Laboratory of Pathology, Center for CancerResearch, National Cancer Institute, Bethesda, Maryland; 6Purdue Uni-versity College of Veterinary Medicine, Comparative Pathobiology Depart-ment,West Lafayette, Indiana; and 7Linus Pauling Institute; 8Department ofAnimal and Rangeland Sciences, Oregon State University, Corvallis,Oregon
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
S.M. Saud and M.R. Young share first authorship of this article.
Corresponding Author: Matthew R. Young, National Cancer Institute,Bldg576Rm. 100D, Frederick,MD21702.Phone: 301-846-6448; Fax: 301-846-6907; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-13-0525
�2013 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 5473
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525
Recently, it was shown that isorhamnetin can suppress skincancer by binding to and inhibiting MAP (mitogen-activatedprotein)/ERK kinase (MEK) 1 and PI3K (14). To further inves-tigate the potential role of flavonols in colorectal cancerprevention, we assessed the effects of four flavonols commonlyconsumed by humans, isorhamnetin, quercetin, rutin, andmyricetin, in a mouse model for colorectal cancer. We foundthat supplementing the diet with isorhamnetin significantlyreduced colorectal tumorigenesis in these mice. The isorham-netin diet seemed to resolve the dextran sodium sulfate (DSS)–induced inflammation as measured by MRI, histopathology,and immunohistochemical analysis (IHC). Isorhamnetin alsoinhibited Src activity and nuclear localization of b-catenin andincreased expression of CSK both in vivo and in colorectalcarcinoma cells. Finally, we showed that inhibition of Src andnuclear b-catenin was dependent on the upregulation of CSKin colon cancer cells and associated with CSK expression inisorhamnetin-fed mice.
Materials and MethodsAnimal studies
All mouse experiments were agreed to and regulated by theAnimal Care and Use Committee of the National CancerInstitute (Frederick, MD). Pathogen-free male FVB/N micewere purchased from the NCI-Frederick Animal ProductionArea at 5 weeks of age and were exposed to a 12:12–hour light/dark cycle. Mice were given an AIN-96G–purified diet fromHarlan Teklad and drinking water ad libitum. At 6 weeks of age(day 0; Fig. 1), mice were injected intraperitoneally withazoxymethane (Sigma) at a dose of 10 mg/kg body weight in0.1 mL of saline (Fig. 1). Seven days later, mice were treatedwith 2% DSS, 36,000 to 50,000 kDa (MP Biomedicals LL),dissolved in normal drinking water (reverse osmosis–purifiedwater) for 7 days and then switched back to normal drinkingwater. Body weights were measured during the DSS treatment.Three days after DSS treatment ended, the mice were evenly
sorted by change in body weight into five diet groups to ensurethat effects of the DSS were evenly represented in each of thedifferent diets. Mice were single caged and started on theindicated diet (day 17; Supplementary Table S1). Isorhamnetinand myricetin were obtained from Jinan Haohua Industry Co.,Ltd. and quercetin and rutin were obtained from Sigma-Aldrich Co., LLC. Purified diets were prepared by HarlanLaboratories. Mice were allowed to eat ad libitum. One daybefore euthanasia (days 20, 25, and 33), 12 mice per group andtime point were imaged by MRI as described previously (15).Mice were euthanized at indicated times and colon tissue wascollected.
HistopathologyColonic tissue was harvested from mice at various times
after azoxymethane or AOM/DSS treatment, fixed in 4% form-aldehyde overnight, and stored in 70% ethanol. Fixed sectionsof colonic tissues were embedded in paraffin, cut into 5-mmsections, and put onto microscopic slides by Histoserv Inc.Slides were either stained with hematoxylin and eosin (H&E)for histologic analysis via light microscopy or deparaffinized inxylene and rehydrated in graded ethanol for IHC. For histo-pathology analysis, a section of approximately 5 mm in lengthof the distal colon was evaluated for each animal. The tissueswere blinded and scored by the pathologist semiquantitativelyfrom 0 to 15 (Supplementary Table S2). Colons were evaluatedbeginning at the colorectal junction and proceeding adorally 5mm to the middle colon. Briefly, the severity of the leukocyticinfiltrate in the mucosa was subjectively assessed as mild,moderate, or severe (1, 2, and 3, respectively); the distributionwas evaluated and denoted as focal/locally extensive, multi-focal, or diffuse (1, 2, and 3, respectively); and the depth wasevaluated and denoted as mucosa only, extends to submucosa,or extends to muscularis mucosa or serosa (1, 2, and 3,respectively). The distribution of atypical glandular hyperpla-sia (AGH)/squamous (sq) metaplasia was assessed as focal,multifocal, or diffuse (1, 2, and 3, respectively). The distribution
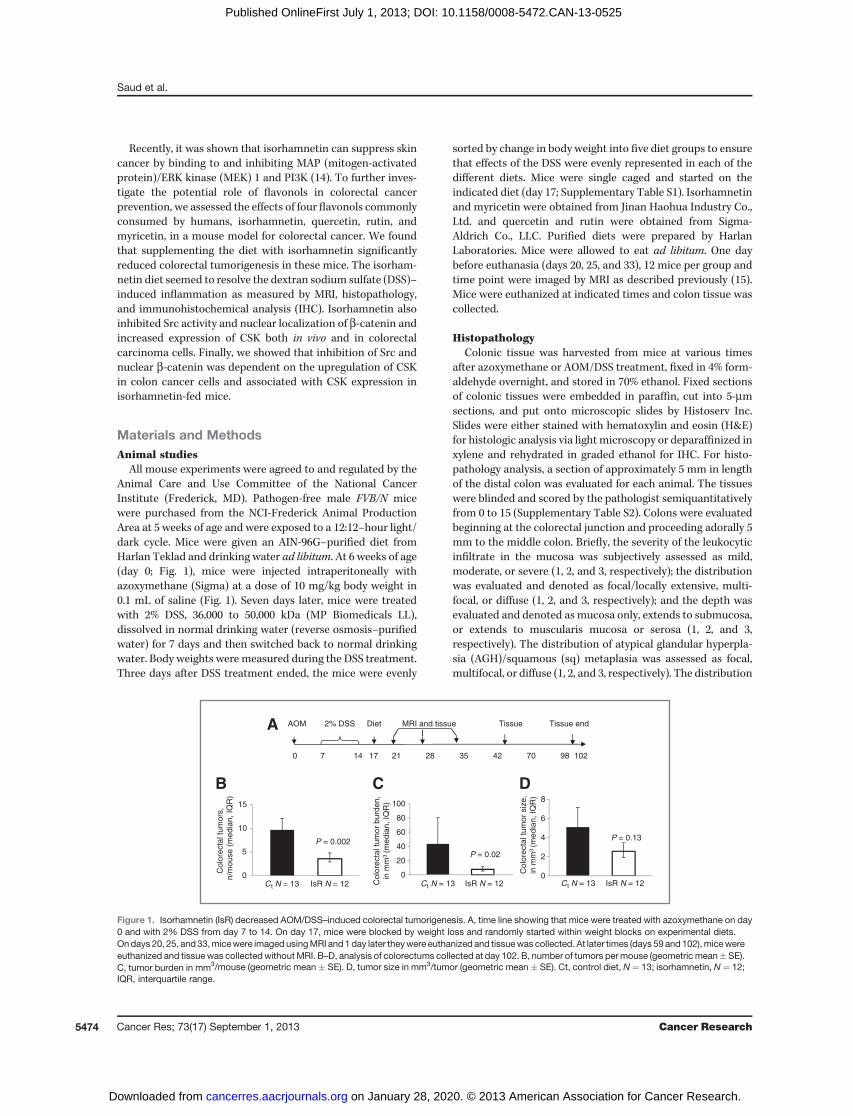
AOM 2% DSS Diet MRI and tissue Tissue endTissue
0 7 14 17 21 28 35 42 70 98 102
A
0
5
10
15
P = 0.002
Colo
recta
l tu
mors
,n/m
ouse (
media
n, IQ
R)
0
20
40
60
80
100
P = 0.02
Colo
recta
l tu
mor
burd
en,
in m
m3
(media
n, IQ
R)
Ct N = 13 IsR N = 12 Ct N = 13 IsR N = 120
2
4
6
8
Colo
recta
l tu
mor
siz
e,
in m
m3
(media
n, IQ
R)
P = 0.13
Ct N = 13 IsR N = 12
B C D
Figure 1. Isorhamnetin (IsR) decreased AOM/DSS–induced colorectal tumorigenesis. A, time line showing that mice were treated with azoxymethane on day0 and with 2% DSS from day 7 to 14. On day 17, mice were blocked by weight loss and randomly started within weight blocks on experimental diets.Ondays 20, 25, and33,micewere imaged usingMRI and1day later theywere euthanized and tissuewas collected. At later times (days 59 and102),micewereeuthanized and tissuewas collectedwithoutMRI. B–D, analysis of colorectums collected at day 102. B, number of tumors permouse (geometric mean�SE).C, tumor burden in mm3/mouse (geometric mean � SE). D, tumor size in mm3/tumor (geometric mean � SE). Ct, control diet, N¼ 13; isorhamnetin, N¼ 12;IQR, interquartile range.
Saud et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5474
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525

of ulceration was assessed as focal, multifocal, or diffuse (1, 2,and 3, respectively). If necrosis was present it was subjectivelyassessed as mild, moderate, or severe and scored accordingly(1, 2, and 3, respectively). A score of 0 was assigned for eachcriterion not represented in the section, which was the case forthe majority of mice for ulceration and necrosis. Total diseasescore per mouse was calculated by summation of the sixparameters for each mouse.
ImmunohistochemistryFor IHC, the sections were placed in low-pH–modified
citrate buffer (Dako) and subjected to antigen retrieval viapressure chamber. The sections were preincubated in 3%H2O2
for 20 minutes and blocked in normal goat serum for 1 hour.Sections were incubated overnight in primary antibody solu-tion diluted as follows: Ki-67, phospho-b-catenin654 (AbcamLaboratories) 1:50, CD45 (BD Biosciences) 1:50, pSrcY417 andb-catenin (Cell Signaling Technology) 1:100 and 1:1,000, res-pectively. Slides were incubated for 1 hour in a biotin-conju-gated secondary antibody diluted 1:200. The sections weretreated with a Vectorstain ABC kit (Vector Laboratories); theresulting activity was detected with diaminobenzidine (Sigma)and counterstained in hematoxylin. For CSK (Abcam Labora-tories), tissues were incubated 1:50 for 1 hour and detectedwith a Ventana iView DAB detection kit (Ventana MedicalSystems) on a Ventana BenchMark.
Cell cultureHuman colon cancer HT29 cell line was obtained from
American Type Culture Collection and cultured as describedpreviously (16).
Western blot analysisHT29 cells were seeded on 100-mm plates and treated with
either dimethyl sulfoxide (DMSO) vehicle (<1%) or isorham-netin (10, 20, and 40 mmol/L) for 24 hours. The media wasremoved and cells were washed with cold PBS. Cells were lysedwith complete radioimmunoprecipitation assay (RIPA) lysisbuffer (Santa Cruz Biotechnology). Protein concentration wascalculated using standard bovine serum albumin (BSA) curve.Approximately, 30 to 80mg of proteinwas loaded and separatedby SDS-PAGE, transferred to nitrocellulose membrane, andprobed with the following antibodies at indicated dilutions:CSK (1:1,000), pSRC417 (1:1,000), pSRC529 (1:1,000), b-catenin(1:2,000), pAKT (1:1,000), pERK (1:2,000), E-cadherin (1:1,000),pGSK3ab (1:1,000; Cell Signaling Technology), and pb-cate-nin654 (Abcam Laboratories) overnight at 4�C, followed byincubation in horseradish peroxidase–labeled secondary anti-bodies for 1 hour. Cells were developed with SuperSignal WestFemto Chemiluminescent Substrate (Thermo Scientific). Eachmembrane was probed with b-actin (1:5,000; Sigma) to ensureconsistent protein loading.
Real-time PCRTotal RNA from HT29 cultured cells was isolated using
TRIzol RNA extraction method. Briefly, chloroform wasadded to the sample, and after shaking and centrifugationthe aqueous layer was isolated. Isopropanol was next added
to the sample to precipitate the RNA. Following centrifu-gation, the RNA pellet was washed with 70% ethanol. RNAwas further purified using an RNeasy kit (Qiagen). ThecDNA was synthesized using Bio-Rad iScript reverse tran-scriptase, and the PCR reactions were carried out using Bio-Rad SYBR Green Master Mix carried out in Bio-Rad iCycler(Bio-Rad Laboratories). The human csk splicing variant 1and 2 were purchased from IDT and sequence is as follows:sense, 50-TCCGGCCCCGTCTCTCTTGG and antisense, 50-ACCCTCACGGGCAGGACAGG. The PCR included nontem-plate control and glyceraldehyde-3-phosphate dehydroge-nase (gapdh) as control primer set. The csk transcriptactivity level was calculated using 2�DDCT , where DCT ¼CT csk � CT gapdh and D (DCT stimulated � DCT control).
CSK siRNA constructHT29 cells were stably transfected with three unique 27mer
siRNA duplexes—2 nmol each against csk siRNA (Trilencer-27;Origene Technologies) or a scrambled control siRNA using themanufacturer's transfection reagent and recommendations.After transfection, cells were incubated for 36 hours. The totalprotein was isolated and subjected to Western blot analysis.
Soft agar assayHT29 cells were seeded onto 100-mm plates and pretreated
with either DMSO vehicle (<1%) or isorhamnetin (10, 20, and 40mmol/L) for 24 hours. Cells were trypsinized and seeded at30,000 cells in 2�Dulbecco'sModified EagleMedium (DMEM).Cell suspension was added 1:1 with 0.5% agarose (2 mL/well ina 6-well plate) and constitutes the top layer. The bottom layerconsisted of 2mL of 1.2% agarose. The cells weremaintained inan incubator for 14 days and the colonies were scanned andcounted with GelCount (Oxford Optronix Ltd.).
Statistical analysesStatistical analyses were done using SAS, version 9.2 (SAS,
Inc.) software. A Student unpaired t test was used to comparetreatment group averages for histopathology and cell culturedata. Generalized linear models were used to compare treat-ment group averages for mouse morbidity (binomial; PROCGLIMMIX) and tumor data (PROC GLM). In addition to diet,the number of DSS rounds was also included in the linearmodels. To achieve normality, tumor number was natural log–transformed and tumor burden and size were twice naturallog–transformed, after adding a 1 to prevent values below zero.Histopathology and cell culture data are shown as raw meanand their SEs; tumor data are shown as least-squaresmean andtheir SEs, which were back transformed to their original scale.All statistical tests were two-sided. The P values were notadjusted for multiple comparisons. Significance was declaredat P � 0.05 and a tendency at 0.05 to 0.10.
ResultsIsorhamnetin prevents colon tumorigenesis in AOM/DSS–treated mice
We previously showed that a high intake of flavonol-enriched foods and beverages, specifically isorhamnetin,kaempferol, and quercetin, was associated with decreased risk
Isorhamnetin-Induced CSK Inhibits Colorectal Cancer
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5475
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525
of advanced adenoma recurrence (13, 17). To verify the che-moprotective effects of individual flavonols on colorectaltumorigenesis, we fed FVB/N male mice previously treatedwith azoxymethane and DSS, diets enriched with four primaryflavonols consumed by humans (Table 1 and SupplementaryTable S1). FVB/N mice are sensitive to the AOM/DSS and onlyrequire one cycle of DSS to promote tumorigenesis. Three daysafter the DSS treatment, mice were started on an AIN93-Gcontrol diet (18) or one supplemented with the indicatedflavonol. Sensitive strains of mice treated with azoxymethaneand 1 week of DSS will develop colon dysplastic lesions,adenomas, and adenocarcinomas; unfortunately some of thesemice will have to be euthanized prematurely due to the largetumor burden and colorectal prolapse (15, 19). In our study,feeding mice isorhamnetin after the DSS exposure produced asignificantly higher survival rate (80%) comparedwith themicefed the control diet (48%, P¼ 0.02; Table 1). The isorhamnetin-fedmice also developed 35% (P¼ 0.03) fewer tumors and a 59%(P¼ 0.04) smaller tumor burden thanmice fed the control diet(Table 1). Because the decreased mortality and tumor numberin the quercetin interventionwas not significant andwe sawnobeneficial effects with rutin or myricetin diets, we conductedan in-depth study of the isorhamnetin intervention. A repeat ofthe isorhamnetin intervention showed 63% decrease in tumornumber (P¼ 0.002) and an 83% decrease in tumor burden (P¼0.02; Fig. 1). In this study, tumors did not progress to adeno-carcinoma on either of the diet interventions. These dataindicate that a diet supplemented with isorhamnetin preventscolorectal tumorigenesis in a mouse model of colitis-associ-ated colorectal cancer.
Isorhamnetin resolves colitis faster in AOM/DSS–treatedmice
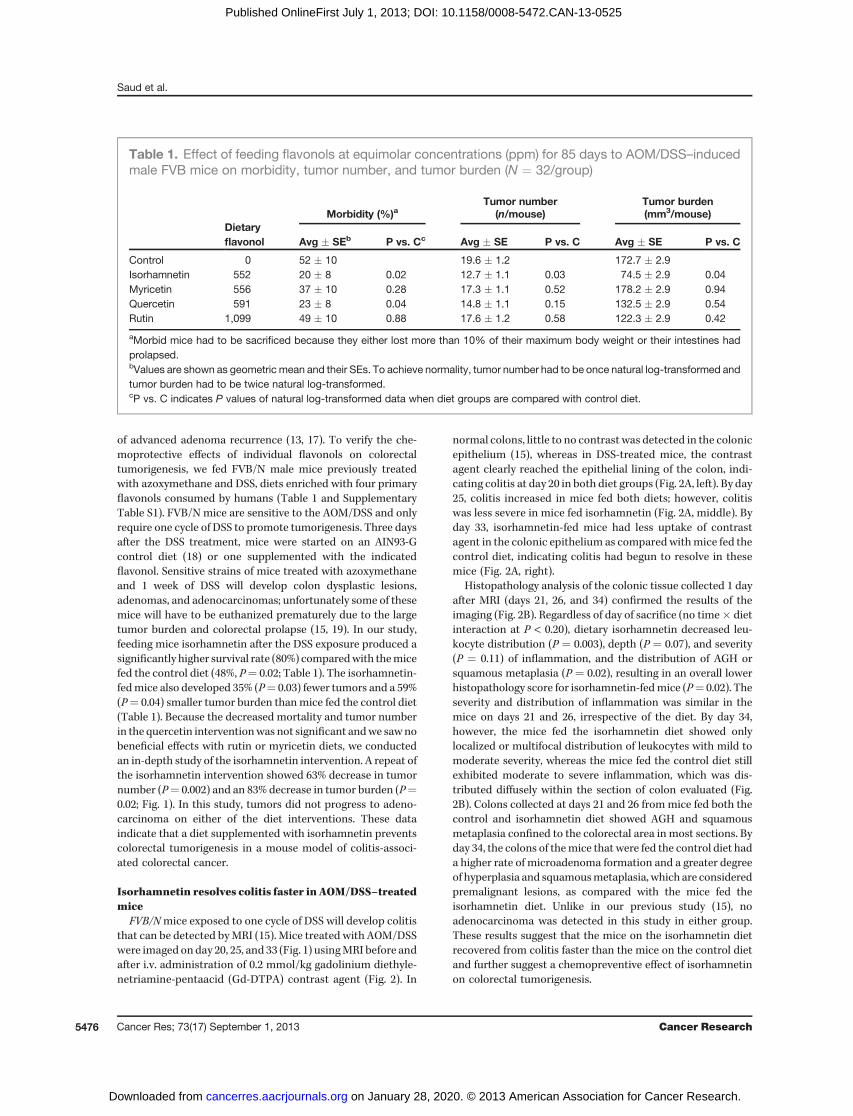
FVB/Nmice exposed to one cycle of DSS will develop colitisthat can be detected byMRI (15). Mice treated with AOM/DSSwere imaged on day 20, 25, and 33 (Fig. 1) usingMRI before andafter i.v. administration of 0.2 mmol/kg gadolinium diethyle-netriamine-pentaacid (Gd-DTPA) contrast agent (Fig. 2). In
normal colons, little to no contrast was detected in the colonicepithelium (15), whereas in DSS-treated mice, the contrastagent clearly reached the epithelial lining of the colon, indi-cating colitis at day 20 in both diet groups (Fig. 2A, left). By day25, colitis increased in mice fed both diets; however, colitiswas less severe in mice fed isorhamnetin (Fig. 2A, middle). Byday 33, isorhamnetin-fed mice had less uptake of contrastagent in the colonic epithelium as comparedwithmice fed thecontrol diet, indicating colitis had begun to resolve in thesemice (Fig. 2A, right).
Histopathology analysis of the colonic tissue collected 1 dayafter MRI (days 21, 26, and 34) confirmed the results of theimaging (Fig. 2B). Regardless of day of sacrifice (no time� dietinteraction at P < 0.20), dietary isorhamnetin decreased leu-kocyte distribution (P ¼ 0.003), depth (P ¼ 0.07), and severity(P ¼ 0.11) of inflammation, and the distribution of AGH orsquamous metaplasia (P ¼ 0.02), resulting in an overall lowerhistopathology score for isorhamnetin-fedmice (P¼ 0.02). Theseverity and distribution of inflammation was similar in themice on days 21 and 26, irrespective of the diet. By day 34,however, the mice fed the isorhamnetin diet showed onlylocalized or multifocal distribution of leukocytes with mild tomoderate severity, whereas the mice fed the control diet stillexhibited moderate to severe inflammation, which was dis-tributed diffusely within the section of colon evaluated (Fig.2B). Colons collected at days 21 and 26 frommice fed both thecontrol and isorhamnetin diet showed AGH and squamousmetaplasia confined to the colorectal area in most sections. Byday 34, the colons of themice that were fed the control diet hada higher rate of microadenoma formation and a greater degreeof hyperplasia and squamousmetaplasia, which are consideredpremalignant lesions, as compared with the mice fed theisorhamnetin diet. Unlike in our previous study (15), noadenocarcinoma was detected in this study in either group.These results suggest that the mice on the isorhamnetin dietrecovered from colitis faster than the mice on the control dietand further suggest a chemopreventive effect of isorhamnetinon colorectal tumorigenesis.
Table 1. Effect of feeding flavonols at equimolar concentrations (ppm) for 85 days to AOM/DSS–inducedmale FVB mice on morbidity, tumor number, and tumor burden (N ¼ 32/group)
DietaryMorbidity (%)a
Tumor number(n/mouse)
Tumor burden(mm3/mouse)
flavonol Avg � SEb P vs. Cc Avg � SE P vs. C Avg � SE P vs. C
Control 0 52 � 10 19.6 � 1.2 172.7 � 2.9Isorhamnetin 552 20 � 8 0.02 12.7 � 1.1 0.03 74.5 � 2.9 0.04Myricetin 556 37 � 10 0.28 17.3 � 1.1 0.52 178.2 � 2.9 0.94Quercetin 591 23 � 8 0.04 14.8 � 1.1 0.15 132.5 � 2.9 0.54Rutin 1,099 49 � 10 0.88 17.6 � 1.2 0.58 122.3 � 2.9 0.42
aMorbid mice had to be sacrificed because they either lost more than 10% of their maximum body weight or their intestines hadprolapsed.bValues are shown as geometric mean and their SEs. To achieve normality, tumor number had to be once natural log-transformed andtumor burden had to be twice natural log-transformed.cP vs. C indicates P values of natural log-transformed data when diet groups are compared with control diet.
Saud et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5476
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525
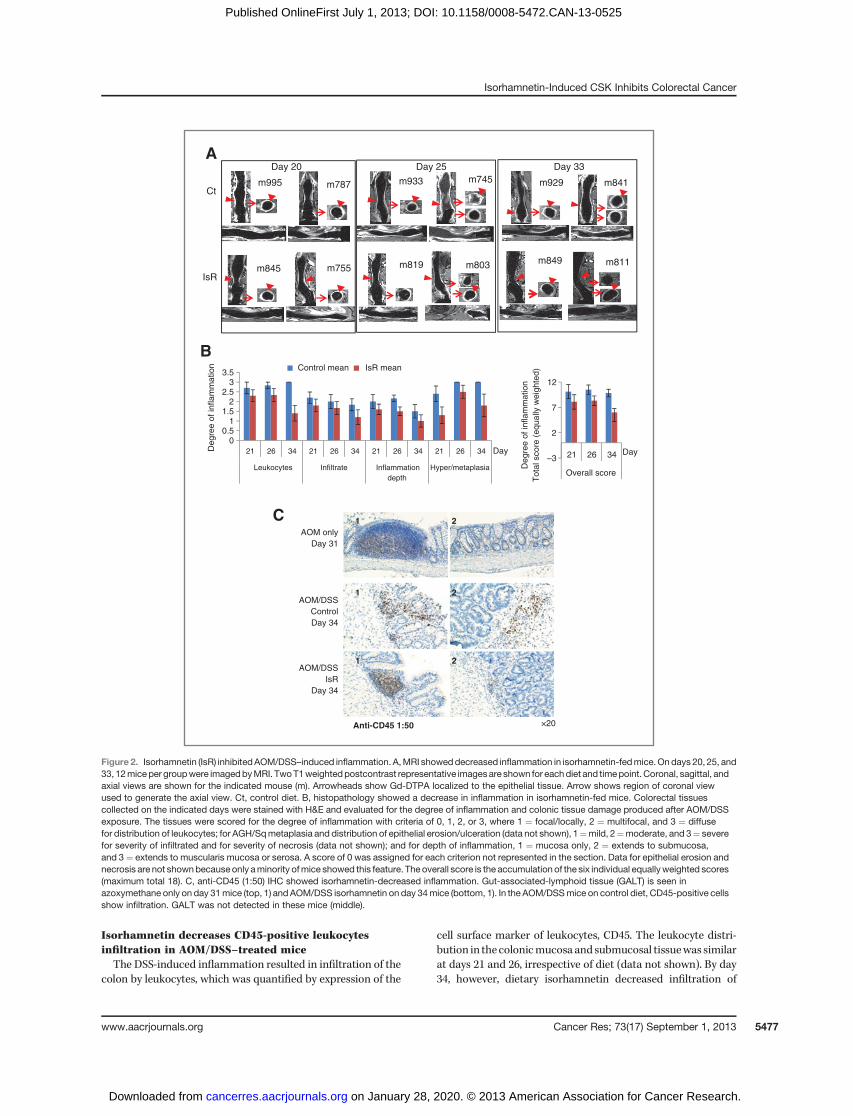
Isorhamnetin decreases CD45-positive leukocytesinfiltration in AOM/DSS–treated miceThe DSS-induced inflammation resulted in infiltration of the
colon by leukocytes, which was quantified by expression of the
cell surface marker of leukocytes, CD45. The leukocyte distri-bution in thecolonicmucosa andsubmucosal tissuewas similarat days 21 and 26, irrespective of diet (data not shown). By day34, however, dietary isorhamnetin decreased infiltration of
21
21
×20
21
Anti-CD45 1:50
AOM only
Day 31
AOM/DSS
IsR
Day 34
AOM/DSS
Control
Day 34
00.5
11.5
22.5
33.5
342621342621342621342621
InflammationInfiltrateLeukocytes
depth
Hyper/metaplasia
Control mean IsR mean
Degre
e o
f in
flam
mation
–3
2
7
12
342621
Overall score
Degre
e o
f in
flam
mation
To
tal sco
re (
eq
ua
lly w
eig
hte
d)
Day Day
A
B
C
Day 20 Day 25 Day 33
IsR
Ctm929
m849
m933
m845 m819
m995 m787m745 m841
m755 m803 m811
Figure 2. Isorhamnetin (IsR) inhibitedAOM/DSS–induced inflammation. A,MRI showeddecreased inflammation in isorhamnetin-fedmice.Ondays 20, 25, and33, 12miceper groupwere imagedbyMRI. TwoT1weightedpostcontrast representative imagesare shown for eachdiet and timepoint. Coronal, sagittal, andaxial views are shown for the indicated mouse (m). Arrowheads show Gd-DTPA localized to the epithelial tissue. Arrow shows region of coronal viewused to generate the axial view. Ct, control diet. B, histopathology showed a decrease in inflammation in isorhamnetin-fed mice. Colorectal tissuescollected on the indicated days were stained with H&E and evaluated for the degree of inflammation and colonic tissue damage produced after AOM/DSSexposure. The tissues were scored for the degree of inflammation with criteria of 0, 1, 2, or 3, where 1 ¼ focal/locally, 2 ¼ multifocal, and 3 ¼ diffusefor distribution of leukocytes; for AGH/Sqmetaplasia and distribution of epithelial erosion/ulceration (data not shown), 1¼mild, 2¼moderate, and 3¼ severefor severity of infiltrated and for severity of necrosis (data not shown); and for depth of inflammation, 1 ¼ mucosa only, 2 ¼ extends to submucosa,and 3¼ extends to muscularis mucosa or serosa. A score of 0 was assigned for each criterion not represented in the section. Data for epithelial erosion andnecrosis are not shownbecauseonly aminority ofmice showed this feature. The overall score is the accumulation of the six individual equallyweighted scores(maximum total 18). C, anti-CD45 (1:50) IHC showed isorhamnetin-decreased inflammation. Gut-associated-lymphoid tissue (GALT) is seen inazoxymethane only on day 31mice (top, 1) and AOM/DSS isorhamnetin on day 34mice (bottom, 1). In the AOM/DSSmice on control diet, CD45-positive cellsshow infiltration. GALT was not detected in these mice (middle).
Isorhamnetin-Induced CSK Inhibits Colorectal Cancer
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5477
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525
CD45-positive cells comparedwithmiceon thecontrol diet (Fig.2C). The expression of CD45was predominantly localized to thegut-associated-lymphoid tissue (GALT) in mice fed isorhamne-tin, similar to what is seen in mice not exposed to DSS. TheGALTprovides immuneprotection to the gut (20). These resultssuggest that isorhamnetin prevents the release of CD45-positiveleukocytes. Consistent with the histopathology and MRI data(Fig. 2A and B), the decrease in CD45-positive leukocyte infil-tration indicates that mice on the isorhamnetin diet recovermore quickly from the colitis than do mice fed the control diet.
Isorhamnetin reduces cell proliferation in AOM/DSS–treated mice
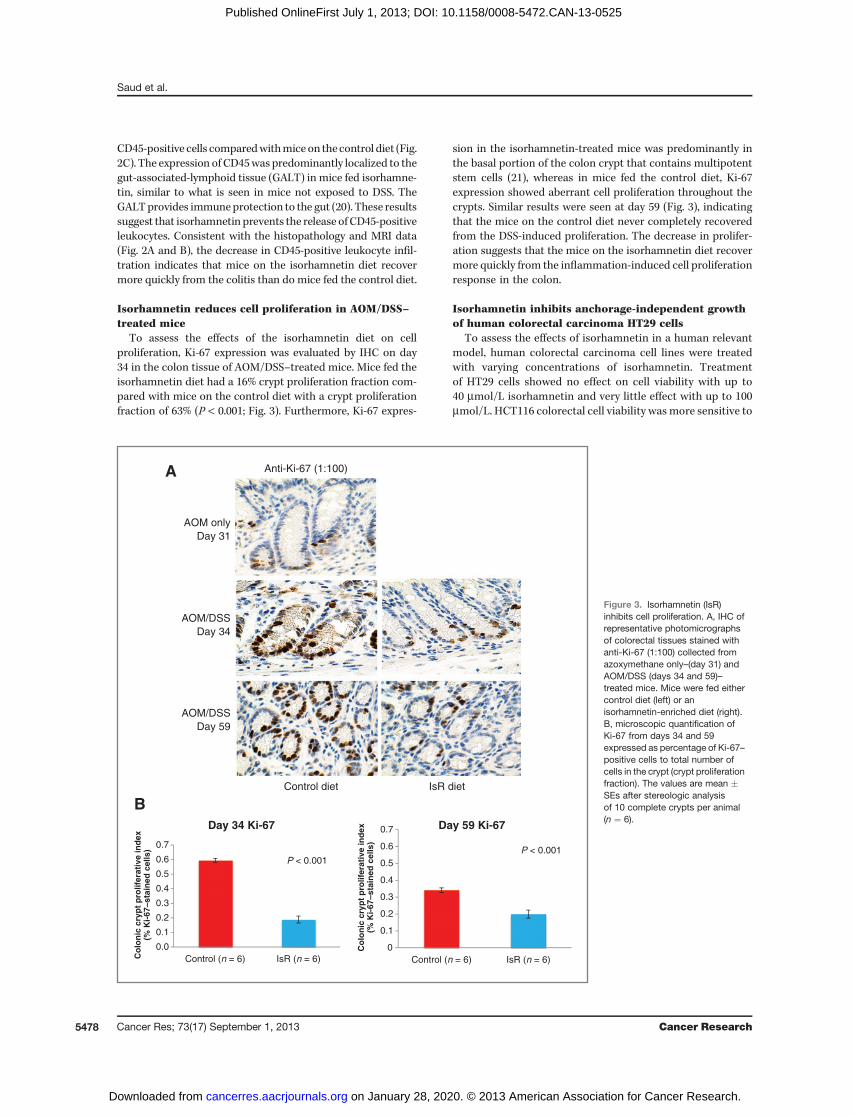
To assess the effects of the isorhamnetin diet on cellproliferation, Ki-67 expression was evaluated by IHC on day34 in the colon tissue of AOM/DSS–treated mice. Mice fed theisorhamnetin diet had a 16% crypt proliferation fraction com-pared with mice on the control diet with a crypt proliferationfraction of 63% (P < 0.001; Fig. 3). Furthermore, Ki-67 expres-
sion in the isorhamnetin-treated mice was predominantly inthe basal portion of the colon crypt that contains multipotentstem cells (21), whereas in mice fed the control diet, Ki-67expression showed aberrant cell proliferation throughout thecrypts. Similar results were seen at day 59 (Fig. 3), indicatingthat the mice on the control diet never completely recoveredfrom the DSS-induced proliferation. The decrease in prolifer-ation suggests that the mice on the isorhamnetin diet recovermore quickly from the inflammation-induced cell proliferationresponse in the colon.
Isorhamnetin inhibits anchorage-independent growthof human colorectal carcinoma HT29 cells
To assess the effects of isorhamnetin in a human relevantmodel, human colorectal carcinoma cell lines were treatedwith varying concentrations of isorhamnetin. Treatmentof HT29 cells showed no effect on cell viability with up to40 mmol/L isorhamnetin and very little effect with up to 100mmol/L. HCT116 colorectal cell viability was more sensitive to
Control diet IsR diet
A Anti-Ki-67 (1:100)
AOM only
Day 31
AOM/DSS
Day 34
AOM/DSS
Day 59
B
P < 0.001
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Control (n = 6) IsR (n = 6)Co
lon
ic c
ryp
t p
rolif
erat
ive
ind
ex(%
Ki-
67–s
tain
ed c
ells
)
Day 34 Ki-67
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Control (n = 6) IsR (n = 6)
Co
lon
ic c
ryp
t p
rolif
erat
ive
ind
ex(%
Ki-
67–s
tain
ed c
ells
)
P < 0.001
Day 59 Ki-67
Figure 3. Isorhamnetin (IsR)inhibits cell proliferation. A, IHC ofrepresentative photomicrographsof colorectal tissues stained withanti-Ki-67 (1:100) collected fromazoxymethane only–(day 31) andAOM/DSS (days 34 and 59)–treated mice. Mice were fed eithercontrol diet (left) or anisorhamnetin-enriched diet (right).B, microscopic quantification ofKi-67 from days 34 and 59expressed as percentage of Ki-67–positive cells to total number ofcells in the crypt (crypt proliferationfraction). The values are mean �SEs after stereologic analysisof 10 complete crypts per animal(n ¼ 6).
Saud et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5478
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525
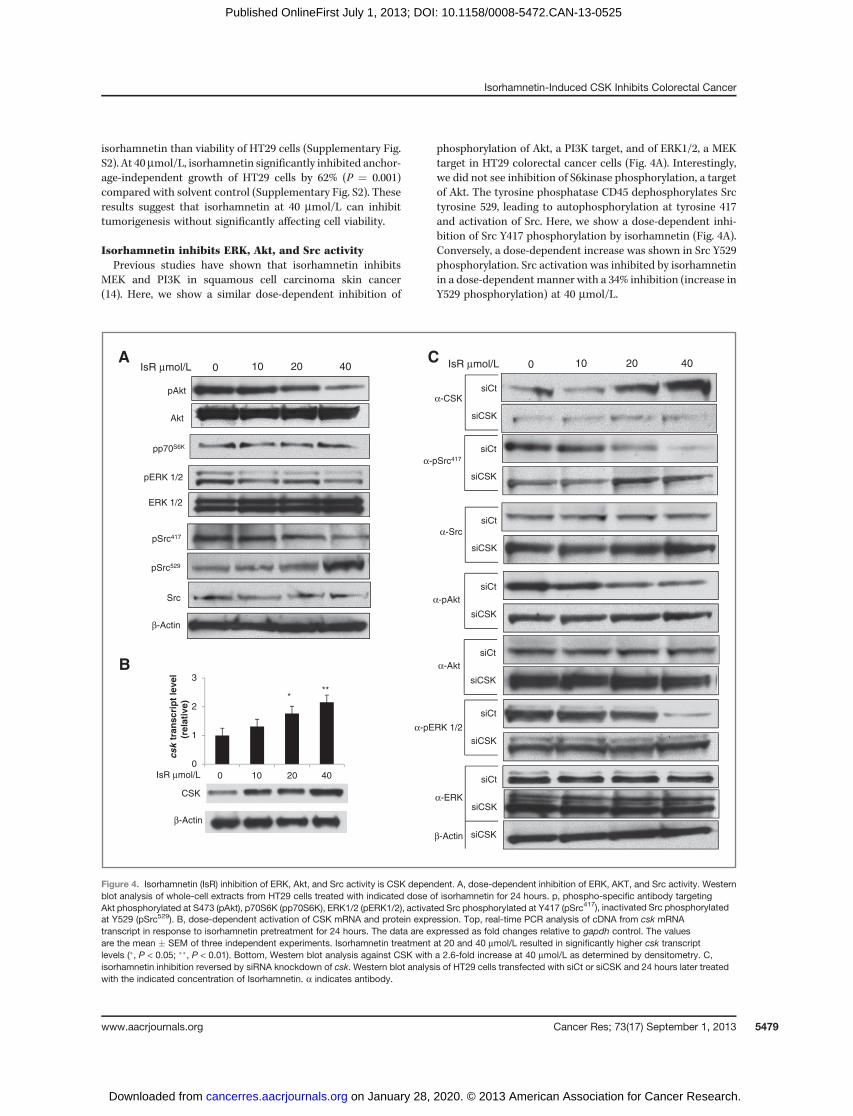
isorhamnetin than viability of HT29 cells (Supplementary Fig.S2). At 40mmol/L, isorhamnetin significantly inhibited anchor-age-independent growth of HT29 cells by 62% (P ¼ 0.001)compared with solvent control (Supplementary Fig. S2). Theseresults suggest that isorhamnetin at 40 mmol/L can inhibittumorigenesis without significantly affecting cell viability.
Isorhamnetin inhibits ERK, Akt, and Src activityPrevious studies have shown that isorhamnetin inhibits
MEK and PI3K in squamous cell carcinoma skin cancer(14). Here, we show a similar dose-dependent inhibition of
phosphorylation of Akt, a PI3K target, and of ERK1/2, a MEKtarget in HT29 colorectal cancer cells (Fig. 4A). Interestingly,we did not see inhibition of S6kinase phosphorylation, a targetof Akt. The tyrosine phosphatase CD45 dephosphorylates Srctyrosine 529, leading to autophosphorylation at tyrosine 417and activation of Src. Here, we show a dose-dependent inhi-bition of Src Y417 phosphorylation by isorhamnetin (Fig. 4A).Conversely, a dose-dependent increase was shown in Src Y529phosphorylation. Src activation was inhibited by isorhamnetinin a dose-dependent manner with a 34% inhibition (increase inY529 phosphorylation) at 40 mmol/L.
CSK
β-Actin
* **
IsR μmol/L 0 10 20 40
0
1
2
3
csk
tran
scri
pt
leve
l (r
elat
ive)
pAkt
Akt
pp70S6K
pERK 1/2
ERK 1/2
pSrc417
Src
β-Actin
pSrc529
A
B
siCSK
α-Src
α-pAkt
α-CSK
α-pSrc417
siCSK
siCt
siCSK
siCt
siCSK
siCt
siCSK
siCt
α-Akt
α-pERK 1/2
siCSK
siCt
α-ERK
β-Actin
0 40 20 10 IsR μmol/L0 40 20 10 IsR μmol/L
siCSK
siCt
siCSK
siCt
C
Figure 4. Isorhamnetin (IsR) inhibition of ERK, Akt, and Src activity is CSK dependent. A, dose-dependent inhibition of ERK, AKT, and Src activity. Westernblot analysis of whole-cell extracts from HT29 cells treated with indicated dose of isorhamnetin for 24 hours. p, phospho-specific antibody targetingAkt phosphorylated at S473 (pAkt), p70S6K (pp70S6K), ERK1/2 (pERK1/2), activated Src phosphorylated at Y417 (pSrc417), inactivated Src phosphorylatedat Y529 (pSrc529). B, dose-dependent activation of CSK mRNA and protein expression. Top, real-time PCR analysis of cDNA from csk mRNAtranscript in response to isorhamnetin pretreatment for 24 hours. The data are expressed as fold changes relative to gapdh control. The valuesare the mean � SEM of three independent experiments. Isorhamnetin treatment at 20 and 40 mmol/L resulted in significantly higher csk transcriptlevels (�, P < 0.05; ��, P < 0.01). Bottom, Western blot analysis against CSK with a 2.6-fold increase at 40 mmol/L as determined by densitometry. C,isorhamnetin inhibition reversed by siRNA knockdown of csk. Western blot analysis of HT29 cells transfected with siCt or siCSK and 24 hours later treatedwith the indicated concentration of Isorhamnetin. a indicates antibody.
Isorhamnetin-Induced CSK Inhibits Colorectal Cancer
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5479
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525
Isorhamnetin induces csk expressionBecause tyrosine 529 on Src is phosphorylated by CSK, we
examined the effect of isorhamnetin on CSK. Isorhamnetininduced a dose-dependent increase in CSK mRNA (2.2-fold change at 40 mmol/L) and protein (2.6-fold change at40 mmol/L; Fig. 4B). Isorhamnetin at 40 mmol/L also inducedCSK expression in the breast cancer cell line MCF7 (datanot shown). These results indicate that isorhamnetin acti-vates gene expression of csk, leading to inhibition of Srcactivity.
Isorhamnetin inhibition of ERK, Akt, and Src activity isCSK dependent
To determine if the isorhamnetin inhibition of Src isdependent on CSK expression, we knocked down csk exp-ression with siRNA and assessed Src activity. Transfectionwith the control siRNA (siCt) did not affect the dose-dependent increase of CSK protein in the isorhamnetin-treated cells, whereas knockdown of the csk gene blockedthe isorhamnetin-induced increase in CSK protein (Fig. 4C).Knockdown of csk expression reversed the inhibition of Srcactivation by isorhamnetin. Transfection with controlsiRNA had no effect on the isorhamnetin-dependent dec-rease in Src Y417 phosphorylation. Conversely, transfection
of HT29 cells with siRNA against csk reversed the isorham-netin-induced decrease in Y417 phosphorylation on Src. Infact, siRNA knockdown of csk resulted in a slight increase inY417 phosphorylation (Fig. 4C), showing that inhibitoryeffects of isorhamnetin on Src is dependent on CSK inHT29 cells.
Transfection of HT29 cells with siCSK also reversed theisorhamnetin inhibition of Akt and ERK activation seen in cellstransfected with the control siRNA (Fig. 4C), indicating thatactivation of the MAPK and PI3K pathways is in part regulatedby Src or an Src family kinase in HT29 cells. Src can activateboth the ERK and the PI3K pathways (22–24). These observa-tions show that in HT29 colorectal cancer cells isorhamnetininduces CSK expression, leading to inactivation of Src byphosphorylation at Y527 and decrease in ERK1/2 and AKTactivity.
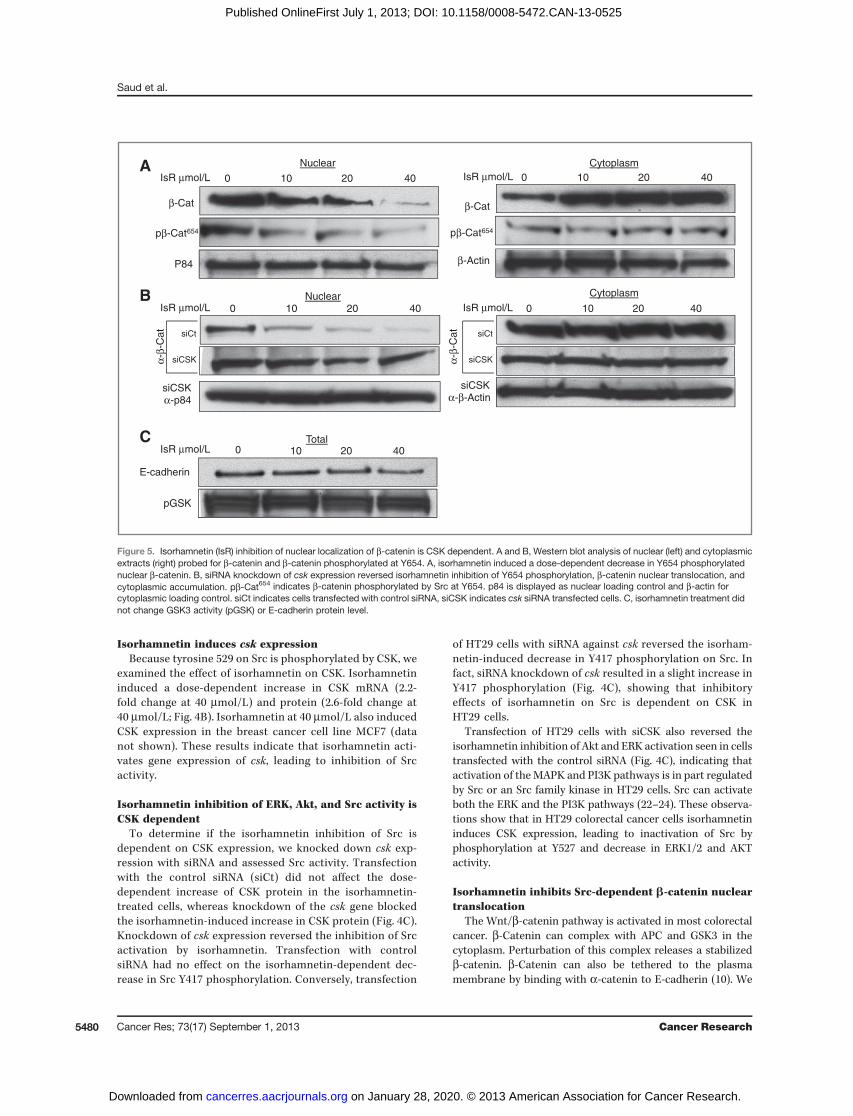
Isorhamnetin inhibits Src-dependent b-catenin nucleartranslocation
The Wnt/b-catenin pathway is activated in most colorectalcancer. b-Catenin can complex with APC and GSK3 in thecytoplasm. Perturbation of this complex releases a stabilizedb-catenin. b-Catenin can also be tethered to the plasmamembrane by binding with a-catenin to E-cadherin (10). We
β-Actin
β-Cat
P84
CytoplasmNuclearα-
β-C
at
siCSKα-p84
β-Cat
siCSKα-β-Actin
pβ-Cat654 pβ-Cat654
Nuclear Cytoplasm
0 40 20 10 0 40 20 10
0 40 20 10 IsR μmol/L
IsR μmol/L IsR μmol/L
IsR μmol/L
IsR μmol/L
0 40 20 10
E-cadherin
pGSK
Total0 40 20 10
siCSK
siCt
siCSK
siCt
α-β-
Ca
t
A
B
C
Figure 5. Isorhamnetin (IsR) inhibition of nuclear localization of b-catenin is CSK dependent. A and B, Western blot analysis of nuclear (left) and cytoplasmicextracts (right) probed for b-catenin and b-catenin phosphorylated at Y654. A, isorhamnetin induced a dose-dependent decrease in Y654 phosphorylatednuclear b-catenin. B, siRNA knockdown of csk expression reversed isorhamnetin inhibition of Y654 phosphorylation, b-catenin nuclear translocation, andcytoplasmic accumulation. pb-Cat654 indicates b-catenin phosphorylated by Src at Y654. p84 is displayed as nuclear loading control and b-actin forcytoplasmic loading control. siCt indicates cells transfected with control siRNA, siCSK indicates csk siRNA transfected cells. C, isorhamnetin treatment didnot change GSK3 activity (pGSK) or E-cadherin protein level.
Saud et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5480
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525

found a dose-dependent reduction in nuclear b-catenin with acorresponding accumulation of cytoplasmicb-catenin inHT29cells treated with isorhamnetin (Fig. 5A). Interestingly, iso-rhamnetin did not change GSK3 activity or E-cadherin levels(Fig. 5C). Alternatively, isorhamnetin inhibited tyrosine phos-phorylation at Y654 on b-catenin (Fig. 5A, nuclear). Tyrosine654 is phosphorylated by Src, which results in release ofb-catenin from its complex with E-cadherin allowing nuclearlocalization (10). The corresponding increase in cytoplasmicb-catenin did not show an increase in phosphorylation at Y654,consistent with a decrease in Src-regulated nuclear transloca-tion of b-catenin. To assess whether nuclear translocation ofb-catenin was Src-dependent, we transfected HT29 cells withcsk siRNA. Knockdown of csk expression reversed the inhib-itory effects of isorhamnetin treatment on Y654 phosphoryla-tion and nuclear translocation of b-catenin (Fig. 5B). Theseresults indicate that nuclear localization of b-catenin in HT29colorectal cancer cells is a consequence of Src phosphorylationof Y654 onb-catenin and that isorhamnetin induces expressionof CSK leads to inhibition of Src phosphorylation of Y654 onb-catenin, blocking its release from E-cadherin and its nuclearlocalization.
Isorhamnetin induces CSK expression and inhibits Srcactivation in vivo
To determine if the effects of isorhamnetin on Src activationand b-catenin nuclear localization seen in HT29 cells can betranslated to the mouse colon, we measured CSK levels, Srcactivity, and b-catenin levels by IHC in AOM/DSS–treatedmice. Similar to the HT29 cells, mice consuming isorhamnetinshowed higher levels of CSK in both the DSS-treated anduntreated groups of mice compared with the mice fed thecontrol diet (Fig. 6A). CSK expression was higher in the laminapropria surrounding the crypts compared with the epithelialtissue. CSK was also detected, although at lower levels, innormal human colon tissue. Furthermore, activated Src (phos-phorylated at Y417) was higher in the crypts of AOM/DSS–treatedmice fed the control diet at both day 34 and 59 (Fig. 6B).Conversely, inactivated Src (phosphorylated at Y529) washigher in crypts of the isorhamnetin-fed mice (Fig. 6B). Theseresults indicate that isorhamnetin induces expression of CSKand inhibits Src activation in vivo in the colons of AOM/DSS–treated mice.
Isorhamnetin inhibits b-catenin nuclear localizationin vivo
Nuclear b-catenin levels were increased in AOM/DSS–trea-ted mice on the control diet, consistent with activation of Src(Fig. 6C, middle). By day 34, AOM/DSS–treated mice on thecontrol diet showed a diffuse staining for b-catenin throughoutthe cells, indicating both cytoplasmic and nuclear accumula-tion of b-catenin. Furthermore, IHC staining with the Y654
Anti-CSK x60
AOM onlyDay 31
AOM/DSSDay 59
Control IsR
AOM/DSSDay 34
Untreated mouse Normal human
IsRControl
AOM only
Anti-pSrc417
AOM/DSS
Anti-pSrc417
Day 34
AOM/DSS
Anti-pSrc417
AOM/DSS
Anti-pSrc529
Day 34
Day 59
Anti-β-catenin Anti-pβ-catenin654
AOM/DSSControlDay 34
AOM/DSSIsR
Day 34
No DSSDay 31
C
B
A
Figure 6. Isorhamnetin (IsR) induces CSK expression and inhibits Srcphosphorylation at Y417, Src phosphorylation of b-catenin at Y654, andb-catenin nuclear localization in vivo. A, IHC analysis of CSK levelsshowed expression was elevated in lamina propria surrounding thecrypts and epithelial tissue of mice fed isorhamnetin diet afterazoxymethane (day 31; top) and after AOM/DSS (days 34 and59;middle).Images are representativephotomicrographsof colonic sections.Normalcolons (no azoxymethane, DSS, or isorhamnetin treatment) from mouse(bottom left) and human colon tissue (bottom right). B, DSS-inducedphosphorylation of Src Y417 was reduced in isorhamnetin-fed mice.Representative photomicrographs depicting activated Src in crypts ofAOM/DSS–treated mice as measured using IHC analysis of pSrc Y417(brown staining). Blue counterstaining of nuclei indicates activated Src isnot nuclear. Isorhamnetin supplementation reduced the amount of Srcactivity as measured at days 34 and 59 (middle left). C-terminalphosphorylated Src (pSrc529) was elevated in isorhamnetin-fed mice atday 34 (bottom). C, nuclear localization and phosphorylation of b-cateninwas reduced in isorhamnetin-fed mice. IHC analysis of b-catenin andY654 phosphorylated b-catenin. b-Catenin and pb-catenin colocalizedwith nuclear counterstaining, indicate phosphorylation and nuclear
localization of b-catenin in AOM/DSS–treated animals (middle). Inisorhamnetin-fed mice, b-catenin staining is predominantly cytoplasmicand with limited p-b-catenin (bottom). Representative serial sections ofcolonic tissue are taken from mice treated with azoxymethane only (day31) or AOM/DSS (day 34).
Isorhamnetin-Induced CSK Inhibits Colorectal Cancer
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5481
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525

phospho-specific b-catenin antibody indicated a high level ofSrc-induced phosphorylation. In contrast, in the isorhamnetin-fed mice, b-catenin remained predominantly in the cytoplasm(Fig. 6C, bottom), with lower amounts of Y654 phosphorylatedb-catenin and decreased nuclear localization. These resultssuggest that Src regulated phosphorylation of b-catenin andthat its nuclear translocation can be inhibited with an iso-rhamnetin-supplemented diet.
DiscussionIn a mouse model of colorectal cancer, we evaluated the
effects of dietary isorhamnetin for colorectal cancer preven-tion. In this model, FVB mice treated with azoxymethaneand DSS develop tumors that will progress to adenoma andadenocarcinomas (Table 1; refs. 15, 19). Dietary isorhamne-tin reduced inflammation, neutrophil infiltration, cell pro-liferation, tumor burden, and mortality associated with theAOM/DSS treatment. Src activation and b-catenin nuclearlocalization induced by AOM/DSS were also reduced in theisorhamnetin-fed mice and in HT29 colon cancer cellstreated with isorhamnetin. Isorhamnetin induced theexpression of CSK, a negative regulator of Src. In HT29 cellsisorhamnetin-induced inhibition of Src activity, and nuclearlocalization of b-catenin was dependent on CSK expression(Fig. 4). Isorhamnetin did not affect the expression of E-cadherin, activation of GSK3, or activation of S6Kinase.These results suggest that the anti-inflammatory and anti-cancer activities of isorhamnetin are linked to inhibition ofoncogenic Src activity, which can phosphorylate b-catenin atY654, leading to its dissociation from the membrane and itsnuclear localization.
b-Catenin signaling in colorectal cancerThe Wnt/b-catenin pathway is activated in most colorectal
cancer (25, 26). This pathway is also activated in the AOM/DSSmouse model (19, 27). b-Catenin can complex with a-cateninand E-cadherin at the cytoplasmic membrane, providingadherin junction communication (10). Activated Src can phos-phorylate b-catenin at Y654, releasing it and a-catenin fromthe E-cadherin complex. Dissociation of b-catenin from thecadherin complex will cause dysregulation of tyrosine kinasesignaling, will affect cell–cell communication, will affectb-catenin–regulated gene expression, and can lead to trans-formation and survival of b-catenin–driven cancer (10).
Activated c-Src in colorectal cancer is an indicator ofpoor prognosis
Members of the SFK family are nonreceptor tyrosine kinasesthat are recruited to the membrane by integrin or receptortyrosine kinase–induced phosphorylation of focal adhesionkinase (FAK). Recruitment of SFK to the membrane providesa molecular switch that is important for regulating prolifera-tion, differentiation, cell adhesion, and cell mobility (28).Activated Src in primary colorectal cancer is an indicator ofpoor prognosis (7), and elevated Src activity can be detected inthe majority of human colon cancer (29). Inhibition of Src orSFK can enhance cell to cell adhesion and can suppressmigration and invasion in vitro andmetastasis in vivo, suggest-
ing an anti-invasive role for Src inhibitors (10). Inhibitors of Srcare currently being tested in clinical trials (10).
CSK negatively regulates SrcThe CSK is a nonreceptor tyrosine kinase that serves as a
negative regulator of Src and SFK. CSK phosphorylates theC-terminal regulatory site of SFK, resulting in a conforma-tion change and inactivation of the kinase activity (30–33). Areduction in CSK mRNA, protein, and kinase activity incolorectal carcinoma has been shown to be correlated withan increase in Src activity, suggesting that a loss in CSKmay influence transformation of colorectal carcinoma (8).Rengifo-Cam and colleagues have shown that CSK regulatessignaling from integrin–SFK–mediated cell adhesion, whichcan influence the metastasis of cancer cells (34). Recruit-ment to the membrane by scaffolding proteins such as CSK-binding protein (Cbp) is required for inactivation of Srcby CSK and is crucial for preventing tumorigenesis (31, 35).The fact that none of the other flavonols tested, includingmycetin, quercetin, and rutin, induced expression of csk inHT29 cells (results not shown) and were not effective forinhibiting carcinogenesis in vivo is consistent with ourconclusion that the chemoprotective effects of isorhamnetinin colon cancer are linked to its anti-inflammatory activitiesand its inhibition of oncogenic Src activity and consequen-tial loss of nuclear b-catenin, activities that are dependenton csk expression. Although csk expression can be regulatedtranslationally (36), very little is known about how csk isregulated transcriptionally. Further investigation is neededto learn how csk expression is lost in colorectal cancer andhow isorhamnetin restores csk expression.
Dietary isorhamnetin has chemoprotective propertiesIsorhamnetin, quercetin, kaempferol, and myricetin are
flavonols that are present in a wide variety of fruits andvegetables and have anticancer activity (37). Computationaland binding assays have shown that isorhamnetin can binddirectly to MEK1 and to PI3K and quercetin can bind toRSK2 (14, 38). Lee and colleagues (39) reported that kaemp-ferol can bind to Src, and Jung and colleagues (40) showedthat myricetin can bind to the SFK Fyn. Our findings showthat isorhamnetin can inhibit Src activity, but that thisinhibition is dependent on CSK expression, suggesting thatunlike the structurally similar flavonols, kaempferol, andmyricetin, isorhamnetin does not bind directly to Src. Themechanism of how isorhamnetin is upregulating CSKexpression is currently under investigation. Our results andthose of others (8, 34, 35) show the importance of CSK as anegative regulator of SFK and as a tumor suppressor, sug-gesting that preventing the loss of or restoring the expres-sion of CSK would be beneficial for preventing tumorigen-esis, tumor progression, and tumor metastasis.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: M.R. Young, N.H. Colburn, Y.S. Kim, G. BobeDevelopment of methodology: M.R. Young, M.O. Evbuomwan, G. Bobe
Saud et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5482
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525

Acquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): S.M. Saud, M.R. Young, Y.L. Jones-Hall, L. Ileva, M.O.Evbuomwan, G. BobeAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): S.M. Saud, M.R. Young, G. BobeWriting, review, and/or revision of the manuscript: S.M. Saud, M.R. Young,N.H. Colburn, G. BobeAdministrative, technical, or material support (i.e., reporting or orga-nizingdata, constructingdatabases): S.M. Saud,M.R. Young, J.Wise, Y.S. Kim,G. BobeStudy supervision: S.M. Saud, M.R. Young, N.H. Colburn, Y.S. Kim, G. Bobe
AcknowledgmentsThe authors thankCraig Driver of the Laboratory Animal Sciences Programof
SAIC-Frederick, Darlene Green and Tammy Beachley of the Pathology/Histo-technology Laboratory of SIAC-Frederick, and Thomas G. McCloud from the
Natural Products Support Group of AppliedDevelopmental Research Program ofSAIC-Frederick.
Grant SupportThis study was funded by the Office of Complementary and Alternative
Medicine, Office of Dietary Supplements, the Division of Cancer Prevention andthe Intramural Research Program, National Cancer Institute, NIH, Departmentof Health and Human Services (DHHS), Bethesda, MD.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicate thisfact.
Received February 28, 2013; revised June 7, 2013; accepted June 8, 2013;published OnlineFirst July 1, 2013.
References1. American CS. Cancer facts & figures 2011. Atlanta, GA: American
Cancer Society; 2011.2. Shacter E, Weitzman SA. Chronic inflammation and cancer. Oncology
2002;16:217–26.3. Shenoy AK, Fisher RC, Butterworth EA, Pi L, Chang LJ, Appelman HD,
et al. Transition from colitis to cancer: high Wnt activity sustains thetumor-initiating potential of colon cancer stem cell precursors. CancerRes 2012;72:5091–100.
4. Eaden JA, Abrams KR, Mayberry JF. The risk of colorectal cancer inulcerative colitis: a meta-analysis. Gut 2001;48:526–35.
5. Gyde S, Prior P, Dew MJ, Saunders V, Waterhouse JA, Allan RN.Mortality in ulcerative colitis. Gastroenterology 1982;83:36–43.
6. LieuC,Kopetz S. TheSRC family of protein tyrosine kinases: a newandpromising target for colorectal cancer therapy. Clin Colorectal Cancer2010;9:89–94.
7. Aligayer H, Boyd DD, Heiss MM, Abdalla EK, Curley SA, Gallick GE.Activationof Src kinase in primary colorectal carcinoma: an indicator ofpoor clinical prognosis. Cancer 2002;94:344–51.
8. Cam WR, Masaki T, Shiratori Y, Kato N, Ikenoue T, Okamoto M,et al. Reduced C-terminal Src kinase activity is correlated inverselywith pp60(c-src) activity in colorectal carcinoma. Cancer 2001;92:61–70.
9. Hermiston ML, Zikherman J, Zhu JW. CD45, CD148, and Lyp/Pep:critical phosphatases regulating Src family kinase signaling networksin immune cells. Immunol Rev 2009;228:288–311.
10. Wadhawan A, Smith C, Nicholson RI, Barrett-Lee P, Hiscox S. Src-mediated regulation of homotypic cell adhesion: implications forcancer progression and opportunities for therapeutic intervention.Cancer Treat Rev 2011;37:234–41.
11. Gill S, Sinicrope FA. Colorectal cancer prevention: is an ounce ofprevention worth a pound of cure? Semin Oncol 2005;32:24–34.
12. American CS. Cancer facts & figures 2010. Atlanta, GA: AmericanCancer Society; 2010.
13. Bobe G, Sansbury LB, Albert PS, Cross AJ, Kahle L, Ashby J, et al.Dietary flavonoids and colorectal adenoma recurrence in the PolypPreventionTrial. Cancer Epidemiol BiomarkersPrev2008;17:1344–53.
14. Kim JE, Lee DE, Lee KW, Son JE, Seo SK, Li J, et al. Isorhamnetinsuppresses skin cancer through direct inhibition of MEK1 and PI3-K.Cancer Prev Res 2011;4:582–91.
15. Young MR, Ileva LV, Bernardo M, Riffle LA, Jones YL, Kim YS, et al.Monitoring of tumor promotion and progression in a mouse model ofinflammation-induced colon cancer with magnetic resonance colono-graphy. Neoplasia 2009;11:237–46.
16. Mitsunaga M, Kosaka N, Choyke PL, Young MR, Dextras CR, SaudSM, et al. Fluorescence endoscopic detection of murine colitis-asso-ciated colon cancer by topically applied enzymatically rapid-activa-table probe. Gut 2013;62:1179–86.
17. Bobe G, Barrett KG, Mentor-Marcel RA, Saffiotti U, Young MR,Colburn NH, et al. Dietary cooked navy beans and their fractionsattenuate colon carcinogenesis in azoxymethane-induced ob/obmice. Nutr Cancer 2008;60:373–81.
18. Reeves PG, Nielsen FH, Fahey GC Jr. AIN-93 purified diets for labo-ratory rodents: final report of the American Institute of Nutrition ad hocwriting committee on the reformulation of the AIN-76A rodent diet.J Nutr 1993;123:1939–51.
19. Tanaka T, Kohno H, Suzuki R, Yamada Y, Sugie S, Mori H. A novelinflammation-related mouse colon carcinogenesis model induced byazoxymethane and dextran sodium sulfate. Cancer Sci 2003;94:965–73.
20. Salminen S, Bouley C, Boutron-Ruault MC, Cummings JH, Franck A,Gibson GR, et al. Functional food science and gastrointestinal phys-iology and function. Br J Nutr 1998;80(Suppl 1):S147–71.
21. Barker N, van Es JH, Kuipers J, Kujala P, van denBornM,CozijnsenM,et al. Identification of stem cells in small intestine and colon by markergene Lgr5. Nature 2007;449:1003–7.
22. Irby RB, Yeatman TJ. Role of Src expression and activation in humancancer. Oncogene 2000;19:5636–42.
23. Karni R, Gus Y, Dor Y, Meyuhas O, Levitzki A. Active Src elevates theexpression of beta-catenin by enhancement of cap-dependent trans-lation. Mol Cell Biol 2005;25:5031–9.
24. Penuel E, Martin GS. Transformation by v-Src: Ras-MAPK and PI3K-mTOR mediate parallel pathways. Mol Biol Cell 1999;10:1693–703.
25. Clevers H, Nusse R. Wnt/beta-catenin signaling and disease. Cell2012;149:1192–205.
26. Kim YS, Milner JA. Dietary modulation of colon cancer risk. J Nutr2007;137:2576S–9S.
27. Salcedo R, Worschech A, Cardone M, Jones Y, Gyulai Z, Dai RM,et al. MyD88-mediated signaling prevents development of adeno-carcinomas of the colon: role of interleukin 18. J Exp Med 2010;207:1625–36.
28. Thomas SM, Brugge JS. Cellular functions regulated by Src familykinases. Annu Rev Cell Dev Biol 1997;13:513–609.
29. Talamonti MS, RohMS, Curley SA, Gallick GE. Increase in activity andlevel of pp60c-src in progressive stages of human colorectal cancer.J Clin Invest 1993;91:53–60.
30. Nada S, Okada M, MacAuley A, Cooper JA, Nakagawa H. Cloning of acomplementary DNA for a protein-tyrosine kinase that specificallyphosphorylates a negative regulatory site of p60c-src. Nature 1991;351:69–72.
31. Okada M. Regulation of the SRC family kinases by Csk. Int J Biol Sci2012;8:1385–97.
32. Okada M, Nada S, Yamanashi Y, Yamamoto T, Nakagawa H. CSK: aprotein-tyrosine kinase involved in regulation of src family kinases.J Biol Chem 1991;266:24249–52.
33. Sabe H, Knudsen B, Okada M, Nada S, Nakagawa H, Hanafusa H.Molecular cloning and expression of chicken C-terminal Src kinase:lack of stable association with c-Src protein. Proc Natl Acad Sci U S A1992;89:2190–4.
34. Rengifo-CamW, Konishi A, Morishita N, Matsuoka H, Yamori T, NadaS, et al. Csk defines the ability of integrin-mediated cell adhesion andmigration in human colon cancer cells: implication for a potential role incancer metastasis. Oncogene 2004;23:289–97.
Isorhamnetin-Induced CSK Inhibits Colorectal Cancer
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5483
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525

35. Oneyama C, Hikita T, Enya K, Dobenecker MW, Saito K, Nada S, et al.The lipid raft-anchored adaptor protein Cbp controls the oncogenicpotential of c-Src. Mol Cell 2008;30:426–36.
36. Liang F, LuoY, DongY,Walls CD, Liang J, JiangHY, et al. Translationalcontrol of C-terminal Src kinase (Csk) expression by PRL3 phospha-tase. J Biol Chem 2008;283:10339–46.
37. Birt DF, Hendrich S, Wang W. Dietary agents in cancer preven-tion: flavonoids and isoflavonoids. Pharmacol Ther 2001;90:157–77.
38. ChenH,YaoK,Nadas J, BodeAM,MalakhovaM,OiN, et al. Predictionof molecular targets of cancer preventing flavonoid compounds usingcomputational methods. PLoS ONE 2012;7:e38261.
39. LeeKM, LeeKW, JungSK, LeeEJ,HeoYS,BodeAM, et al. Kaempferolinhibits UVB-induced COX-2 expression by suppressing Src kinaseactivity. Biochem Pharmacol 2010;80:2042–9.
40. Jung SK, Lee KW, Byun S, Kang NJ, Lim SH, Heo YS, et al. Myricetinsuppresses UVB-induced skin cancer by targeting Fyn. Cancer Res2008;68:6021–9.
Saud et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5484
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525

2013;73:5473-5484. Published OnlineFirst July 1, 2013.Cancer Res Shakir M. Saud, Matthew R. Young, Yava L. Jones-Hall, et al.
-CateninβColorectal Cancer Is Mediated by Oncogenic Src and Chemopreventive Activity of Plant Flavonoid Isorhamnetin in
Updated version
10.1158/0008-5472.CAN-13-0525doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/07/10/0008-5472.CAN-13-0525.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/73/17/5473.full#ref-list-1
This article cites 38 articles, 13 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/73/17/5473To request permission to re-use all or part of this article, use this link
on January 28, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 1, 2013; DOI: 10.1158/0008-5472.CAN-13-0525





























