Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Dec. 2009, p. 7296–7305 Vol. 191, No. 230021-9193/09/$12.00 doi:10.1128/JB.00882-09Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Characterization of the Sporulation Initiation Pathway ofClostridium difficile and Its Role in Toxin Production�†
Sarah Underwood,1 Shuang Guan,1 Vinod Vijayasubhash,1 Simon D. Baines,1,2 Luke Graham,3Richard J. Lewis,3 Mark H. Wilcox,1,2 and Keith Stephenson1*
Institute of Molecular and Cellular Biology, Faculty of Biological Sciences, Leeds University, Leeds LS2 9JT, United Kingdom1;Department of Microbiology, Leeds General Infirmary, Leeds LS1 3EX, United Kingdom2; and Institute for Cell and
Molecular Biosciences, Faculty of Medical Sciences, Newcastle University, Newcastle upon Tyne NE2 4HH, United Kingdom3
Received 7 July 2009/Accepted 17 September 2009
Clostridium difficile is responsible for significant mortality and morbidity in the hospitalized elderly. C.difficile spores are infectious and are a major factor contributing to nosocomial transmission. The Spo0Aresponse regulator is the master regulator for sporulation initiation and can influence many other cellularprocesses. Using the ClosTron gene knockout system, we inactivated genes encoding Spo0A and a putativesporulation-associated sensor histidine kinase in C. difficile. Inactivation of spo0A resulted in an asporogeneousphenotype, whereas inactivation of the kinase reduced C. difficle sporulation capacity by 3.5-fold, suggestingthat this kinase also has a role in sporulation initiation. Furthermore, inactivation of either spo0A or the kinaseresulted in a marked defect in C. difficile toxin production. Therefore, Spo0A and the signaling pathway thatmodulates its activity appear to be involved in regulation of toxin synthesis in C. difficile. In addition, Spo0Awas directly phosphorylated by a putative sporulation-associated kinase, supporting the hypothesis thatsporulation initiation in C. difficile is controlled by a two-component signal transduction system rather than amulticomponent phosphorelay. The implications of these findings for C. difficile sporulation, virulence, andtransmission are discussed.
Clostridium difficile is a major nosocomial enteropathogenand is the primary cause of infectious hospital acquired diar-rhea, mainly occurring in patients with a prior history of anti-biotic therapy (9). The virulence of C. difficile is generallyattributed to the production of toxins, which cause colonicdamage and inflammation, leading to colitis and diarrhea.Transmission of C. difficile in health care institutions is medi-ated primarily by spores (31). Spores are infective and highlyresistant and facilitate C. difficile persistence in aerobic envi-ronments outside of the host. Spores may also contribute tosurvival of C. difficile inside the host during antimicrobial ther-apy and subsequent recrudescence following the cessation oftherapy (42). Interestingly, epidemic strains of C. difficile, suchas ribotypes 001 and 027 (NAP1/BI), have an increased inher-ent sporulation capacity, indicating that this may be a factor intheir spread (1, 10).
Generally, sporulation only occurs when efforts to maintainvegetative growth have failed. The signal transduction pathwaythat controls sporulation initiation has been most extensivelystudied in the genus Bacillus and is composed of an expandedvariant of a two-component signal transduction system (TCS)known as the sporulation phosphorelay (6) (see Fig. S1 in thesupplemental material). Environmental and cellular signalsthat indicate that vegetative growth is no longer possible aresensed by sporulation-associated sensor histidine kinases, re-
sulting in autophosphorylation of a specific histidine in thecatalytic domain. The phosphoryl group is transferred to anaspartate on the Spo0F response regulator and is subsequentlyrelayed to the Spo0A response regulator transcription factorby the Spo0B phosphotransferase, which is transiently phos-phorylated on a histidine. Phosphorylation of the active siteaspartate of Spo0A promotes binding to a specific target se-quence (the “0A box”) in or near the promoters of genes underSpo0A control, resulting in gene activation or repression (34).
Sensor kinases are responsible for activation of TCSs andphosphorelays via the input of phosphoryl groups. Sensor ki-nases have a modular domain organization with an N-terminalsignal input domain and a catalytic C-terminal kinase domaincontaining the dimerization and histidine phosphotransfer(DHpt) subdomain and an ATP binding subdomain. In thesporulation phosphorelay, multiple sensor kinases phosphory-late the same response regulator substrate, Spo0F (35). Forexample, B. subtilis has five sensor kinases, KinA to KinE,which have highly conserved active sites (Fig. 1) and are capa-ble of influencing sporulation (17). Generally, one sensor ki-nase is responsive to one specific signal ligand; thus, havingseveral kinases increases signal sensing diversity and allowsmultiple signals to influence Spo0A activation.
Spo0A is highly conserved in Bacillus and Clostridium spe-cies, and 0A boxes can be identified upstream of genes likely tobe controlled by Spo0A in clostridia (such as spo0A itself),suggesting that the mechanism of Spo0A-mediated gene reg-ulation is also the same. Furthermore, the key residues ofSpo0A known to mediate the interaction with the bases of the0A box are highly conserved in Bacillus and Clostridium species(47). Despite these facts, true Spo0F and Spo0B homologuesdo not appear to be encoded within the genomes of sequenced
* Corresponding author. Mailing address: Institute of Molecularand Cellular Biology, Faculty of Biological Sciences, Leeds University,Leeds LS2 9JT, United Kingdom. Phone: 44(0)113 3435581. Fax:44(0)113 3435638. E-mail: [email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
� Published ahead of print on 25 September 2009.
7296
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

Clostridium species, at least on the basis of protein sequencehomology (35). The generally accepted hypothesis is that inClostridium species the sporulation initiation pathway has notevolved into a multicomponent phosphorelay but has remainedmore like a TCS in which signals are sensed by sporulation-associated sensor kinases that phosphorylate Spo0A directly(39, 45).
Once activated, Spo0A controls many post-exponential-phase phenomena. Spore-forming bacterial pathogens haveevolved to utilize Spo0A to modulate the virulence and survivalresponses that are essential for their pathogenic lifestyle (39).For example, Spo0A is involved in both sporulation and toxinproduction in Clostridium perfringens, presumably by binding tothe putative 0A box upstream from the cpe cytotoxin gene (15).In C. difficile there is conflicting evidence for the existence offirm regulatory links between sporulation and toxin produc-tion. Several reports describing positive correlations betweensporulation, stationary-phase events, and toxin production (8,18, 20) are contradicted by those showing negative correlationsor no link at all (2, 19). This confusing picture has been com-pounded by differences in the strains and the growth mediaused in the studies and by the paucity of C. difficile gene
inactivation systems to construct defined mutants in key genesthat would allow the dissection of the pathways involved. In thepresent study we have used the recently developed ClosTrongene knockout system (14) to inactivate the genes encodingSpo0A and a putative sporulation-associated sensor kinase toinvestigate their involvement in sporulation and toxin produc-tion in C. difficile.
MATERIALS AND METHODS
Bacterial strains, plasmids, DNA manipulation, and oligonucleotide primers.To allow selection of ClosTron mutants the erythromycin-sensitive C. difficilestrain 630 �erm (16) was used. This strain is a spontaneously cured derivative ofthe genome sequencing strain 630, which is a virulent and multidrug-resistantisolate from an outbreak in Switzerland (32). Escherichia coli DH5� was used asthe host for cloning steps and E. coli Rosetta (DE3; Novagen) was used as thehost for overexpression and protein purification. Conjugative transfer of plas-mids to C. difficile was carried out with E. coli CA434 (27) as the donor strain.The ClosTron delivery vector pMTL007 (14) was used as the parental plasmidfor the construction of pMTL007::Cdi-spo0A-178a and pMTL007::Cdi-CD2492-254a, which retarget the group II intron of pMTL007 to spo0A and CD2492 ofC. difficile. pGEX-6PI (GE Healthcare Life Sciences) was used as the vector forthe expression and purification of proteins as GST fusions. All DNA manipula-tions were carried out according to standard molecular biological protocols (30).
FIG. 1. DHpt subdomains of the B. subtilis sporulation-associated sensor kinases and the orphan sensor kinases of C. difficile. (A) Alignmentof the amino acid sequences of the active site regions of the B. subtilis sporulation-associated sensor kinases, the orphan sensor kinases of C.difficile, and HK PPA0945 from Propionibacterium acnes. The active sites of the B. subtilis kinases have been described previously (35), and theactive sites of the C. difficile orphan kinases and HK PPA0945 were predicted by using Pfam (http://pfam.sanger.ac.uk/). Amino acids that areidentical in the kinases of each organism are shaded. (B) Alignment of the amino acid sequences of the DHpt subdomains of the C. difficile orphansensor kinases and HK PPA0945. Amino acids that are known from structural studies (46) or from covariance analysis (33, 44) to be involved ininteraction (*) and/or specificity of interaction (†) with the cognate response regulator are indicated. Putative specificity-determining amino acidsthat are strictly conserved in HK CD1492, HK CD1579, and HK CD2492 but different in HK CD1352 and HK CD1949 are shaded. In all cases,the known or likely phosphorylatable histidine residue is underlined and all amino acid sequences are shown in the N-to-C direction. Alpha-1 andalpha-2 represent the �-helices of one subunit of the DHpt subdomain that form the phosphotransfer four-helix bundle on kinase dimerization.
VOL. 191, 2009 CLOSTRIDIUM DIFFICILE SPORULATION AND TOXIN PRODUCTION 7297
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

The sequences of the primers used in the present study are shown in Table S1 inthe supplemental material.
Cultivation of bacterial strains. Liquid cultures of C. difficile were grown inbrain heart infusion (BHI; Oxoid) broth or Schaedlers anaerobic broth (SAB;Oxoid). Solid C. difficile cultures were grown on fresh blood agar (FBA) or BHIagar. In all cases, C. difficile was grown at 37°C in an anaerobic environment(80% N2, 10% CO2, and 10% H2 [vol/vol]). E. coli was grown aerobically inLuria-Bertani (LB) broth or on LB agar at 37°C. For conjugative transfer to C.difficile, E. coli was grown anaerobically on FBA. Unless stated otherwise, thefollowing antibiotics were included in liquid and solid media as required; cy-closerine (250 �g/ml), cefoxitin (8 �g/ml), chloramphenicol (12.5 �g/ml), thia-mphenicol (15 �g/ml), and erythromycin (5 �g/ml).
Construction of C. difficile mutants by using the ClosTron system. TheClosTron system was used as described previously (14), in conjunction with thecommercially available TargeTron gene knockout system kit (Sigma-Aldrich).Briefly, the TargeTron design software was used to design primers to retarget thegroup II intron on pMTL007 to spo0A (primers Cd-spo0A-178a-IBS, Cd-spo0A-178a-EBS1d, and Cd-spo0A-178a-EBS2; see Table S1 in the supplemental ma-terial) and CD2492 (primers IBS_CD1A, EBS1d_CD1A, and EBS2_CD1A; seeTable S1 in the supplemental material). These primer sets were used with theEBS universal primer (see Table S1 in the supplemental material) and the introntemplate DNA provided with the TargeTron kit to generate a retargeted intronDNA fragment for each gene by overlap PCR according to the manufacturer’sinstructions. The two resultant DNA fragments (353 bp each) were cloned intothe HindIII and BsrGI restriction sites of pMTL007 using E. coli DH5� as thehost to produce pMTL007::Cdi-spo0A-178a and pMTL007::Cdi-CD2492-254a.The fidelity of the cloned inserts was verified by DNA sequencing.
Retargeted plasmids were transformed into E. coli CA434 using chloramphen-icol as the selective marker and subsequently mated with C. difficile 630 �erm onFBA. C. difficile transconjugant colonies were selected by subculturing on BHIagar containing cycloserine, cefoxitin, and thiamphenicol. To make the geneknockouts, C. difficile transconjugant colonies were grown in SAB, and theintegration of the group II intron RNA into the spo0A and CD2492 genes wasinduced by the addition of 1 mM IPTG (isopropyl-�-D-thiogalactopyranoside),followed by plating onto BHI agar containing erythromycin. Erythromycin-resis-tant (and thiamphenicol-sensitive) C. difficile colonies are produced, followingplasmid loss and after insertion of the group II intron into the chromosome,which is accompanied by splicing out of the td group I intron from the ermBretrotransposition-activated marker (RAM).
Extraction of genomic DNA from C. difficile. Cultures of C. difficile were grownovernight in BHI broth containing antibiotics as required. The cells from 1.5 mlof culture were harvested by microcentrifugation, and the cell pellets wereresuspended in 500 �l of solution I (50 mM Tris-HCl [pH 7.5], 20 mM NaCl, 2mg of lysozyme/ml) and incubated at 37°C for 30 min. Then, 20 �l of proteinaseK solution (20 mg/ml) and 350 �l of 2% (wt/vol) sodium dodecyl sulfate (SDS)were added to each sample, followed by incubation at 60°C for 2 h. The DNA wasextracted using phenol-chloroform, precipitated with ethanol, and washed with70% (vol/vol) ethanol. Precipitated DNA pellets were dissolved in 50 �l ofdistilled water, and contaminating RNA was removed by the addition of 1.25 �lof RNase (10 mg/ml), followed by incubation at 37°C for 30 min.
Sporulation assays. C. difficile sporulation was measured by direct microscopiccounting of vegetative cells and spores as described previously (10). Overnightcultures of C. difficile were grown in SAB and used to inoculate BHI broth to anoptical density at 550 nm (OD550) of 0.1. The cultures were incubated for 72 h,after which time samples were removed, and the cells or spores in each samplewere pelleted by microcentrifugation. Cell or spore pellets were resuspended inphosphate-buffered saline (PBS), spread evenly over microscope slides, and airdried. The slides were stained with malachite green and counterstained withcarbol fuchsin over boiling water to discriminate between spores and vegetativecells, respectively. C. difficile sporulation capacity was determined by counting atleast five fields of 100 cells or spores per slide using a light microscope at �1,000magnification, and the number of spores was expressed as a percentage of thetotal number of cells.
SDS-PAGE, TCA precipitation, and Western blotting of proteins. Proteins inC. difficile culture supernatants or cell extracts were separated by SDS-poly-acrylamide gel electrophoresis (PAGE) on a Mini-Protean II electrophoresis cell(Bio-Rad) using 7% (wt/vol) acrylamide gels. To obtain the samples for SDS-PAGE, 1-ml volumes of C. difficile cultures were removed, and the cells andsupernatants were collected after microcentrifugation. Supernatant sampleswere loaded without further manipulation. Cell extracts representing total intra-cellular proteins were prepared by resuspending the cell pellets in 100 �l of lysisbuffer (50 mM Tris-HCl [pH 7.5], 2 mg of lysozyme/ml, 2 mg of DNase I/ml),followed by incubation at 37°C for 30 to 45 min. The samples were boiled for 3
min, and then 900 �l of 50 mM Tris-HCl (pH 7.5) was added. Where required,proteins in C. difficile culture supernatants (1 ml) were precipitated by theaddition of 500 �l of 50% (wt/vol) trichloroacetic acid (TCA; Sigma), followedby incubation on ice for 60 min. Precipitated proteins were pelleted by micro-centrifugation, washed twice with acetone, and resuspended in 100 �l of 20 mMTris-HCl (pH 7.5). For all samples, prior to gel loading, 5� SDS-PAGE samplebuffer (100 mM Tris-HCl [pH 6.8], 5% [vol/vol] �-mercaptoethanol, 4% [wt/vol]SDS, 20% [vol/vol] glycerol, 0.2% bromophenol blue) was added, and the sam-ples were boiled for 3 min. Where required, the volumes loaded onto the gelswere corrected to take into account minor differences in culture ODs. Separatedproteins in gels were stained with Coomassie blue as required.
To detect C. difficile toxin A in culture supernatants and cell extracts, proteinswere separated by SDS-PAGE and transferred to nitrocellulose membranes byusing a Mini-Transblot blotting cell (Bio-Rad). Membranes were blocked inWestern blotting buffer (PBS, 0.1% Tween 20) containing 5% (wt/vol) skim milkand then incubated overnight at room temperature with the primary rabbitanti-toxin A polyclonal antibody (Fitzgerald Industries). After extensive washingin blotting buffer, membranes were incubated for 2 h at room temperature witha secondary goat anti-rabbit peroxidase-conjugated antibody (Jackson Immu-noresearch). The secondary antibody was removed by extensive washing in blot-ting buffer, and the immobilized toxin A on the membranes was detected by usingthe SuperSignal West Pico chemiluminescent substrate (Pierce) according to themanufacturer’s instructions, followed by exposure to X-ray film, scanning, andqualitative image analysis.
C. difficile cytotoxin assay. C. difficile cytotoxin titers (i.e., total toxins A and B)in culture supernatants were determined by using a Vero cell cytotoxicity assayas described previously (5, 12). Briefly, replicates of C. difficile culture superna-tants were 10-fold serially diluted in sterile PBS and applied to Vero cell mono-layers in 96-well microtiter plates. Plates were incubated at 37°C in a 5% (vol/vol)CO2 environment for 48 h and examined by using an inverted microscope.Positive toxin reactions were indicated by the characteristic rounding of Verocells accompanied by parallel neutralization of cytotoxicity with Clostridiumsordellii antitoxin (Prolab Diagnostics). Total cytotoxin titers in C. difficile culturesupernatants were expressed as log10 relative units (RU), where 1 RU represents�80% cell rounding by the undiluted sample, 2 RU represents �80% cellrounding by the 10�1 dilution of the sample, and so on.
Purification of proteins encoded by the C. difficile spo0A and CD1579 genes.DNA fragments encoding full-length products of the spo0A and CD1579 geneswere generated by PCR from C. difficile 630 genomic DNA using primer pairsCDSpo0A_For/CDSpo0A_Rev and CD3B_For/CD3B_Rev, respectively (seeTable S1 in the supplemental material). The resulting DNA fragments wereligated into the BamHI and XhoI restriction sites of pGEX-6P1 to produce genesencoding fusion proteins with glutathione S-transferase (GST) at their N termini.The fidelity of the process was confirmed by DNA sequencing. Recombinantproteins were overexpressed in E. coli Rosetta (DE3) cells grown at 37°C in LBbroth supplemented with ampicillin (270 �M) and chloramphenicol (105 �M).Expression was induced by the addition of 1 mM IPTG at an OD550 of 0.5.Incubation was continued for 4 h, and the cells were harvested by centrifugationat 4,500 � g for 20 min.
Cells were resuspended in 20 ml of buffer A (20 mM Tris-HCl [pH 7.5], 200mM NaCl) and disrupted by sonication. GST-tagged proteins were purified fromcleared lysates by using GST-Sepharose columns (GE Healthcare Life Sciences).Briefly, cell lysates were applied to columns pre-equilibrated with three columnvolumes of buffer A. Columns were washed with five column volumes of bufferA, and proteins were eluted with the same buffer containing 50 mM reducedglutathione. Fractions containing the desired proteins were identified by usingSDS-PAGE, pooled, and then treated with 80 �g of C3 protease at 4°C for 14 hto cleave the GST tag. The proteins were concentrated by using Centriconultrafiltration devices (Millipore) of appropriate molecular weight cutoff andseparated by size exclusion chromatography using Superdex S75 or S200 (GEHealthcare Life Sciences) for Spo0A or CD1579, respectively. Proteins wereconcentrated to �2 mg/ml as before and stored at �80°C until required.
Biochemical assay of sensor histidine kinase and phosphotransfer activity.The autophosphorylation activity of sensor kinases and phosphotransfer toSpo0A were assayed as described previously (21, 40). Reactions (20 �l) werecarried out in 50 mM EPPS buffer (Sigma) containing 20 mM MgCl2, 0.1 mMEDTA, and 5% (vol/vol) glycerol. Autophosphorylation reactions contained 1�M kinase and, where required, Spo0A was included at a final concentration of10 �M to measure the phosphotransfer. In all cases, the reactions were initiatedby the addition of 100 �M ATP containing 50 �Ci of [�-32P]ATP (Perkin-Elmer). At various time intervals, reactions were terminated by the addition of5� SDS-PAGE sample buffer on dry ice. Labeled protein samples were sepa-
7298 UNDERWOOD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

rated by SDS-PAGE on a 10% (wt/vol) acrylamide gel and visualized by phos-phorimaging using a Fuji BAS1000 phosphorimager.
RESULTS
Bioinformatic identification of putative sporulation-associ-ated sensor histidine kinases of C. difficile. In Bacillus speciessporulation-associated phosphorelay sensor kinases have cer-tain characteristics that distinguish them from those of TCSs(39). Unlike most TCSs, the genes encoding sporulation-asso-ciated kinases are chromosomally located as orphans, i.e., theyare not adjacent to a gene encoding their cognate responseregulator. Furthermore, since all of the sporulation kinaseshave the same substrate, Spo0F, they share a high degree ofsequence conservation around the active site histidine and inthe other regions of the DHpt subdomain that interact withSpo0F and/or determine specificity (Fig. 1). Using these fea-tures, it is possible to identify potential sporulation-associatedsensor kinases within the genome sequences of spore-formingbacteria (35).
The genome of C. difficile 630 (32) encodes 50 sensor kinasesand 51 response regulators that are organized into 45 TCSs,with five orphan kinases and six orphan response regulatorsscattered around the chromosome. The orphan kinases areencoded by the CD1352, CD1492, CD1579, CD1949, andCD2492 genes. From here on, sensor histidine kinase proteinsencoded by the relevant genes will be denoted by the prefix HKfollowed by the locus tag.
Sporulation-associated sensor kinases of B. subtilis havehighly variable signal input domains but are generally wellconserved (39 to 50% amino acid identity) in their catalyticdomains. Similarly, HK CD1492, HK CD1579, and HKCD2492 share 39 to 44% amino acid identity in their catalyticdomains, but the signal input domains are variable in se-quence. In addition, in a phylogenetic analysis of all C. difficile630 sensor kinases HK CD1492, HK CD1579, and HK CD2492group together in a single cluster, whereas the other orphankinases fall into separate and unrelated clusters (Y.-F. Lin andK. Stephenson, unpublished data). The DHpt subdomains, in-cluding the active sites of the orphan kinases, were identifiedby using the Pfam database (http://pfam.sanger.ac.uk/) (Fig. 1).The active sites of HK CD1492, HK CD1579, and HK CD2492share a high degree of sequence identity, which is at a levelcomparable to that shared by B. subtilis KinA to KinE (Fig. 1).The active sites of HK CD1352 and HK CD1949 do not exhibitgood homology to each other or to the other C. difficile orphankinases. Furthermore, a number of the amino acids that definethe interaction surface with the response regulator and/or areinvolved in specificity of the interaction (33, 44, 46) are strictlyconserved in HK CD1492, HK CD1579, and HK CD2492 butare different in HK CD1352 and HK CD1949 (Fig. 1). Takentogether, this suggests that HK CD1492, HK CD1579, and HKCD2492 may be able to phosphorylatie the same responseregulator substrate, Spo0A, and are therefore potentially in-volved in C. difficile sporulation initiation.
HK CD1492 and HK CD2492 are putative integral mem-brane proteins, whereas HK CD1579 is likely to be a cytosolickinase (see Fig. S2 in the supplemental material). Domainswith recognized signaling functions were not detected in HKCD1492 and HK CD2492 using Pfam. HK CD1579 was pre-
dicted to contain a PAS domain in its signal input domain (seeFig. S2 in the supplemental material). A PAS domain at the Nterminus of B. subtilis KinA has a critical and indispensablerole in the activity of the kinase (38, 43).
Inactivation of sporulation-associated genes of C. difficileusing the ClosTron system. We used the ClosTron system toconstruct a knockout mutant in which the gene encoding HKCD2492 was inactivated by insertion of the group II intronfrom pMTL007. In parallel, we also constructed a mutant inwhich the spo0A gene had been inactivated as described pre-viously (14). The general gene inactivation process is repre-sented schematically in Fig. 2.
pMTL007::Cdi-spo0A-178a and pMTL007::Cdi-CD2492-254a retarget the group II intron to insert into the spo0A andCD2492 genes in the antisense orientation immediately afterthe 178th or 254th nucleotide in the coding sequence, respec-tively. The plasmids were transferred to C. difficile 630 �ermby conjugation, and intron insertion was induced by IPTG.Genomic DNA was isolated from erythromycin-resistanttransconjugants. This DNA was subjected to PCR using (i)the RAM-F/RAM-R primer pair, (ii) the Target_R_Spo0A/EBS universal primer pair, and (iii) the Target_R_CD1A/EBS universal primer pair. As a control, unmodifiedpMTL007 plasmid DNA was also subjected to PCR with thesame primer pairs.
In the pMTL007 control only the RAM-F/RAM-R primerpair produced a PCR product (Fig. 2). This DNA fragment(1,300 bp) represents the erm RAM prior to splicing out of thetd group I intron. As expected, PCR products were not ampli-fied from the pMTL007 template using the Target_R_Spo0A/EBS primer pair or the Target_R_CD1A/EBS primer pairsince this requires integration of the group II intron into therespective target genes (Fig. 2). When genomic DNA isolatedfrom C. difficile 630 �erm carrying pMTL007::Cdi-spo0A-178aor pMTL007::Cdi-CD2492-254a was used as the template forPCR with the RAM-F/RAM-R primer pair, a product of 900bp was amplified in both cases (Fig. 2B). These products are400 bp smaller than the one amplified from the unmodifiedpMTL007 template with the same primer pair and representthe erm RAM after splicing out of the td group I intron. Sincesplicing out of the group I intron accompanies insertion of thegroup II intron into the chromosome, the presence of thesesmaller RAM-F/RAM-R products confirms the insertionevent. Furthermore, the failure to amplify a 1,300-bp productusing the RAM-F/RAM-R primers confirms the absence ofextrachromosomally replicating plasmid, as would be expecteddue to the known instability of this plasmid replicon in C.difficile (14, 27).
To verify that the group II intron had inserted into thecorrect target gene, PCR was carried out by using both theTarget_R_Spo0A/EBS and the Target_R_CD1A/EBS primerpairs. PCR products could only be amplified from thepMTL007::Cdi-spo0A-178a and pMTL007::Cdi-CD2492-254aDNA templates by using the respective Target_R_Spo0A/EBSand Target_R_CD1A/EBS primer pairs (Fig. 2B). Thesespo0A and CD2492 PCR products were 425 and 246 bp in sizereflecting differences in the relative site of annealing of thegene specific primers within their target genes. Overall, this setof PCRs confirms that the erm RAM inserted into the C.difficile chromosome at the correct site and in the correct
VOL. 191, 2009 CLOSTRIDIUM DIFFICILE SPORULATION AND TOXIN PRODUCTION 7299
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

orientation, resulting in the inactivation of the spo0A andCD2492 genes. For unknown reasons, despite multiple at-tempts, it was not possible to generate knockout mutants ofCD1492 or CD1579 using the ClosTron system.
Effect of inactivation of the genes encoding Spo0A and HKCD2492 on C. difficile growth and sporulation capacity. Batchcultures of C. difficile 630 �erm, C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a, and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a were cultivated in BHI broth, and growth wasmonitored by measuring the OD550. All strains grew well inBHI, and vegetative cells of the parental strain and knockout
mutants were indistinguishable using light microscopy (datanot shown). The spo0A and CD2492 mutants grew with thesame exponential growth rate as the parental strain and thestrains had a mean generation time of 66 min (see Fig. S3 inthe supplemental material). All cultures reached the sameOD550 at the end of the exponential phase and exhibited thesame pattern of stationary phase growth with a gradual reduc-tion in OD over time (see Fig. S3 in the supplemental mate-rial). Therefore, neither inactivation of the genes encodingSpo0A or HK CD2492 or the presence of the integrated groupII intron in the genome had any significant effects on the
FIG. 2. Inactivation of genes encoding Spo0A and HK CD2492 in C. difficile using the ClosTron system. (A) ClosTron gene knockout strategy.The ClosTron delivery system is encoded on plasmid pMTL007 and consists of a group II intron (black arrow) with an internal RAM conferringerythromycin resistance (white arrow), which is itself interrupted by the td group I intron (hatched box). The group II intron is retargeted to thedesired target gene (gray arrow) by altering the sequence of the IBS/EBS region using overlap PCR. After introduction of pMTL700-basedplasmids into C. difficile by conjugation from E. coli, production of group II intron RNA is induced from the Pfac promoter by the addition of IPTG.This results in the splicing of the group II intron into the target gene to disrupt it and the splicing out of the td group I intron from the erm RAMto restore a functional ermB gene, allowing positive selection of mutants. The locations of primers used for screening potential mutants areindicated. (B) Confirmation of gene knockouts using PCR. Chromosomal DNA was extracted from C. difficile erythromycin-resistant colonies andscreened using PCR with the primer pairs indicated. Lane A, RAM-F and RAM-R primers; lane B, spo0A target specific primer (Target_R_spo0A)and EBS universal primer; lane C, CD2492 target specific primer (Target_R_CD1A) and EBS universal primer. As a control pMTL007 unmodifiedparental plasmid DNA was included. PCR products were electrophoresed on a 1% (wt/vol) agarose gel and visualized by staining with ethidiumbromide. M, DNA molecular size standards.
7300 UNDERWOOD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.
growth or viability of vegetative C. difficile when cultured inbatch in a rich growth medium.
In contrast, the sporulation capacities of C. difficile 630�erm/pMTL007::Cdi-spo0A-178a and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a were markedly affected com-pared to the parental strain. After 72 h of incubation in BHI,C. difficile 630 �erm produced 14% spores (Fig. 3). Althoughlow compared to other spore-forming bacteria such as B. sub-tilis (38), this inherent sporulation capacity of C. difficile 630�erm is reproducible and comparable to batch cultures ofother virulent C. difficile isolates cultivated for the equivalenttime period (10). Consistent with previous observations (14),inactivation of spo0A in C. difficile resulted in a complete in-ability to sporulate (Fig. 3). Furthermore, inactivation of thegene encoding HK CD2492 in C. difficile also resulted in areduction in the number of spores produced but did not com-pletely abolish sporulation. The sporulation capacity of C. dif-ficile 630 �erm/pMTL007::Cdi-CD2492-254a was reduced to4% after 72 h of cultivation (Fig. 3).
Involvement of Spo0A and HK CD2492 in C. difficile toxinproduction. For several other spore-forming pathogens,Spo0A is known to influence the production of key virulencefactors such as toxins (13, 15, 22, 29). To determine whether asimilar phenomenon exists in C. difficile, we investigated toxinproduction in batch-grown cultures of C. difficile 630 �erm, C.difficile 630 �erm/pMTL007::Cdi-spo0A-178a, and C. difficile630 �erm/pMTL007::Cdi-CD2492-254a.
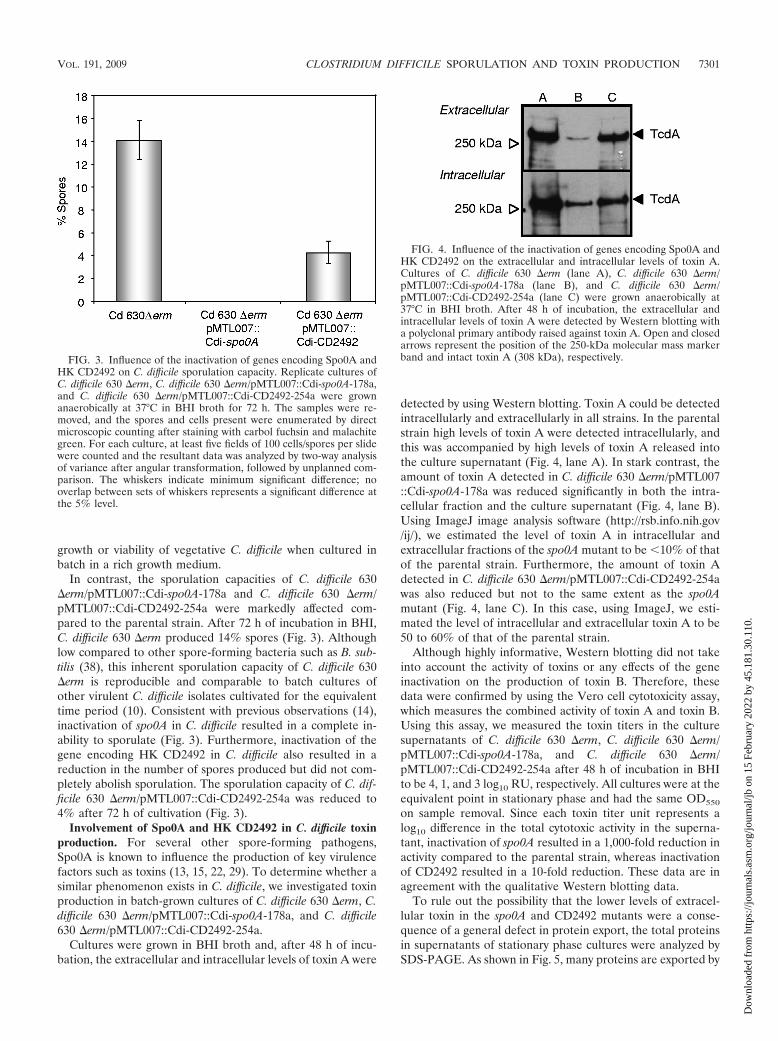
Cultures were grown in BHI broth and, after 48 h of incu-bation, the extracellular and intracellular levels of toxin A were
detected by using Western blotting. Toxin A could be detectedintracellularly and extracellularly in all strains. In the parentalstrain high levels of toxin A were detected intracellularly, andthis was accompanied by high levels of toxin A released intothe culture supernatant (Fig. 4, lane A). In stark contrast, theamount of toxin A detected in C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a was reduced significantly in both the intra-cellular fraction and the culture supernatant (Fig. 4, lane B).Using ImageJ image analysis software (http://rsb.info.nih.gov/ij/), we estimated the level of toxin A in intracellular andextracellular fractions of the spo0A mutant to be 10% of thatof the parental strain. Furthermore, the amount of toxin Adetected in C. difficile 630 �erm/pMTL007::Cdi-CD2492-254awas also reduced but not to the same extent as the spo0Amutant (Fig. 4, lane C). In this case, using ImageJ, we esti-mated the level of intracellular and extracellular toxin A to be50 to 60% of that of the parental strain.
Although highly informative, Western blotting did not takeinto account the activity of toxins or any effects of the geneinactivation on the production of toxin B. Therefore, thesedata were confirmed by using the Vero cell cytotoxicity assay,which measures the combined activity of toxin A and toxin B.Using this assay, we measured the toxin titers in the culturesupernatants of C. difficile 630 �erm, C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a, and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a after 48 h of incubation in BHIto be 4, 1, and 3 log10 RU, respectively. All cultures were at theequivalent point in stationary phase and had the same OD550
on sample removal. Since each toxin titer unit represents alog10 difference in the total cytotoxic activity in the superna-tant, inactivation of spo0A resulted in a 1,000-fold reduction inactivity compared to the parental strain, whereas inactivationof CD2492 resulted in a 10-fold reduction. These data are inagreement with the qualitative Western blotting data.
To rule out the possibility that the lower levels of extracel-lular toxin in the spo0A and CD2492 mutants were a conse-quence of a general defect in protein export, the total proteinsin supernatants of stationary phase cultures were analyzed bySDS-PAGE. As shown in Fig. 5, many proteins are exported by
FIG. 3. Influence of the inactivation of genes encoding Spo0A andHK CD2492 on C. difficile sporulation capacity. Replicate cultures ofC. difficile 630 �erm, C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a,and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a were grownanaerobically at 37°C in BHI broth for 72 h. The samples were re-moved, and the spores and cells present were enumerated by directmicroscopic counting after staining with carbol fuchsin and malachitegreen. For each culture, at least five fields of 100 cells/spores per slidewere counted and the resultant data was analyzed by two-way analysisof variance after angular transformation, followed by unplanned com-parison. The whiskers indicate minimum significant difference; nooverlap between sets of whiskers represents a significant difference atthe 5% level.
FIG. 4. Influence of the inactivation of genes encoding Spo0A andHK CD2492 on the extracellular and intracellular levels of toxin A.Cultures of C. difficile 630 �erm (lane A), C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a (lane B), and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a (lane C) were grown anaerobically at37°C in BHI broth. After 48 h of incubation, the extracellular andintracellular levels of toxin A were detected by Western blotting witha polyclonal primary antibody raised against toxin A. Open and closedarrows represent the position of the 250-kDa molecular mass markerband and intact toxin A (308 kDa), respectively.
VOL. 191, 2009 CLOSTRIDIUM DIFFICILE SPORULATION AND TOXIN PRODUCTION 7301
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

C. difficile, but there were no large-scale reductions in thenumber or amounts of total proteins released by the spo0A andCD2492 mutants that could account for the observed reduc-tions in extracellular toxin. In addition, comparable reductionsin the amount of a high-molecular-mass protein, most likelytoxin A/B, are clearly visible on the stained gel of the proteinsisolated from the supernatants of C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a (Fig. 5, asterisk).
Direct phosphorylation of C. difficile Spo0A by HK CD1579.The genetic evidence discussed above suggests that HKCD2492 is involved in sporulation initiation in C. difficile. Theabsence of Spo0F or Spo0B in C. difficile raises the possibilitythat HK CD1492, HK CD1579, and HK CD2492 phosphory-late Spo0A directly. HK CD1492 and HK CD2492 are large(100 kDa) integral membrane proteins (see Fig. S2 in thesupplemental material). The integral membrane nature ofmany sensor kinases limits investigation of their biochemicalproperties due to problems with heterologous overexpression,solubility, and kinase activity. There are only a few examples ofoverexpression and purification of intact and active full-lengthintegral membrane sensor kinases (4, 11, 26).
In the present study, it was not possible to overexpress andpurify the intact full-length HK CD2492 to investigate its bio-chemical properties. However, due to the high degree of over-all conservation in the active site and DHpt subdomain (Fig.1), we reasoned that it is probable that both HK CD2492 and
HK CD1579 recognize and phosphorylate the same responseregulator in C. difficile. Like the main B. subtilis sporulation-associated sensor kinase KinA (23), HK CD1579 is likely to becytosolic, rendering the full-length protein amenable to heter-ologous expression and purification. The genes encoding HKCD1579 and Spo0A from C. difficile were cloned into pGEX-6PI and purified as described above. On the basis of SDS-PAGE, the HK CD1579 and Spo0A proteins were estimated tobe 80 and 95% pure, respectively (data not shown).
The ability of HK CD1579 to autophosphorylate and trans-fer the phosphoryl group to C. difficile Spo0A was investigatedby using in vitro phosphorylation assays (21, 40, 43). The assaymeasures the incorporation of 32P into sensor kinases from[�-32P]ATP and subsequent transfer to a response regulator.Upon the addition of [�-32P]ATP to HK CD1579 in the ab-sence of Spo0A, a radiolabeled protein of high intensity wasdetected after 20 min of incubation (Fig. 6A, control), whichcorresponds to the phosphorylated kinase (HK CD1579�P).This confirms that the purified protein was active as a kinaseunder these reaction conditions. When Spo0A was included inthe reaction mix from t � 0, the amount of HK CD1579�Pincreased with time, but the level was much reduced comparedto the control, and this was accompanied by a steady increasein the amount of phosphorylated Spo0A (Spo0A�P) with time(Fig. 6A). This demonstrates that HK CD1579 is able to phos-phorylate Spo0A in vitro. In the absence of HK CD1579,
FIG. 5. Effect of inactivation of spo0A and CD2192 on overallprotein export to the culture supernatant. Cultures of C. difficile 630�erm (lane A), C. difficile 630 �erm/pMTL007::Cdi-spo0A-178a (laneB), and C. difficile 630 �erm/pMTL007::Cdi-CD2492-254a (lane C)were grown anaerobically at 37°C in BHI broth. After 48 h of incuba-tion, the total proteins present in culture supernatants were precipi-tated with TCA and subjected to SDS-PAGE and Coomassie bluestaining. The positions of molecular mass standards are indicated byopen arrows. The asterisk indicates a large protein (250 kDa) whichwas at a reduced level in the knockout mutants but not in the parentalstrain.
FIG. 6. Direct phosphorylation of C. difficile Spo0A by HKCD1579. The autophosphorylation and phosphotransfer to Spo0A byHK CD1579 (A) or HK PPA0945 (B) was investigated. Sensor histi-dine kinases (1 �M) were incubated with [�-32P]ATP (50 �Ci in a totalof 100 �M ATP per 20-�l reaction) in the absence (control reactions)or presence of C. difficile Spo0A (10 �M). Control reactions wereallowed to proceed without intervention for 20 min before terminationwith 5� SDS-PAGE sample buffer on dry ice, whereas reactions con-taining Spo0A were terminated at the time points indicated. Sampleswere subjected to SDS-PAGE, and radiolabeled proteins were visual-ized by phosphorimaging. Open arrows indicate phosphorylated pro-teins, whereas the closed arrow indicates the position of the nonphos-phorylated Spo0A that was not detected by phosphorimaging.
7302 UNDERWOOD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

Spo0A does not become phosphorylated by [�-32P]ATP (datanot shown).
To exclude the possibility that the phosphorylation of Spo0Aby HK CD1579 was a result of nonspecific phosphorylation,the assay was carried out using a heterologous sensor kinasefrom Propionibacterium acnes, HK PPA0945, which was puri-fied by using the same vector and conditions as part of anunrelated project in our laboratory. After 20 min of incubationin the absence of Spo0A, a radiolabeled protein correspondingto phosphorylated HK PPA0945 (HK PPA0945�P) was visibleon the gel (Fig. 6B, control) at a level comparable to HKCD1579�P (Fig. 6A, control). This confirms that HKPPA0945 is active under these assay conditions and that theautophosphorylation activity is similar to that of HK CD1579.However, when C. difficile Spo0A was included in the reactionthe level of HK PPA0945�P increased steadily with time butno Spo0A�P could be detected after 20 min (Fig. 6B); at thistime, the level of HK PPA0945�P was comparable to that ofthe control lacking Spo0A. Therefore, in contrast to HKCD1579, HK PPA0945 did not phosphorylate Spo0A. This isstrong evidence that the direct phosphorylation of Spo0A byHK CD1579 is specific. Coomassie blue staining of all gelsconfirmed the presence of proteins at the expected concentra-tions (data not shown).
DISCUSSION
Sporulation is a complex developmental process that is usedby certain bacteria to survive conditions that cannot supportvegetative growth and/or those that threaten integrity or via-bility of the cell. Despite the central importance of spores intransmission, persistence and pathogenesis of C. difficile, thesignal transduction pathway controlling sporulation initiationin this bacterium has not been studied in detail.
Using bioinformatics, we identified three putative C. difficilesporulation-associated sensor kinases, HK CD1492, HKCD1579, and HK CD2492. Inactivation of the gene encodingHK CD2492 had no observable effects on C. difficile growth,viability, or gross cell morphology but reduced the sporulationcapacity by 3.5-fold, providing strong genetic evidence for theinvolvement of HK CD2492 in sporulation initiation. The re-duction in sporulation capacity of the C. difficile CD2492 mu-tant is comparable to the �3-fold reduction in B. subtilis sporu-lation resulting from insertional inactivation of kinA, the geneencoding the main sporulation-associated phosphorelay sensorkinase of this bacterium (23).
Inactivation of CD2492 did not abolish sporulation com-pletely, suggesting that other sensor kinases are likely to beinvolved in sporulation initiation. In B. subtilis, the involve-ment of alternative sensor kinases such as KinB in sporulationonly becomes apparent after inactivation of kinA (41). HKCD1492 and HK CD1579 are the likely candidates for this rolein C. difficile, and the residual sporulation observed in C. dif-ficile 630 �erm/pMTL007::Cdi-CD2492-254a is likely due tothe activity of these alternative kinases. Despite repeated at-tempts to retarget the group II intron to the same and differentinsertion sites, we were unable to obtain knockout mutants inCD1492 or CD1579. Although highly unlikely due to the activesite/DHpt conservation and the fact that HK CD1579 canphosphorylate Spo0A, the possibility that the kinases them-
selves or an additional non-Spo0A response regulator sub-strate for these kinases is essential for viability cannot be ruledout. Nevertheless, the effect of HK CD2492 on C. difficilesporulation is clear and is comparable to the best characterizedof all spore-forming gram-positive bacteria, B. subtilis.
It was not possible to clone, overexpress, and purify HKCD2492 in an intact form from E. coli to study its biochemicalactivity and confirm the genetic data. Similar problems havebeen reported previously with Clostridium botulinum sensorkinases (45). As a tractable alternative, we investigated thebiochemical activity of HK CD1579, a smaller-molecular-masssensor kinase (�72 kDa) lacking membrane spanning seg-ments which, unlike HK CD2492, was amenable to cloning,overexpression, and purification in its full-length intact form.HK CD1579 was active as a kinase and could phosphorylateSpo0A efficiently and at a level similar to that of B. subtilisKinA on Spo0F (40), whereas the heterologous HK PPA0945could not. This suggests that the phosphorylation of Spo0A byHK CD1579 is specific. Under similar reaction conditionsKinA does not readily phosphorylate B. subtilis Spo0A (7). Thedata presented here represent the first report of the directphosphorylation of Spo0A by a sensor kinase in C. difficile orother clostridia and strongly support the hypothesis thatSpo0A is directly phosphorylated by sporulation-associatedsensor kinases in the anaerobic spore formers. Due to theconservation of key residues, HK CD1492 and HK CD2492would also be expected to possess the same specificity and tophosphorylate Spo0A.
The amino acid sequences of the signal input domains of HKCD2492, HK CD1492, and HK CD1579 are highly variable,indicating that each is responsive to a different signal ligand.Furthermore, BLAST analysis of these kinases with microbialgenome sequences in the NCBI database (http://blast.ncbi.nlm.nih.gov/blast.cgi) reveals that orthologues of these proteinsare found in all C. difficile strains currently held in that repos-itory but not in closely related Clostridium species or othergenera of bacteria. The C. difficile strains harboring these sen-sor kinase orthologues include highly virulent strains of currentclinical importance such as ribotype 027 (NAP1/BI). This sug-gests that the signals required to initiate sporulation in C.difficile are unique and specific to this species, as is known to bethe case for members of the genus Bacillus (35). The precisenature of sporulation-inducing signal ligands has generally re-mained elusive. In C. perfringens, inorganic phosphate has beenshown to be a signal capable of promoting spo0A expression,sporulation, and toxin synthesis (25), but any involvement ofsporulation-associated sensor kinases in this process has yet tobe demonstrated.
Inactivation of spo0A in C. difficile 630 �erm resulted in amutant that was completely asporogenous. This is in agree-ment with previous studies on Spo0A in clostridia (3, 14, 15)and highlights the central and essential role of Spo0A in sporu-lation initiation in C. difficile. Inactivation of spo0A also led toa drastic reduction in the level of toxin produced during sta-tionary phase in batch cultures. We have observed similareffects in a chemostat-based model of C. difficile infection ofthe human gut (unpublished data). Inactivation of CD2492resulted in less severe, but nevertheless significant, reductionsin toxin production, as would be expected if HK CD2492 wasone of multiple sensor kinases contributing to Spo0A activa-
VOL. 191, 2009 CLOSTRIDIUM DIFFICILE SPORULATION AND TOXIN PRODUCTION 7303
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

tion. The reduction in extracellular toxin A is mirrored by thedrop in intracellular levels of this protein in the CD2492 andspo0A mutants. The defect in toxin production thus most likelyoccurs at the level of toxin transcription and/or translation, aswould be expected given the well established role of Spo0A intranscriptional control and cellular regulation. Based on thesedata there appears to be a direct correlation between the levelof toxin produced and the relative level of sporulation, whichare influenced by HK CD2492 and Spo0A. This confirms theexistence of firm regulatory links between the sporulation ini-tiation pathway and toxin production in C. difficile.
Although Spo0A is known to be involved in the virulenceresponses of other spore-forming human pathogens, this is, toour knowledge, the first report in C. difficile. Spo0A is a highlypleiotropic cellular regulator that has a dramatic influence onglobal gene expression. Therefore, it is not surprising that C.difficile has adapted this regulatory protein and the pathwaythat controls its activation status to modulate virulence re-sponses. A pertinent question is how does Spo0A exert itseffects on C. difficile toxin production? In the C. difficile ge-nome 200 open reading frames have potential 0A boxeswithin 200 bp of their start codons, indicating direct regulationby Spo0A (28). However, tcdA and tcdB, encoding toxin A andtoxin B, respectively, are not among them. Therefore, Spo0A islikely to have an indirect influence on toxin production, andmany candidate regulators are encoded by the genes putativelyunder the direct control of Spo0A in C. difficile (28). In otherpathogenic spore-forming bacteria the regulatory networksthat influence sporulation initiation and virulence are inti-mately intertwined (24), and it appears that a similar phenom-enon exists in C. difficile. What is clear is that the regulatorynetworks that contribute to both C. difficile sporulation initia-tion and toxin regulation are likely to be highly complex.
Histidine kinase-mediated signal transduction pathways,such as phosphorelays and TCSs, are known to be integralelements of the virulence responses of bacterial pathogens andconsequently are recognized targets for the development ofnovel antimicrobial agents (36, 37). Therefore, selective inhi-bition of Spo0A or other elements of the signaling pathwaythat influences Spo0A activity has the potential to simulta-neously reduce toxin production and inhibit sporulation of C.difficile. Ultimately, this could have important consequencesfor the therapy of C. difficile infections and the control oftransmission and persistence of this highly significant nosoco-mial pathogen.
ACKNOWLEDGMENTS
The study was supported in part by BBSRC, Leeds University, TheRoyal Society, and The Hospital Infection Society.
We thank Nigel P. Minton and John T. Heap for providing theClosTron gene knockout system.
REFERENCES
1. Akerlund, T., I. Persson, M. Unemo, T. Noren, B. Svenungsson, M. Wullt,and L. G. Burman. 2008. Increased sporulation rate of epidemic Clostridiumdifficile type 027/NAP1. J. Clin. Microbiol. 46:1530–1533.
2. Akerlund, T., B. Svenungsson, A. Lagergren, and L. G. Burman. 2006.Correlation of disease severity with fecal toxin levels in patients with Clos-tridium difficile-associated diarrhea and distribution of PCR ribotypes andtoxin yields in vitro of corresponding isolates. J. Clin. Microbiol. 44:353–358.
3. Alsaker, K. V., T. R. Spitzer, and E. T. Papoutsakis. 2004. Transcriptionalanalysis of spo0A overexpression in Clostridium acetobutylicum and its effecton the cells response to butanol stress. J. Bacteriol. 189:1959–1971.
4. Bader, M. W., S. Sanowar, M. E. Daley, A. R. Schneider, U. Cho, W. Xu, R. E.Klevit, M. H. Le, and S. I. Miller. 2005. Recognition of antimicrobial pep-tides by a bacterial sensor kinase. Cell 122:461–472.
5. Baines, S. D., J. Freeman, and M. H. Wilcox. 2005. Effects of piperacillin-tazobactam on Clostridium difficile growth and toxin production in a humangut model. J. Antimicrob. Chemother. 55:974–982.
6. Burbulys, D., K. A. Trach, and J. A. Hoch. 1991. Initiation of sporulation inBacillus subtilis is controlled by a multicomponent phosphorelay. Cell 64:545–552.
7. Cervin, M. A., and G. B. Spiegelman. 1999. The Spo0A sof mutations revealregions of the regulatory domain that interact with a sensor kinase and RNApolymerase. Mol. Microbiol. 31:597–607.
8. Dupuy, B., and A. L. Sonenshein. 1998. Regulated transcription of Clostrid-ium difficile toxins. Mol. Microbiol. 27:107–120.
9. Elliott, B., B. J. Chang, C. L. Golledge, and T. V. Riley. 2007. Clostridiumdifficile-associated diarrhoea. Intern. Med. J. 37:561–568.
10. Fawley, W. N., S. Underwood, J. Freeman, S. D. Baines, K. Saxton, K.Stephenson, R. C. Owens, Jr., and M. H. Wilcox. 2007. Efficacy of hospitalcleaning agents and germicides against epidemic Clostridium difficile strains.Infect. Control Hosp. Epidemiol. 28:920–925.
11. Fleischer, R., R. Heermann, K. Jung, and S. Hunke. 2007. Purification,reconstitution, and characterization of the CpxRAP envelope stress systemof Escherichia coli. J. Biol. Chem. 282:8583–8593.
12. Freeman, J., F. J. O’Neill, and M. H. Wilcox. 2003. Effects of cefotaxime anddesacetylcefotaxime upon Clostridium difficile proliferation and toxin pro-duction in a triple-stage chemostat model of the human gut. J. Antimicrob.Chemo. 52:96–102.
13. Gominet, M., L. Slamti, N. Gilois, M. Rose, and D. Lereclus. 2001. Oligopep-tide permease is required for expression of the Bacillus thuringiensis plcRregulon and for virulence. Mol. Microbiol. 40:963–975.
14. Heap, J. T., O. J. Pennington, S. T. Cartman, G. P. Carter, and N. P. Minton.2007. The ClosTron: a universal gene knockout system for the genus Clos-tridium. J. Microbiol. Methods 70:452–464.
15. Huang, I.-H., M. Waters, R. R. Grau, and M. R. Sarker. 2004. Disruption ofthe gene (spo0A) encoding sporulation transcription factor blocks endosporeformation and enterotoxin production in enterotoxigenic Clostridium perfrin-gens type A. FEMS. Microbiol. Lett. 233:233–240.
16. Hussain, H. A., A. P. Roberts, and P. Mullany. 2005. Generation of anerythromycin-sensitive derivative of Clostridium difficile strain 630 (630�erm) and demonstration that the conjugative transposon Tn916�E entersthe genome of this strain at multiple sites. J. Med. Microbiol. 54:137–141.
17. Jiang, M., W. Shao, M. Perego, and J. A. Hoch. 2000. Multiple histidinekinases regulate entry into stationary phase and sporulation in Bacillus sub-tilis. Mol. Microbiol. 38:535–542.
18. Kamiya, S., H. Ogura, X. Q. Meng, and S. Nakamura. 1992. Correlationbetween cytotoxin production and sporulation in Clostridium difficile. J. Med.Microbiol. 37:206–210.
19. Karlsson, S., L. G. Burman, and T. Akerlund. 1999. Suppression of toxinproduction in Clostridium difficile VPI 10463 by amino acids. Microbiol.145:1683–1693.
20. Karlsson, S., L. G. Burman, and T. Akerlund. 2008. Induction of toxins inClostridium difficile is associated with dramatic changes of its metabolism.Microbiology 154:3430–3436.
21. Kojetin, D. J., R. J. Thompson, L. M. Benson, S. Naylor, J. Waterman, K. G.Davies, C. H. Opperman, K. Stephenson, J. A. Hoch, and J. Cavanagh. 2005.Structural analysis of divalent metals binding to the Bacillus subtilis responseregulator Spo0F: the possibility for in vitro metalloregulation in the initiationof sporulation. Biometals 18:449–466.
22. Lereclus, D., H. Agaisse, C. Grandvalet, S. Salamitou, and M. Gominet.2000. Regulation of toxin and virulence gene transcription in Bacillus thu-ringiensis. Int. J. Med. Microbiol. 290:295–299.
23. Perego, M., S. P. Cole, D. Burbulys, K. Trach, and J. A. Hoch. 1989. Char-acterization of the gene for a protein kinase which phosphorylates the sporu-lation-regulatory proteins Spo0A and Spo0F of Bacillus subtilis. J. Bacteriol.171:6187–6196.
24. Perego, M., and J. A. Hoch. 2008. Commingling regulatory systems followingacquisition of virulence plasmids by Bacillus anthracis. Trends Microbiol.16:215–221.
25. Philippe, V. A., M. B. Mendez, I. H. Huang, L. M. Orsaria, M. R. Sarker, andR. R. Grau. 2006. Inorganic phosphate induces spore morphogenesis andenterotoxin production in the intestinal pathogen Clostridium perfringens.Infect. Immun. 74:3651–3656.
26. Potter, C. A., A. Ward, C. Laguri, M. P. Williamson, P. J. Henderson, andM. K. Phillips-Jones. 2002. Expression, purification, and characterisation offull-length histidine protein kinase RegB from Rhodobacter sphaeroides. J.Mol. Biol. 320:201–213.
27. Purdy, D., T. A. O’Keeffe, M. Elmore, M. Herbert, A. McLeod, M. Bokori-Brown, A. Ostrowski, and N. P. Minton. 2002. Conjugative transfer of clos-tridial shuttle vectors from Escherichia coli to Clostridium difficile throughcircumvention of the restriction barrier. Mol. Microbiol. 46:439–452.
28. Rose, G. 2007. Defining the Spo0A regulon of Clostridium difficile and other
7304 UNDERWOOD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.

clinically and industrially significant clostridia. M.Res. thesis. University ofLeeds, Leeds, United Kingdom.
29. Saile, E., and T. M. Koehler. 2002. Control of anthrax toxin gene expressionby the transition state regulator abrB. J. Bacteriol. 184:370–380.
30. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory Press, New York, NY.
31. Savage, A. M., and R. H. Alford. 1983. Nosocomial spread of Clostridiumdifficile. Infect. Control. 4:31–33.
32. Sebaihia, M., B. W. Wren, P. Mullany, N. F. Fairweather, N. Minton, R.Stabler, N. R. Thomson, A. P. Roberts, A. M. Cerdeno-Tarraga, H. Wang,M. T. Holden, A. Wright, C. Churcher, M. A. Quail, S. Baker, N. Bason, K.Brooks, T. Chillingworth, A. Cronin, P. Davis, L. Dowd, A. Fraser, T. Felt-well, Z. Hance, S. Holroyd, K. Jagels, S. Moule, K. Mungall, C. Price, E.Rabbinowitsch, S. Sharp, M. Simmonds, K. Stevens, L. Unwin, S. Whithead,B. Dupuy, G. Dougan, B. Barrell, and J. Parkhill. 2006. The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaicgenome. Nat. Genet. 38:779–786.
33. Skerker, J. M., B. S. Perchuk, A. Siryaporn, E. A. Lubin, O. Ashenberg, M.Goulian, and M. T. Laub. 2008. Rewiring the specificity of two-componentsignal transduction systems. Cell 133:1043–1054.
34. Spiegelman, G. B., T. H. Bird, and V. Voon. 1995. Transcription regulationby the Bacillus subtilis response regulator Spo0A, p. 159–179. In J. A. Hochand T. J. Silhavy (ed.), Two-component signal transduction. ASM Press,Washington, DC.
35. Stephenson, K., and J. A. Hoch. 2002. Evolution of signalling in the sporu-lation phosphorelay. Mol. Microbiol. 46:297–304.
36. Stephenson, K., and J. A. Hoch. 2004. Developing inhibitors to selectivelytarget two-component and phosphorelay signal transduction systems. Curr.Med. Chem. 11:765–773.
37. Stephenson, K., and J. A. Hoch. 2002. Two-component and phosphorelaysignal transduction systems as therapeutic targets. Curr. Opin. Pharmacol.2:507–512.
38. Stephenson, K., and J. A. Hoch. 2001. PAS-A domain of phosphorelay
sensor kinase A: a catalytic ATP-binding domain involved in the initiation ofdevelopment in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 98:15251–15256.
39. Stephenson, K., and R. J. Lewis. 2005. Molecular insights into the initiationof sporulation in gram-positive bacteria: new technologies for an old phe-nomenon. FEMS. Microbiol. Rev. 29:281–301.
40. Stephenson, K., Y. Yamaguchi, and J. A. Hoch. 2000. The mechanism ofaction of inhibitors of bacterial two-component signal transduction systems.J. Biol. Chem. 275:38900–38904.
41. Trach, K. A., and J. A. Hoch. 1993. Multisensory activation of the phos-phorelay initiating sporulation in Bacillus subtilis: identification and se-quence of the protein kinase of the alternate pathway. Mol. Microbiol.8:69–79.
42. Walters, B. A., R. Roberts, R. Stafford, and E. Seneviratne. 1983. Relapse ofantibiotic associated colitis: endogenous persistence of Clostridium difficileduring vancomycin therapy. Gut 24:206–212.
43. Wang, L., C. Fabret, K. Kanamaru, K. Stephenson, V. Dartois, M. Perego,and J. A. Hoch. 2001. Dissection of the functional and structural domains ofphosphorelay histidine kinase A of Bacillus subtilis. J. Bacteriol. 183:2795–2802.
44. White, R. A., H. Szurmant, J. A. Hoch, and T. Hwa. 2007. Features ofprotein-protein interactions in two-component signaling deduced fromgenomic libraries. Methods Enzymol. 422:75–101.
45. Worner, K., H. Szurmant, C. Chiang, and J. A. Hoch. 2006. Phosphorylationand functional analysis of the sporulation initiation factor Spo0A from Clos-tridium botulinum. Mol. Microbiol. 59:1000–1012.
46. Zapf, J., U. Sen, Madhusudan, J. A. Hoch, and K. I. Varughese. 2000. Atransient interaction between two phosphorelay proteins trapped in a crystallattice reveals the mechanism of molecular recognition and phosphotransferin signal transduction. Struct. Fold. Des. 8:851–862.
47. Zhao, H., T. Msadek, J. Zapf, Madhusudan, J. A. Hoch, and K. I. Varughese.2002. DNA complexed structure of the key transcription factor regulatingdevelopment in sporulating bacteria. Structure 10:1041–1050.
VOL. 191, 2009 CLOSTRIDIUM DIFFICILE SPORULATION AND TOXIN PRODUCTION 7305
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Febr
uary
202
2 by
45.
181.
30.1
10.





























