Embed Size (px)
Citation preview

J Sci Food Agric 1996,72,127-134
Chemical Composition of Pea Fibre Isolates and their Effect on ihe Endogenous Amino Acid Flow at the Ileum of the Pig* Pascal Leterme,"S Andr6 Thkwis," Piet van Leeuwen,b Thierry Monmart" and Joop Huismanb a Faculte des Sciences Agronomiques, UER de Zootechnie, B-5030 Gembloux, Belgium
(Received 21 August 1995; revised version received 6 January 1996; accepted 10 May 1996)
ILOB-TNO, PO Box 15,6700 AA Wageningen, The Netherlands
Abstract: Pea starch, hulls and cotyledon inner fibres, isolated from pea seeds, were incorporated in N-free diets, on a NDF-content basis, in order to study their effect on the ileal endogenous amino acid (AA) excretion in the growing pig. Maize starch and wood cellulose were selected as references. The inner fibre- containing diet was also supplemented with enzymatically hydrolysed casein (EHC) in order to test the peptide alimentation method on a diet containing a fibre source with a high water-holding capacity (WHC = 10.9 g water g-' DM). The fibres were also analysed by different methods (crude fibre, NDF, ADF, enzymatic-gravimetric method, Englyst method). The fibre content of the inner fibre fraction varied widely from one method to another and ranged from 109 g crude fibre to 480 g AOAC fibre kg-' DM. Compared to the 'maize starch + wood cellulose' diet (11.0 g AA excreted kg-' DM intake), pea starch had no effect on ileal AA losses (9.6 g), whereas pea hulls slightly increased them (14.2 g). The AA excretion dramatically increased with the incorporation of inner fibres (28.0 g), because of their very high WHC. The addition of EHC increased the output (48.0 g) further, at a level that can hardly be explained only by an increase of the endogenous secretions. The results suggest that the NDF content of grain legume products is not indicative of their effect on the ileal endogenous AA losses in pigs and that the physiological effects of fibres along the gastro- intestinal tract are due to both their chemical and physical properties.
Key words: pea, fibre, pig, analysis, endogenous amino acid, ileum.
INTRODUCTION
The ileal endogenous nitrogen or amino acid (AA) losses are increasingly studied in pigs because they rep- resent a significant part of the N excreted in the environment and because the presence of endogenous N compounds in the ileal digesta impairs the measurement of the true ileal digestibility of dietary proteins. These losses are often estimated by means of N-free diets, sup- plemented by wood cellulose. However, two studies concluded that this fibre source has no effect on ileal
endogenous AA flows, whatever its content in the diet (Furuya and Kaji 1992; Leterme et a1 1992). On the contrary, other fibre sources-like wheat bran-have a positive effect on ileal endogenous AA losses, in propor- tion to the fibre content of the diet (Schulze et a1 1994, 1995; Mariscal-Landin et al 1995). Relationships between endogenous losses and the proportion of the feed component that induces them would be of great value for the nutritionist and would allow him to com- pensate for the losses.
Peas are widely used in pig nutrition. While present in limited amounts, their antinutritional factors (eg trypsin inhibitors) slightly increase the ileal endogenous N excretion but do not affect the protein digestibility significantly (Huisman et al 1992). So far, the study of
* Presented in part at the 7th International Symposium on Digestive Physiology in Pigs, Bad Doberan, Germany, 1-4 October 1994. 1 To whom correspondence should be addressed.
J Sci Food Agric 0022-5142/96/$09.00 0 1996 SCI. Printed in Great Britain
the gastrointestinal implications of pea fibre intake has 127

128 P Leterme et a1
been neglected and, most of the time, has been limited at the fecal level of rats (Goodlad and Mathers 1992; Hansen et al 1992; Zhao et a1 1995). These studies and others, performed on pigs with pea hulls (Lien et al 1994), demonstrated an effect of pea fibre intake on both protein digestibility and endogenous losses.
Pea fibres are increasingly incorporated in human food, as dietary fibre sources (Ralet et al 1993; Sand- strom et al 1994). In the current context of the healthy diet, increased grain legume consumption is being pro- moted in order to reduce the risk of coronary heart disease, diabetes and obesity (Geil and Anderson 1994). Grain legumes are protein sources but are also con- sidered now for their starch and dietary fibre supply. These fibres are highly fermentable in the large intes- tine, providing a substantial amount of energy in the form of volatile fatty acids (Goodlad and Mathers 1990, 1992). The fermentation also helps to reduce the risks of colon cancer (Geil and Anderson 1994), blood choles- terol level (Costa et al 1994) and post-prandial lipidae- mia (Sandstrom et a1 1994). Pea hulls have been used for years as dietary fibre sources in bread, etc. The coty- ledon inner fibres, which account for 90% of the total seed fibres, are now also available on the market. Their high water-holding capacity (WHC) makes them ideal for meat products, dressings, soups, etc. However, few data are available on the effect of these fibres, either on the digestive processes or on the endogenous secretions in the small intestine.
To study the effect of pea starch, hulls and cotyledon inner fibres on the ileal endogenous AA excretion, an experiment with pigs was carried out. As fibre analysis is difficult on grain legumes (Giger et a1 1987; Mongeau and Brassard 1994), the fibre sources were analysed by five different methods. The peptide alimentation method, suggested by Moughan et al (1990) and Butts et al(1991, 1992, 1993) to avoid the depressive effect of N-free diets on the endogenous secretions, was also tested on the inner fibre-containing diet.
MATERIAL AND METHODS
Animals
Ten Landrace x Yorkshire (mean bodyweight: 25 _+ 2 kg) were fitted with a post-valvular T-cannula (PVTC) as described by van Leeuwen et al (1991). The caecum is removed and replaced by a large T-cannula. After the caecum is cut, the flange of the cannula is introduced in the large intestine and the cannula aper- ture is placed in front of the ileocaecal valve. The flange is fixed to the intestinal wall by tightening a pre-placed purse-string suture. The cannula is then exteriorised through an incision in the body wall, fixed externally by mounting a ring of silicone rubber and closed with a stop, also in silicone rubber. When the cannula is
closed, the digesta coming from the ileum flow into the colon. When it is open, the ileocaecal valve is protruded in the aperture of the cannula, the digesta flow directly in it and are collected with plastic bags.
After surgery, the pigs were kept in large cages (1.2 x 1.0 m) and allowed a 10 day recovery period.
Diets
Pea starch, hulls and inner fibres were provided by Pro- vital (Warcoing, Belgium). Only peas (Pisurn satiuum L subsp hortense) with yellow seeds are utilised. During the industrial process, the hulls were first mechanically separated from the seeds. The cotyledons were then finely ground and plunged into an alkaline solution (pH 8) for protein solubilisation. The proteins were iso- lated by ultrafiltration and starch was allowed to settle and decant. The remaining product, composed of fibres and starch, forms the 'inner fibre' fraction utilised here.
Five diets were formulated in order to provide the same amount of NDF fibres to the pigs (Table 1). Enzy- matically hydrolysed casein (EHC) was added to a diet containing inner fibres, in order to meet the pig's protein requirements for growth (160 g protein kg-') at the expense of sucrose present in the N-free diet. The diets contained approximately the same amounts of starch, either maize starch alone (wood cellulose diet) or a mixture of maize and pea starch (1 : 1) (Table 1). Each meal was mixed with water (1 : 1) immediately prior to feeding, except for the meals containing inner fibres for which the double amount was added in order to com- pensate for the very high water-holding capacity of the fibres. Fresh water was also freely available. Chromic oxide was added for the determination of the digesta flows (3 g Cr,O, kg-' DM).
Experimental procedure
The experiment was conducted as a cross-over design with two periods and two pigs per diet per period. The pigs were fed twice daily (8-20 h) in equal portions, at a feed intake level of 2.6 times the maintenance require- ments (& 1 kg day-') and for five days in each period. The ileal digesta were collected for the last three days from 8 to 20 h with plastic bags attached to the PVTC cannula and immediately frozen. Between the two periods, the pigs were fed a balanced diet and then randomly allotted to a second experimental diet.
Chemical analyses
The pea fibrous fractions were analysed for nitrogen (Kjeldahl method using a Kjeltec 1030 analyser), starch (with amyloglucosidase), crude fibre (Weende method using a Fibertec analyser), neutral (NDF) and acid detergent fibre (ADF) as described by van Soest and

Composition of pea Jibres and eflect on ileal endogenous amino acid flow in pigs 129
TABLE 1 Composition and fibre content of the experimental diets (g kg-' DM)
Diets Wood cellulose Wood cellulose Inner fibre Inner fibre Hulls + peas starch + EHC
Composition Maize starch Pea starch Sucrose Wood cellulose" Inner fibres Hulls EHCb Minerallvitamin premix
Fibre content Crude fibre NDF AOAC Englyst
615
260 80
-
- - - 45
62 80 80 74
295 315 265 80 - - - 45
62 80 80 14
270 190 175
320 -
- - 45
35 80
154 116
270 190 - -
320
175 45
-
35 80
154 116
295 320 220 - - 120
45 -
61 83
110 99
" Wood cellulose: Arbocel@. EHC (Sigma N-4642; N-Z-Case Plus, from bovine milk) (g kg-' DM): 42.4 arg, 25.4 his, 47.7 ile, 85.5
leu, 81.4 lys, 28.6 met, 43.2 phe, 38.6 thr, 13.5 trp, 55.1 Val, 31.3 ala, 66.9 asp, 5.2 cys, 180 glu, 18-2 gly, 90.1 pro, 54.3 ser, 14.9 tyr, sum 922.3; 56% free amino acids, 44% di- and tripeptides.
Wine (1967), using a Fibertec analyser. The NDF and ADF determinations were performed separately and, prior to these analyses and that of crude fibre, the samples were boiled for 1 h in a thermostable a-amylase solution (Termamyl 120L, Novo Nordisk, Denmark; 1%) in order to remove starch.
The total, soluble and insoluble dietary fibre contents were also determined by an enzymatic-gravimetric method (AOAC method described by Prosky et a1 1988, 1994) and by a chromatographic method where dietary fibres are the sum of the constituent sugars of the non- starch polysaccharides (NSP) released by acid hydro- lysis (Englyst and Cummings 1988; Englyst et a1 1992, referred to as the Englyst method). The uronic acids were analysed with the colorimetric method of Scott (1979) using 3,5-dimethylphenol. All the analyses were made in duplicate and, for the Englyst method, the series of analyses included a reference sample provided by Mrs H Johansen (NIAS, Denmark).
The water-holding capacity (WHC) was determined as follows: 1 g DM was placed in a centrifuge tube with 20 ml of distilled water. After 16 h, the tubes were cen- trifuged (2000 x g, 50 min at 20°C) and were kept 8 min before the supernatant water was discarded and the remaining water weighed.
The digesta were analysed for nitrogen, amino acids and chromium. The amino acids were analysed after acid hydrolysis (6 M HC1 + 1% phenol; 24 h at 110°C in glass tubes under N atmosphere) by gas chromatog- raphy or GC (Carlo Erba HRCG 5160; J & W-DB1 column, J & W Scientific Ltd, Folsom, CA, USA) using heptafluorobutyril iso-butyl ester amino acid derivatives
(MacKenzie and Tenaschuk 1979a, b). Cysteine and methionine were determined by ion exchange chroma- tography using a Pharmacia-LKB Plus I1 analyser after oxidation using performic acid. Tryptophan was analysed by the Slump and Schreuder (1969) method, as adapted by Leterme and Monmart (1990). After alkaline hydrolysis of the sample, tryptophan was separated by gel filtration chromatography and measured spectro- photometrically at 570 nm after reaction with ninhy- drin. The chromium content was determined by titration with Mohr's salt after nitroperchloric attack, as described by FranGois et a1 (1978).
Statistical evaluation
Results are presented as mean values and standard errors of the mean. The results of each period were tested statistically in an analysis of variance test using a General Linear Model (GLM) procedure. As no signifi- cant differences were found between the periods, the results from the two periods were pooled and analysed by the multiple comparison test of Newman-Keuls, using a computerised statistical software package (SAS, SAS Institute Inc, Cary, NC, USA).
RESULTS AND DISCUSSION
Chemical composition of the fibre sources
The results of fibre content (Table 2) illustrate the diffi- culty of analysing grain legume products correctly. In their comparative study, Wolters et a1 (1992) concluded

130 P Leterme et a1
TABLE 2 Water holding capacity (g water g-' DM) and chemical composition of
the fibre sources (g kg- DM)
Wood Pea Pea cellulose inner fibre hulls
Water holding capacity 3.5 10.9 4.3
Chemical composition Crude protein (N x 6.25) Starch Crude fibre NDF ADF AOAC
Total Insoluble Soluble
Engl yst Total Insoluble Soluble
ND ND 772 997 937
1000 1000 ND
929 91 1
18
38 422 109 250 152
480 420 60
362 284 78
53 13
51 1 695 628
920 88 1 39
824 664 160
that the NDF and ADF methods are not suited for determination of dietary fibres in soya products. The ADF fraction, for example, contained significant amounts of xylose and uronic acids, whereas it was sup- posed to contain only cellulose and lignin. The NDF residues may also contain some resistant starch. Before treatment with detergents, the samples were treated with a thermostable a-amylase. However, Giger et al (1987) demonstrated that the treatment with an a- amylase is partially inefficient for grain legumes : the NDF residue still contained from 15 to 25% of starch with faba beans and from 36 to 40% with pea samples.
The AOAC and Englyst methods are better adapted for such products (Wolters et a1 1992). However, there are still differences between both methods: the values of total dietary fibres are systematically lower for Englyst's method (Periago et a1 1994). In the latter, the constitu- ent sugars of the fibres are released by acid hydrolysis, before being measured by gas chromatography. This hydrolysis causes some losses of each sugar that could lead to an underestimation of the fibre content. These losses were taken into account here because we analysed a standard mixture of the sugars and applied a correction for their respective loss. However, the recovery of the sugars is subjected to the conditions used for acid hydrolysis of the polymers (Englyst et al 1992).
The AOAC fibre content of the pea inner fibres is probably slightly overestimated. With this method, starch is supposed to be removed by amylase treatment and the resistant starch is considered as dietary fibre (Periago et a1 1994). When inner fibres are isolated, the most available starch of the seed is allowed to settle and decant. The remaining fraction is partly held within the
unaltered cells and must be distinguished from the retrograded amylose. Despite the grinding of the samples, it may escape the action of amylase and con- tribute to overestimating the fibre content. Mongeau and Brassard (1994) demonstrated that discrepancies among gravimetric methods for fibre analysis of canned grain legumes were reduced by 60-98% when the starch digestion problem was resolved. With the Englyst method, starch is scattered with dimethylsulfoxide and the glucose residues detected by GC are considered to come from cellulose only. From all this, it may be concluded-in agreement with Wolters et a1 (1992) and Periago et al (1994bthat the AOAC method slightly overestimates the fibre content of the pea products and that the Englyst method slightly underestimates it.
The proportion of soluble fibres in the pea products is quite variable from one technique to another (Table 2). The data available in literature are also quite differ- ent: from 4.5 to 9.4% of the pea hull fibres with the AOAC method (Ralet et al 1993) and 44% of the inner fibres (% total fibres) with the Englyst method (Hansen et al 1992). In both methods, soluble fibres are precipi- tated with cooled ethanol and the proportion of soluble fibre is calculated by difference between the total and insoluble fractions. Such approaches can lead to inaccu- racies. Moreover, the grinding of the samples modifies the proportion of their soluble fraction (Ralet et al 1993).
The sugar profile of the polymers (Table 3) is in agreement with the literature data for inner fibre (Hansen et a1 1992) and pea hulls (Ralet et a1 1993; Auffret et a1 1994; Lien et a1 1994). The so-called wood cellulose contains, in fact, other polysaccharides, as demonstrated by the presence of arabinose and
Composition of pea fibres and efSect on ileal endogenous amino acid flow in pigs 131
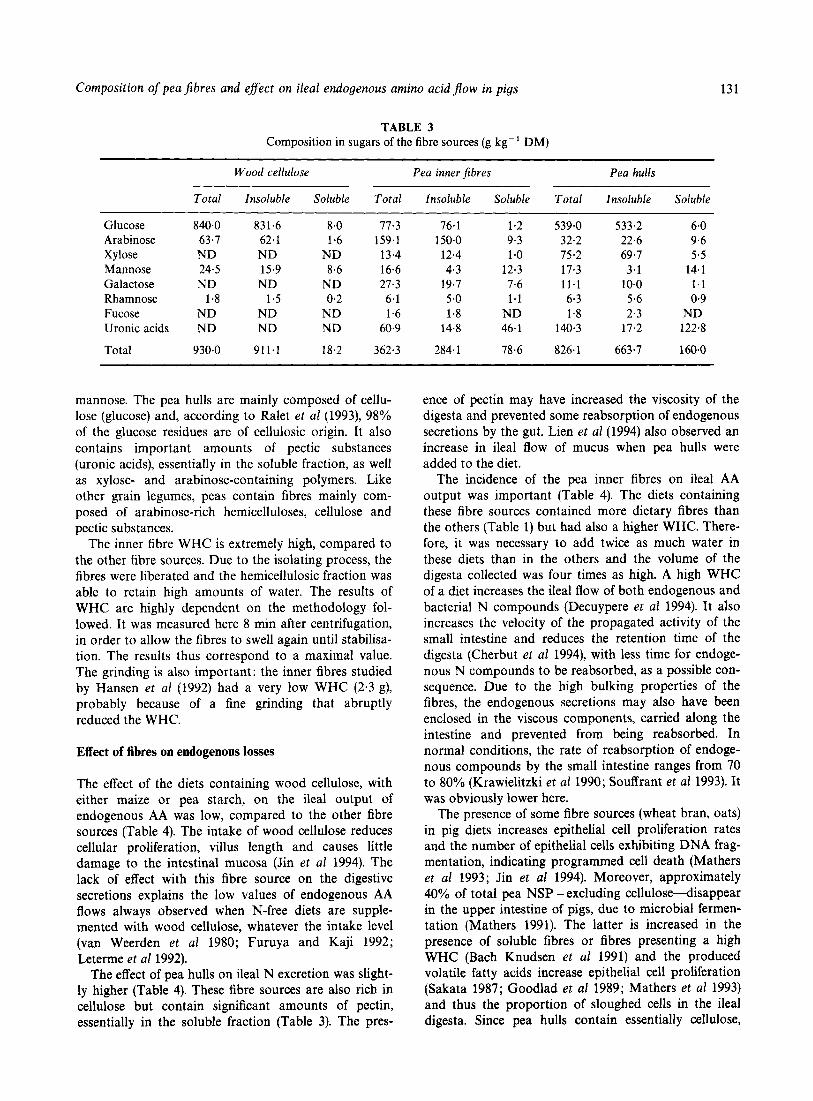
TABLE 3 Composition in sugars of the fibre sources (g kg-' DM)
Wood cellulose Pea inner Jibres Pea hulls
Total Insoluble Soluble Total Insoluble Soluble Total Insoluble Soluble
Glucose Arabinose Xylose Mannose Galactose Rharnnose Fucose Uronic acids Total
840.0 63.7 N D 24.5 ND
ND ND
1.8
930.0
831.6 62- 1 N D 15.9 ND
1.5 ND ND
911.1
8.0 1 *6
N D 8.6
ND 0.2
ND ND
18.2
77-3 159.1 13.4 16.6 27.3 6.1 1.6
60.9
362.3
~~
76- 1 150.0 12.4 4.3
19.7 5.0 1.8
14.8
284.1
~~~
1 *2 9.3 1 .o
12.3 7.6 1.1
N D 46.1
78.6
539.0 32.2 75.2 17.3 11.1 6.3 1.8
140.3
826.1
533.2 22.6 69.7 3.1
10.0 5.6 2.3
17.2
663.7
6.0 9.6 5.5
14.1 1.1 0.9
ND 122.8
160.0
mannose. The pea hulls are mainly composed of cellu- lose (glucose) and, according to Ralet et al (1993), 98% of the glucose residues are of cellulosic origin. It also contains important amounts of pectic substances (uronic acids), essentially in the soluble fraction, as well as xylose- and arabinose-containing polymers. Like other grain legumes, peas contain fibres mainly com- posed of arabinose-rich hemicelluloses, cellulose and pectic substances.
The inner fibre WHC is extremely high, compared to the other fibre sources. Due to the isolating process, the fibres were liberated and the hemicellulosic fraction was able to retain high amounts of water. The results of WHC are highly dependent on the methodology fol- lowed. It was measured here 8 min after centrifugation, in order to allow the fibres to swell again until stabilisa- tion. The results thus correspond to a maximal value. The grinding is also important: the inner fibres studied by Hansen et a1 (1992) had a very low WHC (2.3 g), probably because of a fine grinding that abruptly reduced the WHC.
Effect of fibres on endogenous losses
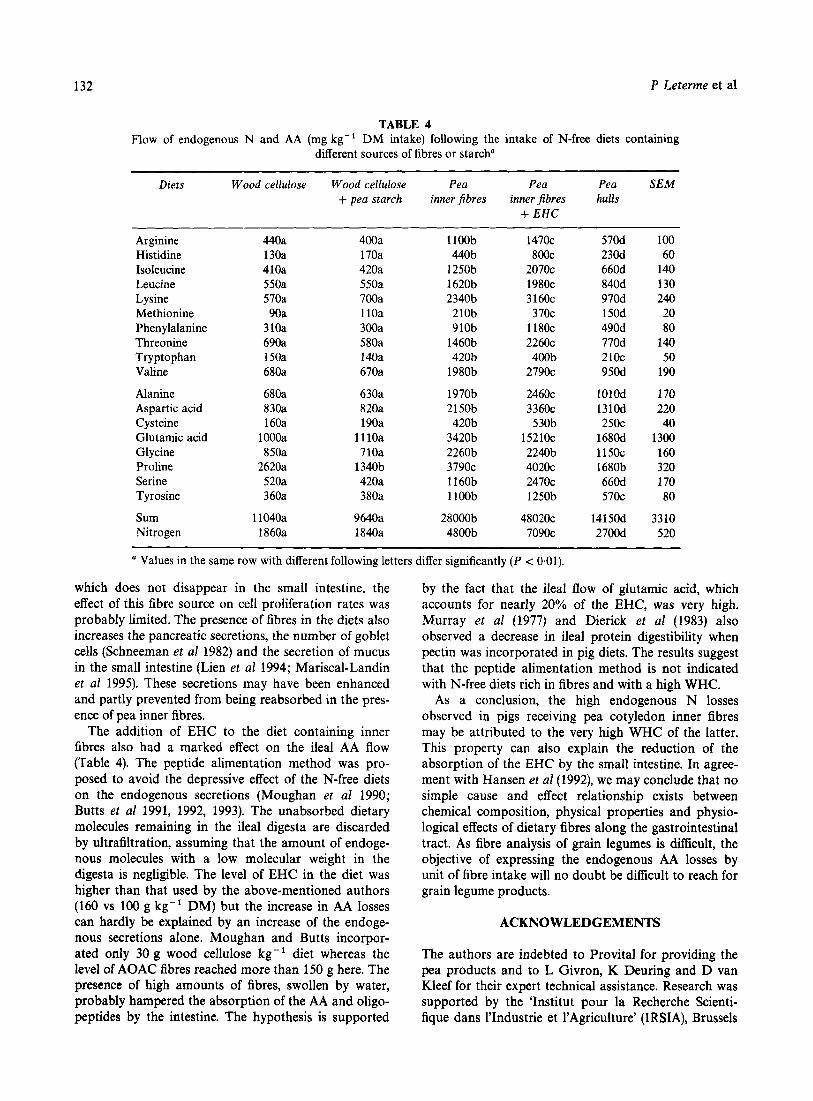
The effect of the diets containing wood cellulose, with either maize or pea starch, on the ileal output of endogenous AA was low, compared to the other fibre sources (Table 4). The intake of wood cellulose reduces cellular proliferation, villus length and causes little damage to the intestinal mucosa (Jin et a1 1994). The lack of effect with this fibre source on the digestive secretions explains the low values of endogenous AA flows always observed when N-free diets are supple- mented with wood cellulose, whatever the intake level (van Weerden et al 1980; Furuya and Kaji 1992; Leterme et a1 1992).
The effect of pea hulls on ileal N excretion was slight- ly higher (Table 4). These fibre sources are also rich in cellulose but contain significant amounts of pectin, essentially in the soluble fraction (Table 3). The pres-
ence of pectin may have increased the viscosity of the digesta and prevented some reabsorption of endogenous secretions by the gut. Lien et a1 (1994) also observed an increase in ileal flow of mucus when pea hulls were added to the diet.
The incidence of the pea inner fibres on ileal AA output was important (Table 4). The diets containing these fibre sources contained more dietary fibres than the others (Table 1) but had also a higher WHC. There- fore, it was necessary to add twice as much water in these diets than in the others and the volume of the digesta collected was four times as high. A high WHC of a diet increases the ileal flow of both endogenous and bacterial N compounds (Decuypere et a1 1994). It also increases the velocity of the propagated activity of the small intestine and reduces the retention time of the digesta (Cherbut et al 1994), with less time for endoge- nous N compounds to be reabsorbed, as a possible con- sequence. Due to the high bulking properties of the fibres, the endogenous secretions may also have been enclosed in the viscous components, carried along the intestine and prevented from being reabsorbed. In normal conditions, the rate of reabsorption of endoge- nous compounds by the small intestine ranges from 70 to 80% (Krawielitzki et a1 1990; Souffrant et a1 1993). It was obviously lower here.
The presence of some fibre sources (wheat bran, oats) in pig diets increases epithelial cell proliferation rates and the number of epithelial cells exhibiting DNA frag- mentation, indicating programmed cell death (Mathers et a1 1993; Jin et a1 1994). Moreover, approximately 40% of total pea NSP-excluding cellulose-disappear in the upper intestine of pigs, due to microbial fermen- tation (Mathers 1991). The latter is increased in the presence of soluble fibres or fibres presenting a high WHC (Bach Knudsen et a1 1991) and the produced volatile fatty acids increase epithelial cell proliferation (Sakata 1987; Goodlad et a1 1989; Mathers et a1 1993) and thus the proportion of sloughed cells in the ileal digesta. Since pea hulls contain essentially cellulose,
132 P Leterme et a1
TABLE 4 Flow of endogenous N and AA (mg kg-' DM intake) following the intake of N-free diets containing
different sources of fibres or starch" ~~ ~
Diets Wood cellulose Wood cellulose Pea Pea Pea SEM + pea starch inner jibres inner fibres hulls
+ EHC
Arginine Histidine Isoleucine Leucine Lysine Methionine Phenylalanine Threonine Tryptophan Valine
Alanine Aspartic acid Cysteine Glutamic acid Gly cine Proline Serine Tyrosine
Sum Nitrogen
440a 130a 410a 550a 570a 90a
310a 690a 150a 680a
680a 830a 160a
1 OOOa 850a
2620a 520a 360a
11040a 1860a
400a 170a 420a 550a 700a 110a 300a 580a 140a 670a
630a 820a 190a
11 10a 710a
1340b 420a 380a
9640a 1840a
1 l00b 440b
1250b 1620b 2340b 210b 910b
1460b 420b
1980b
1970b 2150b 420b
3420b 2260b 3790c 1160b 1 l00b
28000b 4800b
1470c 800c
2070c 1980c 3160c 370c
1 180c 2260c 400b
2790c
2460c 3360c 530b
15210c 2240b 4020c 2470c 1250b
48020c 7090c
570d 230d 660d 840d 970d 150d 490d 770d 210c 950d
lOlOd 1310d 250c
1680d 1150c 1680b 660d 570c
14150d 2700d
100 60
140 130 240 20 80
140 50
190
170 220 40
1300 160 320 170 80
3310 520
a Values in the same row with different following letters differ significantly ( P < 0.01).
which does not disappear in the small intestine, the effect of this fibre source on cell proliferation rates was probably limited. The presence of fibres in the diets also increases the pancreatic secretions, the number of goblet cells (Schneeman et al 1982) and the secretion of mucus in the small intestine (Lien et a1 1994; Mariscal-Landin et a1 1995). These secretions may have been enhanced and partly prevented from being reabsorbed in the pres- ence of pea inner fibres.
The addition of EHC to the diet containing inner fibres also had a marked effect on the ileal AA flow (Table 4). The peptide alimentation method was pro- posed to avoid the depressive effect of the N-free diets on the endogenous secretions (Moughan et a1 1990; Butts et a1 1991, 1992, 1993). The unabsorbed dietary molecules remaining in the ileal digesta are discarded by ultrafiltration, assuming that the amount of endoge- nous molecules with a low molecular weight in the digesta is negligible. The level of EHC in the diet was higher than that used by the above-mentioned authors (160 vs 100 g kg-' DM) but the increase in AA losses can hardly be explained by an increase of the endoge- nous secretions alone. Moughan and Butts incorpor- ated only 30 g wood cellulose kg-' diet whereas the level of AOAC fibres reached more than 150 g here. The presence of high amounts of fibres, swollen by water, probably hampered the absorption of the AA and oligo- peptides by the intestine. The hypothesis is supported
by the fact that the ileal flow of glutamic acid, which accounts for nearly 20% of the EHC, was very high. Murray et a1 (1977) and Dierick et a1 (1983) also observed a decrease in ileal protein digestibility when pectin was incorporated in pig diets. The results suggest that the peptide alimentation method is not indicated with N-free diets rich in fibres and with a high WHC.
As a conclusion, the high endogenous N losses observed in pigs receiving pea cotyledon inner fibres may be attributed to the very high WHC of the latter. This property can also explain the reduction of the absorption of the EHC by the small intestine. In agree- ment with Hansen et a1 (1992), we may conclude that no simple cause and effect relationship exists between chemical composition, physical properties and physio- logical effects of dietary fibres along the gastrointestinal tract. As fibre analysis of grain legumes is difficult, the objective of expressing the endogenous AA losses by unit of fibre intake will no doubt be difficult to reach for grain legume products.
ACKNOWLEDGEMENTS
The authors are indebted to Provital for providing the pea products and to L Givron, K Deuring and D van Kleef for their expert technical assistance. Research was supported by the 'Institut pour la Recherche Scienti- fique dans l'Industrie et 1'Agriculture' (IRSIA), Brussels

Composition of pea fibres and effect on ileal endogenous amino acid pow in pigs 133
(contract no 5460) and the TNO-ILOB, Wageningen, The Netherlands. For his stay in Wageningen, P Leterme received a short-term training grant from the European Union (GT 920155).
REFERENCES
Auffret A, Guillon F, Barry J, Ralet M, Thibault J 1994 Effect of grinding and experimental conditions on the measure- ment of hydration properties of dietary fibres. Lebensm Wiss Techno1 12 247-251.
Bach Knudsen K, Borg Jensen B, Andersen J, Hansen I 1991 Gastrointestinal implications in pigs of wheat and oat frac- tions. 2. Microbial activity in the gastrointestinal tract. Br J Nutr 65 233-248.
Butts C, Moughan P, Smith W 1991 Endogenous amino acid flow at the terminal ileum of the rat determined under con- ditions of peptide alimentation. J Sci Food Agric 55 175- 187.
Butts C, Moughan P, Smith W 1992 Protein nitrogen, peptide nitrogen and free amino acid nitrogen in endogenous digesta nitrogen at the terminal ileum of the rat. J Sci Food Agric 59 291-298.
Butts C, Moughan P, Smith W, Carr D 1993 Endogenous lysine and other amino acid flows at the terminal ileum of the growing pig (20 kg bodyweight): the effect of protein- free, synthetic amino acid, peptide and protein alimentation. J Sci Food Agric 61 31-40.
Cherbut C, Bruley des Varannes S, Schnee M, Rival M, Bamiche J, Delort-Lava1 J 1994 Involvment of small intesti- nal motility in blood glucose response to dietary fibre in man. Br J Nutr 71 675-685.
Costa N, Low G, Walker A, Owen R, Englyst H 1994 Effect of baked on steroid metabolism and non-starch poly- saccharide output of hypercholesteroleamic pigs with or without an ileo-rectal anastomosis. Br J Nutr 71 871-886.
Decuypere J, Spriet S, van Gils L 1994 Influence of the water- holding capacity (WHC) of the feed on the precaecal and fecal apparent digestibility in pigs. In: Proc VIth Int Symp on Digestive Physiology in Pigs, eds Souffrant W & Hage- meister H. EAAP Publ no 80, pp 125-128.
Dierick N, Vervaeke I, Decuypere J, Henderickx H 1983 Influ- ence de la nature et du niveau des fibres brutes sur la digest- ibilite ileale et fecale apparente de la matriere seche, des proteines et des acides amines et sur la retentions azotee chez les porcs. Rev Agric (Brussels) 36 1691-1711.
Englyst H, Cummings J 1988 Improved method for measure- ment of dietary fibre as non-starch polysaccharides in plant
Englyst H, Quigley M, Hudson G, Cummings J 1992 Deter- mination of dietary fibre as non-starch polysaccharides by gas-liquid chromatography. Analyst 117 1707-1714.
Franqois E, Thill N, Thewis A 1978 Methode rapide de dosage de l’oxyde de chrome dans les aliments, les fkces et les contenus digestifs par titrage apres oxydation nitro- perchlorique. Ann Zootech 27 355-361.
Furuya S, Kaji Y 1992 The effects of feed intake and purified cellulose on the endogenous ileal amino acid flow in growing pigs. Br J Nutr 68 463-472.
Geil P, Anderson J 1994 Nutrition and health implications of dry beans: a review. J Am College Nutr 13 549-558.
Giger S, Thivend P, Sauvant D, Dorleans M, Hournaix P 1987 Etude de I’influence prealable de differentes enzymes amylolytiques sur la teneur en residu NDF d’aliments du betail. Ann Zootech 36 39-48.
Goodlad J, Mathers J 1990 Large bowel fermentation in rats given diets containing raw peas. Br J Nutr 64 569-587.
foods. J AOAC 71 808-814.
Goodlad J, Mathers J 1992 Digestion of complex carbo- hydrates and large bowel fermentation in rats fed on raw and cooked peas. Br J Nutr 67 475-488.
Goodlad R, Ratcliffe B, Fordham J, Wright N 1989 Does dietary fibre stimulate intestinal epithelial cell proliferation in germ-free rats. Gut 30 820-825.
Hansen I, Bach Knudsen K, Eggum B 1992 Gastrointestinal implications in the rat of wheat bran, oat bran and pea fibre. Br J Nutr 68 451-462.
Huisman J, Heinz T, Poel A van der, Leeuwen P van, Souf- frant W, Verstegen M 1992 True ileal digestibility and amounts of endogenous protein measured with the I5N- dilution technique in piglets fed on peas (Pisum sativum) and common beans (Phaseolus vulgaris). Br J Nutr 68 101-110.
Jin L, Reynolds L, Redmer D, Caton J, Crenshaw J 1994 Effects of dietary fibre on intestinal growth, cell prolifer- ation and morphology in growing pigs. J Anim Sci 72 2270- 2278.
Krawielitzki K, Zebrowska T, Schadereit R, Kowalczyk J, Wiinsche J, Hermann U 1990 Determining of nitrogen absorption and nitrogen secretion in different sections of the pig’s intestine by digesta exchange between I5N-labelled and unlabelled animals. Arch Tierernahrung 40 25-37.
Leeuwen P van, Kleef D van, Kempen G van, Huisman J, Verstegen M 1991 The post-valve T-caecum cannulation technique: an alternative method for chyme collection in pigs. J Anim Physiol Anim Nutr 65 183-193.
Leterme P, Monmart M 1990 Importance du tryptophane en alimentation et son dosage dans les aliments. Bull Rech Agron (Gembloux) 25 29-344.
Leterme P, Pirard L, Thtwis A 1992 A note on the effect of wood cellulose level in protein-free diets on the recovery and amino acid composition of endogenous protein col- lected from the ileum in pigs. Anim Prod 54 163-165.
Lien K, Sauer W, McBurney M 1994 The effect of pea fibre addition on ileal and fecal amino acid digestibilities, carbo- hydrate recoveries and ileal mucus output. In: Proc VIth Int Symp on Digestive Physiology in Pigs, eds Souffrant W & Hagemeister H. EAAP Publ, pp 75-78.
MacKenzie S, Tenaschuk D 1979a Quantitative formation of N (0, S)-heptafluorobutyryl isobutyl amino acids for gas chromatographic analyses. I. Esterification. J Chromatogr
MacKenzie S, Tenaschuk D 1979b Quantitative formation of N (0, S)-heptafluorobutyryl isobutyl amino acids for gas chromatographic analyses. 11. Acylation. J Chromatogr 173
Mariscal-Landin G, S h e B, Colleaux Y, Lebreton Y 1995 Endogenous amino nitrogen collected from pigs with end-to-end ileorectal anastomosis is affected by the method of estimation and altered by dietary fibre. J Nutr 125 136- 146.
Mathers J 1991 Digestion of non-starch polysaccharides by non-ruminant omnivores. Proc Nutr Soc 50 161-172.
Mathers J, Kennard J, James 0 1993 Gastrointestinal responses to oats consumption in young adult and elderly rats : digestion, large bowel fermentation and crypt cell pro- liferation rates. Br J Nutr 70 567-584.
Mongeau R, Brassard R 1994 Comparison and assessment of the difference in total dietary fibre in cooked dried legumes as determined by five methods. J AOAC 77 1197-1202.
Moughan P, Darragh A, Smith W, Butts C 1990 Perchloric and trichloroacetic acids as precipitants of protein in endogenous ileal digesta from the rat. J Sci Food Agric 52
Murray A, Fuller M, Pine A 1977 The effect of fibre in the form of various polysaccharides on the apparent digest- ibility of protein in the pig. Anim Prod 24 139-144.
171 195-208.
53-63.
13-21.

134 P Leterme et a1
Periago M, Ros G, Englyst H, Rincon F 1994 Variation in dietary fibre content of peas as function of variety, size and analytical method. Reuista Espariola de Ciencia y Tecnologia de Alimentos 34 565-575.
Prosky L, Asp N, Schweizer T, De Vries J, Furda I 1988 Determination of insoluble, soluble and total dietary fibre in foods and food products. J AOAC 71 1017-1023.
Prosky L, Asp N, Schweizer T, De Vries J, Furda I, Lee S 1994 Determination of soluble dietary fibre in foods and food products: collaborative study. J AOAC 77 690-694.
Ralet M, Della Valle G, Thibault J 1993 Raw and extruded fibre from pea hulls. Part I: Composition and physico- chemical properties. Carbohydr Polym 20 17-23.
Sakata T 1987 Stimulatory effect of short-chain fatty acids on epithelial cell proliferation in the rat intestine: possible explanation for the trophic effects of fermentable fibre, gut microbes and luminal trophic factors. Br J Nutr 58 95-103.
Sandstrom B, Hansen L, Ssrensen A 1994 Pea fibre lowers fasting and postprandial blood triglyceride concentrations in humans. J Nutr 124 2386-2396.
Schneeman B, Richter D, Jacobs L 1982 Response to dietary wheat bran in the exocrine pancreas and intestine of rats. J Nutr 112 283-290.
Schulze H, Leeuwen P van, Verstegen M, Huisman J, Souf- frant W, Ahrens F 1994 Effects of level of dietary neutral detergent fibre on ileal apparent digestibility and ileal nitro- gen losses in pigs. J Anim Sci 72 2362-2368.
Schulze H, Leeuwen P van, Verstegen M, van den Berg J 1995 Dietary level of neutral detergent fibre and ileal endogenous nitrogen flow in pigs. J h i m Sci 73 441-448.
Scott R 1979 Colorimetric determination of hexuronic acids in plant materials. Anal Chem 31 936-941.
Slump P, Schreuder H 1969 Determination of tryptophan in foods. Anal Biochem 27 182-185.
Soest J van, Wine R 1967 Use of detergents in the analysis of fibrous feeds. Determination of plant cell-wall constituents. J AOAC 50 50-55.
Souffrant W, Rtrat A, Laplace J P, Darcy-Vrillon B, Kohler R, Corring T, Gebhardt G 1993 Exogenous and endoge- nous contributions to nitrogen fluxes in the digestive tract of pigs fed a casein diet. 111. Recycling of endogenous nitro- gen. Reprod Nutr Divelop 33 373-382.
Weerden E van, Slump P, Huisman J 1980 Amino acid digestion in different parts of the intestinal tract of pigs. In: Proc 3rd EAAP-Symposium on Protein and Metabolism and Nutrition, eds Oslage H & Rohr K. EAAP Publ, pp 207- 214.
Wolters G, Verbeek C, van Westerop J, Hermus R, Voragen A 1992 Comparison of different methods for determination of dietary fibre. J AOAC 75 626-634.
Zhao X, Jsrgensen H, Eggum B 1995 The influence of dietary fibre on body composition, visceral organ weight, digest- ibility and energy balance in rats housed in different thermal environments. Br J Nutr 73 687-699.












![PEA-RP250GA PEA-RP400GA PEA-RP500GA - …H]-RP/2010-2009/... · PEA-RP250GA PEA-RP400GA PEA-RP500GA ... Cautions for units utilising refrigerant R410A ... It is also possible to attach](https://img.pdfslide.us/doc/110x75/5ad5679d7f8b9a075a8cd92b/pea-rp250ga-pea-rp400ga-pea-rp500ga-h-rp2010-2009pea-rp250ga-pea-rp400ga.jpg)
















