Embed Size (px)
Citation preview

European Journal of Soil Science, 2012 doi: 10.1111/j.1365-2389.2012.01457.x
Characterization of dissolved organic matterin decomposing Norway spruce and silver birch litter
O . K i i k k i l a , V . K i t u n e n , P . S p e t z & A . S m o l a n d e rFinnish Forest Research Institute, Vantaa Research Unit, PO Box 18, FIN 01301 Vantaa, Finland
Summary
Dissolved organic matter (DOM) plays an important role in transport, storage and cycling of carbon (C) andnitrogen (N) in forest soils where litter is one of the main sources. The aim was to study the amount andcharacteristics of DOM leached from freshly fallen litters of silver birch (Betula pendula Roth.), Norway spruce(Picea abies (L.) Karst.) and their mixture during decomposition. DOM was collected after irrigation on eightoccasions during 252 days incubation in the laboratory at about 18◦C, including one freeze-thaw cycle. Duringthe incubation about 33–35% of C from birch and spruce litter and 40% of C from their mixture was lost.The total cumulative flux of dissolved organic carbon (DOC) from the mixture of litters was approximately40% larger than that from single litters. The flux of DOC, DON, phenolic compounds and proteins followed atwo-stage pattern during decomposition. In the first stage the initially large fluxes decreased gradually. In thesecond stage, after freezing and thawing, the fluxes tended to increase again. Mixing birch and spruce littersand a freeze-thaw cycle seems to increase the decomposition of litter and result in the increased flux of DOC,DON and phenolic compounds. The flux of hemicelluloses and the degradability of DOM were large at the firstleaching occasion and decreased during the incubation. Birch had a 40% larger total flux of easily degradableDOM than spruce, supporting the previous consistent signs of greater microbial biomass and activities relatedto C and N cycling in soil under birch than under spruce. It is known that recalcitrant DOM might be stabilizedwhereas labile DOM may promote microbial activity and nutrient cycling. We conclude that the storage andcycling of C and N is affected by both tree species and degradation stage of litter in forest soils.
Introduction
The rate of litter decomposition can be predicted from environ-mental factors as well as from the chemical composition of litter,as documented in recent reviews (Zhang et al., 2008; Prescott,2010). The main findings are that deciduous litter generallydecomposes faster than coniferous litter and that litter decompo-sition is largely controlled by lignin degradation. However, litterdecomposition is a dynamic process in which compounds alterand degrade in close connection to each other and decompos-ing organisms. The decomposition rate is known to be affectedby the concentration and molecular structure of N-compounds,lignin, phenolic compounds and carbohydrates (Valachovic et al.,2004). The role of carbohydrates such as cellulose and hemi-celluloses is not clear. They are generally regarded as easilydegradable. However, cellulose and hemicelluloses that are chem-ically connected to lignin seem to be resistant to decomposi-tion (Thevenot et al., 2010). Water-soluble hemicelluloses weredegraded rapidly at the early stages of decomposition when
Correspondence: O. Kiikkila. E-mail: [email protected]
Received 25 May 2011; revised version accepted 8 March 2012
degradation of cellulose and lignin was slow (Sariyildiz & Ander-son, 2003; Snajdr et al., 2011). However, after 2 years of decom-position, some of the hemicelluloses remained whereas cellulosehad decomposed almost completely (Snajdr et al., 2011).
Litter is one of the main sources of dissolved organic matter(DOM) in forest soils. DOM plays an important role in transport,storage and cycling of C and N (Kalbitz et al., 2007). Theturnover, production and degradation of dissolved organic carbon(DOC) and nitrogen (DON) is an important process linked tomicrobial activities, and thus to C and N cycling in soil whichhas been observed to be greater under silver birch (Betula pendulaRoth.) than Norway spruce (Picea abies (L.) Karst.) (Smolander& Kitunen, 2011). There is a lack of knowledge concerning theamount and degradability of DOM leached from decomposinglitter in boreal forest soils. A large part of easily degradable, water-soluble carbohydrates and phenolic compounds has been observedto leach in early stages of litter decomposition and more refractory,lignin-derived phenolic compounds have been observed to leach inlater stages of litter decomposition (Don & Kalbitz, 2005; Kalbitzet al., 2006; Hagedorn & Machwitz, 2007). Fresh litter of silverbirch has been found to release more water-extractable phenolic
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science 1

2 O. Kiikkila et al.
compounds than that of Norway spruce (Kuiters & Sarink, 1986).In addition, different patterns of leaching of DOM during thedecomposition have been observed for deciduous and coniferousspecies in general. Deciduous species had a larger DOC flux atthe beginning of decomposition whereas coniferous species, whichhave greater lignin content, showed increasing DOC flux when themass loss exceeded 20% (Kalbitz et al., 2006).
Litter mixtures have been reported to decompose faster thansingle species litter in about half of the cases reviewed by Gartner& Gordon (2004). Mixing birch with spruce litter has beensuggested to increase the spruce litter decomposition (Brandtberg& Lundkvist, 2004). Thus mixing of litters might have an effecton quantity and properties of DOM leached from decomposinglitter. Also, freeze-thaw cycles have been observed to increaselitter decomposition (Wu et al., 2010). A rapid pulse of respirationfollowing thawing is thought to result partly from the easilydegradable substrates released by microbial death during freezing(Matzner & Borken, 2008) but clear evidence for the lysisof microorganisms is lacking (Hentschell et al., 2008). To ourknowledge there are no studies where the leaching of DOM afterfreeze-thaw cycles or from litter mixtures has been measured.
The aim of this study was to measure the quantity and quality ofDOM produced during the decomposition of litter. We studied thelitters of two important tree species in Finland, Norway spruceand silver birch. It was hypothesized that the tree species andmixing of litters affect the quality and quantity of DOM leachedfrom decomposing litters. We performed a long-term litter-columnexperiment in the laboratory and studied compound groups thatare known to include labile compounds. The concentrations of C,N and hemicellulotic and phenolic compounds were measured inDOM leaching from decomposing litter on eight occasions duringthe decomposition as well as in initial and decomposed solid lit-ter. In order to assess the turnover of DOM we also measured thedegradability of DOM and calculated the total amount of easilydegradable DOM leached from litters. As a measure of degrad-ability we used the growth rate of bacteria and fungi in DOM,which we also related to the chemical composition of DOM.
Materials and Methods
Study site and sampling
Representative samples of freshly fallen litter were collected fromthe silver birch-Norway spruce experiment in Eno, central-easternFinland. Three replicate plots (40 × 40 m) for both tree specieswere sampled separately. Stand characteristics were determined in2002 and presented in detail by Smolander et al. (2005). As birchplots had understorey vegetation consisting of grey alder (Alnusincana) and rowan (Sorbus aucuparia), a negligible amount ofthese leaves was included in the birch litter. The aim was tolose as little DOM from litter in the field as possible. However,an unknown amount of DOM was lost from birch litter becausethe collection period was rainy. The litter was collected duringthe main litter fall, birch leaf litter in late September and spruceneedle litter in March–April, by spreading nets (area 3 m2) over
the forest floor for approximately two weeks. Snowfall containingspruce litter was brought into the laboratory and melted at roomtemperature. Litter was air-dried, larger branches were removedand litter was kept frozen (−18◦C) until the experiment.
Litter column experiment
The decomposition of birch, spruce and a mixture of birch andspruce litter (1:1, according to mass loss on ignition, LOI: birch83%, spruce 88%) was followed in litter columns for 252 days at18–20◦C in the laboratory. Litter, intact leaves or needles equalto 90 g LOI (about 100 g dry mass), was weighed to each column(volume 2 l, diameter 18 cm). Two litter bags containing 1 g drymass (40◦C) of litter were placed in the middle of each column inorder to measure the mass loss of litter. Two laboratory replicatesof columns were prepared for each of the six sampling plots.DOM and litter of the laboratory replicates were combined infurther analysis and the means of the mass losses in the two litterbags per sampling were used as observations. Litter mixtures wereprepared respectively by combining one randomly selected birchand spruce plot. A microbial inoculum (30 ml) was added to eachcolumn. The inoculum was prepared from a mixture of birch andspruce humus layers as described in Kiikkila et al. (2011).
In order to keep the litter moist, the columns were moistenedevery 3 days with artificial rainwater (pH 4.7), prepared accordingto Lindroos et al. (2007). After 74 days of incubation the columnswere frozen (−18◦C) for 2 weeks. Samples were collected on13 leaching occasions (first after 10 days) when the columns wereirrigated with a volume of cold (4◦C) artificial rainwater thatcorresponded to 20–40 mm rain. DOM leached first by gravityand after water suction for 30 s was collected. The total volumecollected was measured in order to calculate the amount ofcompounds leached from the litter (flux). The flux during theincubation was calculated per g C in initial litter. In addition,the total cumulative flux was calculated per g C in decomposedlitter. The solution was filtered finally through a 0.45-μm PESmembrane (Pall Corporation, Port Washington, NY, USA) andfrozen (−18◦C) until analysis.
The DOC was measured on all 13 sampling occasions whereasthe other analyses of DOM were measured only on eight leachingoccasions (four before and four after freezing; the first after14 days of incubation). The total flux was based on all 13 leachingoccasions in the experiment. The volumes of the five leachingoccasions, when only DOC was measured, were combined withthe next leaching occasion.
Determination of chemical characteristics of litter
The litter bags were sampled after 74 and 252 days of decompo-sition and the mass loss after drying was determined. Carbon, N,phenolic compounds and hemicelluloses were measured initiallyand after the 252 days of decomposition. The initial content of thelitter mixture was calculated as the mean of the values of spruceand birch. The samples were measured for the moisture content
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science

DOM from tree litter 3
(dry mass; 105◦C for 20 hour) and LOI (500◦C, 4 hour). Total Cand N were analysed on a LECO CHN-2000 analyser (St. Joseph,MI, USA).
The concentration of total water-soluble phenolic compoundswas measured from initial and decomposed litters by spectropho-tometry. We used the Folin-Ciocalteu method (Box, 1981) asdescribed by Kanerva et al. (2007) with 0.5 g dry and ground sam-ple and tannic acid as standard. High-molecular-weight (HMW)phenolic compounds were precipitated with casein, the total andlow-molecular-weight (LMW) phenolic compounds were mea-sured and the concentration of HMW phenolic compounds wascalculated.
The concentration of hemicellulotic compounds was determinedfrom initial and decomposed litters using the method described bySundberg et al. (1996) for wood fibres. A dried and ground sample(10 mg dry mass) was subjected to acid methanolysis, whichdegrades non-cellulosic polysaccharides into their monomericsugar units. The concentration of neutral and acidic sugar unitswas measured by gas chromatograph (GC- 2010, ShimadzuCorporation, Duisburg, Germany). The concentration of totalhemicellulotic compounds (hemicelluloses) was calculated as thesum of individual sugars: arabinose (Ara), rhamnose (Rha), xylose(Xyl), mannose (Man), galactose (Gal), glucose (Glc) and uronicacids: glucuronic acid (GlcA), galacturonic acid (GalA) and 4-O-Me-glucuronic acid (Me-glc) including pectin.
Determination of DOM properties
The concentrations of total and inorganic dissolved C and totaldissolved N were measured with a Multi N/C analyser (Ana-lyticjena, Wembley, UK) and inorganic N with a flow injectionanalyser (FIA 5012, Tecator, Hillerød, Denmark). DOC and DONwere calculated as a difference between total and inorganic Cand N, respectively. The concentration of proteins was measuredwith a Bio-Rad protein assay, based on the method of Brad-ford (1976) using spectrophotometry and bovine serum albumin(BSA) as a standard. The pH was measured from the DOMsolutions after adjusting the DOC concentration to 30 mg l−1
DOC. The concentrations of total phenolic compounds were mea-sured in 0.5–2 ml DOM samples as described above. HMW andLMW phenolic compounds could not be separated. The concen-tration of hemicellulotic sugar units in DOM were determinedin 0.5–2.0 ml samples after freeze-drying and methanolysis asdescribed above. Monomeric sugar units in DOM were similarlydetermined by GC after freeze-drying and silylation according toWillfor et al. (2005).
The availability of DOM to bacteria was assessed by measuringthe bacterial growth rate in the solution with the 3H-thymidineincorporation technique modified to DOM studies by Kiikkilaet al. (2006). Bacterial growth rates were measured after adjustingthe DOC concentration to 30 mg l−1 DOC. Ultrapure water wasused as a control sample. The natural logarithm of the ratioof incorporation of 3H-thymidine (that is radioactivity in DOMsolution) to the incorporation in water was used as a measure
of the relative availability of DOM to bacteria (Tdr). Tdr wasmultiplied by the amount of DOC that leached from litter C(Tdr × mg DOC g−1 C) in order to assess the total amountof easily degradable DOM (cumulative Tdr) leached from thelitters.
The availability of DOM to fungi was assessed as fungal growthrate in DOM solution using the 14C-acetate-in-ergosterol techniquemodified by Kiikkila et al. (2006). Fungal growth rates weremeasured after adjusting the DOC concentration to 10 mg l−1
DOC and the pH to 4.5. The natural logarithm of the ratio (14C-acetate in ergosterol in DOM solution):(concentration of totalergosterol) was used as the measure of relative availability ofDOM to fungi (Ac-erg). The ratio Ac-erg:Tdr was calculated inorder to assess the importance of bacteria and fungi in degradingDOM during decomposition.
Data analysis
One-way Anova followed by Tukey’s test was used to studythe differences in the chemical characteristics of litter and littersolution among birch, spruce and their mixture. The necessarylogarithmic transformations were made in order to normalize thedistribution of the variables. The statistical significance for alltests was set at P < 0.05.
The sugars that were lost from the litters were calculated as thedifference in the concentrations between initial and decomposedlitter. The proportional distribution of sugars in initial, decom-posed and lost litter, as well as in total DOM, were analysedtogether by principal component analyses (PCA) using correlationmatrix with SPSS 15.0 software (SPSS Inc., Chicago, IL, USA).Sample scores are plotted along PC1 and PC2. In the same figurethe correlations between the original variables and the new axesare presented as vectors. Another PCA was performed on the pro-portional distribution of sugars in DOM at each leaching occasion.
Correlation analyses were used to assess the availability ofdifferent compound groups to bacteria or fungi. A Pearsoncorrelation for Tdr and Ac-erg against the concentration of totalphenolic compounds (mg g−1 DOC) and total hemicelluloses(mg g−1 DOC) in incubation solution was calculated. Theconcentration of individual sugars in DOM against Tdr and Ac-erg was combined with canonical correlation analysis (CCA) usingSAS 9.1 software (SAS Institute Inc., Cary, NC, USA). The largestcorrelations between sugars and their canonical variables weredrawn as vectors on respective axes.
Results
Composition of litter
At the beginning of the experiment the concentrations of C, N,water-soluble total: HMW or LMW phenolic compounds andtotal hemicelluloses did not differ significantly between birch andspruce litters (Table 1). The distribution of hemicellulotic sugarsdiffered and is discussed later.
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science
4 O. Kiikkila et al.
Table 1 Composition of initial and decomposed spruce and birch litterand the litter mixture (mean ± SD, n = 3)
Spruce Birch Mixture
C / % Initial 48.0 ± 0.2 50.8 ± 0.2 49.5 ± 0.2Decomposed 47.5 ± 0.4 47.9 ± 0.4 47.5 ± 0.2
N / % Initial 1.0 ± 0.1 1.0 ± 0.1 1.0 ± 0.1Decomposed 1.3 ± 0.1 1.4 ± 0.1 1.5 ± 0.1
C:N ratio Initial 47 ± 3 51 ± 1 49 ± 2Decomposed 35 ± 1 35 ± 1 31 ± 1
Phenolic compounds / mg TA g−1 CTotal Initial 19.2 ± 3.4 16.9 ± 1.4 18.1 ± 1.0
Decomposed 4.2 ± 0.3 2.3 ± 0.3 3.5 ± 0.1Low molecular
weightInitial 7.5 ± 1.2 6.8 ± 0.5 7.1 ± 0.4
Decomposed 2.3 ± 0.1 1.9 ± 0.2 2.5 ± 0.1High molecular
weightInitial 11.7 ± 2.4 10.2 ± 1.1 10.9 ± 0.7
Decomposed 1.7 ± 0.2 0.4 ± 0.2 0.9 ± 0.2
Hemicelluloses / mg g−1 CInitial 400 ± 7 380 ± 15 391 ± 6Decomposed 320 ± 4 361 ± 1 320 ± 14
TA = tannic acid.
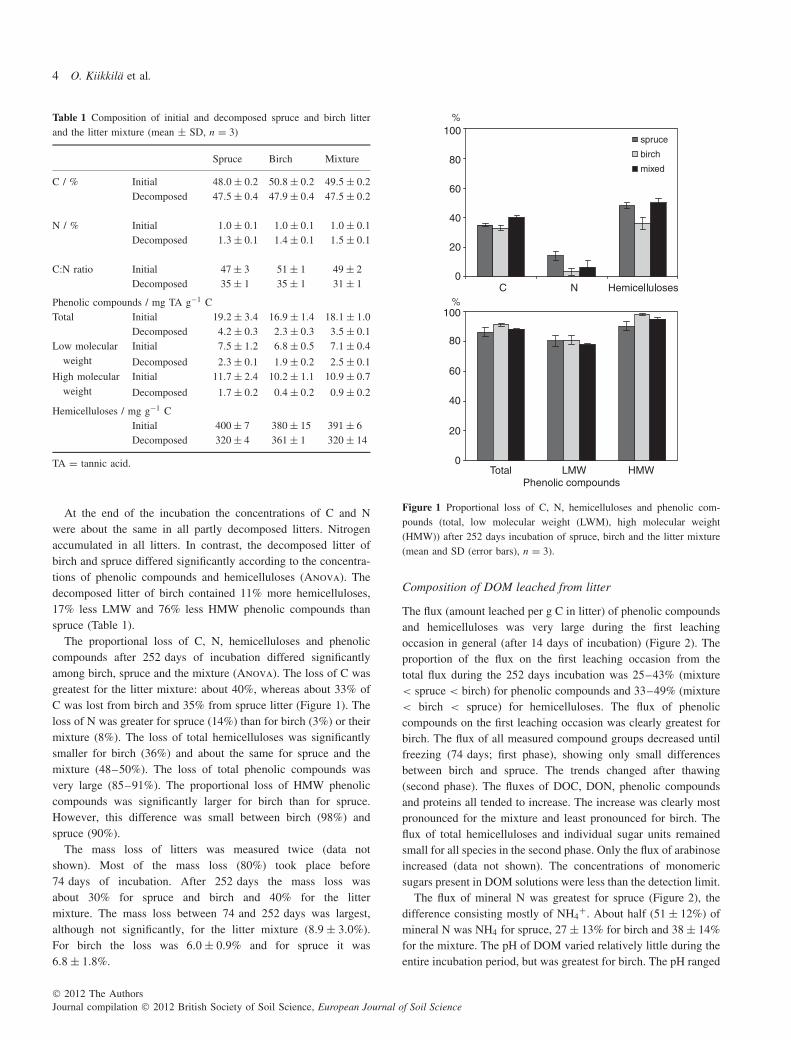
At the end of the incubation the concentrations of C and Nwere about the same in all partly decomposed litters. Nitrogenaccumulated in all litters. In contrast, the decomposed litter ofbirch and spruce differed significantly according to the concentra-tions of phenolic compounds and hemicelluloses (Anova). Thedecomposed litter of birch contained 11% more hemicelluloses,17% less LMW and 76% less HMW phenolic compounds thanspruce (Table 1).
The proportional loss of C, N, hemicelluloses and phenoliccompounds after 252 days of incubation differed significantlyamong birch, spruce and the mixture (Anova). The loss of C wasgreatest for the litter mixture: about 40%, whereas about 33% ofC was lost from birch and 35% from spruce litter (Figure 1). Theloss of N was greater for spruce (14%) than for birch (3%) or theirmixture (8%). The loss of total hemicelluloses was significantlysmaller for birch (36%) and about the same for spruce and themixture (48–50%). The loss of total phenolic compounds wasvery large (85–91%). The proportional loss of HMW phenoliccompounds was significantly larger for birch than for spruce.However, this difference was small between birch (98%) andspruce (90%).
The mass loss of litters was measured twice (data notshown). Most of the mass loss (80%) took place before74 days of incubation. After 252 days the mass loss wasabout 30% for spruce and birch and 40% for the littermixture. The mass loss between 74 and 252 days was largest,although not significantly, for the litter mixture (8.9 ± 3.0%).For birch the loss was 6.0 ± 0.9% and for spruce it was6.8 ± 1.8%.
Figure 1 Proportional loss of C, N, hemicelluloses and phenolic com-pounds (total, low molecular weight (LWM), high molecular weight(HMW)) after 252 days incubation of spruce, birch and the litter mixture(mean and SD (error bars), n = 3).
Composition of DOM leached from litter
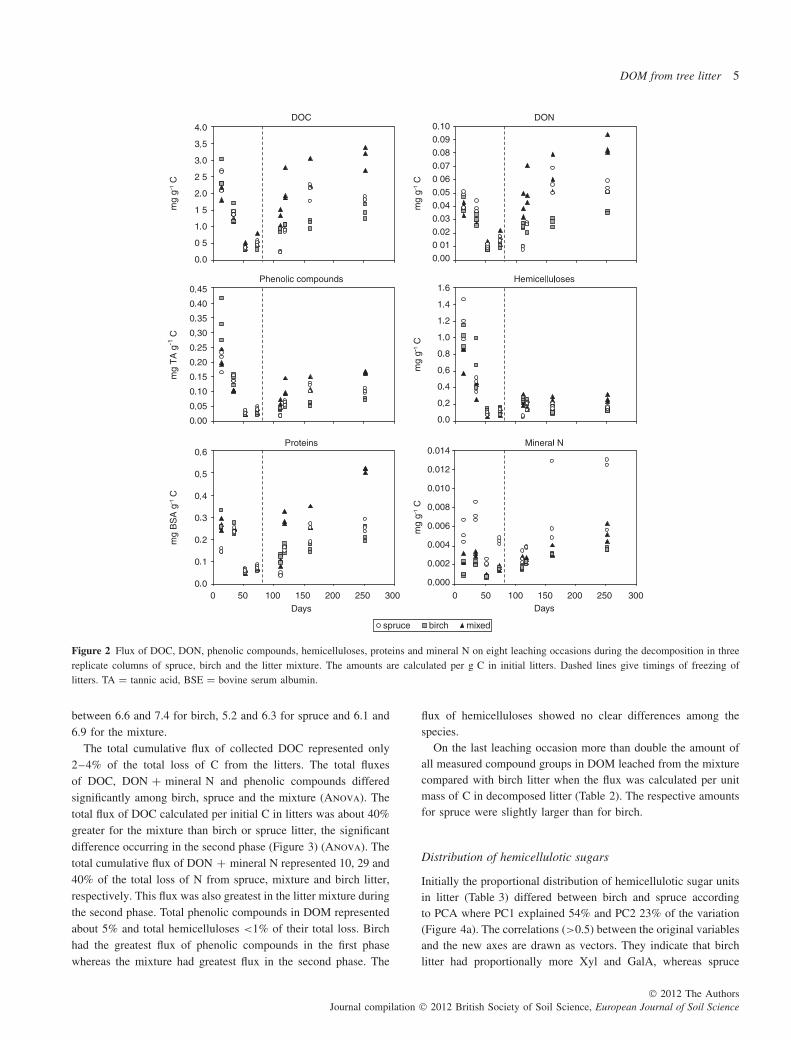
The flux (amount leached per g C in litter) of phenolic compoundsand hemicelluloses was very large during the first leachingoccasion in general (after 14 days of incubation) (Figure 2). Theproportion of the flux on the first leaching occasion from thetotal flux during the 252 days incubation was 25–43% (mixture< spruce < birch) for phenolic compounds and 33–49% (mixture< birch < spruce) for hemicelluloses. The flux of phenoliccompounds on the first leaching occasion was clearly greatest forbirch. The flux of all measured compound groups decreased untilfreezing (74 days; first phase), showing only small differencesbetween birch and spruce. The trends changed after thawing(second phase). The fluxes of DOC, DON, phenolic compoundsand proteins all tended to increase. The increase was clearly mostpronounced for the mixture and least pronounced for birch. Theflux of total hemicelluloses and individual sugar units remainedsmall for all species in the second phase. Only the flux of arabinoseincreased (data not shown). The concentrations of monomericsugars present in DOM solutions were less than the detection limit.
The flux of mineral N was greatest for spruce (Figure 2), thedifference consisting mostly of NH4
+. About half (51 ± 12%) ofmineral N was NH4 for spruce, 27 ± 13% for birch and 38 ± 14%for the mixture. The pH of DOM varied relatively little during theentire incubation period, but was greatest for birch. The pH ranged
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science
DOM from tree litter 5
Figure 2 Flux of DOC, DON, phenolic compounds, hemicelluloses, proteins and mineral N on eight leaching occasions during the decomposition in threereplicate columns of spruce, birch and the litter mixture. The amounts are calculated per g C in initial litters. Dashed lines give timings of freezing oflitters. TA = tannic acid, BSE = bovine serum albumin.
between 6.6 and 7.4 for birch, 5.2 and 6.3 for spruce and 6.1 and6.9 for the mixture.
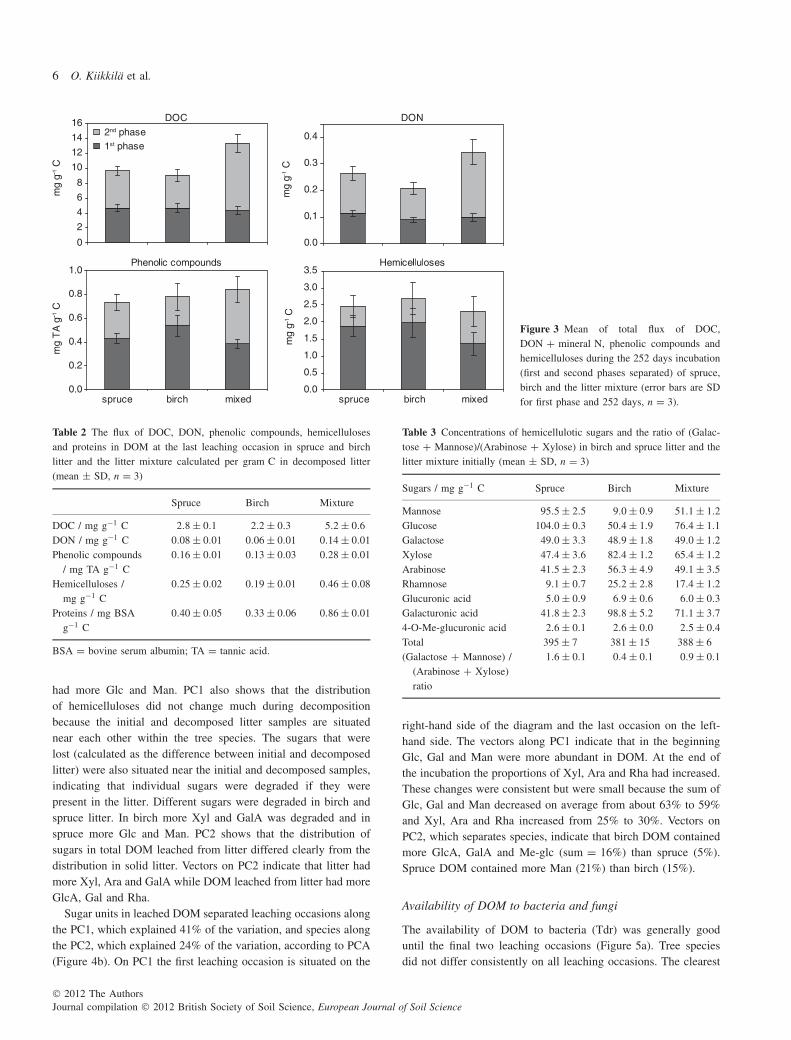
The total cumulative flux of collected DOC represented only2–4% of the total loss of C from the litters. The total fluxesof DOC, DON + mineral N and phenolic compounds differedsignificantly among birch, spruce and the mixture (Anova). Thetotal flux of DOC calculated per initial C in litters was about 40%greater for the mixture than birch or spruce litter, the significantdifference occurring in the second phase (Figure 3) (Anova). Thetotal cumulative flux of DON + mineral N represented 10, 29 and40% of the total loss of N from spruce, mixture and birch litter,respectively. This flux was also greatest in the litter mixture duringthe second phase. Total phenolic compounds in DOM representedabout 5% and total hemicelluloses <1% of their total loss. Birchhad the greatest flux of phenolic compounds in the first phasewhereas the mixture had greatest flux in the second phase. The
flux of hemicelluloses showed no clear differences among thespecies.
On the last leaching occasion more than double the amount ofall measured compound groups in DOM leached from the mixturecompared with birch litter when the flux was calculated per unitmass of C in decomposed litter (Table 2). The respective amountsfor spruce were slightly larger than for birch.
Distribution of hemicellulotic sugars
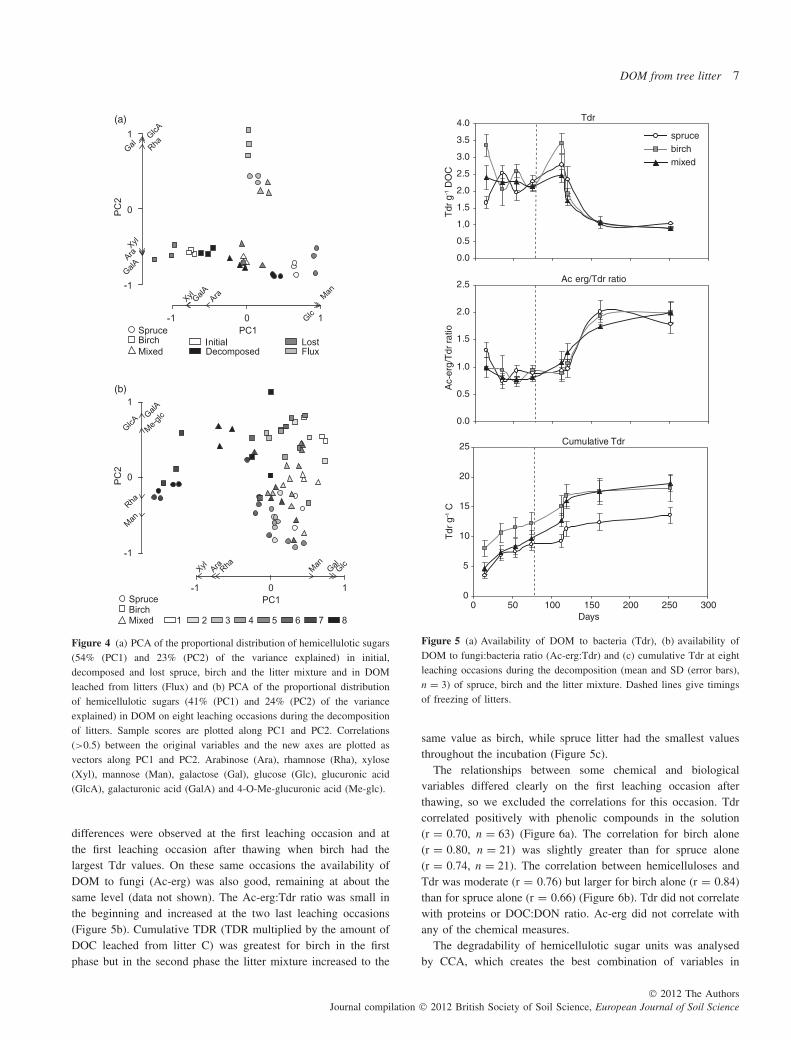
Initially the proportional distribution of hemicellulotic sugar unitsin litter (Table 3) differed between birch and spruce accordingto PCA where PC1 explained 54% and PC2 23% of the variation(Figure 4a). The correlations (>0.5) between the original variablesand the new axes are drawn as vectors. They indicate that birchlitter had proportionally more Xyl and GalA, whereas spruce
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science
6 O. Kiikkila et al.
Figure 3 Mean of total flux of DOC,DON + mineral N, phenolic compounds andhemicelluloses during the 252 days incubation(first and second phases separated) of spruce,birch and the litter mixture (error bars are SDfor first phase and 252 days, n = 3).
Table 2 The flux of DOC, DON, phenolic compounds, hemicellulosesand proteins in DOM at the last leaching occasion in spruce and birchlitter and the litter mixture calculated per gram C in decomposed litter(mean ± SD, n = 3)
Spruce Birch Mixture
DOC / mg g−1 C 2.8 ± 0.1 2.2 ± 0.3 5.2 ± 0.6DON / mg g−1 C 0.08 ± 0.01 0.06 ± 0.01 0.14 ± 0.01Phenolic compounds
/ mg TA g−1 C0.16 ± 0.01 0.13 ± 0.03 0.28 ± 0.01
Hemicelluloses /mg g−1 C
0.25 ± 0.02 0.19 ± 0.01 0.46 ± 0.08
Proteins / mg BSAg−1 C
0.40 ± 0.05 0.33 ± 0.06 0.86 ± 0.01
BSA = bovine serum albumin; TA = tannic acid.
had more Glc and Man. PC1 also shows that the distributionof hemicelluloses did not change much during decompositionbecause the initial and decomposed litter samples are situatednear each other within the tree species. The sugars that werelost (calculated as the difference between initial and decomposedlitter) were also situated near the initial and decomposed samples,indicating that individual sugars were degraded if they werepresent in the litter. Different sugars were degraded in birch andspruce litter. In birch more Xyl and GalA was degraded and inspruce more Glc and Man. PC2 shows that the distribution ofsugars in total DOM leached from litter differed clearly from thedistribution in solid litter. Vectors on PC2 indicate that litter hadmore Xyl, Ara and GalA while DOM leached from litter had moreGlcA, Gal and Rha.
Sugar units in leached DOM separated leaching occasions alongthe PC1, which explained 41% of the variation, and species alongthe PC2, which explained 24% of the variation, according to PCA(Figure 4b). On PC1 the first leaching occasion is situated on the
Table 3 Concentrations of hemicellulotic sugars and the ratio of (Galac-tose + Mannose)/(Arabinose + Xylose) in birch and spruce litter and thelitter mixture initially (mean ± SD, n = 3)
Sugars / mg g−1 C Spruce Birch Mixture
Mannose 95.5 ± 2.5 9.0 ± 0.9 51.1 ± 1.2Glucose 104.0 ± 0.3 50.4 ± 1.9 76.4 ± 1.1Galactose 49.0 ± 3.3 48.9 ± 1.8 49.0 ± 1.2Xylose 47.4 ± 3.6 82.4 ± 1.2 65.4 ± 1.2Arabinose 41.5 ± 2.3 56.3 ± 4.9 49.1 ± 3.5Rhamnose 9.1 ± 0.7 25.2 ± 2.8 17.4 ± 1.2Glucuronic acid 5.0 ± 0.9 6.9 ± 0.6 6.0 ± 0.3Galacturonic acid 41.8 ± 2.3 98.8 ± 5.2 71.1 ± 3.74-O-Me-glucuronic acid 2.6 ± 0.1 2.6 ± 0.0 2.5 ± 0.4Total 395 ± 7 381 ± 15 388 ± 6(Galactose + Mannose) /
(Arabinose + Xylose)ratio
1.6 ± 0.1 0.4 ± 0.1 0.9 ± 0.1
right-hand side of the diagram and the last occasion on the left-hand side. The vectors along PC1 indicate that in the beginningGlc, Gal and Man were more abundant in DOM. At the end ofthe incubation the proportions of Xyl, Ara and Rha had increased.These changes were consistent but were small because the sum ofGlc, Gal and Man decreased on average from about 63% to 59%and Xyl, Ara and Rha increased from 25% to 30%. Vectors onPC2, which separates species, indicate that birch DOM containedmore GlcA, GalA and Me-glc (sum = 16%) than spruce (5%).Spruce DOM contained more Man (21%) than birch (15%).
Availability of DOM to bacteria and fungi
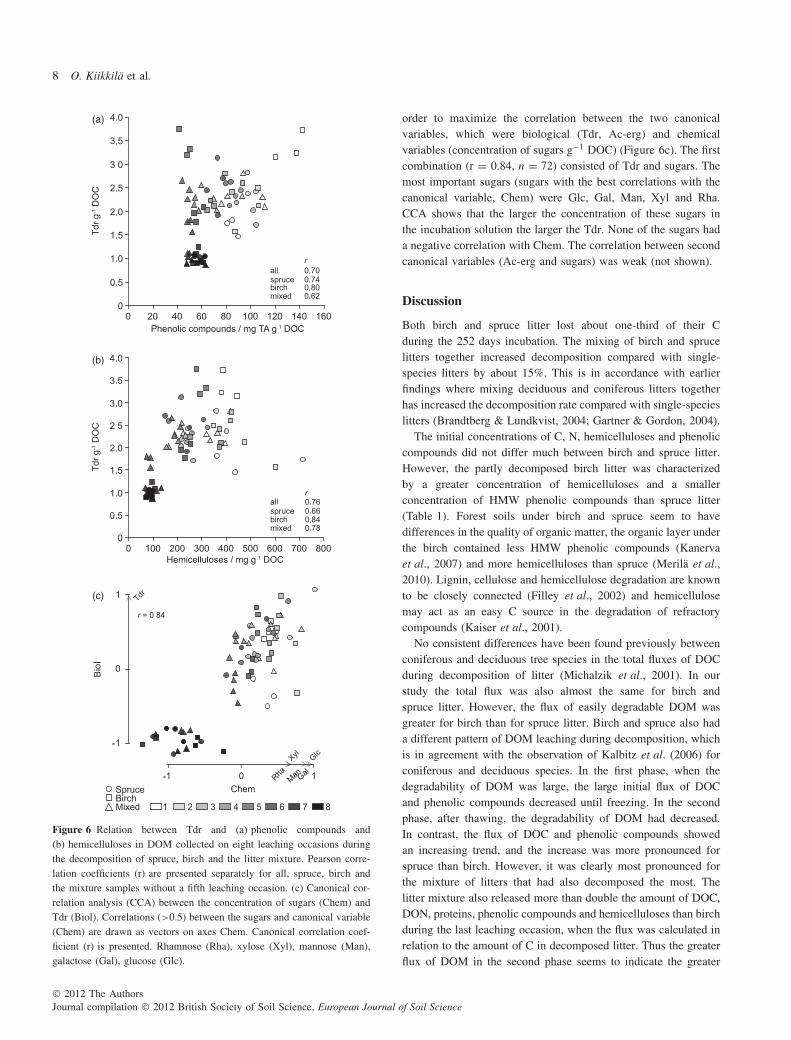
The availability of DOM to bacteria (Tdr) was generally gooduntil the final two leaching occasions (Figure 5a). Tree speciesdid not differ consistently on all leaching occasions. The clearest
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science
DOM from tree litter 7
(a)
(b)
Figure 4 (a) PCA of the proportional distribution of hemicellulotic sugars(54% (PC1) and 23% (PC2) of the variance explained) in initial,decomposed and lost spruce, birch and the litter mixture and in DOMleached from litters (Flux) and (b) PCA of the proportional distributionof hemicellulotic sugars (41% (PC1) and 24% (PC2) of the varianceexplained) in DOM on eight leaching occasions during the decompositionof litters. Sample scores are plotted along PC1 and PC2. Correlations(>0.5) between the original variables and the new axes are plotted asvectors along PC1 and PC2. Arabinose (Ara), rhamnose (Rha), xylose(Xyl), mannose (Man), galactose (Gal), glucose (Glc), glucuronic acid(GlcA), galacturonic acid (GalA) and 4-O-Me-glucuronic acid (Me-glc).
differences were observed at the first leaching occasion and atthe first leaching occasion after thawing when birch had thelargest Tdr values. On these same occasions the availability ofDOM to fungi (Ac-erg) was also good, remaining at about thesame level (data not shown). The Ac-erg:Tdr ratio was small inthe beginning and increased at the two last leaching occasions(Figure 5b). Cumulative TDR (TDR multiplied by the amount ofDOC leached from litter C) was greatest for birch in the firstphase but in the second phase the litter mixture increased to the
Figure 5 (a) Availability of DOM to bacteria (Tdr), (b) availability ofDOM to fungi:bacteria ratio (Ac-erg:Tdr) and (c) cumulative Tdr at eightleaching occasions during the decomposition (mean and SD (error bars),n = 3) of spruce, birch and the litter mixture. Dashed lines give timingsof freezing of litters.
same value as birch, while spruce litter had the smallest valuesthroughout the incubation (Figure 5c).
The relationships between some chemical and biologicalvariables differed clearly on the first leaching occasion afterthawing, so we excluded the correlations for this occasion. Tdrcorrelated positively with phenolic compounds in the solution(r = 0.70, n = 63) (Figure 6a). The correlation for birch alone(r = 0.80, n = 21) was slightly greater than for spruce alone(r = 0.74, n = 21). The correlation between hemicelluloses andTdr was moderate (r = 0.76) but larger for birch alone (r = 0.84)than for spruce alone (r = 0.66) (Figure 6b). Tdr did not correlatewith proteins or DOC:DON ratio. Ac-erg did not correlate withany of the chemical measures.
The degradability of hemicellulotic sugar units was analysedby CCA, which creates the best combination of variables in
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science
8 O. Kiikkila et al.
(a)
(b)
(c)
Figure 6 Relation between Tdr and (a) phenolic compounds and(b) hemicelluloses in DOM collected on eight leaching occasions duringthe decomposition of spruce, birch and the litter mixture. Pearson corre-lation coefficients (r) are presented separately for all, spruce, birch andthe mixture samples without a fifth leaching occasion. (c) Canonical cor-relation analysis (CCA) between the concentration of sugars (Chem) andTdr (Biol). Correlations (>0.5) between the sugars and canonical variable(Chem) are drawn as vectors on axes Chem. Canonical correlation coef-ficient (r) is presented. Rhamnose (Rha), xylose (Xyl), mannose (Man),galactose (Gal), glucose (Glc).
order to maximize the correlation between the two canonicalvariables, which were biological (Tdr, Ac-erg) and chemicalvariables (concentration of sugars g−1 DOC) (Figure 6c). The firstcombination (r = 0.84, n = 72) consisted of Tdr and sugars. Themost important sugars (sugars with the best correlations with thecanonical variable, Chem) were Glc, Gal, Man, Xyl and Rha.CCA shows that the larger the concentration of these sugars inthe incubation solution the larger the Tdr. None of the sugars hada negative correlation with Chem. The correlation between secondcanonical variables (Ac-erg and sugars) was weak (not shown).
Discussion
Both birch and spruce litter lost about one-third of their Cduring the 252 days incubation. The mixing of birch and sprucelitters together increased decomposition compared with single-species litters by about 15%. This is in accordance with earlierfindings where mixing deciduous and coniferous litters togetherhas increased the decomposition rate compared with single-specieslitters (Brandtberg & Lundkvist, 2004; Gartner & Gordon, 2004).
The initial concentrations of C, N, hemicelluloses and phenoliccompounds did not differ much between birch and spruce litter.However, the partly decomposed birch litter was characterizedby a greater concentration of hemicelluloses and a smallerconcentration of HMW phenolic compounds than spruce litter(Table 1). Forest soils under birch and spruce seem to havedifferences in the quality of organic matter, the organic layer underthe birch contained less HMW phenolic compounds (Kanervaet al., 2007) and more hemicelluloses than spruce (Merila et al.,2010). Lignin, cellulose and hemicellulose degradation are knownto be closely connected (Filley et al., 2002) and hemicellulosemay act as an easy C source in the degradation of refractorycompounds (Kaiser et al., 2001).
No consistent differences have been found previously betweenconiferous and deciduous tree species in the total fluxes of DOCduring decomposition of litter (Michalzik et al., 2001). In ourstudy the total flux was also almost the same for birch andspruce litter. However, the flux of easily degradable DOM wasgreater for birch than for spruce litter. Birch and spruce also hada different pattern of DOM leaching during decomposition, whichis in agreement with the observation of Kalbitz et al. (2006) forconiferous and deciduous species. In the first phase, when thedegradability of DOM was large, the large initial flux of DOCand phenolic compounds decreased until freezing. In the secondphase, after thawing, the degradability of DOM had decreased.In contrast, the flux of DOC and phenolic compounds showedan increasing trend, and the increase was more pronounced forspruce than birch. However, it was clearly most pronounced forthe mixture of litters that had also decomposed the most. Thelitter mixture also released more than double the amount of DOC,DON, proteins, phenolic compounds and hemicelluloses than birchduring the last leaching occasion, when the flux was calculated inrelation to the amount of C in decomposed litter. Thus the greaterflux of DOM in the second phase seems to indicate the greater
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science

DOM from tree litter 9
decomposition stage of the litter mixture. Don & Kalbitz (2005)and Kalbitz et al. (2006) suggested that the increase in the fluxof DOC in the second phase indicates the degradation of ligninbecause the increase in DOC flux was accompanied by an increasein lignin degradation products and a poorer degradability of DOM.
The degradability of DOM and the flux of DOC, DON andproteins increased after freezing and thawing of litters and this canbe explained by the physical destruction of microbial and plantcells. For phenolic compounds and hemicelluloses the increasein the flux was not so clear. Little is known about the effects offreeze-thaw cycles on litter chemistry. Decomposition of celluloseand mineralization of N increased and decomposition of lignindecreased in litter during the freeze-thaw season (Wu et al.,2010). Freeze-thaw cycles have been observed to decrease theconcentration of sugars but not lignin (Schmitt et al., 2008), aswell as to increase the release of DOC but not DON (Hentschellet al., 2008) in forest soils. The complicated relationships of bioticand abiotic factors connected to freeze-thaw cycles depend onthe properties of organic matter and are not fully understood.In our study the effects of the freeze-thaw cycle are difficult todistinguish from the effects of increasing decomposition stage.
The C:N ratio of litters decreased from about 50 to 35 (Table 1)during the decomposition and thus N was accumulated during thedecomposition in general, which has also been reported earlier(Berg, 1986; Couteaux et al., 1997). Spruce litter had a greaterloss of N than birch, as also observed by Brandtberg & Lundkvist(2004). The loss of N from solid litters during the incubationshould have been at a similar rate to the total flux of N. However,the flux was markedly less than the loss and this gap was larger forspruce than for birch. Filtration removed some solid material andit is possible that sorption of DON and mineral N with particles orprecipitating compounds is different for birch and spruce becauseof the differences in the chemical composition of both solid anddissolved compounds. Also, the immobilization of N in microbialbiomass may have been different between litters. Neither can weexclude the possibility of N losses as gases, although large rates ofdenitrification were unlikely. Birch litter columns do not mimic theforest floor particularly well because underground vegetation andearthworms are absent and this may also explain why birch litterdid not decompose more than spruce litter as found in previousstudies (Prescott et al., 2000; Wardle et al., 2003).
The birch and spruce litters had a different initial distributionof individual sugar units and the distribution did not change muchduring decomposition. Some of the sugars that were lost were notleached as DOM. Glucose and mannose were more abundant inspruce litter. They also correlated strongly with the availabilityof DOM to bacteria according to CCA and thus seemed to beeasily degradable. Birch litter contained more galacturonic acidthat had possibly degraded quickly because it was almost absentin the DOM solutions. Another explanation for the disappearanceof sugars is chemical alteration of sugar molecules leading to SOMstabilization, as suggested by Schmitt et al. (2008). On the whole,the concentrations of several individual and total hemicelluloses inDOM were positively correlated with the availability of DOM to
bacteria (Figure 6b). Therefore we suggest that hemicelluloses inDOM had good degradability that did not differ markedly betweenbirch and spruce despite their different patterns in sugar contents.
Leached DOM includes also microbial products such assugars. The ratio of (galactose + mannose):(arabinose + xylose)has been used successfully as an indicator of sugars of microbialorigin (Rumpel et al., 2010). When the ratio is <0.5 sugars aresuggested to be more of plant origin and when >2 of microbialorigin. Initially the ratio in birch litter was 0.4. However, sprucelitter contained a remarkable amount of mannose, giving a ratio of1.6. The ratio in DOM was always >2 and thus we may explainthe difference in sugars between solid litter and DOM in part bythe greater proportion of microbial sugars in DOM than in thesolid litter, as suggested also by Kaiser et al. (2001).
The ratio of fungal to bacterial growth rate in DOM solutionincreased during the decomposition of litter. Thus it seems thatbacteria played a greater part in decomposition at the initialstages and fungi at the later stages of litter decomposition, assuggested by Moore-Kucera & Dick (2008). The freeze-thaw cycleincreased the growth rate of both bacteria and fungi: thus bothbacteria and fungi seemed to be able to use the substrates released.Contradictory observations, where fungi are more susceptible tosoil frost than bacteria, have been reported by Schmitt et al.(2008). It has also been observed that bacteria benefit from asmall and fungi from a large C:N ratio of the substrate (Rousket al., 2010). We, however, did not have solutions with very smallC:N ratios: the values were mostly 25–60 and may not explainthe fungal:bacterial growth ratio.
The compounds in DOM are primary products leached fromplants or secondary products produced during the dynamic processof litter decomposition. We cannot separate these fractions. DOMcan mineralize, degrade to smaller particles, change to othercompounds (oxidation, reduction, condensation), or bind withother molecules into insoluble, refractory forms. It has beenobserved that recalcitrant DOM provides remarkable C storageafter sorption by soil particles whereas labile DOM promotesmicrobial activities and C cycling (Kalbitz et al., 2007). Thedegradability of DOM leached from decomposing litter differedbetween birch and spruce and between degradation stages. Thusboth tree species and degradation stage contribute to soil C andN storage and cycling.
The total flux of sugars and phenolic compounds did not differmuch between birch and spruce litter. Birch and spruce litters hadprobably about the same amount of C respired because the lossof C and total DOC leached were about the same. In contrast, thetotal amount of easily degradable leached DOM was greater forbirch than for spruce. This supports the earlier results where DOMderived from birch stands was more degradable than DOM derivedfrom spruce stands (Kiikkila et al., 2011). The degradability ofDOM can be one explanation for the larger microbial biomass andactivities related to C and N transformations in the organic layerunder birch than under spruce (Smolander & Kitunen, 2011). Wealso assume that, in the field, birch litter generally decomposesfaster than spruce litter and the annual litterfall of birch may
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science

10 O. Kiikkila et al.
be slightly larger than that of spruce (Saarsalmi et al., 2007;A., Saarsalmi, personal communication), increasing further thedifference in labile DOM between birch and spruce.
We conclude that the greater amount of easily degradableDOM leached from birch than spruce litter may explain theearlier observed greater microbial activity under birch than spruce.Because the degradability of DOM leached from decomposinglitter differed between birch and spruce and between degradationstages we conclude that both tree species and degradation stagecontribute to the storage and cycling of C and N in forest soil.
Acknowledgements
We are grateful to R. Lievonen for collecting the litter samples,P. Kinnunen and A. Rautiainen for laboratory work, to A. Siikaand S. Elomaa for the figures and M. Waller for checking theEnglish. The research was supported by the Academy of Finland.
References
Berg, B. 1986. Nutrient release from litter and humus in coniferous forestsoils - a mini review. Scandinavian Journal of Forest Research, 1,359–369.
Box, J.D. 1981. Investigation of the Folin-Ciocalteu phenol reagent forthe determination of polyphenol substances in natural waters. Water
Research, 17, 511–525.Bradford, M.M. 1976. A rapid and sensitive method for the quantification
of microgram quantities of protein utilizing the principle of protein-dyebinding. Analytical Biochemistry, 72, 248–254.
Brandtberg, P.O. & Lundkvist, H. 2004. Does an admixture of Betulaspecies in Picea abies stands increase organic matter quality and nitro-gen release? Scandinavian Journal of Forest Research, 19, 127–141.
Couteaux, M.M., McTiernan, K.B., Berg, B., Szuberla, D., Dardenne, P.& Botther, P. 1997. Chemical composition and carbon mineralizationpotential of Scots pine needles at different stages of decomposition. Soil
Biology & Biochemistry, 5, 583–595.Don, A. & Kalbitz, K. 2005. Amount and degradability of dissolved
organic carbon from foliar litter at different decomposition stages. Soil
Biology & Biochemistry, 37, 2171–2179.Filley, T.R., Cody, G.D., Goodell, B., Jellison, J., Noser, C. & Ostrof-
sky, A. 2002. Lignin demethylation and polysaccharide decompositionin spruce sapwood degraded by brown rot fungi. Organic Geochemistry,33, 111–124.
Gartner, T.B. & Gordon, Z.G. 2004. Decomposition dynamics in mixedleaf litter. Oikos, 104, 230–246.
Hagedorn, F. & Machwitz, M. 2007. Controls on dissolved organic matterleaching from forest litter grown under elevated atmospheric CO2. Soil
Biology & Biochemistry, 39, 1759–1769.Hentschell, K., Borken, W. & Matzner, E. 2008. Repeated freeze-thaw
events affect leaching losses of nitrogen and dissolved organic matterin a forest soil. Journal of Plant Nutrition & Soil Science, 171, 699–706.
Kaiser, K., Guggenberger, G., Haumaier, L. & Zech, W. 2001. Seasonalvariations in the chemical composition of dissolved organic matter inorganic forest floor layer leachates of old-growth Scots pine (Pinus
sylvestris L.) and European beech (Fagus sylvatica L.) stands innortheastern Bavaria, Germany. Biogeochemistry, 55, 103–143.
Kalbitz, K., Kaiser, K., Bargholz, J. & Dardenne, P. 2006. Lignindegradation controls the production of dissolved organic matter indecomposing foliar litter. European Journal of Soil Science, 57,504–516.
Kalbitz, K., Meyer, A., Yang, R. & Gerstberger, P. 2007. Response ofdissolved organic matter in the forest floor to long-term manipulationof litter and throughfall inputs. Biogeochemistry, 86, 301–318.
Kanerva, S., Kitunen, V., Loponen, J. & Smolander, A. 2007. Phenoliccompounds and terpenes in soil organic horizon layers under silverbirch, Norway spruce and Scots pine. Biology & Fertility of Soils, 44,547–556.
Kiikkila, O., Kitunen, V. & Smolander, A. 2006. Dissolved organic matterfrom surface organic horizons under birch and conifers: degradation inrelation to chemical characteristics. Soil Biology & Biochemistry, 38,737–746.
Kiikkila, O., Kitunen, V. & Smolander, A. 2011. Properties of dissolvedorganic matter derived from silver birch and Norway spruce stands:degradability combined with chemical characteristics. Soil Biology &Biochemistry, 43, 421–430.
Kuiters, A.T. & Sarink, H.M. 1986. Leaching of phenolic compoundsfrom leaf and needle litter of several deciduous and coniferous trees.Soil Biology & Biochemistry, 5, 475–480.
Lindroos, A.J., Derome, J. & Derome, K. 2007. Open area bulk depositionand stand throughfall in Finland during 2001–2004. In: ForestCondition Monitoring in Finland, National Report 2002–2005 (edsP. Merila, T. Kilponen & J. Derome), pp. 81–92. Working Papers ofthe Finnish Forest Research Institute, Vantaa.
Matzner, E. & Borken, W. 2008. Do freeze-thaw events enhance C and Nlosses from soils of different ecosystems? A review. European Journalof Soil Science, 59, 274–284.
Merila, P., Malmivaara-Lamsa, M., Spetz, P., Stark, S., Vierikko, K.,Derome, J. et al. 2010. Soil organic matter quality as a link betweenmicrobial community structure and vegetation composition along asuccessional gradient in a boreal forest. Applied Soil Ecology, 46,259–267.
Michalzik, B., Kalbitz, K., Park, J.H., Solinger, S. & Matzner, E. 2001.Fluxes and concentrations of dissolved organic carbon and nitrogen – asynthesis for temperate forests. Biogeochemistry, 52, 173–205.
Moore-Kucera, J. & Dick, R.P. 2008. Application of 13C-labeled litter androot materials for in situ decomposition studies using phospholipid fattyacids. Soil Biology & Biochemistry, 40, 2485–2493.
Prescott, C.E. 2010. Litter decomposition: what controls it and how canwe alter it to sequester more carbon in forest soils? Biogeochemistry,101, 133–149.
Prescott, C.E., Zabek, L.M., Staley, C.L. & Kabzems, R. 2000. Decom-position of broadleaf and needle litter in forests of British Columbia:influences of litter type, forest type and litter mixtures. Canadian Jour-nal of Forest Research, 30, 1742–1750.
Rousk, J., Brookes, P.C. & Baath, E. 2010. Investigating the mechanismsfor the opposing pH relationships of fungal and bacterial growth in soil.Soil Biology & Biochemistry, 42, 926–934.
Rumpel, C., Eusterhues, K. & Kogel-Knabner, I. 2010. Non-cellulosicneutral sugar contribution to mineral associated organic matter intop- and subsoil horizons of two acid forest soils. Soil Biology &Biochemistry, 42, 379–382.
Saarsalmi, A., Starr, M., Hokkanen, T., Ukonmaanaho, L., Kukkola, M.,Nojd, P. et al. 2007. Predicting annual canopy litterfall productionfor Norway spruce (Picea abies (L.) Karst.) stands. Forest Ecology &Management, 242, 578–586.
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science

DOM from tree litter 11
Sariyildiz, T. & Anderson, J.M. 2003. Interactions between litter quality,decomposition and soil fertility: a laboratory study. Soil Biology &
Biochemistry, 35, 391–399.Schmitt, A., Glaser, B., Borken, W. & Matzner, E. 2008. Repeated freeze-
thaw cycles changed organic matter quality in a temperate forest soil.Journal of Plant Nutrition & Soil Science, 171, 707–718.
Smolander, A. & Kitunen, V. 2011. Comparison of tree species effectson microbial C and N transformations and dissolved organic matterproperties in boreal forest floors. Applied Soil Ecology, 49, 224–233.
Smolander, A., Loponen, J., Suominen, K. & Kitunen, V. 2005. Organicmatter characteristics and C and N transformations in the humus layerunder two tree species, Betula pendula and Picea abies. Soil Biology &
Biochemistry, 37, 1309–1318.Snajdr, J., Cajthaml, T., Valaskova, V., Merhautova, V., Petrankova, M.,
Spetz, P. et al. 2011. Transformation of Quercus petraea litter:successive changes in litter chemistry are reflected in differentialenzyme activity and changes in the microbial community composition.FEMS Microbiology Ecology, 75, 291–303.
Sundberg, A., Sundberg, K., Lilland, A. & Holmbom, B. 1996. Determi-nation of hemicelluloses and pectins in wood and pulp fibres by acid
methanolysis and gas chromatography. Nordic Pulp & Paper Research
Journal, 4, 216–226.Thevenot, M., Dignac, M.F. & Rumpel, C. 2010. Fate of lignins in soils:
a review. Soil Biology & Biochemistry, 42, 1200–1211.Valachovic, Y.S., Caldwell, B.A., Cromack, K. Jr & Griffiths, R.P. 2004.
Leaf litter chemistry controls on decomposition of Pacific Northwesttrees and woody shrubs. Canadian Journal of Forest Research, 34,2131–2147.
Wardle, D.A., Nilsson, M.C., Zackrisson, O. & Gallet, C. 2003. Determi-nants of litter mixing effects in a Swedish boreal forest. Soil Biology &
Biochemistry, 35, 827–835.Willfor, S., Sundberg, A., Hemming, J. & Holbom, B. 2005. Polysaccha-
rides in some industrially important softwood species. Wood Science &
Technology, 39, 245–258.Wu, F., Yang, W., Zhang, J. & Deng, R. 2010. Litter decomposition in
two subalpine forests during the freeze-thaw season. Acta Oecologica,36, 135–140.
Zhang, D., Hui, D., Luo, Y. & Zhou, G. 2008. Rates of litter decompo-sition in terrestrial ecosystems: global patterns and controlling factors.Journal of Plant Ecology, 1, 85–93.
© 2012 The AuthorsJournal compilation © 2012 British Society of Soil Science, European Journal of Soil Science