Embed Size (px)
Citation preview

Characterization of a positive transcriptional control element in the Cd4 gene in mice using transient transfection of double positive thymoma cells
Darina Spasova and Sophia D. SarafovaBiology Department, Davidson College, Davidson, NC 28036
Abstract
Methods
Acknowledgments
References
A B
C
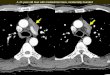
Figure 1:
AKR1G1 double positive thymoma cells successfully transfected with 4e/p-eGFP-sil fluoresce green (A). Transfection efficiency using the DEAE dextran method is low, regardless of the amount of DNA (B) 4g/mL, (C) 6 g/mL, or the time allowed for recovery and eGFP expression after transfection - 24h (B) vs. 48h (C)
Higher level of eGFP expression was observed at the later time points.
Figure 2: Flow cytometry data from a DEAE dextran transient transfection of AKR1G1 and RLM11 double positive thymoma cell lines, using 4e/p-eGFP-sil as the experimental transfection plasmid and TIPflag(CD8) as the transfection control plasmid (A, B, C, D, E, F). Lipofection of AKR1G1 double positive thymoma cell line using 4e/p-eGFP-sil as experimental transfection plasmid and no plasmid as experimental control (G). A flow cytometry assay was performed on the double transfection using the eGFP fluorescence, which the transfection plasmid was supposed to introduce into the cells (A, D). The single transfections were not stained (B, E). Cells were stained with CD8FITC when transfection efficiency was to be tested for the control plasmid - TIPflag(CD8) (C, F). Even though they were visible under a fluorescent microscope, AKR1G1 and RLM11 transfected cells with 4e/p-eGFP-sil were not detectable by FACS (B, E). CD8 transfection is readily visible by FACS in the AKR1G1 cell line (C) but not in the RLM11 cells (F). Blue line represents no plasmid transfection. Red line represents plasmid transfection (stained with CD8-FITC in C, F). Black line represents no plasmid transfection stained with CD8-FITC (stained with CD8-FITC in C, F).
Transient Transfection using DEAE dextran
Transfection plasmid
RLM-11 Double Positive Thymoma
AKR1G1 Double Positive Thymoma
DEAE Dextran
Transfection plasmid
Incubate in 7.5% CO2
at 37°C
24, 48, 72 hour time points
Fluorescent Microscope Examination and FACS analysis to check for transfection success and
efficiency
Transient Transfection using Lipofectamine
Fluorescent Microscope Examination and FACS analysis to check for transfection success and efficiencyAKR1G1 Double
Positive Thymoma
LipofectamineTransfection plasmid
Transfect cells with mixture
Incubate in 7.5%
CO 2 at 3
7°C
DEAE dextran and lipofection are not suitable for transfection of DNA into AKR1G1 DP thymoma cells
Low efficiency of DEAE dextran transfection of DNA into AKR1G1 DP thymoma cells
4e/p-eGFP-silDEAE dextran transfection
4e/p-eGFP-sil and TIPflag(CD8)DEAE dextran transfection
TIPflag(CD8)DEAE dextran transfection
AKR1G1
RLM11
eGFP CD8
4e/p-eGFP-sil lipofection
eGFP
4e/p-eGFP-silDEAE dextran transfection
4e/p-eGFP-sil and TIPflag(CD8)DEAE dextran transfection
TIPflag(CD8)DEAE dextran transfection
AKR1G1
RLM11
eGFP CD8
4e/p-eGFP-sil lipofection
eGFP
Conclusions and Future Directions DEAE dextran and lipofection are both not highly efficient techniques for transfecting DNA into AKR1G1 double positive thymoma cells. More protocols need to be examined in order to achieve high efficiency of transfection. One possibility is using an electroporator. When a high efficency protocol is selected, the newly made transfection plasmids will be able to demonstrate whether the transcriptional control element (Pr2) is an enhancer or a promoter.
pSK.Cd4Pr1
Pr1
Am
pr
Making Transfection Plasmids
XbaI and EcoRV double digest, forming 1 blunt, 1 sticky end
XbaIXhoI
4e/p-eGFP-sil
1. XhoI digest
2. Klenow
Pr1
4e/p-eGFP-sil
Pr1XbaI
XbaI digest
pSK
pSK
4e/p-eGFP-sil
Pr1 Quick
Ligat
ion
Amp
rAm
pr
pSK.Cd4Pr1 pSK.Cd4Pr1eGFPPr2
HindIII digest, CIP, CIP Inactivate Ligate
MigRI
eGFP
Am
pr
HindIII digest to excise eGFP
eGFP
pSK.Cd4Pr1
Pr1
Am
pr
pSK.Cd4Pr1eGFP
Pr1
Am
pr
eGFP
Ligate
pSK.Cd4Pr1eGFPPr2
Pr1
Am
pr
Pr2
eGF
P
pNEB206A.Cd4Pr2
Am
pr
Ehlers, M., K. Laule-Kilian, M. Petter, C. Aldrian, B. Grueter, A. Würch, N. Yoshida, T. Watanabe, M. Satake, and V. Steimle. 2003. Morpholino antisense oligonucleotide-mediated gene knockdown during thymocyte development reveals role of Runx3 transcription factor in Cd4 silencing during development of Cd4-CD8+ thymocytes. J. Immunol. 171: 3594-3604.
McCready, P., R. Hansen, S. Burke, and J. Sands. 1997. Multiple negative and positive cis-acting elements control the expression of the murine Cd4 gene. Biochim. Biophys. Acta. 1351:181-191
Sawada, S., J.D. Scarborough, N. Killeen, Littman D. 1994. A lineage-specific transcriptional silencer regulates Cd4 gene expression during T lymphocyte development. Cell. 77:917-929.
Wildt, K., G. Sun, B. Grueter, M. Fischer, M. Zamisch, M. Ehlers, and R. Bosselut. 2007. The transcription factor Zbtb7b promotes Cd4 expression by antagonizing Runx-mediated activation of the Cd4 silencer. J. Immunol. 179: 4405-4414.
Regulation of Cd4 gene expression cannot be fully explained by the transcriptional control elements described to date. Previous research in the lab revealed that there are two different transcripts present in developing and in mature T cells, which, we speculated, showed the presence of another positive control element within intron 1 of the Cd4 gene. In this study, we describe an in vitro approach to determine if this positive regulatory element is a promoter or an enhancer. We cloned a piece of intron 1, immediately upstream of exon 2, and wished to examine its promoter and enhancer capabilities in a transient transfection assay. For that purpose, we started constructing plasmids in which we placed the DNA of interest in front of a reporter gene (eGFP) in the presence or absence of other known Cd4 transcriptional control elements. Furthermore, we attempted to establish a high efficiency transient transfection protocol, such that successfully transfected cells can be visualized by flow cytometry. Neither of the two different protocols gave us high efficiency transfection. More protocols will be tested in the future, while the reporter plasmid construction is finished.
We would like to thank Chris Healey for providing us with technical support, the NIH – for all the reagents they donated to our lab, Karmella Haynes for the insight and reagents for lipofection, and Dr. Yuan Zhuang at Duke University Medical Center for providing us with transfection and control plasmids.
A B C G
D E F
Pr2
Quick
Ligatio
n
Sil
Am
pr
eGF
P
4e/p-eGFP-sil