Embed Size (px)
Citation preview

Intensive Care Med (2005) 31:129–137DOI 10.1007/s00134-004-2487-y E X P E R I M E N T A L
Hans JacobsSteven N. MinkKrika DukeDeepak BoseZhao-Qin ChengSusan HowlettGregory R. FerrierR. Bruce Light
Characterization of membrane N-glycanbinding sites of lysozymefor cardiac depression in sepsis
Received: 31 March 2004Accepted: 8 October 2004Published online: 17 December 2004� Springer-Verlag 2004
Supported by the Manitoba Heart andStroke Foundation, Health Sciences CentreFoundation, and Biology of BreathingGroup Manitoba Institutes of Child Health
Electronic Supplementary MaterialSupplementary material is available in theonline version of this article at http://dx.doi.org/10.1007/s00134-004-2487-y
S. N. Mink ())Health Sciences Centre GF-221,700 William Ave, Winnipeg, Manitoba,R3E-0Z3, Canadae-mail: [email protected].: +1-204-7872914Fax: +1-204-7871932
H. JacobsDepartment of Biochemistryand Medical Genetics,University of Manitoba,Winnipeg, Manitoba, Canada
S. N. Mink · D. BoseDepartment of Pharmacologyand Therapeutics,University of Manitoba,Winnipeg, Manitoba, Canada
S. N. Mink · K. Duke · D. Bose ·Z.-Q. Cheng · R. B. LightDepartment of Internal Medicine,University of Manitoba,Winnipeg, Manitoba, Canada
D. BoseDepartment of Anesthesiology,University of Manitoba,Winnipeg, Manitoba, Canada
S. Howlett · G. R. FerrierDepartment of Pharmacology,Dalhousie University,Halifax, Nova Scotia, Canada
Abstract Purpose: In sepsis, revers-ible myocardial depression has beenascribed to the release of mediators ofinflammation. We previously foundthat lysozyme released from leuko-cytes from the spleen and other or-gans mediated myocardial depressionin an Escherichia coli model of septicshock in dogs. We hypothesize thatlysozyme binds to or cleaves a car-diac surface membrane N-glycopro-tein to cause depression. The objec-tives of the present study were: 1) todetermine whether the binding of ly-sozyme is reversible; 2) to assess theN-glycan structure to which lyso-zyme binds; 3) to examine whethernonenzymatic proteins, termed lec-tins, with a binding specificity similarto that of lysozyme could also causedepression; and 4) to assess whetherthe membrane to which lysozymebinds is affected by the enzymesprotease type XIV and collagenase A,
that are used to prepare single cellmyocyte experiments. Methods: Wemeasured isometric contraction in aright ventricular trabecular prepara-tion. Results: We found that lyso-zyme binds in a reversible manner tothe Man b(1–4) GlcNAc b(1–4)Glc-NAc moiety in the tri-mannosyl corestructure of high mannose/hybrid andtri-antennary carbohydrate classeswhere GlcNAc is N-acetylglu-cosamine and Man is mannose. Lec-tins with a specificity similar to thatof lysozyme also caused depression,and lysozyme’s depressant activitywas eliminated by protease type XIVand collagenase A. Conclusions:These results indicate that lysozymereversibly binds to a membrane gly-coprotein to cause myocardial de-pression in sepsis. We further localizeits binding site to a variant of thechitotriose structure in the tri-man-nosyl core of the membrane glyco-protein.
Keywords Myocardial depressantfactor · Lectins · Oligosaccharides ·Glycoproteins · Septic shock ·Endotoxemia · Dog

130
Introduction
Reversible myocardial depression in sepsis has been as-cribed to the release of mediators of inflammation [1, 2].In an Escherichia coli model of sepsis in dogs, we pre-viously found that continuous arterial venous hemofil-tration could reverse myocardial depression by removal ofa substance of <30,000 molecular weight from plasma [3].We used a right ventricular trabecular (RVT) preparationas a bioassay to identify the plasma fraction wheremyocardial depressant activity was contained. After var-ious separation procedures and protein sequencing tech-niques, we determined that this depressant substance waslysozyme c, consistent with that originating from thespleen [4, 5]. We further showed that specific inhibitors oflysozyme could prevent myocardial depression in in vitroand in vivo canine preparations [4, 5].
Lysozyme is a bacteriolytic enzyme originally identi-fied in secretions by Fleming [7]. It catalyzes the hy-drolysis of N-acetylglucosamine (GlcNAc) to N-acetyl-muramic acid (NAM) linkages of bacterial cell wallsthereby disrupting the bacterial cell [8, 9]. Lysozymeexists among the cells of the blood exclusively in neu-trophilic granulocytes and monocytes, whereas in tissues,the greatest concentrations are found in the bone marrow,lungs, intestines, spleen, and kidneys [6]. The increase inplasma lysozyme that occurs in sepsis is probably relatedto its release from disintegrating leukocytes from thespleen or other organs [6]. In the present study, we haveused the term Lzm-S rather than lysozyme c, since lyso-zyme identified by the sequencing analyses could havearisen from the source organ, i.e., the spleen or leukocytesarising from that organ, or possibly from other organs.
The mechanism of action of Lzm-S in causing cardiacdepression is not clear. In the heart, most surface mem-brane proteins are heavily N-glycosylated [10, 11], and inturn, changes in the N-glycosylated regions may affectmembrane function and alter myocardial contraction [11].We consider two possibilities about how Lzm-S may af-fect membrane function. In one mechanism, Lzm-S bindsto a surface glycoprotein and hydrolyses the bond be-tween two monosaccharide units that irreversibly leads tomyocardial depression. Alternatively, Lzm-S binds to anextracellulary orientated saccharide bearing structure, isunable to hydrolyze it, but because of its binding inducesa conformational change of the carbohydrate tree and theglycoprotein bearing it that mediates the biological effectof myocardial depression.
N-glycans to which Lzm-S may bind may be classifiedinto three major groups termed high-mannose, complex,and hybrid [12] (see Fig. 1 and Discussion). All threetypes contain a common structure, Mana1-6(Mana1-3)Manb1-4GlcNAcb1-4GlcNAcb1-Asn where Man ismannose and Asn is asparagine (see dotted lines inFig. 1). This common structure is called the “core” or the
tri-mannosyl core that is fairly uniform among speciesand proteins [12].
The objectives of the present study were multifold. Inour right ventricular trabecular preparation [3, 4, 5], wefirst tried to determine whether Lzm-S binds reversibly orwhether it irreversibly cleaves the N-glycan structure onthe cardiac membrane. In a second approach, we tried toassess whether the N-glycan structure to which Lzm-Sbinds represents a complex carbohydrate, a high-mannosestructure, or a variety of types. In a third part, we deter-mined whether nonenzymatic proteins termed lectins [13]that reversibly bind to the carbohydrate structure with aspecificity similar to that of Lzm-S may also causemyocardial depression. Finally, we assessed whethertreatment of RVT with enzymes [14] used to preparesingle myocytes would inhibit Lzm-S’s myocardial de-pressant effect. Such an effect would therefore limit thepotential use of the myocyte preparation in determiningLzm-S’s mechanism of action. The overall objective ofthis study was to further examine the role of Lzm-S incontributing to myocardial depression in sepsis.
Methods
These experiments were approved by the University Animal CareCommittee and conform with the Guide for the Care and Use ofLaboratory Animals published by the US National Institutes ofHealth (NIH Publication No 85–23, 1996) [15].
The right ventricular trabecular (RVT) preparation used in thisstudy has previously been described [3 ,4, 5]. Briefly, mongrel dogs(15–25 kg) were anesthetized with pentobarbital. The hearts wereremoved, flushed with 50 ml of Krebs-Henseleit solution (KH), andplaced in ice-cold KH bubbled with a mixture of 95%O2 and 5%CO2. Three to four thin trabeculae (<1 mm diameter) were obtainedfrom the right ventricle and were tied at each end with 6-O silkthread. Each thin muscle was suspended in a 5-ml vertical constanttemperature bath that contained KH. The muscle was stimulatedelectrically via punctate platinum bipolar electrodes with rectan-gular pulses (1 ms duration) at an intensity of 50% above thresholddelivered at intervals of 2,000 ms. Isometric contraction wasmeasured while the trabeculae were stretched to optimal length.Furthermore, right ventricular trabeculae rather than left ventricletrabeculae were used in this study, since it is possible to obtain agreater number of thin muscle strips from the right ventricle, andbecause it was previously shown that septic plasma caused similardecline in isometric contraction when trabeculae were obtainedfrom both ventricles [3].
The plasma concentration of Lzm-S attained in our sepsis modelis approximately 10�8 M [4], and the 10�7 M–10�9 M range wasused in the experiments described below. Lzm-S was purified fromthe spleens of nonseptic dogs as previously described [4]. Fur-thermore, when Lzm-S, lectin, or KH (see below) was added to theRVT preparation, measurements were determined at 5-min inter-vals unless otherwise indicated.
Reversibility experiments
Reversibility of Lzm-S’s myocardial depressant effect in the RVTpreparation was assessed by repeatedly washing the muscle withKH. Four groups of trabeculae were examined under differentconditions. In two of the four groups included, after baseline

131
measurements were obtained, 10�7M Lzm-S was added to the RVTpreparation and isometric contraction was measured 5 min later(treatment condition). Repeat measurements were then performedfor three consecutive intervals at 15 min apart. In group 1, nofurther intervention was performed, while in group 2 the trabeculaewere washed (1 bath exchange) with KH at the beginning of each15 min interval. The other two groups (groups 3 and 4) were controlgroups in which KH rather than Lzm-S was added to the prepara-tion in the treatment condition. In group 3, the trabeculae wererepeatedly washed with KH at 15 min intervals, while in group 4,no further intervention was performed.
Assessment of N-glycan structure to which Lzm-S binds
Regardless of the type of carbohydrate structure to which Lzm-Smay bind, it is possible to deglycosylate the carbohydrate structureby means of exoglycosidases in which a specific sugar residue isremoved from the terminal end of the carbohydrate tree [16]. Theextent to which removal of a residue is associated with a reductionin the depressant effect of Lzm-S can be used to ascertain thefunctional importance of a sugar residue. Based on the work ofprevious investigators [9], Lzm-S would preferentially bind to anoligosaccharide that contains a chitotriose type structure defined by(GlcNAcb1-4GlcNAcb1-4GlcNAc), but to a lesser extent couldalso bind to one with a chitobiose type structure. The chitobiosestructure is the GlcNAcb1-4GlcNAc entity that is linked to theasparagine residue shown in Fig. 1. Since the chitobiose moietyrather than the chitotriose moiety is present as part of the corestructure of the carbohydrate types delineated in Fig. 1, it wasconsidered that Lzm-S would bind to a chitobiose type structure on
a membrane glycoprotein. In this case, Lzm-S’s depressant activitywould still be evident when all of the terminal sugar residues,except for the chitobiose structure were removed from the carbo-hydrate structure in terms of the protocol described below.
In the exoglycosidase experiments, six groups of trabeculaewere studied. In group 1, no treatment was administered and theeffect of Lzm-S was examined at 10�9M, 10�8M, and 10�7M con-centrations. In group 2, the trabeculae were initially bathed insialidase (60 ml; 5 U/ml) for 2 h to remove any N-acetylneuramicacid (NeuNAc) residues (see Fig. 1), after which the effect of Lzm-S was determined (all kits from Prozyme, San Leandro, Calif.,USA; kit GE 23; Sialidase A). In group 3, the trabeculae werebathed with sialidase and b-galactosidase (60 ml; 3 U/ml) to removepossible NeuNAc and galactose (Gal) residues, after which theeffect of Lzm-S was examined (Kit GE 12; b galactosidase). Ingroup 4, the effect of Lzm-S was examined after the trabeculaewere incubated with sialidase, galactosidase, and b-glu-cosaminidase (60 ml; 45 U/ml) to remove the NeuNAc, Gal, andGlcNAc residues (Kit 31: b glucosaminidase), respectively. Ingroup 5, the trabeculae were bathed with the three previous en-zymes as well as enzymes to remove all of the mannose residues[(Kit GE 60; a(1-6) core mannosidase (60 ml; 1 U/ml); Kit 62; a (1-3,4 mannosidase (60 ml; 11 U/ml); Kit Glyco X-5015: b1-4 man-nosidase) (60 ml; 1 U/200ml)] and most possible fucose attachments[Kit 70; a 1-3,4 fucosidase (60 ml; 0.5 U/ml) and Kit 73; a (1-2)fucosidase (60 ml; 100 m U/ml)]; in group 5, note that since only thechitobiose structure would remain attached to the membrane, noreduction in Lzm-S’s depressant effect was predicted to occur inthis group. In group 6, the exact same protocol was performed asthat described in group 5, except that the b(1-4) mannosidase en-
Fig. 1 Carbohydrates may beclassified into high-mannoseoligosaccharides, hybrid, andcomplex types. Of the complextypes described, bi-antennaryand tri-antennary structures areshown in the figure. The dottedbox shows the tri-mannosylcore. Results suggest that lyso-zyme binds to the Manb1—4GlcNAcb1—4GlcNAc moietyin the core structure (see boldedand underlined residues; seetext for discussion). The threeendo-b-N-acetylglu-cosaminidases, endo-F1, endo-F2, and endo-F3 have specifici-ties for distinct oligosaccharidestructures and cleave the chito-biose structure (see arrows).The results indicate that Lzm-Sbinds to the high mannose/hybrid and tri-antennary carbo-hydrate type structures (seetext). (Man mannose, GlcNAcN-acetylglucosamine, NeuNAcN-acetylneuramic acid, Gal ga-lactose, Asn asparagine,GlcNAcb1-4GlcNAc chitobioseresidue in the core).

132
zyme that cleaved the Manb1-4 linkage was not included in theenzyme mix (see Fig. 1 and Discussion).
In other experiments, endo-b-N-acetylglucosaminidases wereused to define the type of carbohydrate structure to which lysozymebinds. Endo-b-N-acetylglucosaminidases cleave the linkage be-tween the two GlcNAcb1-4GlcNAc residues in the chitobiose core[17] (see bold “F” in Fig. 1). Flavobacterium meningosepticumsecretes three endo-b-N-acetylglucosaminidases, termed endo-F1,endo-F2, and endo-F3 that have specificities for distinct oligosac-charide structures. Endo-F1 cleaves only high-mannose and hybridoligosaccharides. Endo-F2 preferentially processes bi-antennarycomplex oligosaccharides. Endo-F3 is specific for bi-and tri-an-tennary complex oligosaccharides. In respective groups, the ven-tricular trabeculae were treated with either endo-F1(0.3 unit), endo-F2(0.1 unit), and endo-F3(0.1 unit) for 2 h (as per guidelines de-scribed in Native Protein De-glycosylation Kit; Product Code N-Degly, Sigma Canada, Oakville, Ontario). The enzyme was washedoff of the muscle with KH. The effect of Lzm-S on isometriccontraction was compared between the respective endo-F treatedand nontreated groups. Control experiments were also performed inwhich the effect of KH rather than that of Lzm-S was examinedover a similar interval in the three endo-F treated groups.
Lectin experiments
Lectins are nonenzymatic proteins that bind to carbohydrates andcan be classified by their specificity for a particular sugar orpolysaccharide [13]. Lectins do not hydrolyze the bond betweentwo monosaccharide units and therefore binding is reversible. Thus,if hydrolysis of the oligosaccharide were not required for Lzm-S’smyocardial depressant effect, then lectins with a binding specificitysimilar to that of lysozyme would be expected to produce depres-sion in the RVT preparation, while lectins without this specificitywould not produce such an effect. Lycopersicon esculentum lectin(LEL) and Datura stramonium lectin (DSL) have preferential af-finity for the chitotriose structure (i.e., the GlcNAcb1-4GlcNAcb1-4GlcNAc entity) [18, 19] in a manner similar to that described forlysozyme. Triticum vulgaris lectin (TVL) [20] has a predominantaffinity for the related chitobiose structure (i.e., the GlcNAcb1-4GlcNAc entity). On the other hand, Erythrina corallodendronlectin (ECL) [21] and Helix pomatia lectin (HPL) have affinity fortotally unrelated structures that include for the former N-ace-tyllactosamine, N acetyl-galactosamine, lactose, and D-galactose[21] and for the latter N-acetyl galactosaminyl residues [22]. ECLand HPL were included to serve as negative controls in this study.
In the lectin experiments, after baseline isometric contractionwas determined, either the lectin or control (i.e., KH) was placedinto respective organ baths in which the concentrations of thelectins used were similar to those described for Lzm-S. In the caseof DSL, moreover, it was assessed whether pre-treatment withN,N’,N” triacetylchitotriose (10�4M), a known competitive inhibi-tor of Lzm-S’s myocardial depressant effect [4, 5] would preventthe depression caused by this lectin.
To examine whether the carbohydrate portion of the membraneglycoprotein to which lysozyme binds is affectedby the enzymes collagenase A and protease XIV
Because of the potential to use a single myocyte preparation infuture experiments to assess the mechanism by which Lzm-Scauses myocardial depression, there was concern that some of theenzymes used in this preparation, namely collagenase A and pro-tease XIV, would injure the N-glycan to which Lzm-S binds. In theright ventricular trabecular preparation, the same enzymes wereused in a similar manner as that described for the single myocytepreparation after which the effect of Lzm-S on isometric contrac-
tion was determined to see if its depressant activity was stillmaintained. Ventricular trabeculae were treated with collagenase A(25 mg/50 ml buffer, Boehringer-Mannheim) and protease typeXIV (4.8 mg/50 ml buffer, Sigma) for 5 min [14] after which thetrabeculae were washed with KH. The effects of Lzm-S and KH onisometric contraction were compared in respective groups.
Statistics
Statistical analyses included two way repeated measures analysis ofvariance (between-within ANOVA) and Student Newman-Keuls’multiple comparison test. Of the three to four ventricular trabeculaeobtained from a given dog, trabeculae were randomized among thedifferent subset of groups used in a particular experiment. Theresults are expressed as mean (€1SD).
Results
In the reversibility experiments (see Fig. 2), the resultsshow that when Lzm-S was added to the organ prepara-tion in which no intervention was performed, isometriccontraction measured at the 3rd measurement intervaldecreased to � 40% of baseline. On the other hand, whenthe Lzm-S treated group was repeatedly washed withbuffering solution, isometric contraction determined atthe 2nd and 3rd measurement intervals had returned to thebaseline level. In control experiments in which KH ratherthan Lzm-S was added to the preparation, there was littlechange in isometric contraction over the course of theexperiment.
Figure 3 shows the exoglycosidase experiments inwhich the effect of Lzm-S on isometric contraction wasdetermined after terminal sugars of the membrane gly-coprotein were sequentially removed. When no treatmentwas administered, Lzm-S at 10�7M caused a decrease inisometric contraction to approximately 40% of baseline(see group 1). Treatments with sialidase or the combina-tion of a sialidase, galactosidase, and glucosaminidasehad no inhibitory effect on the response to Lzm-S (seegroups 2, 3, 4). On the other hand, in group 5, whentreatment was performed with all enzymes that includedsialidase, galactosidase, glucosaminidase, the three typesof mannosidases (a1-3,4); (a1-6); (b1-4)], and the fu-cosidases (i.e., all terminal sugars except the chitobiosestructure was removed), it was found that the myocardialdepressant effect of Lzm-S was eliminated. In group 6,when the b1-4 mannosidase was omitted from the en-zymes included in group 5, Lzm-S’s myocardial depres-sant effect was restored to that found in groups 1–4 (seeDiscussion and also Fig. 1).
In Fig. 4 (upper panel), the effect of Lzm-S on ven-tricular trabeculae treated with endo-F1, endo-F2, andendo-F3 is shown. Without any treatment, Lzm-S at10�7M decreased isometric contraction to 48% of base-line. When the ventricular trabeculae were incubated withendo-F1, the decrease in isometric contraction at 10�7Mwas significantly less to 70% of baseline. Similar results

133
were obtained with endo-F3 in which isometric contrac-tion decreased to 74% of baseline. On the other hand,treatment with endo-F2 did not significantly attenuate thedepression in isometric contraction as compared to thelysozyme-alone treated group.
In control experiments (Fig. 4 lower panel), in whichKH rather than Lzm-S was added to the preparation, theendo-F enzymes by themselves displayed some depres-sant effect over time that was only slightly greater thantime controls (�10% vs �20%). For the endo-F1 and endo-F3 treatments, no difference in the depressant effect ob-served when the results in the respective Lzm-S and KHgroups shown in Fig. 4 were compared by two-wayANOVA, while there was a significant difference foundfor the end-F2 treatment. In addition, when expressedrelative to the baseline value, the percent decline in iso-metric contraction obtained at a Lzm-S concentration of10�7M was very similar for the Lzm-S vs KH groups forthe endo-F1 (70€11% vs 71€ 3%) and the endo-F3 ex-periments (74€6% vs 78€3%), while for the endo-F2experiments, the depressant effect obtained with Lzm-Swas significantly greater as compared with KH (62€5%vs 80€8%; P<.05). These results indicate that endo-F2 hadlittle attenuating effect on Lzm-S’s myocardial depressantactivity as compared with endo-F1 and endo-F3.
The lectin experiments are shown in F.S1 (see Elec-tronic Supplementary material). DSL (F.S1 left upperpanel) has a primary specificity for the chitotriose struc-ture [19] and caused a decline in isometric contractionsimilar to that found for Lzm-S. Moreover, this decline inisometric tension could be prevented by pre-treatmentwith the inhibitor N,N’,N”-triacetylchitotriose (FS1 leftlower panel) [4, 5]. LEL also has affinity for the chito-
triose structure [18] and similarly produced a markeddecrease in isometric contraction (F.S1 right upper panel).However, TVL which has a specificity to the chitobiosestructure [20], and both ECL [21] and HPL [22] whichhave specificities for entirely different moieties causedmuch less a decline in isometric contraction as comparedwith DSL and LEL (right lower panel).
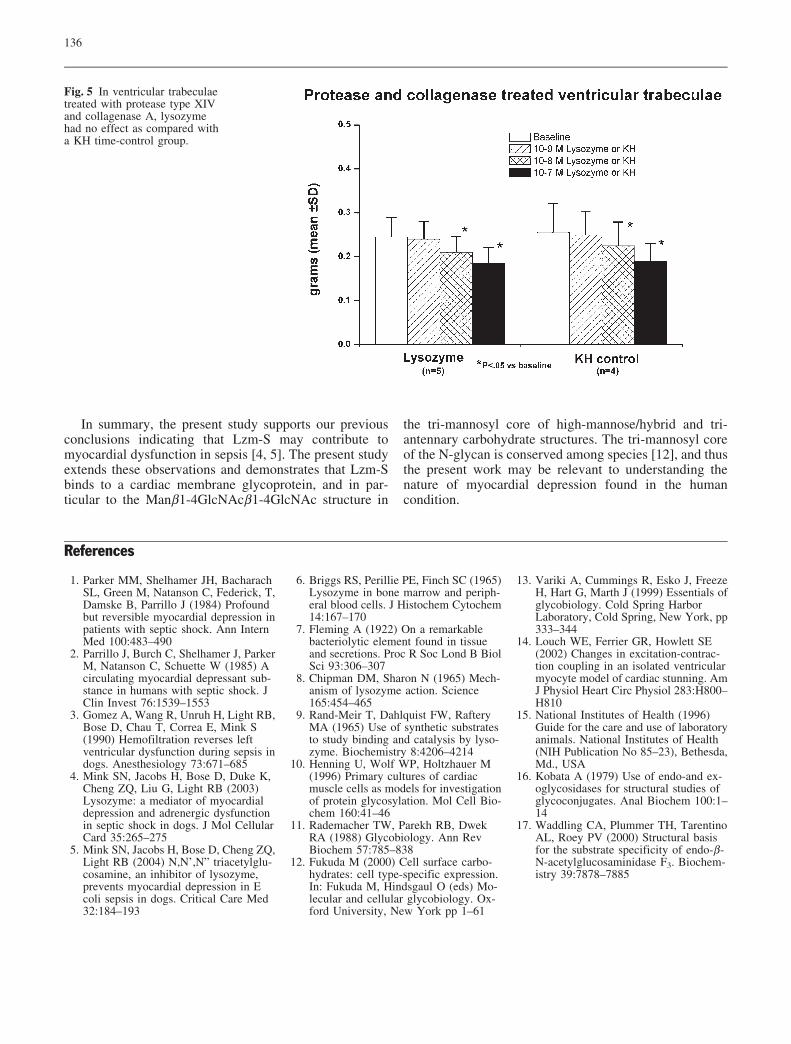
In Fig. 5, the results show that when the ventriculartrabeculae were treated with protease type XIV and col-lagenase A in a manner similar to that described for thesingle myocyte preparation [14], the effect of Lzm-S onisometric contraction was lost, and the results were notdifferent as compared to that observed with KH treatment.
Discussion
In this study, we characterized the N-glycan to whichLzm-S binds to cause myocardial depression in sepsis.Based on the types of carbohydrate structures to whichLzm-S could theoretically bind, our primary hypothesiswas that the binding of Lzm-S would occur to the Glc-NAcb1-4GlcNAc moiety (i.e., the chitobiose structure) inthe region of the tri-mannosyl core (see Fig. 1). However,when all of the sugar residues except for the GlcNAcb1-4GlcNAc structure were removed by exoglycosidasetreatments (see group 5 in Fig. 3), all depressant activityof was Lzm-S was eliminated. Thus, our primary hy-pothesis was incorrect and further examination was re-quired.
Rand-Meir et al. [9] reported that the chitotriosestructure (i.e., the GlcNAcb1-4GlcNAcb1-4GlcNAc en-tity) was a better inhibitor of Lzm-S’s enzymatic activity
Fig. 2 In the reversibility ex-periments, isometric contraction(mean€SD) obtained in theright ventricular trabecularpreparation is plotted on theordinate against measurementinterval for lysozyme treatedand Krebs-Henseleit (KH)treated groups. In the lysozymetreatment alone group, no in-tervention was performed andisometric contraction fell overthe three measurement intervalsperformed 15 min apart. In thelysozyme group in which re-peated washes with bufferingsolution were performed, iso-metric contraction was restoredto baseline levels. In the KHtreated groups, there was littlechange in isometric contractionover the course of the experi-ment.

134
than the two unit chitobiose structure [8, 9]. Furthermore,they showed that Lzm-S cannot only accommodateGlcNAc in its binding, but that the enzyme also bindsglucose and deoxyglucose. In deoxyglucose, the oxygenof the number 2 position of glucose is missing. On theother hand, the N-acetyl moiety in GlcNAc is linked viathe number 2 carbon atom to the rest of the glucosemolecule. We therefore considered that the active site ofLzm-S might be tolerant of changes to the number 2position of the monosaccharide unit that is accommo-dated. Glucose and mannose differ only in the orientationof the hydroxyl group at the number 2 position of themolecule, in one sugar the OH-group is above the plane,in the other it is below the plane of the pyranose ring.Thus, we hypothesized that the Man residue linked by b1-4 linkages to the GlcNAcb1-4GlcNAc in the core struc-ture may be important for the myocardial depressant ef-fect of Lzm-S. When the Manb1-4 GlcNAcb1-4GlcNAcstructure was left intact in group 6 (see Fig. 3), weshowed that Lzm’s depressant activity was again appar-ent, the results supporting the conclusion that Lzm-Sbinds to a variant of the chitotriose structure in the tri-mannosyl core of a membrane glycoprotein (see under-lined structure in Fig. 1).
In other experiments, we used different endo-b-N-acetylglucosaminidases to determine the particular type ofcarbohydrate structure to which Lzm-S may bind [17].Lzm-S’s depressant activity was attenuated by endo-F1,which has specificity for high mannose/hybrid types, andby endo-F3, which has specificity for bi- and tri-antennarycomplex oligosaccharides. On the other hand, there waslittle effect for endo-F2 which preferentially processes bi-antennary complex oliogosaccharides. Stiles et al. [23]examined the carbohydrate subtypes of the mammalian b-adrenergic receptors in hamster lung and rat erythrocytes,and also provided evidence for the existence of both high-mannose and complex type carbohydrate chains on b2-adrenergic receptors. Our results show that Lzm-S bindsto similar types of carbohydrate structures in sepsis.
The washing experiments (Fig. 2) and the lectin ex-periments (F.S1) indicate that the myocardial depressioncaused by Lzm-S is reversible, since lectins do not havethe capability of hydrolyzing the bond between twomonosaccharide units [12]. Lectins such as LEL and DSLthat have a specificity to the chitotriose structure caused adecrease in isometric contraction in a manner similar toLzm-S, while the other lectins used did not cause myo-cardial depression. The lectin study adds to the results
Fig. 3 In the exoglycosidase experiments, isometric contraction(mean€SD) measured in the right ventricular trabecular preparationis plotted on the ordinate. In group 1, no enzymes were added, andlysozyme was instilled at 10�9M, 10�8M, and 10�7M at 5 min in-tervals. In group 2, a sialidase was added to remove any N-acetylneuramic acids present and no difference in isometric tensionwas noted as compared to group 1; in group 3, both sialidase andgalactosidase were added to remove any N-acetylneuramic andgalactose residues and no change was noted as compared with
group 1; in group 4, the enzymes administered consisted of asialidase, galactosidase, and a glucosaminidase, and no significanteffect was observed as compared to the preceding groups; in group5, in addition to the latter enzymes, a and b(1–4) mannosidases andfucosidases were added, and lysozyme’s depressant activity wascompletely eliminated; in group 6, the b1-4 mannosidase wasomitted from the enzyme mixture used in group 5, and the de-pressant activity increased to that found in groups 1-4.

135
obtained in the exoglycosidase study, since both studiesindicate that in order for myocardial depression to occur,Lzm-S needs to bind to an oligosaccharide that contains achitotriose type structure. The exoglycosidase study fur-ther points out that binding to a chitotriose variant may besufficient in which case a Man residue linked by b1-4linkage to the GlcNAcb1-4GlcNAc moiety may substitutefor the GlcNAc residue found in chitotriose (see under-lined structure in Fig. 1).
The present study also suggests that the glycoprotein towhich Lzm-S binds may be disrupted by some of the
agents used in preparing the single myocyte preparation[14]. Thus, this fact must be considered when data fromthe intact preparation and isolated myocytes are com-pared. Further work will have to be performed to deter-mine the precise receptor to which Lzm-S binds, to dis-tinguish whether this receptor is found on the myocardialcell per se or the endothelial cells that line the ventriculartrabeculae, as well as to define the biochemical signalingmechanism responsible for the myocardial depressionobserved.
Fig. 4 The effect of lysozyme(upper panel) on isometriccontraction (mean€SD) isshown for the respective groupsof ventricular trabeculae bathedwith the three endo-b-N-acetylglucosaminidases. Endo-F1 and endo-F3 attenuated ly-sozyme’s depressant effect tothe greatest extent, while endo-F2 had no significant effect. Inthe lower panel, when KrebsHenseleit solution (KH) ratherthan lysozyme was added to thepreparation, there was a modestdepressant of the enzymes bythemselves that was slightlygreater than that found in thetime-control group. For theendo-F1 and endo-F3 treat-ments, no difference in the de-pressant effect observed whenrespective Lzm-S and KHgroups were compared by two-way ANOVA. On the otherhand, for the endo-F2 experi-ments, the depressant effectobtained with KH was muchless than that found when Lzm-S was used.
136
References
1. Parker MM, Shelhamer JH, BacharachSL, Green M, Natanson C, Federick, T,Damske B, Parrillo J (1984) Profoundbut reversible myocardial depression inpatients with septic shock. Ann InternMed 100:483–490
2. Parrillo J, Burch C, Shelhamer J, ParkerM, Natanson C, Schuette W (1985) Acirculating myocardial depressant sub-stance in humans with septic shock. JClin Invest 76:1539–1553
3. Gomez A, Wang R, Unruh H, Light RB,Bose D, Chau T, Correa E, Mink S(1990) Hemofiltration reverses leftventricular dysfunction during sepsis indogs. Anesthesiology 73:671–685
4. Mink SN, Jacobs H, Bose D, Duke K,Cheng ZQ, Liu G, Light RB (2003)Lysozyme: a mediator of myocardialdepression and adrenergic dysfunctionin septic shock in dogs. J Mol CellularCard 35:265–275
5. Mink SN, Jacobs H, Bose D, Cheng ZQ,Light RB (2004) N,N’,N” triacetylglu-cosamine, an inhibitor of lysozyme,prevents myocardial depression in Ecoli sepsis in dogs. Critical Care Med32:184–193
6. Briggs RS, Perillie PE, Finch SC (1965)Lysozyme in bone marrow and periph-eral blood cells. J Histochem Cytochem14:167–170
7. Fleming A (1922) On a remarkablebacteriolytic element found in tissueand secretions. Proc R Soc Lond B BiolSci 93:306–307
8. Chipman DM, Sharon N (1965) Mech-anism of lysozyme action. Science165:454–465
9. Rand-Meir T, Dahlquist FW, RafteryMA (1965) Use of synthetic substratesto study binding and catalysis by lyso-zyme. Biochemistry 8:4206–4214
10. Henning U, Wolf WP, Holtzhauer M(1996) Primary cultures of cardiacmuscle cells as models for investigationof protein glycosylation. Mol Cell Bio-chem 160:41–46
11. Rademacher TW, Parekh RB, DwekRA (1988) Glycobiology. Ann RevBiochem 57:785–838
12. Fukuda M (2000) Cell surface carbo-hydrates: cell type-specific expression.In: Fukuda M, Hindsgaul O (eds) Mo-lecular and cellular glycobiology. Ox-ford University, New York pp 1–61
13. Variki A, Cummings R, Esko J, FreezeH, Hart G, Marth J (1999) Essentials ofglycobiology. Cold Spring HarborLaboratory, Cold Spring, New York, pp333–344
14. Louch WE, Ferrier GR, Howlett SE(2002) Changes in excitation-contrac-tion coupling in an isolated ventricularmyocyte model of cardiac stunning. AmJ Physiol Heart Circ Physiol 283:H800–H810
15. National Institutes of Health (1996)Guide for the care and use of laboratoryanimals. National Institutes of Health(NIH Publication No 85–23), Bethesda,Md., USA
16. Kobata A (1979) Use of endo-and ex-oglycosidases for structural studies ofglycoconjugates. Anal Biochem 100:1–14
17. Waddling CA, Plummer TH, TarentinoAL, Roey PV (2000) Structural basisfor the substrate specificity of endo-b-N-acetylglucosaminidase F3. Biochem-istry 39:7878–7885
In summary, the present study supports our previousconclusions indicating that Lzm-S may contribute tomyocardial dysfunction in sepsis [4, 5]. The present studyextends these observations and demonstrates that Lzm-Sbinds to a cardiac membrane glycoprotein, and in par-ticular to the Manb1-4GlcNAcb1-4GlcNAc structure in
the tri-mannosyl core of high-mannose/hybrid and tri-antennary carbohydrate structures. The tri-mannosyl coreof the N-glycan is conserved among species [12], and thusthe present work may be relevant to understanding thenature of myocardial depression found in the humancondition.
Fig. 5 In ventricular trabeculaetreated with protease type XIVand collagenase A, lysozymehad no effect as compared witha KH time-control group.

137
18. Nachbar MS, Oppenheim JD, ThomasJO (1980) Lectins in the US Diet. Iso-lation and characterization of a lectinfrom the tomato (Lycopersicon escu-lentum). J Biol Chem 255:2056–2061
19. Crowley JI, Goldstein IJ, Arnarp J,Lonngren J (1984) Carbohydrate bind-ing studies on the lectin from Daturastramonium seeds. Arch Biochem Bio-phys 231:524–533
20. Nagata Y, Burger MM (1974) Wheatgerm agglutinin. Molecular character-istics and specificity for sugar binding.J Biol Chem 249:3116–3122
21. Gilboa-Garber N, Mizrahi L (1981) Anew mitogenic D-galactosephilic lectinisolated from seeds of the coral-treeErythrina corallodendron. Comparisonswith Glycine max (soybean) and Pseu-domonas aeruginosa lectins. Can JBiochem 59:315–320
22. Hammarstrom S, Westoo A, Bjork I(1972) Subunit structure of Helix po-matia A hemagglutinin. Scand J Im-munol 1:295–309
23. Stiles GL, Benovic JL, Caron MG,Lefkowitz RJ (1984) Mammalian b-adrenergic receptors. Distinct glyco-protein populations containing highmannose or complex type carbohydratechains. J Biol Chem 259:8566–8663