Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Jan. 2007, p. 76–82 Vol. 189, No. 10021-9193/07/$08.00�0 doi:10.1128/JB.00788-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Characterization of the Moraxella catarrhalis Opa-Like Protein, OlpA,Reveals a Phylogenetically Conserved Family of
Outer Membrane Proteins�
Michael J. Brooks,1 Cassie A. Laurence,2 Eric J. Hansen,2 and Scott D. Gray-Owen1*Department of Medical Genetics and Microbiology, University of Toronto, Toronto, Canada M5S 1A8,1 and Department of
Microbiology, University of Texas Southwestern Medical Center, Dallas, Texas 753902
Received 1 June 2006/Accepted 5 October 2006
Moraxella catarrhalis is a human-restricted pathogen that can cause respiratory tract infections. In thisstudy, we identified a previously uncharacterized 24-kDa outer membrane protein with a high degree ofsimilarity to Neisseria Opa protein adhesins, with a predicted �-barrel structure consisting of eight antiparallel�-sheets with four surface-exposed loops. In striking contrast to the antigenically variable Opa proteins, theM. catarrhalis Opa-like protein (OlpA) is highly conserved and constitutively expressed, with 25 of 27 strainscorresponding to a single variant. Protease treatment of intact bacteria and isolation of outer membranevesicles confirm that the protein is surface exposed yet does not bind host cellular receptors recognized byneisserial Opa proteins. Genome-based analyses indicate that OlpA and Opa derive from a conserved familyof proteins shared by a broad array of gram-negative bacteria.
Moraxella catarrhalis is an obligate parasite of humans thatcan cause disease in both the upper and lower respiratorytracts. In the upper respiratory tract, M. catarrhalis is respon-sible for cases of sinusitis and is the third leading cause of otitismedia in infants, after only Haemophilus influenzae and Strep-tococcus pneumoniae (5, 10, 18, 24). In the lower respiratorytract, M. catarrhalis can cause respiratory infections in adultswith chronic obstructive pulmonary disease, resulting in in-creased morbidity and mortality of these patients (27, 28). M.catarrhalis is thought to be responsible for 2 to 4 million infec-tious exacerbations of chronic obstructive pulmonary diseasein the United States each year (25), and there is significant costin treating disease related to M. catarrhalis, which is compli-cated in part by a rise in antibiotic-resistant strains (2, 30). Thismakes development of a vaccine an exciting and importantgoal.
The mechanisms involved in pathogenesis and virulence ofM. catarrhalis remain poorly defined. Attachment to host mu-cosal surfaces is an important step in colonization. Recently,M. catarrhalis genes encoding the type IV pilus were identifiedand characterized (23), leading to speculation that this struc-ture mediates initial attachment to mucosal epithelia in a man-ner similar to that of the closely related pathogenic Neisseriaspp. (9). M. catarrhalis can also interact with the A549 humanlung epithelial carcinoma cell line through the bacterium’souter membrane protein CD (16) and Hag (17) adhesins. An-other adhesin, UspA1, promotes attachment to Chang humanconjunctiva-derived epithelial cells (21) and was recentlyshown to mediate adherence via the host cellular receptorCEACAM1 (15). Human CEACAM1 is also engaged by theadhesins of other bacteria, including the Haemophilus influen-
zae P5 protein (14) and the Opa proteins of pathogenic andcommensal Neisseria spp. (12, 29, 35). Remarkably, the UspA1,P5, and Opa proteins are not related, indicating a clear exam-ple of convergent evolution by these various human-restrictedbacteria.
Considering the importance of the Opa family of outermembrane proteins in neisserial pathogenesis, we sought todetermine whether homologues of neisserial Opa proteins ex-ist in M. catarrhalis. A genome-wide screen of M. catarrhalisrevealed a single gene with high homology to the neisserialOpa proteins. Herein, we characterize this previously uniden-tified 24-kDa Opa-like protein A of M. catarrhalis (OlpA) andestablish it as an outer membrane protein that is highly con-served among clinical strains. Based upon broader analyses, wealso reveal that OlpA is a member of a highly conserved familyof proteins conserved across a wide variety of gram-negativebacteria.
MATERIALS AND METHODS
Bacterial strains and culture conditions. The M. catarrhalis strains employedin this study are listed in Table 1. Several of these strains, including O35E (1),O46E (21), TTA37 (21), 7169 (22), and 4223 (19), have been described previ-ously. For preparation of whole-cell lysates, M. catarrhalis strains were grown at37°C on brain heart infusion (Difco/Becton Dickinson, Sparks, MD) agar platesin an atmosphere of 5% CO2 or in liquid broth. Escherichia coli strains employedin this study were strain EPI300 with plasmids indicated under “Cloning thegene” below. E. coli strains were grown in Luria-Bertani (LB) agar plates orbroth with standard antibiotic supplementation as required.
General DNA methods. M. catarrhalis genomic DNA was isolated from agarplate-grown cells by using the Easy-DNA kit (Invitrogen, Carlsbad, CA) withminor modifications. The preparation of plasmids and purification of PCR prod-ucts were performed using kits manufactured by QIAGEN (Santa Clarita, CA).
DNA sequencing and analysis. The M. catarrhalis strain ATTC 43617 genomesequence was previously published (NCBI patent number WO0078968). Thissequenced genome was scanned using Gene Tool (Biotools Inc.) in all openreading frames to search for proteins with homology to neisserial Opa proteinsequences. The identified sequence was submitted to BLASTP (Entrez-PubMed)to examine it for homologous proteins. Where indicated, DNA fragments weresequenced by the York University DNA Sequencing Facility (http://www.biol.yorku.ca/cm/autoseq.htm), using the same primers. Mult-Alin (8) was used to
* Corresponding author. Mailing address: Department of Medical Ge-netics and Microbiology, Room 4381, Medical Sciences Building, 1 King’sCollege Circle, Toronto, Ontario M5S 1A8, Canada. Phone: (416) 946-5307. Fax: (416) 978-6885. E-mail: [email protected].
� Published ahead of print on 13 October 2006.
76
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

generate sequence alignments. Two-dimensional (2D) and 3D modeling wasperformed using 3D-JIGSAW (3, 4, 6). Neighbor-joining tree diagrams weredesigned using ClustalW (http://clustalw.genome.jp/)-based sequence alignmentsand Phlyodraw v0.82.
Cloning the gene. PCR primers Sgo114 (AACCCAATGCCACAAGGAC)and Sgo115 (AGCATTTGGACTGTGTGGG) were designed using the putativeM. catarrhalis olpA gene identified on the basis of our analyses of the M.catarrhalis genome (NCBI patent number WO0078968). A PCR product wasamplified using primers Sgo114 and Sgo115 from chromosomal DNAs of strains035E, 046E, ATTC 25238, 7169, 4223, and TTA37 and then cloned into thepCC1 (Blunt Cloning-Ready) vector (Epicenter Biotechnologies) to createpCC.035E, pCC.046E, pCC.25238, pCC.7169, pCC.4223, and pCC.TTA37, re-spectively. A kanamycin resistance cassette was obtained and cloned into pCC1to generate the pCCKan control. Plasmids were electroporated into electrocom-petent TransforMax E. coli EPI300 as described in the Epicenter CopyControlcDNA, gene, and PCR cloning kit to create OlpA-expressing E. coli strains. Theperiplasmic maltose binding protein (MBP)-encoding plasmid pMAL was ob-tained from New England Biolabs and introduced into E. coli EPI300 by elec-troporation.
Preparation of polyclonal OlpA antiserum. The translated OlpA protein se-quence from M. catarrhalis strain O35E was analyzed for hydrophilic sequencestretches by using the Kyte-Doolittle method (20). The peptide SNLEAKYNDNRPNDNKLEDK was selected and synthesized, with a cysteine residue addedto its N terminus, by the Protein Chemistry Technology Center at University ofTexas Southwestern Medical Center. This peptide was covalently coupled toImject maleimide-activated mariculture keyhole limpet hemocyanin (Pierce,Rockford, IL). A 100-�g portion of the peptide-keyhole limpet hemocyaninconjugate was mixed 50:50 with Freund’s complete adjuvant (Difco, Detroit, MI)and used to immunize five BALB/c mice. Four weeks later, these mice receiveda booster immunization with 30 �g of this conjugate mixed 1:1 with Freund’sincomplete adjuvant. These animals were euthanatized and exsanguinated ap-proximately 2 weeks later, and serum was then prepared from this blood bystandard methods.
Western blot analysis. For detection of the OlpA protein, whole-cell lysates(1) were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) in 12.5% (wt/vol) polyacrylamide separating gels, transferred toImmobilon-P membranes (Millipore, Bedford, MA), and probed with the indi-cated primary antibody. OlpA was detected using the mouse polyclonal OlpAantisera described herein. UspA1 and CopB were detected using the monoclonalantibodies 245B5 (7) and 10F3 (13), respectively. Maltose binding protein was
detected using the MBP-specific monoclonal antibody (New England Biolabs).The FbpA-specific rabbit polyclonal antiserum was generously provided by An-thony Schryvers (University of Calgary, Alberta, Canada). The secondary anti-body used was goat anti-mouse immunoglobulin G or anti-rabbit immunoglob-ulin G conjugated to horseradish peroxidase (Jackson ImmunoResearch, WestGrove, PA). Antigen-antibody complexes were visualized by chemiluminescenceby use of the Western Lightning Chemiluminescence Reagent Plus (New En-gland Nuclear, Boston, MA).
Protease treatment of intact cells. M. catarrhalis or E. coli cells were grown inbroth overnight at 37°C. Cells were pelleted and washed three times in phos-phate-buffered saline (PBS) containing 10 mM MgCl2 and 5 mM CaCl2 (PBS-Mg/Ca) and resuspended to a concentration of 1 � 109 cells/ml. Then, 250 �l ofcells was incubated with increasing proteinase K (Pharmacia) before incubationat room temperature for 10 min. The reaction was stopped by adding 10 �l of a10-mg/ml stock of phenylmethylsulfonyl fluoride (in isopropanol) to stop thereaction. The cells were then centrifuged and resuspended in PBS-Mg/Ca twice,and the final washed pellet was then boiled in SDS-PAGE sample buffer con-taining �-mercaptoethanol.
Heat modifiability. Bacterial cultures were grown overnight at 37°C. Cells werethen pelleted and resuspended in SDS-PAGE sample buffer. Samples were thenincubated either at room temperature or in a boiling water bath for 20 minbefore Western blot analysis.
OMVs. Outer membrane vesicles (OMVs) of M. catarrhalis were isolatedthrough heating in the presence of EDTA, as described by Murphy and Loeb(26). Briefly, 50 ml brain heart infusion broth was inoculated with M. catarrhalisand cultured overnight with shaking at 37°C. The cells were centrifuged at10,000 � g for 15 min at 4°C and the resulting pellet then resuspended in 0.05 MNa2HPO4, 0.15 M NaCl, 0.01 M EDTA (pH 7.4) before incubation at 56°C withshaking. The bacteria were removed by two consecutive spins of 10,000 � g for15 min at 4°C, and the resulting supernatant was centrifuged at 50,000 � g for 90min at 4°C to recover the outer membrane vesicles, which were then resuspendedin PBS.
Nucleotide sequence accession numbers. The olpA sequences determined inthis study were submitted via the Entrez-PubMed protein/nucleotide database,and accession numbers are listed in Table 2.
RESULTS
Identification of OlpA. Given the close relationship betweenM. catarrhalis and the Neisseria spp., we sought to determinewhether proteins related to the neisserial Opa proteins arepresent in M. catarrhalis. A search of the M. catarrhalis ATCC43617 genome was conducted using Gene Tool software bysearching with a combination of neisserial Opa protein anti-genic variants. The screen identified an open reading frameencoding a predicted protein with a high degree of sequenceidentity to neisserial Opa proteins. The predicted protein is234 amino acids in length with a possible leader peptide of 30amino acids. Oligonucleotide primers Sgo114 and Sgo115 (seeMaterials and Methods) were designed based on this predictedopen reading frame and used to screen a variety of M. catarrha-lis laboratory strains by PCR. A single fragment of �1.6 kb insize was amplified from each of the strains and then sequenced.GenBank accession numbers for each sequence are listed in
TABLE 1. M. catarrhalis strains used in this study
Strain Source
ATCC 43617...............................ATCCO35E ...........................................John D. Nelson, TexasO46E ...........................................John D. Nelson, TexasTTA37.........................................Steven L. Berk, Tennessee7169 .............................................Anthony A. Campagnari, New YorkATCC 25238...............................ATCC4223 .............................................Timothy F. Murphy, New YorkV1171 ..........................................Frederick Henderson, North CarolinaV1156 ..........................................Frederick Henderson, North CarolinaV1145 ..........................................Frederick Henderson, North CarolinaV1120 ..........................................Frederick Henderson, North CarolinaV1118 ..........................................Frederick Henderson, North CarolinaV1069 ..........................................Frederick Henderson, North CarolinaP44...............................................Steven L. Berk, TennesseeFR3227........................................Richard J. Wallace, TexasFR2336........................................Richard J. Wallace, TexasFR2213........................................Richard J. Wallace, TexasFIN2406 ......................................Merja Helminen, FinlandFIN2344 ......................................Merja Helminen, FinlandFIN2341 ......................................Merja Helminen, FinlandFIN2265 ......................................Merja Helminen, FinlandETSU-26 .....................................Steven L. Berk, TennesseeETSU-22 .....................................Steven L. Berk, TennesseeETSU-17 .....................................Steven L. Berk, TennesseeETSU-5 .......................................Steven L. Berk, TennesseeB59911 ........................................David Goldblatt, EnglandB59504 ........................................David Goldblatt, England
TABLE 2. Accession numbers of olpA genes from strainsused for cloning
M. catarrhalis strain olpA accession no.
035E ...............................................................................DQ996458046E ...............................................................................DQ99645925238 ..............................................................................DQ9964607169 ................................................................................DQ9964614223 ................................................................................DQ996462TTA37............................................................................DQ996463V1156 .............................................................................DQ996464
VOL. 189, 2007 CHARACTERIZATION OF M. CATARRHALIS OlpA 77
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
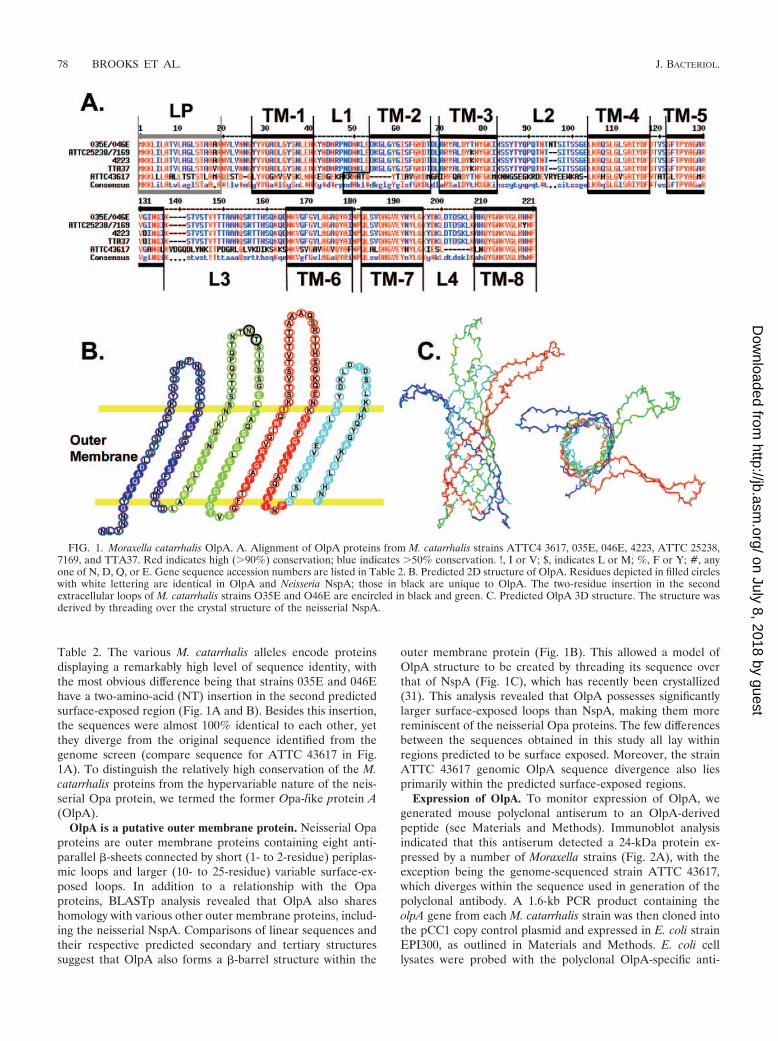
Table 2. The various M. catarrhalis alleles encode proteinsdisplaying a remarkably high level of sequence identity, withthe most obvious difference being that strains 035E and 046Ehave a two-amino-acid (NT) insertion in the second predictedsurface-exposed region (Fig. 1A and B). Besides this insertion,the sequences were almost 100% identical to each other, yetthey diverge from the original sequence identified from thegenome screen (compare sequence for ATTC 43617 in Fig.1A). To distinguish the relatively high conservation of the M.catarrhalis proteins from the hypervariable nature of the neis-serial Opa protein, we termed the former Opa-like protein A(OlpA).
OlpA is a putative outer membrane protein. Neisserial Opaproteins are outer membrane proteins containing eight anti-parallel �-sheets connected by short (1- to 2-residue) periplas-mic loops and larger (10- to 25-residue) variable surface-ex-posed loops. In addition to a relationship with the Opaproteins, BLASTp analysis revealed that OlpA also shareshomology with various other outer membrane proteins, includ-ing the neisserial NspA. Comparisons of linear sequences andtheir respective predicted secondary and tertiary structuressuggest that OlpA also forms a �-barrel structure within the
outer membrane protein (Fig. 1B). This allowed a model ofOlpA structure to be created by threading its sequence overthat of NspA (Fig. 1C), which has recently been crystallized(31). This analysis revealed that OlpA possesses significantlylarger surface-exposed loops than NspA, making them morereminiscent of the neisserial Opa proteins. The few differencesbetween the sequences obtained in this study all lay withinregions predicted to be surface exposed. Moreover, the strainATTC 43617 genomic OlpA sequence divergence also liesprimarily within the predicted surface-exposed regions.
Expression of OlpA. To monitor expression of OlpA, wegenerated mouse polyclonal antiserum to an OlpA-derivedpeptide (see Materials and Methods). Immunoblot analysisindicated that this antiserum detected a 24-kDa protein ex-pressed by a number of Moraxella strains (Fig. 2A), with theexception being the genome-sequenced strain ATTC 43617,which diverges within the sequence used in generation of thepolyclonal antibody. A 1.6-kb PCR product containing theolpA gene from each M. catarrhalis strain was then cloned intothe pCC1 copy control plasmid and expressed in E. coli strainEPI300, as outlined in Materials and Methods. E. coli celllysates were probed with the polyclonal OlpA-specific anti-
FIG. 1. Moraxella catarrhalis OlpA. A. Alignment of OlpA proteins from M. catarrhalis strains ATTC4 3617, 035E, 046E, 4223, ATTC 25238,7169, and TTA37. Red indicates high (�90%) conservation; blue indicates �50% conservation. !, I or V; $, indicates L or M; %, F or Y; #, anyone of N, D, Q, or E. Gene sequence accession numbers are listed in Table 2. B. Predicted 2D structure of OlpA. Residues depicted in filled circleswith white lettering are identical in OlpA and Neisseria NspA; those in black are unique to OlpA. The two-residue insertion in the secondextracellular loops of M. catarrhalis strains O35E and O46E are encircled in black and green. C. Predicted OlpA 3D structure. The structure wasderived by threading over the crystal structure of the neisserial NspA.
78 BROOKS ET AL. J. BACTERIOL.
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

serum to confirm that the recombinant protein was expressedin E. coli (Fig. 2B). The OlpA protein was expressed in E. coliwithout any induction, indicating that expression is likely con-stitutively driven by its native M. catarrhalis promoter (Fig. 2Band data not shown).
Conservation of OlpA among M. catarrhalis strains. Consid-ering that the cloned olpA gene sequences were highly con-served among the M. catarrhalis strains tested in this study, it isremarkable that the strain ATTC 43617 genome sequence-derived allele is so distinct. Since the OlpA antiserum distin-guishes between these two variants, we collected M. catarrhalisstrains from a variety of geographic locations (Table 1) andprobed bacterial lysates by immunoblotting. In total, 25 of the27 strains tested during this study reacted with the M. catarrha-lis antibody (Fig. 2A and C), with only strains ATTC 43617 andV1156 displaying no cross-reactive OlpA. To understand therelationship between these two proteins, we sequenced theV1156 allele. As expected, the V1156 OlpA (GenBank acces-sion no. DQ996464) is closely related to that of ATTC 43617,suggesting that these two represent an uncommon second vari-ant of OlpA.
Surface accessibility and heat modifiability. Our structuralanalyses suggested that OlpA is a surface-exposed outer mem-brane protein (Fig. 1B and C). Consistent with its specificity fora peptide sequence predicted to lie within the transmembranesequence (Fig. 1B), the OlpA-specific antiserum did not reactwith intact bacteria (data not shown). To localize the OlpAprotein, M. catarrhalis strain 035E was treated with heat andEDTA to liberate OMVs (26). Immunoblot analysis confirmedthat the OMV preparations contained OlpA and other well-characterized outer membrane proteins, including CopB andUspA1 (Fig. 3A), while the abundant periplasmic ferric bind-ing protein (FbpA) was barely detectable (data not shown),consistent with an outer membrane localization of OlpA. Toconfirm exposure at the bacterial cell surface, whole-cell ly-sates from recombinant OlpA-expressing E. coli and M.catarrhalis that had been treated with proteinase K were sub-jected to immunoblot analysis. Protease treatment caused adose-dependent degradation of OlpA without affecting the
integrity of the periplasmic maltose binding protein (MbpA) orFbpA (Fig. 3B), confirming surface exposure of OlpA.
The �-barrel structure of many gram-negative outer mem-brane proteins, including the neisserial Opa proteins, tends toresist unfolding in SDS-containing sample buffer unless theyare boiled. The electrophoretic mobility of OlpA was affectedby heating (Fig. 3C), consistent with a �-barrel conformation.The heat-modifiable nature of recombinant OlpA expressed inE. coli is indistinguishable from that observed in the respectiveM. catarrhalis strains, suggesting that the expression and fold-ing of OlpA in E. coli are comparable to those of its nativeform.
M. catarrhalis OlpA does not bind to CEACAM recep-tors. The M. catarrhalis UspA1 protein attaches to humanCEACAM receptors (15). Given the similarity between OlpAand the neisserial Opa proteins, we sought to determinewhether these Opa homologues might represent anothermechanism by which M. catarrhalis binds to CEACAM and/orother host receptors. E. coli expressing the OlpA from strain046E or ATTC 25238 did not interact with either solubleCEACAM1 or CEACAM5 in either bacterial pull-down orsolid-phase binding assays, while E. coli expressing the Neis-seria gonorrhoeae Opa57 protein bound both proteins (datanot shown). Moreover, a UspA1-deficient derivative of M.catarrhalis 035E does not bind to CEACAM-expressing celllines (data not shown) despite expressing OlpA (Fig. 2 and 3).We were also unable to detect any difference in heparin bind-ing between the recombinant OlpA-expressing E. coli and theparental strain, suggesting that OlpA does not function as aheparan sulfate proteoglycan-specific adhesin in a manner sim-ilar to the related N. gonorrhoeae Opa50 (32), the Neisseriameningitidis OpcA (34), or the enterotoxigenic E. coli Tia pro-tein (11). To ascertain whether any adhesin function could beattributed to OlpA in M. catarrhalis, we generated an olpA-deficient strain of M. catarrhalis O35E by insertional mutagen-esis. Next, we performed in vitro infection assays to comparewild-type, UspA1-deficient, and OlpA-deficient M. catarrhalis035E, as well as the OlpA-expressing versus parental (emptyexpression vector-containing) E. coli strains. These studies pro-
FIG. 2. Detection of OlpA protein by immunoblotting. Western blots of bacterial lysates were probed with the OlpA-specific antiserum. A. M.catarrhalis cell lysates. B. Cell lysates from recombinant E. coli strains expressing either OlpA cloned from the indicated M. catarrhalis strains ora kanamycin cassette (Kan) as a negative control. C. Immunoblot analysis to detect reactivity with M. catarrhalis strain O35E-specific OlpAantiserum with lysates prepared from the indicated strains. Molecular mass position markers (in kilodaltons) are shown on the left.
VOL. 189, 2007 CHARACTERIZATION OF M. CATARRHALIS OlpA 79
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

vided no evidence that OlpA facilitates bacterial associationwith HeLa (human endocervical), A549 (human lung-derived),Caco-2 (human colon-derived), or Lec11 (Chinese hamsterovary-derived) cell lines, in either the presence or absence ofserum (data not shown).
Homologues. Considering the presence of the OlpA in M.catarrhalis and the presence of the closely related Opa andNspA proteins in Neisseria spp., we next sought to determinethe phylogenetic distribution of Opa-like proteins via mining ofavailable genome sequences. Interestingly, H. influenzae, whichalso exclusively colonizes the human upper respiratory tractand causes a localized and disseminated infection, encodes aprotein that has significant sequence identity to OlpA but other-
wise remains uncharacterized (accession no. ZP_00154421) (Fig.4). Other respiratory pathogens, such as Actinobacillus succino-genes and Pasteurella multocida, also appear to have highly relatedhomologues (Fig. 4). Each of these Opa-like proteins is moreclosely related to the M. catarrhalis OlpA than to the neisserialOpa proteins (Fig. 4A). Related proteins are also evident inthe Enterobacteriaceae, including a homologue present in en-terotoxigenic E. coli that functions as an adhesin involved inpathogenesis (11).
DISCUSSION
In searching for homologues of the well-characterized Opaprotein adhesins from the pathogenic Neisseria spp., we haveidentified a previously uncharacterized but highly conservedouter membrane protein in M. catarrhalis. While the OlpAprotein shares a high degree of similarity with neisserial Opaproteins, we found no evidence that they share a similar func-tion. While neisserial Opa proteins bind to CEACAM and/orHSPG receptors (12, 32, 33), neither OlpA-expressing butUspA1-deficient M. catarrhalis nor the OlpA-expressing E. colidisplayed any binding to either of these receptor types. In fact,we did not detect OlpA-mediated adherence to any of a varietyof cell lines tested here.
Sequence alignments and our experimental results describedhere indicate that OlpA forms an eight-stranded �-barrel inthe M. catarrhalis outer membrane. This allowed us to predictthe 3D structure by threading over the closely related N. men-ingitidis NspA, which has been previously crystallized (31).Curiously, the OlpA proteins appear to have surface-exposedloops that are much larger than those of NspA yet shorter thanthose found in the Opa adhesins. The similarity of OlpA to thewell-characterized NspA and Opa proteins make it very likelythat these structural predictions provide an accurate depictionof OlpA in the membrane-spanning regions. However, thesoftware used cannot predict the tertiary structure within thesurface-exposed loops.
While the search for a vaccine protecting against M. catar-rhalis infection has led to the characterization of other outermembrane proteins, this is the first description of OlpA. Toensure that the OlpA gene is expressed, we generated an an-tibody against a peptide in the predicted OlpA gene and de-tected expression in M. catarrhalis strains and recombinant E.coli in which the gene is being expressed from its native pro-moter. We also confirmed that OlpA was present within outermembrane fractions, had a heat-modifiable native structurecharacteristic of � barrel structures, and was surface exposed.
The level of conservation within the OlpA genes sequencedwas remarkably high, with only two different variants apparentamong the 27 strains tests. Of note, these variants are highlyconserved among membrane-spanning sequences, with signif-icant deviation apparent only within the surface-exposed loops.However, 25 of 27 strains contained the variant typified bystrain 035E, with �99% identity among the six sequencedalleles.
Considering the lack of CEACAM and HSPG binding andthe relatively high conservation of OlpA compared to neisse-rial Opa proteins, we wondered whether similar proteins existin more distantly related bacterial species. Our survey of avail-able genome sequences indicates that there are uncharacter-
FIG. 3. A. Detection of OlpA in EDTA-induced outer membranevesicles of M. catarrhalis. OMVs prepared from either strain O35E orthe USPA1-deficient 035E mutant (O35E�U) were probed with spe-cific antibodies to detect OlpA, USPA1, and CopB. B. Surface expo-sure of OlpA. Intact E. coli expressing either OlpA or the periplasmicMBP were subjected to increasing doses of trypsin and proteinase K.Lysates were resolved and probed for either OlpA or MBP, as indi-cated. C. Heat-modifiability of OlpA was detected by using SDS-PAGE and immunoblot analysis following incubation of bacterial celllysates for 20 min at either 100°C or 23°C. Molecular mass positionmarkers (in kilodaltons) are shown on the left.
80 BROOKS ET AL. J. BACTERIOL.
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
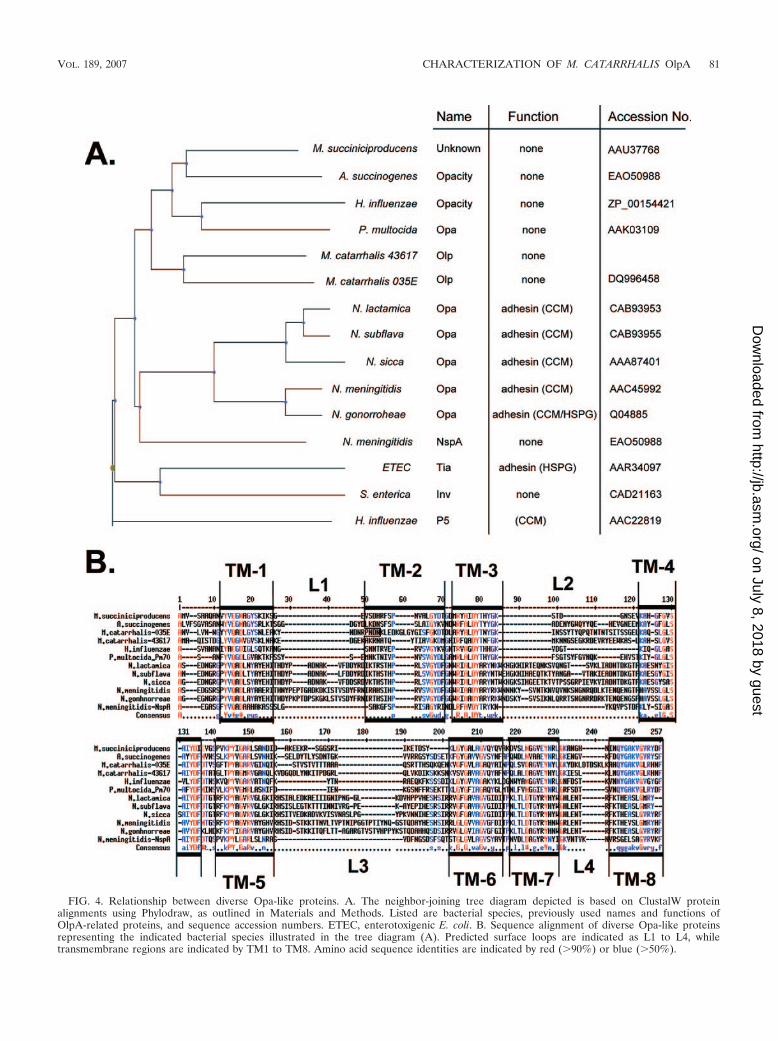
FIG. 4. Relationship between diverse Opa-like proteins. A. The neighbor-joining tree diagram depicted is based on ClustalW proteinalignments using Phylodraw, as outlined in Materials and Methods. Listed are bacterial species, previously used names and functions ofOlpA-related proteins, and sequence accession numbers. ETEC, enterotoxigenic E. coli. B. Sequence alignment of diverse Opa-like proteinsrepresenting the indicated bacterial species illustrated in the tree diagram (A). Predicted surface loops are indicated as L1 to L4, whiletransmembrane regions are indicated by TM1 to TM8. Amino acid sequence identities are indicated by red (�90%) or blue (�50%).
VOL. 189, 2007 CHARACTERIZATION OF M. CATARRHALIS OlpA 81
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ized outer membrane proteins with a high degree of similarityto OlpA in a wide variety of species (Fig. 4). Our alignmentsindicate that each of these is an outer membrane protein witheight antiparallel �-strands and four surfaced-exposed loops,and some species contain homologues in two different genomicloci. As with the M. catarrhalis OlpA variants, these homo-logues possess high sequence conservation within the trans-membrane sequences but vary in size and linear sequencewithin the surface-exposed loops. It will be interesting to es-tablish whether the strict conservation of certain residueswithin the membrane-spanning sequences reveals residues thatare required for proper folding or membrane insertion of thebasic eight-stranded �-barrel structure versus those that areevolutionarily related.
While the function of OlpA proteins remains unknown, theirconservation across phylogenetically disparate bacterial spe-cies suggests a basic essential function. This, along with the factthat the M. catarrhalis OlpA protein is surface exposed andhighly conserved, makes it an obvious vaccine candidate. Fi-nally, when considering the evolutionary relationship amongthe Opa and Opa-like proteins, it is enticing to speculate thatevolution has taken advantage of the minimal �-barrel struc-ture represented by proteins such as NspA to provide a basicframework on which host cell binding adhesins or proteinspossessing other necessary functions can be derived.
ACKNOWLEDGMENTS
This work was supported by funding from Canadian Institutes forHealth research grant no. MOP-15499 to S.D.G.-O. and U.S. PublicHealth Service grant no. AI36344 to E.J.H. S.D.G. is supported byNew Investigator Awards from the Canadian Institutes of Health Re-search and is a recipient of the Province of Ontario Premier’s ResearchExcellence Award.
We thank John Nelson, Anthony Campagnari, David Goldblatt,Richard Wallace, Steven Berk, Merja Helminen, and Frederick Hen-derson for providing the wild-type isolates of M. catarrhalis and An-thony B. Schryvers for the FbpA-specific antiserum used in this study.
REFERENCES
1. Aebi, C., I. Maciver, J. L. Latimer, L. D. Cope, M. K. Stevens, S. E. Thomas,G. H. McCracken, Jr., and E. J. Hansen. 1997. A protective epitope ofMoraxella catarrhalis is encoded by two different genes. Infect. Immun. 65:4367–4377.
2. Bandak, S. I., M. R. Turnak, B. S. Allen, L. D. Bolzon, D. A. Preston, S. K.Bouchillon, and D. J. Hoban. 2001. Antibiotic susceptibilities among recentclinical isolates of Haemophilus influenzae and Moraxella catarrhalis fromfifteen countries. Eur. J. Clin. Microbiol. Infect. Dis. 20:55–60.
3. Bates, P. A., L. A. Kelley, R. M. MacCallum, and M. J. Sternberg. 2001.Enhancement of protein modeling by human intervention in applying theautomatic programs 3D-JIGSAW and 3D-PSSM. Proteins 5(Suppl.):39–46.
4. Bates, P. A., and M. J. Sternberg. 1999. Model building by comparison atCASP3: using expert knowledge and computer automation. Proteins3(Suppl.):47–54.
5. Christensen, J. J. 1999. Moraxella (Branhamella) catarrhalis: clinical, micro-biological and immunological features in lower respiratory tract infections.APMIS 88(Suppl.):1–36.
6. Contreras-Moreira, B., and P. A. Bates. 2002. Domain fishing: a first step inprotein comparative modelling. Bioinformatics 18:1141–1142.
7. Cope, L. D., E. R. Lafontaine, C. A. Slaughter, C. A. J. Hasemann, C. Aebi,F. W. Henderson, G. H. J. McCracken, and E. J. Hansen. 1999. Character-ization of the Moraxella catarrhalis uspA1 and uspA2 genes and their encodedproducts. J. Bacteriol. 181:4026–4034.
8. Corpet, F. 1988. Multiple sequence alignment with hierarchical clustering.Nucleic Acids Res. 16:10881–10890.
9. Craig, L., M. E. Pique, and J. A. Tainer. 2004. Type IV pilus structure andbacterial pathogenicity. Nat. Rev. Microbiol. 2:363–378.
10. Faden, H., J. Hong, and T. Murphy. 1992. Immune response to outer mem-brane antigens of Moraxella catarrhalis in children with otitis media. Infect.Immun. 60:3824–3829.
11. Fleckenstein, J. M., J. T. Holland, and D. L. Hasty. 2002. Interaction of anouter membrane protein of enterotoxigenic Escherichia coli with cell surfaceheparan sulfate proteoglycans. Infect. Immun. 70:1530–1537.
12. Gray-Owen, S. D., C. Dehio, A. Haude, F. Grunert, and T. F. Meyer. 1997.CD66 carcinoembryonic antigens mediate interactions between Opa-ex-pressing Neisseria gonorrhoeae and human polymorphonuclear phagocytes.EMBO J. 16:3435–3445.
13. Helminen, M. E., I. Maciver, J. L. Latimer, L. D. Cope, G. H. McCracken,Jr., and E. J. Hansen. 1993. A major outer membrane protein of Moraxellacatarrhalis is a target for antibodies that enhance pulmonary clearance of thepathogen in an animal model. Infect. Immun. 61:2003–2010.
14. Hill, D. J., M. A. Toleman, D. J. Evans, S. Villullas, L. Van Alphen, and M.Virji. 2001. The variable P5 proteins of typeable and non-typeable Hae-mophilus influenzae target human CEACAM1. Mol. Microbiol. 39:850–862.
15. Hill, D. J., and M. Virji. 2003. A novel cell-binding mechanism of Moraxellacatarrhalis ubiquitous surface protein UspA: specific targeting of the N-domain of carcinoembryonic antigen-related cell adhesion molecules byUspA1. Mol. Microbiol. 48:117–129.
16. Holm, M. M., S. L. Vanlerberg, I. M. Foley, D. D. Sledjeski, and E. R.Lafontaine. 2004. The Moraxella catarrhalis porin-like outer membrane pro-tein CD is an adhesin for human lung cells. Infect. Immun. 72:1906–1913.
17. Holm, M. M., S. L. Vanlerberg, D. D. Sledjeski, and E. R. Lafontaine. 2003.The Hag protein of Moraxella catarrhalis strain O35E is associated withadherence to human lung and middle ear cells. Infect. Immun. 71:4977–4984.
18. Karalus, R., and A. Campagnari. 2000. Moraxella catarrhalis: a review of animportant human mucosal pathogen. Microbes Infect. 2:547–559.
19. Kyd, J. M., A. W. Cripps, and T. F. Murphy. 1998. Outer-membrane antigenexpression by Moraxella (Branhamella) catarrhalis influences pulmonaryclearance. J. Med. Microbiol. 47:159–168.
20. Kyte, J., and R. F. Doolittle. 1982. A simple method for displaying thehydropathic character of a protein. J. Mol. Biol. 157:105–132.
21. Lafontaine, E. R., L. D. Cope, C. Aebi, J. L. Latimer, G. H. McCracken, Jr.,and E. J. Hansen. 2000. The UspA1 protein and a second type of UspA2protein mediate adherence of Moraxella catarrhalis to human epithelial cellsin vitro. J. Bacteriol. 182:1364–1373.
22. Luke, N. R., and A. A. Campagnari. 1999. Construction and characterizationof Moraxella catarrhalis mutants defective in expression of transferrin recep-tors. Infect. Immun. 67:5815–5819.
23. Luke, N. R., A. J. Howlett, J. Shao, and A. A. Campagnari. 2004. Expressionof type IV pili by Moraxella catarrhalis is essential for natural competenceand is affected by iron limitation. Infect. Immun. 72:6262–6270.
24. Murphy, T. F. 1996. Branhamella catarrhalis: epidemiology, surface antigenicstructure, and immune response. Microbiol. Rev. 60:267–279.
25. Murphy, T. F., A. L. Brauer, B. J. Grant, and S. Sethi. 2005. Moraxellacatarrhalis in chronic obstructive pulmonary disease: burden of disease andimmune response. Am. J. Respir. Crit. Care Med. 172:195–199.
26. Murphy, T. F., and M. R. Loeb. 1989. Isolation of the outer membrane ofBranhamella catarrhalis. Microb. Pathog. 6:159–174.
27. Murphy, T. F., and S. Sethi. 1992. Bacterial infection in chronic obstructivepulmonary disease. Am. Rev. Respir. Dis. 146:1067–1083.
28. Murphy, T. F., and S. Sethi. 1997. A national strategy for research in chronicobstructive pulmonary disease. JAMA 277:1596.
29. Toleman, M., E. Aho, and M. Virji. 2001. Expression of pathogen-like Opaadhesins in commensal Neisseria: genetic and functional analysis. Cell. Mi-crobiol. 3:33–44.
30. Turnak, M. R., S. I. Bandak, S. K. Bouchillon, B. S. Allen, and D. J. Hoban.2001. Antimicrobial susceptibilities of clinical isolates of Haemophilus influ-enzae and Moraxella catarrhalis collected during 1999–2000 from 13 coun-tries. Clin. Microbiol. Infect. 7:671–677.
31. Vandeputte-Rutten, L., M. P. Bos, J. Tommassen, and P. Gros. 2003. Crystalstructure of neisserial surface protein A (NspA), a conserved outer mem-brane protein with vaccine potential. J. Biol. Chem. 278:24825–24830.
32. van Putten, J. P., and S. M. Paul. 1995. Binding of syndecan-like cell surfaceproteoglycan receptors is required for Neisseria gonorrhoeae entry into hu-man mucosal cells. EMBO J. 14:2144–2154.
33. Virji, M., K. Makepeace, D. J. P. Ferguson, and S. Watt. 1996. Carcinoem-bryonic antigens (CD66) on epithelial cells and neutrophils are receptors forOpa proteins of pathogenic neisseriae. Mol. Microbiol. 22:941–950.
34. Virji, M., K. Makepeace, I. R. A. Peak, D. J. P. Ferguson, M. P. Jennings,and E. R. Moxon. 1995. Opc- and pilus-dependent interactions of meningo-cocci with human endothelial cells: molecular mechanisms and modulationby surface polysaccharides. Mol. Microbiol. 18:741–754.
35. Virji, M., S. Watt, K. Barker, K. Makepeace, and R. Doyonnas. 1996. TheN-domain of the human CD66a adhesion molecule is a target for Opaproteins of Neisseria meningitidis and Neisseria gonorrhoeae. Mol. Microbiol.22:929–939.
82 BROOKS ET AL. J. BACTERIOL.
on July 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





![Moraxella (Branhamella)catarrhalis BRO β-Lactamase: a … · BRO-1 (1), we did not succeed in isolating [3H]palmitate-labeled BRO-2. Considering the lower level of BRO-2 produc-tion,](https://img.pdfslide.us/doc/110x75/5fde2baa87bc80396c591fc4/moraxella-branhamellacatarrhalis-bro-lactamase-a-bro-1-1-we-did-not-succeed.jpg)












![BMC Microbiology BioMed Central · 2017. 8. 27. · Moraxella catarrhalis is an exclusively human, mucosal res-piratory tract commensal and pathogen causing between 5% [1] and 20%](https://img.pdfslide.us/doc/110x75/60b9d1b25ab06638794a37be/bmc-microbiology-biomed-central-2017-8-27-moraxella-catarrhalis-is-an-exclusively.jpg)










