Embed Size (px)
Citation preview

GENOMICS, TRANSCRIPTOMICS, PROTEOMICS
Characterization of Lactobacillus salivarius CECT 5713,a strain isolated from human milk: from genotypeto phenotype
Susana Langa & Antonio Maldonado-Barragán &
Susana Delgado & Rebeca Martín & Virginia Martín &
Esther Jiménez & José L. Ruíz-Barba & Baltasar Mayo &
Ruth I. Connor & Juan Evaristo Suárez &
Juan M. Rodríguez
Received: 20 January 2012 /Revised: 8 March 2012 /Accepted: 15 March 2012 /Published online: 19 April 2012# Springer-Verlag 2012
Abstract Lactobacillus salivariusCECT 5713, isolated fromhuman milk, has immunomodulatory, anti-inflammatory andantiinfectious properties, as revealed by several in vitro and invivo assays, which suggests a strong potential as a probioticstrain. In this work, the relationships between several geneticfeatures of L. salivarius CECT 5713 and the correspondingphenotypes were evaluated. Although it contains a plasmid-
encoded bacteriocin cluster, no bacteriocin biosynthesis wasobserved, possibly due to a 4-bp deletion at the beginning ofthe histidine kinase determinant abpK. The genome of L.salivarius CECT 5713 harbours two apparently completeprophages of 39.6 and 48 kbp. Upon induction, the 48-kbpprophage became liberated from the bacterial genome, but noDNA replication took place, which resulted in lysis of thecultures but not in phage progeny generation. The strain wassensitive to most antibiotics tested and no transmissible genespotentially involved in antibiotic resistance were detected.Finally, the genome of L. salivarius CECT 5713 containedfour ORFs potentially involved in human molecular mime-tism. Among them, protein 1230 was considered of particularrelevance because of its similarity with dendritic cell-relatedproteins. Subsequently, in vitro assays revealed the ability ofL. salivarius CECT 5713 to stimulate the maturation of im-mature dendritic cells and to inhibit the in vitro infectivity ofHIV-1.
Keywords Lactobacillus salivarius . Probiotic . Genome .
Phage . Antibiotic resistance . Bacteriocin . Dendritic cell .
HIV
Introduction
Lactobacillus salivarius is part of the indigenous microbiotaof the gastrointestinal tract, oral cavity and milk of humans(Casey et al. 2004;Martín et al. 2006; Rogosa et al. 1953). Theproperties associated with some strains, and particularly to L.salivarius UCC118 (Neville and O’Toole 2010), have lead toan increasing interest in the probiotic utility of this species.
S. Langa :V. Martín : E. Jiménez : J. M. Rodríguez (*)Departamento de Nutrición, Bromatología y Tecnología de losAlimentos, Universidad Complutense de Madrid,28040, Madrid, Spaine-mail: [email protected]
S. LangaDepartamento de Tecnología de Alimentos, Instituto Nacional deInvestigación y Tecnología Agraria y Alimentaria,Carretera de la Coruña Km 7.5, 28040, Madrid, Spain
A. Maldonado-Barragán : J. L. Ruíz-BarbaDepartamento de Biotecnología de Alimentos,Instituto de la Grasa-CSIC,41012, Sevilla, Spain
S. Delgado : B. Mayo : J. E. SuárezInstituto de Productos Lácteos de Asturias (IPLA-CSIC),Carretera de Infiesto s/n,33300, Villaviciosa, Spain
R. Martín : J. E. SuárezArea de Microbiología e Instituto de Biotecnología,Universidad de Oviedo,33006, Oviedo, Spain
R. I. ConnorDepartment of Microbiology and Immunology,Dartmouth Medical School,Lebanon 03756 NH, USA
Appl Microbiol Biotechnol (2012) 94:1279–1287DOI 10.1007/s00253-012-4032-1

L. salivarius CECT 5713 was isolated from human milkand infant feces of a healthy mother–child pair (Martín et al.2006) and has been shown to be a good probiotic candidatedue to its repertoire of desirable properties. This includes ahigh survival rate when exposed to gastrointestinal tractconditions, bile salt hydrolase activity, adherence to mucusand intestinal cells, ability to stimulate the expression ofmucin-encoding genes, and in vitro and in vivo antimicro-bial, anti-inflammatory, and immunomodulatory properties(Díaz-Ropero et al. 2006, Martín et al. 2006; Olivares et al.2006; Pérez-Cano et al., 2010). Furthermore, it has beenshown to be well tolerated and safe in animal models (Lara-Villoslada et al. 2007) and in human clinical assays (Arroyoet al. 2010; Jiménez et al. 2008; Maldonado et al. 2010).
Administration of probiotic bacteria benefits the hostthrough a wide array of mechanisms that are increasinglyrecognized as being species- and strain-specific. Therefore,a better knowledge of the genetic basis for common andstrain-specific traits in potentially probiotic strains is re-quired. A recent comparative genomics study that included33 L. salivarius strains isolated from humans, animals orfood revealed that this species is genomically diverse (Raftiset al. 2011). The genome of L. salivarius CECT 5713 wassequenced recently (Jiménez et al. 2010). It consists of acircular chromosome of 1,828,169 bp, two plasmids (pHN1,44,581 bp; pHN2, 20,426 bp), and a megaplasmid (pHN3,242,962 bp) and includes 52 genes that are not present in thegenome of L. salivarius UCC118 (Claesson et al. 2006).The objective of this work was to evaluate the relationshipbetween the genetic background of L. salivarius CECT5713, and certain phenotypic traits, including bacteriocinproduction, presence of prophages, occurrence of antibioticresistance genes, and potential molecular mimetism in hu-man hosts.
Materials and methods
Bacterial strain and growth conditions
L. salivarius CECT 5713was grown inMRSmedium (Oxoid,Basingstoke, UK) containing 0.05 % (w/v) L-cysteine–HCl(AppliChem; MRS-Cys) at 30 °C under anaerobiosis using aDG250 Anaerobic Workstation (Don Whitley Scientific Ltd.,Shipley, West Yorkshire, UK) with a gas mixture consisting of10 % H2, 10 % CO2, and 80 % N2.
Bacteriocin production testing
To check for antimicrobial activities on solid medium, over-night cultures of L. salivarius CECT 5713 were spotted ontoMRS-Cys agar plates, incubated at 30 °C for 18 h in an-aerobiosis, and overlaid with 4.5 ml soft agar inoculated
with ~105 CFU/ml of the selected indicator strain. Plateswere further incubated at 30 °C for 24 h and examined forclear halos of inhibition. Bacteriocin activity in neutralized,cell-free supernatants was assayed by using the agar dropdiffusion test as previously described (Maldonado et al.2003). The following bacterial strains were used as indica-tors: Lactobacillus acidophilus NCDO 1748, Lactobacillusbrevis LB9, Lactobacillus casei ATCC 334, Lactobacilluscoryniformis Q8, Lactobacillus fermentum ATCC 9338,Lactobacillus hilgardii LB76, Lactobacillus jensenii Ov2967, Lactobacillus paraplantarum CNRZ 1885, Lactoba-cillus pentosus 128/2 and 55–1, Lactobacillus plantarumNC8 and ATCC 8014, Lactobacillus reuteri DSM 20016,Lactobacillus salivarius NCFB 2747, Lactobacillus sakeiNCFB 2714, Lactococcus lactis MG 1363, L. lactis CNRZ117, L. lactis IPLA 838, L. lactis IL 1403, Streptococcusthermophilus ST 112, Leuconostoc citreum IPLA 616, L.mesenteroides INRA 33, Pediococcus damnosus NCDO1832, Pediococcus parvulus P339, Pediococcus pentosa-ceus FBB63, P. pentosaceus PC1, Enterococcus faeciumLP6T1a-20, Enterococcus faecalis EFI, Listeria innocuaBL86/26, and Staphylococcus carnosus MC1.
Analysis of the histidin kinase gene abpK
The primers AbpIP-F (5′-GCAACCAAAAAGGGTGGC-3′)and AbpHK-R (5′-TGCTCATAACCATAAGGAAAGT-3′)were designed to span the autoinducing peptide AbpIP(abpIP) and its histidine protein kinase AbpK (abpK) deter-minants (GenBank accession number, CP002037; Fig. 1).DNA was amplified in 50 μl reaction mixtures containing2.5 mM MgCl2, 1× reaction buffer, 200 μM of each deoxy-nucleotide triphosphate, 1 μM of each primer, and 1.25 U TaqDNA polymerase (Ecotaq; Ecogen, Barcelona, Spain). Am-plification was as follows: denaturation at 94 °C for 4 min, 30cycles of denaturation at 94 °C for 30 s, annealing at 58 °C for30 s, polymerization at 72 °C for 90 s, and a final polymeri-zation step at 72 °C for 5 min. The amplified fragment waspurified using the Nucleospin® Extract II kit (Macherey-Nagel, Düren, Germany), and sequenced using primersAbpIP-F and AbpHK-R (Newbiotechnics, Seville, Spain).
Prophage induction
Exponential cultures of L. salivarius CECT 5713 (O.D.6000.4) were treated with 0.25, 0.5 (minimal inhibitory concen-tration; MIC), and 1 μg/ml mitomycin C (final concentra-tion) and incubation was continued for up to 5 h. Aliquots ofthe supernatants were placed on lawns of presumably sus-ceptible L. salivarius strains growing in soft MRS (0.75 %agar) supplemented with 1 % haemoglobin, 10 mM CaCl2and 10 mM MgSO4, placed on top of plates with the samemedium (1.5 % agar). After incubation for 24 h, the
1280 Appl Microbiol Biotechnol (2012) 94:1279–1287

generation of plaques of lysis was recorded. Real-time PCRwas performed with primers internal to the putative majorhead and tail protein genes and others flanking the putativeattP sites of the prophages, under the conditions describedpreviously (Soberón et al. 2007), using the PCR Q SYBRGreen Supermix (Bio-Rad). Searching for virions producedby induced cultures was performed as described previously(Martín et al. 2009). In short, cleared supernatants weretreated with polyethylene glycol and the resulting pelletwas successively subjected to discontinuous and continuousCsCl gradient centrifugations. Aliquots of the resultingbands were negatively stained with 2 % (w/v) uranyl acetateand observed by electron microscopy.
Antibiotic resistance/susceptibility
Initially, a search for antibiotic resistance genes was per-formed in the genome of L. salivarius CECT 5713. BLAST,BLASTX, and BLASTN programs (http://blast.ncbi.nlm.nih.gov/Blast.cgi) were used to compare nucleotide and de-duced amino acid sequences with those stored in the GenBankdatabase. The MIC of the 16 antibiotics included in this study(Table 1) was determined on cultures grown overnight inlactic acid bacteria susceptibility test medium (LSM; Klareet al. 2005) and diluted to obtain a density corresponding toMcFarland standard 1 (equivalent to ~3×108 CFU/ml). Thesuspension was further adjusted to 3×105 CFU/ml with LSM,and 100 μl were inoculated to wells of microtiter VetMICplates for lactic acid bacteria (National Veterinary Institute ofSweden, Uppsala, Sweden). The plates were incubated at37 °C for 48 h and the MIC was defined as the lowestconcentration at which no growth was observed.
Genes potentially involved in host mimetism
The genome of L. salivarius CECT 5713 was searched forthe presence of ORFs encoding putative proteins potentiallyinvolved in human molecular mimetism. For this purpose,the WIP (Words In Proteins) algorithm developed by Era7
Information Technologies (Granada, Spain) was used. Thisalgorithm selects proteins that, despite a distant evolutionaryorigin, share stretches above the expected level that is cal-culated doing, for each protein pair, a million of compar-isons between different randomizations of their sequences.
Stimulation of immature dendritic cells
Mouse immature dendritic cells (DC) were isolated from thespleen of female C57BL/6 mice (6–10-week age) and char-acterized as described previously (Lu et al. 1995). Forpropagation of isolated DC, they were routinely cultured at
1000
L. salivarius CECT 5713 (pHN3)
abpIM
salB abp118a
abpK2
DpbaTpbaRpba
blp1
abpIP
L. salivarius UCC118 (pMP118)
abpK
abpK1
blp3
abp118b
blp2
LSL_1924
orf1 orf4orf3 orf5
orf1
1 2
Fig. 1 Genetic maps of the bacteriocin clusters harboured in themegaplasmids pHN3 and pUCC118 of L. salivarius CECT 5713 andL. salivarius UCC 118, respectively. Dark arrows indicate genes that
share a high degree of homology or are virtually identical. The cleararrows indicate genes of L. salivarius CECT 5713 that differ fromthose of L. salivarius UCC 118
Table 1 Minimal inhibitory concentration (MIC) values of 16 anti-biotics to Lactobacillus salivarius CECT 5713
Antibiotic MIC (μg/ml)
CECT 5713 Microbiological breakpointsa
Penicillin 0.25 –
Ampicillin 1 4
Ciprofloxacin 2 –
Gentamicin 2 16
Kanamycin 128 64
Streptomycin 32 64
Neomycin 4 –
Tetracycline 2 8
Erythromycin 0.12 1
Clindamycin 0.25 1
Chloramphenicol 2 4
Vancomycin >128 –
Virginiamycin 0.5 –
Linezolid 1 –
Trimethoprim 0.25 –
Rifampicin 1 –
aMicrobiological breakpoints are defined as the MIC values that clearlydeviate from those displayed by the normal susceptible populations(EFSA 2008)
Appl Microbiol Biotechnol (2012) 94:1279–1287 1281

37 °C in a humidified 5 % CO2 atmosphere in IMDMmedium with 10 % heat-inactivated FBS, 2 mM L-gluta-mine, 100 IU/ml penicillin G, 100 μg/ml streptomycin, and20 pM β-mercaptoethanol (complete IMDM; all the com-ponents were from Sigma) and supplemented with 0.4 ng/mlmouse rGM-CSF (R&D Systems). The culture medium waschanged every four days and granulocytes and mature DCswere removed by gentle washes, after which the cultureswere replenished with new medium containing fresh rGM-CSF (Lu et al. 1995).
Two markers were selected to study the ability of L.salivarius CECT 5713 to activate DC: the class II majorhistocompatibility complex (MHC II) which participates inantigen presentation to naïve T cells and B7.2 (CD86), themajor functional ligand for CD28 and CTLA-4, two criticalT-cell signalling molecules (Freeman et al. 1993; Linsleyand Ledbetter 1993; Caux et al. 1994). The presence ofthese two cell markers in the surface of DCs is significantlyincreased following DC activation (Hart 1997). The mono-clonal anti-mouse antibodies IA/Ed (2G9) and CD86/B7.2(GL1), which specifically recognise mouse MHC II andB7.2, respectively, were purchased from PharMingen (SanDiego, USA). Phycoerythrin and streptavidin were obtainedfrom Sigma.
MRS-Cys overnight cultures of L. salivarius CECT 5713were recovered by centrifugation at 6,000×g for 5 min, andwashed twice with PBS. Then, 2×107 cfu were distributedin 100 μl aliquots of Isocove’s modified Dulbecco’s medi-um (IMDM) devoid of antibiotics and added to 10 ml freshDC cultures containing 2×106 cells. The cocultures wereincubated for 90 min at 37 °C and non-inoculated DCcultures were included as negative controls. After the incu-bation period, cells were washed with PBS and 2 mM EDTAand maintained for 18 h at 37 °C in complete IMDMsupplemented with gentamicin (250 μg/ml) and tetracycline(10 μg/ml) to kill the remaining bacteria. Then, cells werewashed twice with PBS and stained with the anti-MHC classII and anti-B7.2 antibodies in order to detect both DC andthe potential activation of the surface markers. Staining wasperformed according to standard immunofluorescence tech-niques while labelling of the antibodies with phycoerythrinwas carried out following the manufacturer instructions(Sigma). Finally, flow cytometry analysis was performedwith a FACS scan (Becton Dickinson, San Jose, CA) andthe resulting data were analyzed with the WinMDI 2.8software. A total of 10,000 cells were analyzed through aviable cell gate determined by forward and right angle lightscatter parameters to exclude subcellular particles.
Activity against HIV infectivity
HIV-1 isolates used in this study included R5-tropicHIV-1BaL and two primary isolates, X4-tropic HIV-1HC4
and R5/X4 HIV-1C7/86. All HIV-1 isolates were propa-gated in PHA-activated peripheral blood mononuclearcells (PBMC) and titered on TZM-bl cells [NIH AIDSResearch and Reference Reagent Program (Germantown,MD), contributed by Dr. John C. Kappes, Dr. XiaoyunWu and Tranzyme Inc.] prior to be used in infectivityexperiments.
Subsequently, TZM-bl cells were used to assess theeffect of heat-killed bacteria and cell-free culture super-natants (CF-CS) on HIV-1 infectivity, as described previ-ously (Martín et al. 2010b). TZM-bl cells are derived fromHeLa cells and engineered to stably express the HIV-1receptor (CD4) and co-receptors (CCR5 and CXCR4) re-quired for infection along with reporter genes for luciferaseand β-galactosidase under the control of the HIV-1 pro-moter. Controls included TZM-bl cells infected with therespective HIV-1 isolates in the absence of added bacteriaor supernatants.
Results
Analysis of the L. salivarius CECT 5713 bacteriocin cluster
The pHN3 megaplasmid harbours a DNA stretch that issimilar to the bacteriocin cluster located in the pMP118plasmid of L. salivarius UCC118 (Flynn et al. 2002)(Fig. 1). Namely, genes putatively encoding five bacteriocinprecursors (salb, blp1, blp2, blp3, and abp118α andabp118β, which code for salivaricin Abp118, a two compo-nent bacteriocin), three determinants involved in bacteriocinexport (orf1, abpT, and abpD), one in immunity (abpIM)and those presumptively involved in quorum sensing reg-ulation of production (abpIL, abpK, and abpR). Theircommon origin is evidenced by the gene order in thecluster, and by the high homology (identity between90 % and 100 %) both at the DNA and protein levels.However, L. salivarius CECT 5713 did not show anyantimicrobial activity when challenged against 26 strainsof Gram-positive bacteria, including a variety of lactobacilli,lactococci, leuconostocs, pediococci, listeria, enterococci,streptococci, and staphylococci.
To understand the reason of this lack of bacteriocinogenicactivity, the bacteriocin cluster of L. salivarius CECT 5713was analyzed, and compared to the corresponding DNAsequence of L. salivarius UCC118. The only significantdifference was related to the abpK gene, which encodesthe histidine kinase determinant (429 amino acids) involvedin the quorum sensing system required for the regulation ofbacteriocin synthesis in strain UCC118. However, there is a4 nucleotides deletion at the beginning of the abpK gene ofCECT 5713, which introduces a frameshift mutation result-ing in a premature stop codon.
1282 Appl Microbiol Biotechnol (2012) 94:1279–1287
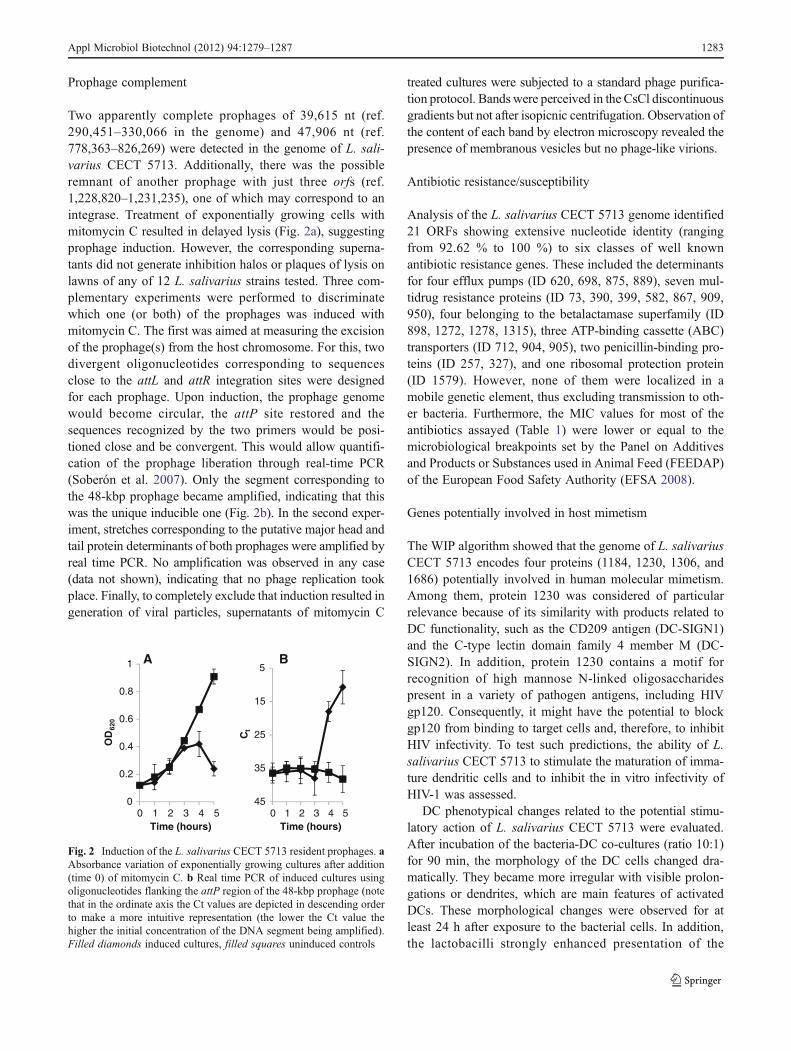
Prophage complement
Two apparently complete prophages of 39,615 nt (ref.290,451–330,066 in the genome) and 47,906 nt (ref.778,363–826,269) were detected in the genome of L. sali-varius CECT 5713. Additionally, there was the possibleremnant of another prophage with just three orfs (ref.1,228,820–1,231,235), one of which may correspond to anintegrase. Treatment of exponentially growing cells withmitomycin C resulted in delayed lysis (Fig. 2a), suggestingprophage induction. However, the corresponding superna-tants did not generate inhibition halos or plaques of lysis onlawns of any of 12 L. salivarius strains tested. Three com-plementary experiments were performed to discriminatewhich one (or both) of the prophages was induced withmitomycin C. The first was aimed at measuring the excisionof the prophage(s) from the host chromosome. For this, twodivergent oligonucleotides corresponding to sequencesclose to the attL and attR integration sites were designedfor each prophage. Upon induction, the prophage genomewould become circular, the attP site restored and thesequences recognized by the two primers would be posi-tioned close and be convergent. This would allow quantifi-cation of the prophage liberation through real-time PCR(Soberón et al. 2007). Only the segment corresponding tothe 48-kbp prophage became amplified, indicating that thiswas the unique inducible one (Fig. 2b). In the second exper-iment, stretches corresponding to the putative major head andtail protein determinants of both prophages were amplified byreal time PCR. No amplification was observed in any case(data not shown), indicating that no phage replication tookplace. Finally, to completely exclude that induction resulted ingeneration of viral particles, supernatants of mitomycin C
treated cultures were subjected to a standard phage purifica-tion protocol. Bandswere perceived in the CsCl discontinuousgradients but not after isopicnic centrifugation. Observation ofthe content of each band by electron microscopy revealed thepresence of membranous vesicles but no phage-like virions.
Antibiotic resistance/susceptibility
Analysis of the L. salivarius CECT 5713 genome identified21 ORFs showing extensive nucleotide identity (rangingfrom 92.62 % to 100 %) to six classes of well knownantibiotic resistance genes. These included the determinantsfor four efflux pumps (ID 620, 698, 875, 889), seven mul-tidrug resistance proteins (ID 73, 390, 399, 582, 867, 909,950), four belonging to the betalactamase superfamily (ID898, 1272, 1278, 1315), three ATP-binding cassette (ABC)transporters (ID 712, 904, 905), two penicillin-binding pro-teins (ID 257, 327), and one ribosomal protection protein(ID 1579). However, none of them were localized in amobile genetic element, thus excluding transmission to oth-er bacteria. Furthermore, the MIC values for most of theantibiotics assayed (Table 1) were lower or equal to themicrobiological breakpoints set by the Panel on Additivesand Products or Substances used in Animal Feed (FEEDAP)of the European Food Safety Authority (EFSA 2008).
Genes potentially involved in host mimetism
The WIP algorithm showed that the genome of L. salivariusCECT 5713 encodes four proteins (1184, 1230, 1306, and1686) potentially involved in human molecular mimetism.Among them, protein 1230 was considered of particularrelevance because of its similarity with products related toDC functionality, such as the CD209 antigen (DC-SIGN1)and the C-type lectin domain family 4 member M (DC-SIGN2). In addition, protein 1230 contains a motif forrecognition of high mannose N-linked oligosaccharidespresent in a variety of pathogen antigens, including HIVgp120. Consequently, it might have the potential to blockgp120 from binding to target cells and, therefore, to inhibitHIV infectivity. To test such predictions, the ability of L.salivarius CECT 5713 to stimulate the maturation of imma-ture dendritic cells and to inhibit the in vitro infectivity ofHIV-1 was assessed.
DC phenotypical changes related to the potential stimu-latory action of L. salivarius CECT 5713 were evaluated.After incubation of the bacteria-DC co-cultures (ratio 10:1)for 90 min, the morphology of the DC cells changed dra-matically. They became more irregular with visible prolon-gations or dendrites, which are main features of activatedDCs. These morphological changes were observed for atleast 24 h after exposure to the bacterial cells. In addition,the lactobacilli strongly enhanced presentation of the
0
0.2
0.4
0.6
0.8
1
0 1 2 3 4 5
OD
620
Time (hours)
5
15
25
35
450 1 2 3 4 5
Ct
Time (hours)
BA
Fig. 2 Induction of the L. salivarius CECT 5713 resident prophages. aAbsorbance variation of exponentially growing cultures after addition(time 0) of mitomycin C. b Real time PCR of induced cultures usingoligonucleotides flanking the attP region of the 48-kbp prophage (notethat in the ordinate axis the Ct values are depicted in descending orderto make a more intuitive representation (the lower the Ct value thehigher the initial concentration of the DNA segment being amplified).Filled diamonds induced cultures, filled squares uninduced controls
Appl Microbiol Biotechnol (2012) 94:1279–1287 1283
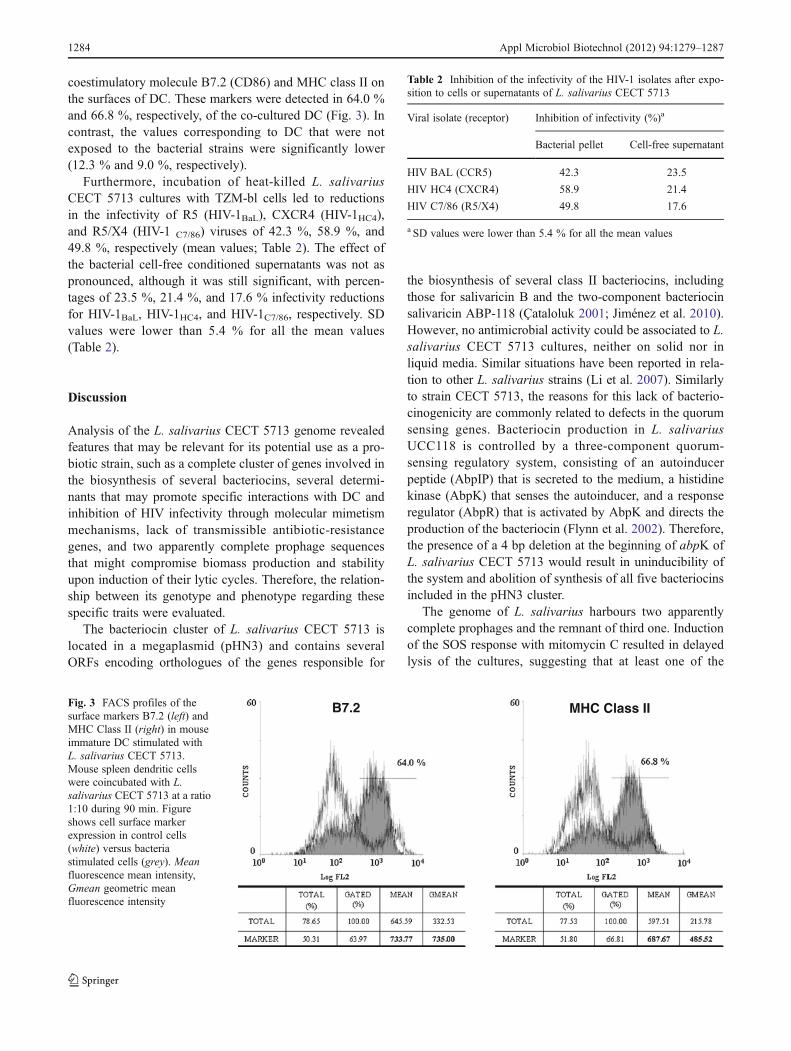
coestimulatory molecule B7.2 (CD86) and MHC class II onthe surfaces of DC. These markers were detected in 64.0 %and 66.8 %, respectively, of the co-cultured DC (Fig. 3). Incontrast, the values corresponding to DC that were notexposed to the bacterial strains were significantly lower(12.3 % and 9.0 %, respectively).
Furthermore, incubation of heat-killed L. salivariusCECT 5713 cultures with TZM-bl cells led to reductionsin the infectivity of R5 (HIV-1BaL), CXCR4 (HIV-1HC4),and R5/X4 (HIV-1 C7/86) viruses of 42.3 %, 58.9 %, and49.8 %, respectively (mean values; Table 2). The effect ofthe bacterial cell-free conditioned supernatants was not aspronounced, although it was still significant, with percen-tages of 23.5 %, 21.4 %, and 17.6 % infectivity reductionsfor HIV-1BaL, HIV-1HC4, and HIV-1C7/86, respectively. SDvalues were lower than 5.4 % for all the mean values(Table 2).
Discussion
Analysis of the L. salivarius CECT 5713 genome revealedfeatures that may be relevant for its potential use as a pro-biotic strain, such as a complete cluster of genes involved inthe biosynthesis of several bacteriocins, several determi-nants that may promote specific interactions with DC andinhibition of HIV infectivity through molecular mimetismmechanisms, lack of transmissible antibiotic-resistancegenes, and two apparently complete prophage sequencesthat might compromise biomass production and stabilityupon induction of their lytic cycles. Therefore, the relation-ship between its genotype and phenotype regarding thesespecific traits were evaluated.
The bacteriocin cluster of L. salivarius CECT 5713 islocated in a megaplasmid (pHN3) and contains severalORFs encoding orthologues of the genes responsible for
the biosynthesis of several class II bacteriocins, includingthose for salivaricin B and the two-component bacteriocinsalivaricin ABP-118 (Çataloluk 2001; Jiménez et al. 2010).However, no antimicrobial activity could be associated to L.salivarius CECT 5713 cultures, neither on solid nor inliquid media. Similar situations have been reported in rela-tion to other L. salivarius strains (Li et al. 2007). Similarlyto strain CECT 5713, the reasons for this lack of bacterio-cinogenicity are commonly related to defects in the quorumsensing genes. Bacteriocin production in L. salivariusUCC118 is controlled by a three-component quorum-sensing regulatory system, consisting of an autoinducerpeptide (AbpIP) that is secreted to the medium, a histidinekinase (AbpK) that senses the autoinducer, and a responseregulator (AbpR) that is activated by AbpK and directs theproduction of the bacteriocin (Flynn et al. 2002). Therefore,the presence of a 4 bp deletion at the beginning of abpK ofL. salivarius CECT 5713 would result in uninducibility ofthe system and abolition of synthesis of all five bacteriocinsincluded in the pHN3 cluster.
The genome of L. salivarius harbours two apparentlycomplete prophages and the remnant of third one. Inductionof the SOS response with mitomycin C resulted in delayedlysis of the cultures, suggesting that at least one of the
B7.2 MHC Class IIFig. 3 FACS profiles of thesurface markers B7.2 (left) andMHC Class II (right) in mouseimmature DC stimulated withL. salivarius CECT 5713.Mouse spleen dendritic cellswere coincubated with L.salivarius CECT 5713 at a ratio1:10 during 90 min. Figureshows cell surface markerexpression in control cells(white) versus bacteriastimulated cells (grey). Meanfluorescence mean intensity,Gmean geometric meanfluorescence intensity
Table 2 Inhibition of the infectivity of the HIV-1 isolates after expo-sition to cells or supernatants of L. salivarius CECT 5713
Viral isolate (receptor) Inhibition of infectivity (%)a
Bacterial pellet Cell-free supernatant
HIV BAL (CCR5) 42.3 23.5
HIV HC4 (CXCR4) 58.9 21.4
HIV C7/86 (R5/X4) 49.8 17.6
a SD values were lower than 5.4 % for all the mean values
1284 Appl Microbiol Biotechnol (2012) 94:1279–1287

prophages was responsive to this stress signal. However, noproficient phages were generated. One of the prophagesappeared to be non-inducible, while the other became inde-pendent from the host chromosome but was unable to rep-licate its own DNA. Lysis of the induced cultures waspossibly due to the coordinate expression of two phagegenes that encode the holin, which opens pores in themembrane, and the lysin, which attacks the peptidoglycanlayer (Young and Wang 2006). These genes form an operonwhose expression is independent of the formation of newviral particles, thus accounting for the observed drop in theabsorbance of the induced L. salivarius cultures. Residentprophages are frequently found in the genomes of lactic acidbacteria (Brussow and Suárez 2006; Deveau et al. 2006) andthey may account for most of the differences observedbetween strains. In many cases, obvious deletions that pre-clude completion of a lytic cycle are observed but, evenwhen apparently complete phage genomes are found, theymay not be able to generate an infective progeny (Denou etal. 2008; Martín et al. 2010a).
L. salivarius CECT5713 appeared to be very susceptibleto most of the 16 antimicrobials tested, the exceptions beingkanamycin and vancomycin. In fact, their MICs were lowerthan the cut-offs established for lactobacilli to seven out ofeight antibiotics (Table 1) by the European Food SafetyAuthority (EFSA 2008). Resistance of lactobacilli to kana-mycin and other aminoglycosides (such as neomycin orstreptomycin) has been repeatedly reported (Charteris et al.1998; Danielsen and Wind 2003) and it is thought to be dueto the lack of cytochrome-mediated transport of this class ofantibiotics (Bryan and Kwan 1981); in the case of L. sali-varius CECT 5713, this is further supported by the absencein its genome of genes encoding aminoglycoside modifyingenzymes. Similarly, most Lactobacillus species are intrinsi-cally resistant to vancomycin (Handwerger et al. 1994).Accordingly, the analysis of the L. salivarius genome didnot reveal the presence of orfs a encoding D-Ala–D-Alasynthetase. As recognized by EFSA (2008), these types ofintrinsic resistances do not represent a human health risk. Inaddition, no transmissible genes involved on antibiotic re-sistance could be found in the L. salivarius CECT 5713genome. Therefore, this strain can be considered as safefrom this point of view.
In this work, the cell surface expression of B7-2 andMHC class II determinants by mouse DC was highly stim-ulated by L. salivarius CECT 5713, which is in accordancewith previous results obtained with other mouse or human-derived DC when stimulated with different bacterial species(Rescigno et al. 1998; Rescigno et al. 2001; Braat et al.2004; Evrard et al. 2011). Bacterial induction of DC matu-ration is an active process since it has been shown that deadbacteria or inert particles, such as latex beads, do not acti-vate DCs even if they are rapidly phagocyted (Rescigno et
al. 1998). The number of studies on interactions betweenDC and lactic acid bacteria is rapidly increasing (Christensenet al. 2002; Drakes et al. 2004; Davies et al. 2009; Evrard et al.2011; Weiss et al. 2011). Recently, an in silico gene-traitmatching (the Random Forest algorithm) led to the identifica-tion of eight candidate genes in the L. plantarum genome thatmight modulate the DC cytokine response to this species(Meijerink et al. 2010).
L. salivarius CECT 5713 was isolated from breast milkand it has been suggested that at least some of the lactic acidbacteria present in the maternal gut may reach the mucosalsurfaces of the lactating mammary gland through an endoge-nous pathway involving DC–bacteria interactions (Martín etal. 2004; Jiménez et al. 2008; Arroyo et al. 2010). Studiescarried out both in vitro and in vivo have shown that DC play acritical role in the uptake of non-invasive bacteria and theirsubsequent translocation across the epithelium (Vázquez-Torres et al. 1999; Rescigno et al. 2001). In this context,bacteria must be able to activate DC, since this is the first steprequired for bacterial recognition and translocation through aDC-based mechanism.
HIV-1 infection does not occur in a sterile environment,suggesting that the mucosa-associated microbiota may in-fluence susceptibility to HIV-1 infection, either by prevent-ing or by promoting the acquisition of the virus (Lei et al.2005; Margolis and Shattock 2006). HIV-positive womencan transmit the virus to their children through breastfeeding,followed by translocation across mucosal surfaces in the oraland gastrointestinal epithelia (Wilkinson and Cunningham2006). Similarly to L. salivarius CECT 5713, it was recentlyshown that other lactic acid bacteria strains isolated frombreast milk of healthy women have the ability to inhibitinfection of HIV-1 in vitro (Martín et al. 2010b). The highestlevels of HIV-inhibitory activity were associated to four bac-terial strains, including a L. salivarius isolate, with distinctinhibitory effects against R5- and X4-tropic HIV-1. HIV-inhibitory activity was associated with both killed bacteriaand conditioned cell-free supernatant from the bacterial cul-tures, suggesting that more than one mechanism may be usedby LAB to block viral infectivity. The results of this worksuggest that molecular mimetism displayed by L. salivariusCECT 5713 may have an antagonist role towards HIV gp120and, therefore, may explain, at least partly, its activity againstHIV infectivity in vitro.
Work is now in progress to confirm the ability of certainbacterial strains present in breast milk to inhibit the infec-tivity of HIV-1 and to elucidate the mechanisms by whichthey exert such an effect, including specific interactionsbetween certain domains of the surface HIV gp120 proteinand DC receptors. Globally, the results of this work providesome clues on the relationship between some phenotypicproperties of L. salivarius CECT 5713 and their subjacentmolecular basis; in the future, such an approach may
Appl Microbiol Biotechnol (2012) 94:1279–1287 1285

contribute to a more rational selection, design and/or appli-cation of probiotic bacteria.
Acknowledgements This work was supported by CSD2007-00063(FUN-C-FOOD, Consolider-Ingenio 2010), AGL2010-15420 andAGL2010-15097 projects from the Ministerio de Ciencia e Innovación(Spain), by a research contract funded by Puleva, and by a Spain-Italybilateral collaboration program (references IT2009-0080 andIT105MD12L). S. Delgado was granted with a research contract fromJuan de la Cierva program (JCI-2008-02391). A. Maldonado was therecipient of a postdoctoral contract by the Spanish National ResearchCouncil (CSIC) through the JAE Doc programme.
References
Arroyo R, Martín V, Maldonado A, Jiménez E, Fernández L, RodríguezJM (2010) Treatment of infectious mastitis during lactation: anti-biotics versus oral administration of lactobacilli isolated from breastmilk. Clin Infect Dis 50:1551–1558
Braat H, Jong EC, van den Brande JM, Kapsenberg ML, PeppelenboschMP, van Tol EA, van Deventer SJ (2004) Dichotomy betweenLactobacillus rhamnosus and Klebsiella pneumoniae on dendriticcell phenotype and function. J Mol Med 82:197–205
Brussow H, Suárez JE (2006) Lactobacillus phages. In: Calendar R(ed) The bacteriophages, 2nd edn. Plenum Publishing Corp, NewYork, pp 653–666
Bryan LE, Kwan S (1981) Mechanisms of aminoglycoside resistanceof anaerobic bacteria and facultative bacteria grown anaerobically.J Antimicrob Chemother 8:S1–S8
Casey PG, Casey GD, Gardiner GE, Tangney M, Stanton C, Ross RP,Hill C, Fitzgerald GF (2004) Isolation and characterization ofanti-Salmonella lactic acid bacteria from the porcine gastrointes-tinal tract. Lett Appl Microbiol 39:431–438
Çataloluk O (2001) Molecular characterization of the gene encodingfor salivaricin B activity and its flanking sequences. Turk J Biol25:379–386
Caux C, Vanbervliet B, Massacrier C, Azuma M, Okumura K, LanierLL, Banchereau J (1994) B70/B7-2 is identical to CD86 and is themajor functional ligand for CD28 expressed on human dendriticcells. J Exp Med 180:1841–1847
Charteris WP, Kelly PM, Morelli L, Collins JK (1998) Antibioticsusceptibility of potentially probiotic Lactobacillus species. JFood Protect 61:1636–1643
Christensen HR, Frokiaer H, Pestka JJ (2002) Lactobacilli differential-ly modulate expression of cytokines and maturation surfacemarkers in murine dendritic cells. J Immunol 168:171–178
Claesson MJ, Li Y, Leahy S, Canchaya C, van Pijkeren JP, Cerdeno-Tarraga AM, Parkhill J, Flynn S, O'Sullivan GC, Collins JK,Higgins D, Shanahan F, Fitzgerald GF, van Sinderen D, O'ToolePW (2006) Multireplicon genome architecture of Lactobacillussalivarius. Proc Natl Acad Sci USA 103:6718–6723
Danielsen M, Wind A (2003) Susceptibility of Lactobacillus spp. toantimicrobial agents. Int J Food Microbiol 82:1–11
Davies JM, Sheil B, Shanahan F (2009) Bacterial signalling overridescytokine signalling and modifies dendritic cell differentiation.Immunology 128:e805–e815
Denou E, Pridmore RD, Ventura M, Pittet AC, Zwahlen MC, Berger B,Barretto C, Panoff JM, Brüssow H (2008) The role of prophagefor genome diversification within a clonal lineage of Lactobacil-lus johnsonii: characterization of the defective prophage LJ771. JBacteriol 190:5806–5813
Deveau H, Labrie SJ, Chopin MC, Moineau S (2006) Biodiversity andclassification of lactococcal phages. Appl Environ Microbio72:4338–4346
Díaz-Ropero M, Martin R, Sierra S, Lara-Villoslada F, Rodriguez JM,Xaus J, Olivares M (2006) Two Lactobacillus strains, isolatedfrom breast milk, differently modulate the immune response. JAppl Microbiol 102:337–343
Drakes M, Blanchard T, Czinn S (2004) Bacterial probiotic modulationof dendritic cells. Infect Immun 72:3299–3309
EFSA (2008) Update of the criteria used in the assessment of bacterialresistance to antibiotics of human or veterinary importance. EFSAJ 732:1–15
Evrard B, Coudeyras S, Dosgilbert A, Charbonnel N, Alamé J, TridonA, Forestier C (2011) Dose-dependent immunomodulation ofhuman dendritic cells by the probiotic Lactobacillus rhamnosusLcr35. PLoS One 6:e18735
Flynn S, van Sinderen D, Thornton GM, Holo H, Nes IF, Collins JK(2002) Characterization of the genetic locus responsible for theproduction of ABP-118, a novel bacteriocin produced by theprobiotic bacterium Lactobacillus salivarius subsp. salivariusUCC118. Microbiology 148:973–984
Freeman GJ, Gribben JG, Boussiotis VA, Ng JW, Restivo VA Jr,Lombard LA, Gray GS, Nadler LM (1993) Cloning of B7-2: ACTLA-4 counter-receptor that costimulates human T cell prolif-eration. Science 262:909
Handwerger S, Pucci MJ, Volk KJ, Liu J, Lee MS (1994) Vancomycin-resistant Leuconostoc mesenteroides and Lactobacillus casei syn-thesize cytoplasmic peptidoglycan precursors that terminate inlactate. J Bacteriol 176:260–264
Hart DN (1997) Dendritic cells: unique leukocyte populations whichcontrol the primary immune response. Blood 90:3245–3287
Jiménez E, Fernández L, Maldonado A, Martín R, Olivares M, Xaus J,Rodríguez JM (2008) Oral administration of lactobacilli strains iso-lated from breast milk as an alternative for the treatment of infectiousmastitis during lactation. Appl Environ Microbiol 74:4650–4655
Jiménez E, Martín R, Maldonado A, Martín V, De Segura Gomez A,Fernández L, Rodríguez JM (2010) Complete genome sequenceof Lactobacillus salivarius CECT 5713, a probiotic strain isolatedfrom human milk and infant feces. J Bacteriol 192:5266–5267
Klare I, Konstabel C, Müller-Bertling S, Reissbrodt R, Huys G,Vancanneyt M, Swings J, Goossens H, Witte W (2005) Evaluationof new broth media for microdilution antibiotic susceptibility testingof lactobacilli, lactococci, pediococci, and bifidobacteria. ApplEnviron Microbiol 71:8982–8986
Lara-Villoslada F, Sierra S, Díaz-Ropero M, Olivares M, Xaus J (2007)Safety assessment of the human milk-isolated probiotic Lactoba-cillus salivarius CECT5713. J Dairy Sci 90:3583–3589
Lei J, Wu C, Wang X, Wang H (2005) p38 MAPK-dependent andYY1-mediated chemokine receptors CCR5 and CXCR4 up-regulation in U937 cell line infected by Mycobacterium tubercu-losis or Actinobacillus actinomycetemcomitans. Biochem Bio-phys Res Commun 329:610–615
Li Y, Canchaya C, Fang F, Raftis E, Ryan KA, van Pijkeren J-P, vanSinderen D, O'Toole PW (2007) Distribution of Megaplasmids inLactobacillus salivarius and other lactobacilli. J Bacteriol189:6128–6139
Linsley PS, Ledbetter JA (1993) The role of the CD28 receptor duringT cell responses to antigen. Annu Rev Immunol 11:191
Lu L, Hsieh M, Oriss TB, Morelf PA, Starzl TE, Rao AS, ThomsonAW (1995) Generation of DC from mouse spleen cell cultures inresponse to GM-CSF: immunophenotypic and functional analy-ses. Immunology 84:127–134
Maldonado A, Ruiz-Barba JL, Jiménez-Díaz R (2003) Purification andgenetic characterization of plantaricin NC8, a novel coculture-inducible two-peptide bacteriocin from Lactobacillus plantarumNC8. Appl Environ Microbiol 69:383–389
1286 Appl Microbiol Biotechnol (2012) 94:1279–1287

Maldonado J, Lara-Villoslada F, Sierra S, Sempere L, Gómez M,Rodriguez JM, Boza J, Xaus J, Olivares M (2010) Safety andtolerance of the humanmilk probiotic strain Lactobacillus salivariusCECT5713 in 6-month-old children. Nutrition 26:1082–1087
Margolis L, Shattock R (2006) Selective transmission of CCR5-utilizing HIV-1: the ‘gatekeeper’ problem resolved? Nat RevMicrobiol 4:312–317
Martín R, Langa S, Reviriego C, Jiménez E, Marín ML, Olivares M,Boza J, Jiménez J, Fernández L, Xaus J, Rodríguez JM (2004)The commensal microflora of human milk: new perspectives forfood bacteriotherapy and probiotics. Trends Food Sci Technol15:121–127
Martín R, Jiménez E, Olivares M, Marín ML, Fernández L, Xaus J,Rodríguez JM (2006) Lactobacillus salivarius CECT 5713, apotential probiotic strain isolated from infant feces and breastmilk of a mother-child pair. Int J Food Microbiol 112:35–43
Martín R, Soberón N, Escobedo S, Suárez JE (2009) Bacteriophageinduction versus vaginal homeostasis: role of H2O2 in the selectionof Lactobacillus defective prophages. Int Microbiol 12:131–136
Martín R, Escobedo S, Suárez JE (2010a) Induction, structural charac-terization and genome sequence of Lv1, a prophage from a humanvaginal. L jensenii strain Int Microbiol 13:113–121
Martín V, Maldonado A, Fernández L, Rodríguez JM, Connor RI(2010b) Inhibition of human immunodeficiency virus type 1 bylactic acid bacteria from human breastmilk. Breastfeeding Med5:153–158
Meijerink M, van Hemert S, Taverne N, Wels M, de Vos P, Bron PA,Savelkoul HF, van Bilsen J, Kleerebezem M, Wells JM (2010)Identification of genetic loci in Lactobacillus plantarum thatmodulate the immune response of dendritic cells using compara-tive genome hybridization. PLoS One 5:e10632
Neville BA, O’Toole PW (2010) Probiotic properties of Lactobacillussalivarius and closely related Lactobacillus species. FutureMicrobiol 5:759–774
Olivares M, Díaz-Ropero MP, Martín R, Rodríguez JM, Xaus J (2006)Antimicrobial potential of four Lactobacillus strains isolated frombreast milk. J Appl Microbiol 101:72–79
Pérez-Cano FJ, Dong H, Yaqoob P (2010) In vitro immunomodulatoryactivity of Lactobacillus fermentum CECT5716 and Lactobacillussalivarius CECT5713: two probiotic strains isolated from humanbreast milk. Inmunobiology 12:996–1004
Raftis EJ, Salvetti E, Torriani S, Felis GE, O'Toole PW (2011) Genomicdiversity of Lactobacillus salivarius. Appl Environ Microbiol77:954–965
Rescigno M, Citterio S, Thery C, Rittig M, Medaglini D, Pozzi G,Amigorena S, Ricciardi-Castagnoli P (1998) Bacteria-inducedneo-biosynthesis, stabilization, and surface expression of func-tional class I molecules in mouse dendritic cells. Proc Natl AcadSci USA 95:5229–5234
Rescigno M, Urbano M, Valzasina B, Francolín M, Rotta G, BonasioR, Granucci F, Kraehenbuhl J, Ricciardi-Castagnoli P (2001)Dendritic cells express tight junction proteins and penetrate gutepithelial monolayers to sample bacteria. Nature Immunol 2:361–367
Rogosa M, Wiseman RF, Mitchell JA, Disraely MN, Beaman AJ(1953) Species differentiation of oral lactobacilli from man in-cluding descriptions of Lactobacillus salivarius nov. spec. andLactobacillus cellobiosus nov. spec. J Bacteriol 65:681–699
Soberón N, Martin R, Suarez JE (2007) New method for evaluation ofgenotoxicity, based on the use of real-time PCR and lysogenicGram-positive and Gram-negative bacteria. Appl Environ Micro-biol 73:2815–2819
Vázquez-Torres A, Jones-Carson J, Baumler AJ, Falkow S, Valdivia R,Brown W, Le M, Berggren R, Parks WT, Fang FC (1999) Extra-intestinal dissemination of Salmonella by CD18-expressing phag-ocytes. Nature 401:804–808
Weiss G, Christensen HR, Zeuthen LH, Vogensen FK, Jakobsen M,Frøkiær H (2011) Lactobacilli and bifidobacteria induce differen-tial interferon-β profiles in dendritic cells. Cytokine 56:520–530
Wilkinson J, Cunningham AL (2006) Mucosal transmission of HIV-1:first stop dendritic cells. Curr Drug Targets 7:1563–1569
Young R, Wang IN (2006) Phage lysis. In: Calendar R (ed) Thebacteriophages, 2nd edn. Plenum Publishing Corp, New York,pp 104–125
Appl Microbiol Biotechnol (2012) 94:1279–1287 1287





























