Embed Size (px)
Citation preview

Curr Genet (1995) 28:346-352 �9 Springer-Verlag 1995
Raman Kumar �9 Jan Drouaud - Monique Raynal Ian Small
Characterization of the nuclear gene encoding chloroplast ribosomal protein $13 from Arabidopsis thaliana
Received: 2 February/20 March 1995
A b s t r a c t We have characterised a cDNA clone and a nuclear gene encoding the chloroplast 30 s ribosomal protein S13 from Arabidopsis thaliana. The identifica- tion is based on the high similarity of the predicted amino-acid sequence with eubacterial S13 protein se- quences, and immunodetection of a 14.5-kDa chloro- plast ribosomal polypeptide using antibodies raised against the polypeptide produced from part of the cDNA expressed in bacteria. The predicted amino-acid sequence contains an N-terminal extension which has several features characteristic of chloroplast transit peptides. Experiments suggest there is a single copy of this gene in A. thaliana and multiple copies in Brassica species. The origin of the mitochondrial S13 polypept- ide in crucifers is also discussed.
Key words Arabidopsis thaIiana �9 Ribosomal protein �9 Chloroplast S13 �9 Mitochondrial $13
Introduction
Ribosomes are RNA-protein complexes involved in protein synthesis in all living cells. The ribosomes in the cytosol, chloroplasts, and mitochondria differ from each other with regard to the kind, number, and size of constituent ribosomal proteins (r-proteins) and rRNAs (Subramanian 1993; Kumar 1995). Although all the r-protein genes of the eubacterium Escherichia coli
R. Kumar " J. Drouard �9 I. Small (5:~) Station de G6n6tique et d'Am61ioration des Plantes, INRA, Route de St-Cyr, F-78026 Versailles Cedex, France
M. Raynal Laboratoire de Physiologie et Biologie Mol6culaire V6g6tale, CNRS-Universit6, Avenue de Villeneuve, F-66025 Perpignan Cedex, France
Communicated by C.J. Leaver
have been cloned and characterised (Nomura 1990), similar information on plant ribosomes is still incom- plete. Plant organelle ribosomes are complexes of about 55-70 r-proteins (Capel and Bourque 1982; Douce 1985; Subramanian et al. 1991). Most of these have counterparts in eubacterial ribosomes (Shinozaki et al. 1986; Ohyama et al. 1988; Hiratsuka et al. 1989; Takemura et al. 1992; Subramanian 1993; Wakasugi et al. 1994), and this similarity provides some of the strongest evidence in favour of the endosymbiotic ori- gin of chloroplasts and mitochondria (reviewed in Gray 1992). Recently, however, examples of chloroplast r-proteins with no homologs in eubacterial ribosomes have been described (reviewed in Subramanian 1993). Of the many proteins involved in organellar ribosomal structure and function, genes for only a few are present on their respective genomes, the majority being encoded by nuclear genes. The mRNAs from these nuclear genes are translated in the cytosol to give precursor proteins containing amino-terminal transit peptides which direct import of the protein into the organelles. These nuclear genes are thought to have arisen from gene transfer between organelles and the nucleus during evolution (reviewed in Brennicke et al. 1993; Subramanian 1993). Functional expression of a nuclear gene encoding an organellar protein usually leads to the rapid loss of the corresponding organellar gene. Examples are provided by the genes encoding organellar S13 ribosomal proteins. None of the se- quenced chloroplast genomes contain a gene coding for this polypeptide, and the mitochondrial gene rps13 has apparently been lost at least twice during angiosperm evolution; once in legumes, and once in Brassicaceae (Nugent and Palmer 1993). As a start towards unravell- ing the intriguing situation surrounding the origins of organellar S13 polypeptides in different plant species, in this paper we describe the characterization of a cDNA clone originally sequenced as part of the effort to provide a transcript map of the Arabidopsis thaliana genome (H6fte et al. 1993) and presumed to encode an

347
organellar S13 homolog because of its similarity to bacterial S13 sequences.
Materials and methods
Plant materials. A. thaIiana var. Columbia, rapeseed (Brassiea napus var. Brutor), cauliflower (B. oleracea), turnip (B. rapa var. Tobin), maize (Zea mays), tobacco (Nicotiana tabacum cv Xanthi), broad bean (Phaseolus vulgaris cv Jalo) and wheat (Triticum aestivum cv. Soissons) were used in the present study.
Isolation of total DNA and total and poly(A) RNA. Total DNA of greenhouse-grown plants of A. thaliana and three Brassica species was extracted according to Dellaporta et al. (1983) and further purified by centrifugation in a CsC1/ethidium bromide gradient. Total RNA from A. thaIiana was extracted using guanidine hydro- chloride (Logemann et al. 1987) and purified through a CsC1 cushion (Sambrook et al. 1989). The poly(A) + fraction was re- covered using a mRNA isolation kit (Pharmacia Biotech SA, St. Quentin-en-Yvelines, France) following the manufacturer's instuc- tions.
Cytosolic, chloroplast and mitochondrial ribosome isolation. About 100 g of leaves of A. thaliana and B. napus were homogenised in extraction buffer (2 ml/g tissue) containing 0.4 M sucrose, 50 mM Tris-HC1 pH 7.5, 25 mM KC1, 5 mM MgCI2, 5 mM 13-mercapto- ethanol, 4~ and filtered through four layers of miracloth (Cal- biochem, La Jolla, Calif., USA). After a slow spin at 360 g (1500 rpm, Kontron A 6.14 rotor) for 10 min to remove nuclei, the supernatant was centrifuged at 1000g (2500rpm, Kontron A 6.14 rotor) for 10 min. The pellet was lysed in cold 10 mM Tris-HC1 pH 7.5, 10 mM MgC12, 50 mM KC1, 2.0% Triton X-100, centrifuged in a microfuge for 10 rain at 4~ and the supernatant layered on to a 1-M sucrose cushion prepared in 10 mM Tris-HC1 pH 7.5, 50 mM KC1, 10 mM MgC12 and centrifuged at 110 000 g (25 000 rpm, Kontron TST 41.14 rotor) for 15 h. The pellet enriched in chloroplast ribosomes was precipitated in 10% (v/v) trichloroacetic acid in acetone, incubated at -20 ~ for 1 h, centrifuged in a microfuge to collect the protein pellet, washed with cold acetone, dried and resuspended in 1 x Laemmli treatment buffer (Laemmli 1970). To obtain cytosolic ribosomes, the post-chloroplast supernatant was spun at 3500g (4600 rpm, Kontron A 6.14 rotor) for 10 min to remove contamina- ting plastids and then centrifuged at 24000 g (12000 rpm, Kontron A 6.14 rotor) for 10 rain to collect mitochondria, and the super- natant directly loaded onto the cushion. The pellet enriched in cytosolic ribosomes was processed as above. For separating frac- tions enriched in mitochondrial ribosomes, etiolated seedlings of wheat, rapeseed, maize and bean germinated at 25 ~ were used. For tobacco, however, greenhouse-grown plants were kept in the dark for 48 h before their leaves were de-ribbed and used for extraction. In each case, tissue was homogenised in a large excess of extrac- tion buffer and mitochondria purified according to the method detailed by Kumar et al. (1995). Purified mitochondria were pro- cessed in the same way as chloroplasts in order to obtain the ribosomal fraction.
Total proteins. Total proteins were extracted from A. thaliana and B. napus leaves essentially according to Damerval et al. (1986). Protein concentration was assayed according to Bradford (1976).
Isolation of cDNA and 9enomic clones. The cDNA clone (YAP046T) was obtained by random sequencing of clones from a cDNA library of developing siliques prepared in ~ZAPII as part of a combined effort to prepare a transcript map of A. thaliana (H6fte et al. 1993). Genomic clones were obtained from a library of A. thaliana var.
Columbia DNA, prepared in )~GEM11 (Promega Corporation, Madison, WIS., USA) by Dr J.T. Mulligan (Stanford University) and distributed via the EEC-BRIDGE Arabidopsis DNA stock Centre, K61n, by Dr. Jeff Dangl. The library was screened with digoxigenin- labelled transcripts of YAP046T using an alkaline phosphatase detection protocol following the manufacturer's instructions (Boeh- ringer Mannheim GmbH, Mannheim, Germany). Suitable frag- ments from these inserts were subcloned into Bluescript KS + or K S - (Stratagene, La Jolla, Calif.), and sequenced using single- stranded DNA and fluorophore-labelled universal primers (Applied Biosystems, Foster City, Calif.) in an Applied Biosystems 373A sequencing system.
Southern- and Northern-blot analyses. A 550-bp fragment of YAP046T from the EcoRI cloning site to an internal PstI site (nt 546; Fig. 1), which included the 5' end of the cDNA and most of the coding region, was purified by the Gtassmitk method (Bio 101, Calil) and labelled with [~ 3zp~ dCTP (Amersham France SA, Les Ulis, France) using a random primer labelling kit (Pharmacia). For Southern-blot analysis, total DNAs digested with different re- striction enzymes were electrophoresed on a 0.7% agarose gel in 1 x TBE (0.045 M Tris-borate, 0.001 M EDTA), and blotted onto a Hybond-N nylon membrane (Amersham) by capillary blotting in 10 x SSC. Membranes were pre-hybridised and hybridised in 6 x SSC, 50% formamide, 0.1% N-lauroylsarcosine, 0.02% SDS and 2% blocking agent (Boehringer Mannheim). The final wash for membranes with A. thaliana samples was in 0.1 x SSC with 0.1% SDS at 60~ for 1 h and for membranes with Brassica samples was in 2xSSC with 0.1% SDS at 55~ for l h . For Northern-blot analysis, total and poly(A) § RNA from A. thaliana inflorescences were separated on a 1.2% denaturing gel contain- ing 6% formaldehyde and transferred to Hybond-N + (Amer- sham) by capillary blotting in 20 x SSC. Pre-hybridisation, hy- bridisation and washing was similar to that for Southern blots on Arabidopsis DNA.
Raisin9 of an antibody and Western-blot analysis. A 387 bp ScaI- DraI fragment (nt. 284-670; Fig. 1) of YAP046T was cloned in-frame at a filled-in BamHI site in the pET14 expression vector (Novagen, Madison, WIS). A fragment containing the junction was subcloned and sequenced to confirm an in-frame cloning. The construct, origin- ally established in E. coli strain NM522, was transformed into the expression-host strain BL21(DE3)pLysS (Novagen). The cells we.re grown at 37 ~ in the presence of carbenicillin and c h!orampllenico,1 and induced by adding IPTG to a final concentration of 0,4 mM when the cell density of the bacterial culture reached an absorbance of 0.5 at 600 nm. The cells were harvested and the protein purifie d ̀o.n Ni-NTA resin (Qiagen Inc., Calif) according to the manufacturer's instructions. About 1 mg of purified protein homogenised with com- plete Freund's adjuvant was injected into a New Zealand male rabbit and then booster injections (1 mg of protein in incomplete Freund's adjuvant) were administered three times at 2-week inter- vals. Blood was collected 7 days after the last injection. Antiserum at 1 : 5000 dilution and an incubation time of 1 h gave a strong specific signal, whereas pre-immune serum gave no significant signal under the same conditions. Western blotting and immunostaining was done essentially as described earlier (Kumar et al. t995). Pre-stained low-molecular-weight markers (BRL Life Technologies SARL, Eragny, France) were used to monitor protein transfers onto nitro- cellulose C (Amersham) and to estimate the sizes of the im- munodetected proteins. Antibody against plant mitochondrial S13 was kindly provided by Dr. Koh-ichi Kadowaki, National Institute of Agrobiological Resources (Japan), and Prof. C.S. Levings III, NCSU, N.C. (USA).
Computer analysis. Computer analysis of sequences was carried out using the GCG package (Genetics Computer Group 1991) and the GenBank (release 84) and EMBL (release 39) databases.

348
Results
c D N A sequence and t ranscript analysis
The c D N A clone (YAP046T) contains a single uninter- rupted reading frame of 169 codons, coding for a basic protein of 19.1 k D a (Fig. 1). The first A T G codon is in the perfect consensus context ( A A C A A T G G C ) found from a study of functional plant initiation codons (Liitcke et al. 1987) and is thus presumed to be the initiation codon. In this case the c D N A contains 96 bp of untransla ted 5' sequence, 134 bp of 3' untransla ted sequence and a 28 bp poly(A) tract at the 3' end, and can be assumed to be a lmost full-length. The deduced amino-acid sequence of this clone shows mos t sim- ilarity (up to 53% identity) with eubacterial S13 r-proteins. A distance tree of different S13 homologs places YAP046T with the eubacterial S13 sequences, ra ther than with cytosolic or mi tochondr ia l sequences
i0 30 50 CG~IGC CTTATCCG~QAGC~ATCTTCq~PCTTCT?CTTCGTCACCTCTGAATTAGTIT
70 90 ii0 C CCAGAATCO~ TTCCTAGGAAGAGAACATAACAATGG CG CAGATGGTTG CGATGCC C
M A Q M V A M P
130 150 170 G TAGCGCAC TCG CTCTCACTCA2~f ~G CA ATTGGGCAAAATCCAATC CTCTCTCTCGCAAC V A H S L S L I C N W A K S N P L S R N
intron 1 190 210 IV 230
ACTCTq]3 CCCTC CCAG CTTCAAACAC TC CCAATAAG CAGAGTCTAAGCATCAGATGTGC T T L A L P A S N T P N K Q S L S I R C
250 270 290 CGTG TTGGTC~GTGGAGATTCCGGC TAATAAGAGAATTGAG TACTCTCITCAGTACATT
D v I A D R D V V I
310 330 350 CATGGGATI~TCGAACTAGGGCTCG TCAGAq'PC T?GTTSATCTTCAGATGGAGAACAAG
F I G R T Q V K E A VS D T
370 390 410 ATTAC TAAAGACATGG CTGAGGAAGAACTCATCA Tr CTTC GTGATGAAGTCTC CAAG TAC I T K B M A ~ I I L ~'--~ E V S ~] Y R V R L T G K I I I D L
in,on 2 430 �9 450 470
ATGATTGAGGGTGATCTGAGGAGG TfCAATGCTC TTGCAATCAAGAGGTTGAAGGAGATC
K V G D L R E V S N I
49Q 510 530 CAGTG T~ACOSTGGGGTAAG GCACATCCAAGGGC TG CCATGTCGGGGACAG AGAACCAAG
C ~ N S v
Q T I R
550 570 590 AACAACTGCAGGACTC qTAAAGGCAAGAAAATTG CTATTG CAGGGAAGAAGAAAGTTTCC ~ - - ~ C ~ - - ~ L ~ K K I A I ~] G ~ V S
A R P R R T V N
610 630 650 AAG TAATCA TCTTCATGATCTCCTTC TTC~TGTAATAC TC TI[[TTAGTTTG TC TAGTC K
670 690 710 ATC TC TG TTTAAATGTC&q~PTG TAATTCCCAAACCTTTGTTCGAACTGAGC CC TI~ CAG
730 TTTATCTTCGGCqTTITAG CT
Fig. 1 DNA sequence of the cDNA clone YAP046T. The predicted amino-acid sequence is shown below the nucleotide sequence (database accession Z17611), and the complete sequence of Bacillus subtilis S13 has been aligned with it. Conserved amino acids are boxed. The sites where the two introns were found in the genomic sequence (database accession Z47986) are marked by filled arrow- heads. The oligopyrimidine tract at the 5' end of the cDNA is underlined.
(Fig. 2). S13 is a componen t of the small (30s) r ibosomal subunit, and the topology of the tree matches phylogenetic trees based on another small subunit component , 16 s 18 s r R N A (Gray 1992), even to the extent of placing the a rchaebac te r ium I talobac- terium marismortui with eukaryotes rather than with eubacteria. This close match to the well-established r R N A trees suggests that the grouping of YAP046T with eubacterial S13 sequences is reliable. To confirm that the sequence corresponds to an A. thaliana m R N A , par t of the c D N A was used as a hybridisat ion probe on Nor the rn blots (Fig. 3). I t hybridised to a single band at
[ _ ~ human 1 mouse eukaryetic drosophila cytosolic $18 sequences
- - Arabidopais thaliana
Halobactariurn marismortui
B. staarothermophilua eubacterial $13 E. coil sequences Chlamydia trachomatis
maize
wheat
,, [ L . ~ tobacco rnitochondrial $13 sequences
_J I :2o. Prototheca wickerhamii
Fig. 2 Clustering relationships of $13 sequences. The tree shows the relationship between the different amino-acid sequences based on their sequence similarity. The tree was generated with the program PILEUP from the GCG package using the default parameters. The sequences used were the following (database accession numbers are in brackets): human (P25232), mouse (M76762), Drosophila (L22959), A. thaliana (Z23165), Halobacterium (Q00861), B. subtilis (P20282), B. stearothermophiIus (P15757), E. cell (P02369), Chlamydia (L23478), A. thaliana cDNA clone YAP046T (Z17611), and mitochondrial se- quences from maize (P08977), wheat (P0~/924), tobacco (P05488), carrot (P23209), the liverwort Marchantia polymorpha (P26872) and the colourless alga Prototheca wickerhamii (U02970).
Fig. 3 Northern blot analysis using YAP046T as a probe; Four gg of poly(A) + RNA (lane 1), 16 gg of total RNA (lane 2) and 8 gg of total RNA (lane 3) from A. thaliana inflorescences were used. The blot was probed with a 550-bp fragment of cDNA that included the 5' end and most of the coding region of the gene. A single transcript of 850 nt hybridized to the probe. The sizes of the molecular-weight markers are given in kb
approximately 850 nucleotides in total RNA from A. thaliana inflorescences and was much enriched in the poly(A) + fraction, proving that the mRNA comes from a nuclear gene. The similarity of the amino-acid sequence predicted from the cDNA sequence to eubac- terial S13 r-proteins, and the long N-terminal exten- sion, suggested that the cDNA encodes an organellar polypeptide.
Immunodetect ion of chloroplast S13
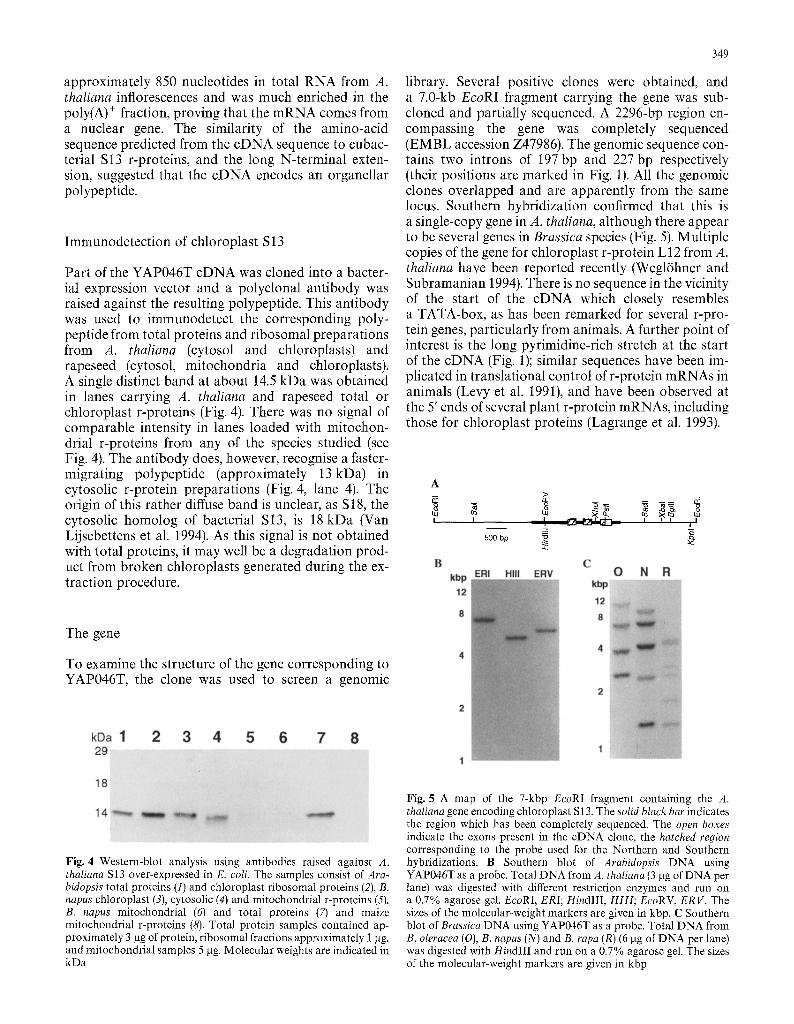
Part of the YAP046T cDNA was cloned into a bacter- ial expression vector and a polyclonal antibody was raised against the resulting polypeptide. This antibody was used to immunodetect the corresponding poly- peptide from total proteins and ribosomal preparations from A. thaliana (cytosol and chloroplasts) and rapeseed (cytosol, mitochondria and chloroplasts). A single distinct band at about 14.5 kDa was obtained in lanes carrying A. thaliana and rapeseed total or chloroplast r-proteins (Fig. 4). There was no signal of comparable intensity in lanes loaded with mitochon- drial r-proteins from any of the species studied (see Fig. 4). The antibody does, however, recognise a faster- migrating polypeptide (approximately 13kDa) in cytosolic r-protein preparations (Fig. 4, lane 4). The origin of this rather diffuse band is unclear, as S18, the cytosolic homolog of bacterial S13, is 18 kDa (Van Lijsebettens et al. 1994). As this signal is not obtained with total proteins, it may well be a degradation prod- uct from broken chloroplasts generated during the ex- traction procedure.
349
library. Several positive clones were obtained, and a 7.0-kb EcoRI fragment carrying the gene was sub- cloned and partially sequenced. A 2296-bp region en- compassing the gene was completely sequenced (EMBL accession Z47986). The genomic sequence con- tains two introns of 197 bp and 227 bp respectively (their positions are marked in Fig. 1). All the genomic clones overlapped and are apparently from the same locus. Southern hybridization confirmed that this is a single-copy gene in A. thaliana, although there appear to be several genes in Brassica species (Fig. 5). Multiple copies of the gene for chloroplast r-protein L12 from A. thaliana have been reported recently (Wegl6hner and Subramanian 1994). There is no sequence in the vicinity of the start of the cDNA which closely resembles a TATA-box, as has been remarked for several r-pro- tein genes, particularly from animals. A further point of interest is the long pyrimidine-rich stretch at the start of the cDNA (Fig. 1); similar sequences have been im- plicated in translational control of r-protein mRNAs in animals (Levy et al. 1991), and have been observed at the 5' ends of several plant r-protein mRNAs, including those for chloroplast proteins (Lagrange et al. 1993).
A
~a t~
_= s00 bp ~
The gene
To examine the structure of the gene corresponding to YAP046T, the clone was used to screen a genomic
Fig. 4 Western-blot analysis using antibodies raised against A. thaliana S13 over-expressed in E. coli. The samples consist of Ara- bidopsis total proteins (1) and chloroplast ribosomal proteins (2), B. napus chloroplast (3), cytosolic (4) and mitochondrial r-proteins (5), B. napus mitochondrial (6) and total proteins (7) and maize mitochondrial r-proteins (8). Total protein samples contained ap- proximately 3 lag of protein, ribosomal fractions approximately 1 lag, and mitochondrial samples 5 lag. Molecular weights are indicated in kDa
Fig. 5 A map of the 7-kbp EcoRI fragment containing the A. thaliana gene encoding chloroplast S13. The solid black bar indicates the region which has been completely sequenced. The open boxes indicate the exons present in the cDNA clone, the hatched region corresponding to the probe used for the Northern and Southern hybridizations. B Southern blot of Arabidopsis DNA using YAP046T as a probe. Total DNA from A. thaliana (3 gg of DNA per lane) was digested with different restriction enzymes and run on a 0.7% agarose gel. EcoRI, ERI; HindlII, ItIII; EcoRV, ERV. The sizes of the molecular-weight markers are given in kbp. C Southern blot of Brassica DNA using YAP046T as a probe. Total DNA from B. oleracea (0), B. napus (N) and B. rapa (R) (6 lag of DNA per lane) was digested with HindIII and run on a 0.7% agarose gel. The sizes of the molecular-weight markers are given in kbp

350
Discussion
The results clearly show that the cDNA clone YAP046T encodes a chloroplast r-protein. The de- duced amino-acid sequence suggests that this protein is the chloroplast equivalent of eubacterial S13. The bac- terial polypeptide has been found to have two func- tional domains; the highly basic C-terminus binds weakly to 16 s rRNA, whilst the N-terminal two-thirds contains a binding site for ribosomal protein S19 (Schwarzbauer and Craven 1985). S13/S19 dimers bind tightly and specifically to 16 s rRNA (Dijk et al. 1977) and, although the exact role of S13 is unclear, ribo- somes deficient in S13 lose 25% of their protein-syn- thesis activity (Schwarzbauer and Craven, t985).
The cDNA sequence predicts a polypeptide of 19.1 kDa, considerably larger than the 14.5 kDa ob- served. However, the amino-acid sequence similarity with eubacterial counterparts starts with alanine 48 (corresponding to the conserved N-terminal alanine of all the sequenced eubacterial S13 polypeptides), sugges- ting that the N-terminal 47 amino acids may constitute a transit peptide which is cleaved off on entry into chloroplasts. Alanine is frequently the N-terminal amino acid of mature nuclear-encoded chloroplast r-proteins (for examples see Smooker et al. 1990; Elhag and Bourque 1992), and alanine 48 may be at the N-terminus in this case too. Given this assumption, the cDNA sequence predicts that the mature polypeptide should be 14.0 kDa, which corresponds reasonably well to the band seen in Western blots at about 14.5 kDa (Fig. 4). The putative transit-peptide sequence has fea- tures characteristic of most chloroplast transit-pep- tides: a net positive charge (+ 5), an abundance of the hydroxylated amino acids serine and threonine (19%) and the small hydrophobic amino acids alanine and valine (17%), and a stretch of ten uncharged amino acids including the dipeptide methionine-alanine at the amino-terminus (Keegstra et al. 1989; yon Heijne et al. 1989).
The similarity of the amino-acid sequence with eu- bacterial homologs is striking, far exceeding the sim- ilarity with its cytosolic equivalent, recently cloned from Arabidopsis (Van Lijsebettens et al. 1994). Trans- fer of sequences from organelles to the nucleus, fol- lowed by the acquisition of expression signals and the in-frame transit peptide sequence necessary to target protein products back to the organelle, has now been observed several times (reviewed in Brennicke et al. 1993; Subramanian 1993). Hypotheses explaining such organelle-to-nucleus transfers have been discussed (Sun and Callis 1993) and evidence suggests that intracellu- lar gene transfer is an ongoing process (Baldauf and Palmer 1990; Gantt et al. 1991). Exon shuffling (Gilbert 1987) may play a role in re-assembling transferred sequences into functional genes (Gantt et al. 1991; Nugent and Palmer 1991), as exemplified by the fre-
quent examples where an intron separates the exon encoding the transit peptide from the exon(s) encoding the catalytic domain(s), both for mitochondrial (Lagac6 et al. 1987; Tsuzuki et al. 1987; Dyer et al. 1989; Hass et al. 1989; Nugent and Palmer 1991) and chloroplast proteins (Wolter et al. 1988; Gantt et al. 1991), although the intron is rarely placed exactly at the cleavage site. In the case of the gene described here there is also an intron which separates the sequences encoding the transit-peptide domain from the rest of the gene, imply- ing that exon shuffling has occurred at some point.
The absence of a gene encoding S13 from the chloro- plast genomes of tobacco (Shinozaki et al. 1986), liver- wort (Ohyama et al. 1988), rice (Hiratsuka et al. 1989), maize (Subramanian 1993) and black pine (Wakasugi et al. 1994) probably also indicates its presence in the nucleus of these species. In E. coli, the gene for S13 is clustered with those of Sll , $4, L17 and the alpha subunit of RNA polymerase (rpoA) in the alpha operon (Bedwetl et al. 1985). Out of these four clustered r-protein genes, only the Sl l gene is associated with rpoA in the chloroplast genomes mentioned above; the $4 gene is located elsewhere and the genes for S13 and L17 are absent. The gene for L17, like that for S13, has perhaps been translocated to the nucleus. The similar arrangement for these genes in all chloroplast genomes studied to-date (including the bryophyte Marchantia) suggests that the transfer of the S13 gene to the nucleus and subsequent loss from the chloroplast genome took place very early, before the evolution of flowering plants.
In contrast, many plant mitochondrial genomes in- cluding those of maize, tobacco, wheat, Daucus, Oenothera and Marchantia (Bland et al. 1986; Bonen 1987; Schuster and Brennicke 1987; Wissinger et al. 1990; Takemura et al. 1992), contain a gene coding for the mitochondrial S13. Its absence from the mitochon- drial genomes of broad bean, pea, mung bean, common bean, and five crucifers (B. rapa, Raphanus sativus, Crambe abyssinica, A. thaliana and Capsella bursa-pas- toris) has also been reported (Bland et al. 1986; Nugent and Palmer 1993; Klein et al. 1994), implying two independent and relatively recent gene losses. The pres- ence of a distinct signal on Western blots of bean mitochondrial proteins, using an antibody against plant mitochondrial S13 at a position comparable to that seen with maize and tobacco, suggests that this gene has been transferred to the nucleus of legumes (Fig. 6). This antibody recognises mitochondrial pro- tein from rapeseed and Arabidopsis very weakly or not at all, suggesting the possibility that either the hypo- thetical nuclear gene has diverged sufficiently from its mitochondrial progenitor so that the polypeptides no longer contain common epitopes or that the mito- chondrial S13 function has been replaced by another protein. One instance where a prokaryotic-type chloro- plast r-protein has been replaced by its eukaryotic cytosolic homolog has been reported (Bubunenko et al.

351
kDa 1 2 3 4 5 6 7 8
Fig. 6 Western-blot analysis of mitochondrial $13. The samples are mitochondrial proteins (approximately 4 gg) from maize (1), B. napus (7), and bean (8), and mitochondrial ribosomal proteins (ap- proximately 1 gg) from maize (2), tobacco (3), wheat (4), bean (5) and B. napus (6). The sizes of the molecular-weight markers are indicated in kDa. The band corresponding to S13 migrates at about 13 kDa
1994), b u t i t w o u l d seem m o r e l ike ly t ha t m i t o c h o n d r i a l r - p r o t e i n s w o u l d be r e p l a c e d b y c h l o r o p l a s t h o m o l o g s a n d vice versa ; it has been s h o w n t h a t one l e a d e r p e p t i d e is c a p a b l e o f d i r ec t ing p r o t e i n s to b o t h or - gane l les ( H u a n g et al. 1990). H o w e v e r , the resul t s in the p r e s e n t p a p e r c lea r ly s h o w t h a t c h l o r o p l a s t S13 is n o t d e t e c t a b l e in cruci fer m i t o c h o n d r i a , a n d thus the o r ig in of the m i t o c h o n d r i a l S13 in crucifers r e m a i n s an en igma .
Acknowledgements We are grateful to Drs. Charles S. Levings III and Koh-ichi Kadowaki for mitochondrial S 13 antibody. We thank Drs J.T. Mulligan and Jeff Dangl for an aliquot of the Arabidosis genomic library, and Dr. M. Renard for B. rapa seeds. We also thank Olivier Grandjean for assistance with photography and Marie Marchand for her help during the present study.
References
Baldauf SL, Palmer JD (1990) Evolutionary transfer of the chloro- plast tufA gene to the nucleus. Nature 344:262-265
Bedwell D, Davis G, Gosink M, Post L, Nomura M, Kestler H, Zengel JM, Lindahl (1985) Nucleotide sequence of the alpha ribosomal protein operon of Escherichia coli. Nucleic Acids Res 13:3891-3903
Bland MM, Levings CS III, Matzinger DF (1986) The tobacco mitochondrial ATPase subunit 9 gene is closely linked to an open reading frame for a ribosomal protein. Mol Gen Genet 204:8 16
Bonen L (1987) The mitochondrial S13 ribosomal protein gene is silent in wheat embryos and seedlings. Nucleic Acids Res 15:10393-10404
Bradford MM (1976) A rapid and sensitive method for the quantitat- ion of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248-254
Brennicke A, Grohmann L, Hiesel R, Knoop V, Schuster W (1993) The mitochondrial genome on its way to the nucleus: different stages of gene transfer in higher plants. FEBS Lett 325:140-145
Bubunenko MG, Schmidt J, Subramanian AR (1994) Protein substi- tution in chloroplast ribosome evolution. A eukaryotic cytosolic protein has replaced its organelle homologue (L23) in spinach. J Mol Biol 240:28-41
Capel MS, Bourque DP (1982) Characterization of Nicotiana tabacum chloroplast and cytoplasmic ribosomal proteins. J Biol Chem 257:7746-7755
Damerval C, Vienne de D, Zivy M, Thiellement H (1986) Technical improvements in two-dimensional electrophoresis increase the
level of genetic variation detected in wheat-seedling proteins. Electrophoresis 7:52-54
Dellaporta SL, Wood J, Hicks JB (1983) A plant DNA miniprepara- tion:version II. Plant Mol Biol Rep 1:19-21
Dijk J, Littlechild J, Garrett RA (1977) The RNA binding properties of 'native' protein protein complexes isolated from the Es- cherichia coli ribosome. FEBS Lett 77:295 300
Douce R (1985) Mitochondria in higher plants: structure, function, and biogenesis. Academic Press, New York
Dyer MR, Gay NJ, Powell SJ, Walker JE (1989) ATP synthase from bovine mitochondria: complementary DNA sequence of the mitochondrial import precursor of the ~/-subunit and the genomic sequence of mature protein. Biochemistry 28:3670 3680
Elhag GA, Bourque DP (1992) Nuclear-encoded chloroplast ribosomal protein L27 of Nicotiana tabacum: cDNA sequence and analysis of mRNA and genes. Biochemistry 31:6856-6864
Gantt JS, Baldauf SL, Calie PJ, Weeden NF, Palmer JD (1991) Transfer of rpl22 to the nucleus greatly preceded its loss from the chloroplast and involved the gain of an intron. EMBO J 10:3073-3078
Genetics Computer Group (1991) Program manual for the GCG package (Version 7). Madison, Wisconsin, USA
Gilbert W (1987) The exon theory of genes. Cold Spring Harbor Symp Quant Biol 52:901 905
Gray MW (1992) The endosymbiont hypothesis revisited. Int Rev Cytol 141:233-357
Haas RC, Korenfeld C, Zhang Z, Perryman B, Roman D, Strauss AW (1989) Isolation and characterization of the gene and cDNA encoding human mitochondrial creatine kinase. J Biol Chem 264:2890 2897
Heijne yon G, Steppuhn J, Herrmann RG (1989) Domain structure of mitochondrial and chloroplast peptides. Eur J Biochem 180:535-545
Hiratsuka J, Shimada H, Whittier R, Ishibashi T, Sakamoto M, Mori M, Kondo C, Honji Y, Sun C-R, Meng B-Y, Li Y-Q, Kanno A, Nishizawa Y, Hirai A, Shinozaki K, Sugiura M (1989) The complete sequence of the rice (Oryza sativa) chloroplast genome: intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol Gen Genet 217:185-194
H/Sfte H, Desprez T, Amselem J, Chiapello H, Caboche M, Moisan A, Jourjon M-F, Charpenteau J-L, Berthomieu P, Guerrier D, Giraudat J, Quigley F, Thomas F, Yu D-Y, Mache R, Raynal M, Cooke R, Grellet F, Delseny M, Parmentier Y, de Marcillac G, Gigot C, Fleck J, Philipps G, Axelos M, Bardet C, Tremousaygue D, Lescure B (1993) An inventory of 1152 expressed sequence tags obtained by partial sequencing of cDNAs from Arabidopsis thaIiana. Plant J 4:1051-1061
Huang J, Hack E, Thornburg RW, Myers AM (1990) A yeast mitochondrial leader peptide functions in vivo as a dual targeting signal for both chloroplasts and mitochondria. Plant Cell 2:1249-1260
Keegstra K, Olsen LJ, Theg SM (1989) Chloroplastic precursors and their transport across the envelope membranes. Annu Rev Plant Physiol Plant Mol Biol 40:471-501
Klein M, Eckert-Ossenkopp U, Schmiedeberg I, Brandt P, Unseld M, Brennicke A, Schuster W (1994) Physical mapping of the mitochondrial genome of Arabidopsis thaliana by cosmid and YAC clones. Plant J 6:447-455
Kumar R (1995) Mitochondrial ribosomes and their proteins. In: Levings CS III, Vasil IK (eds) Molecular biology of the mitochondria. Kluwer Academic Publishers, The Netherlands (in press)
Kumar R, Lelu M-A, Small ID (1995) Purification of mitochondria and mitochondrial nucleic acids from embryogenic suspension cultures of a gymnosperm, Larix x leptoeuropaea. Plant Cell Rep 14:534-538
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685

352
Lagac6 M, Howell BW, Burak R, Lusty CJ, Shore GC (1987) Rat carbamyl-phosphate synthetase I gene. Promoter sequence and tissue-specific transcriptional regulation in vitro. J Biol Chem 262:10415-10418
Lagrange T, Franzetti B, Axelos M, Mache R, Lerbs-Mache S (1993) Structure and expression of the nuclear gene coding for the chloroplast ribosomal protein L21: developmental regulation of a housekeeping gene by alternative promoters. Mol Cell Biol 13:2614 2622
Levy S, Avni D, Hariharan N, Perry RP, Meyuhas O (1991) Oligopyrimidine tract at the 5' end of mammalian ribosomal protein mRNAs is required for their translational control. Proc Natl Acad Sci USA 88:3319 3323
Logemann J, Schell J, Willmitzer L (1987) Improved method for the isolation of RNA from plant tissues. Anal Biochem 163:16-20
Ltitcke HA, Chow KC, Mickel FS, Moss KA, Kern HF, Scheele GA (1987) Selection of AUG initiation codons differs in plants and animals. EMBO J 6:43 48
Nomura M (1990) History of ribosome research: a personal account. In: Hill WE, Dahlberg A, Garrett RA, Moore PB, Schlessinger D, Warner JR (eds) The ribosome: structure, function and evolu- tion. American Society for Microbiology, Washington D.C., pp3 55
Nugent JM, Palmer JD (1991) RNA-mediated transfer of the gene coxII from mitochondrion to the nucleus during flowering plant evolution. Cell 66:473-481
Nugent JM, Palmer JD (1993) Evolution of gene content and gene organization in flowering plant mitochondrial DNA: a general survey and further studies on coxII gene transfer to the nucleus. In: Brennicke A, Kiick U (eds) Plant mitochondria with emphasis on RNA editing and cytoplasmic male sterility. VCH, Germany, pp 163-170
Ohyama K, Fukuzawa H, Kohchi T, Sano T, Sano S, Shirai H, Umesono K, Shiki Y, Takeuchi M, Chang Z, Aota S, Inokuchi H, Ozeki H (1988) Structure and organization of Marchantia poly- morpha chloroplast genome. I. Cloning and gene identification. J Mol Biol 203:281-298
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
Schuster W, Brennicke A (1987) Plastid DNA in the mitochondrial genome of Oenothera: intra- and inter-organellar rearrangements involving part of the plastid ribosomal cistron. Mol Gen Genet 210:44-51
Schwarzbauer J, Craven GR (1985) Evidence that E. coli ribosomal protein S13 has two separable functional domains involved in 16 s RNA recognition and protein S19 binding. Nucleic Acids Res 13:6767-6786
Shinozaki K, Ohme M, Tanaka M, Wakasugi T, Hayashida N, Matsubayashi T, Zaita N, Chunwongse J, Obokata J, Yamaguchi-Shinozaki K, Ohto C, Torazawa K, Meng BY, Sugita M, Deno H, Kamogashira T, Yamada K, Kusuda J, Takaiwa F, Kato A, Tohdoh N, Shimada H, Sugiura M (1986) The complete nucleotide sequence of the tobacco chloroplast genome: its gene organization and expression. EMBO J 5:2043-2049
Smooker PM, Kruft V, Subramanian AR (1990) A ribosomal pro- tein is encoded in the chloroplast DNA in a lower plant but in the nucleus in angiosperms. J Biol Chem 265:16699 16703
Subramanian AR (1993) Molecular genetics of chloroplast ribosomal proteins. Trends Biochem Sci 18:177-181
Subramanian AR, Stahl D, Prombona A (1991) Ribosomal proteins, ribosomes, and translation in plastids. In: Bogorad L, Vasil IK (eds) The molecular biology of plastids: cell culture and somatic cell genetics of plants, volume 7A. Academic Press, New York, pp 191-215
Sun C-W, Callis J (1993) Recent stable insertion of mitochondrial DNA into an Arabidopsis polyubiquitin gene by nonhomologous recombination. Plant Cell 5:97-107
Takemura M, Oda K, Yamato K, Ohta E, Nakamura Y, Nozato N, Akashi K, Ohyama K (1992) Gene clusters for ribosomal pro- teins in the mitochondrial genome of a liverwort, Marchantia polymorpha. Nucleic Acids Res 20:3199-3205
Tsuzuki T, Obaru K, Setoyama C, Shimada K (1987) Structural organization of the mouse mitochondrial aspartate aminotran- sferase gene. J Mol Biol 198:21-31
Van Lijsebettens M, Vanderhaeghen R, Block de M, Bauw G, Villarroel R, Van Montagu M (1994) ArtS18 ribosomal protein gene copy at the Arabidopsis PFL locus affects plant develop- ment by its specific expression in meristems. EMBO J 13:3378-3388
Wakasugi T, Tsudzuki J, Ito S, Nakashima K, Tsudzuki T, Sugiura M (1994) Loss of all ndh genes as determined by sequencing the entire chloroplast genome of black pine Pinus thunbergii. Proc Natl Acad Sci USA 91:9794-9798
Wegl~Shner W, Subramanian AR (1994) Multicopy GTPase center protein L12 of Arabidopsis chloroplast ribosome is encoded by a clustered nuclear gene family with the expressed members closely linked to tRNA pr~ genes. J Biol Chem 269:7330-7336
Wissinger B, Schuster W, Brennicke A (1990) Species-specific RNA editing patterns in the mitochondrial rpsl3 transcripts of Oenoth- era and Daucus. Mol Gen Genet 224:389 395
Wolter FP, Fritz CC, Willmitzer L, Schell J, Schreier PH (1988) rbcS genes in Solarium ruberosum: conservation of transit peptide and exon shuffling during evolution. Proc Natl Acad Sci USA 85:846 850








![[20] Methods for Obtaining and Analyzing Whole …cwd.huck.psu.edu/pdf/meth_enz_2005.pdfgene expression, with the remainder being transfer RNA or ribosomal [20] analyzing chloroplast](https://img.pdfslide.us/doc/110x75/5f110e32e5a4ab5b9255a7cd/20-methods-for-obtaining-and-analyzing-whole-cwdhuckpsuedupdfmethenz2005pdf.jpg)




















