Embed Size (px)
Citation preview

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 1993, p. 962-9690066-4804/93/050962-08$02.00/0Copyright X 1993, American Society for Microbiology
Characterization of a Novel Extended-Spectrum 1-Lactamasefrom Pseudomonas aeruginosa
PATRICE NORDMANN,12t * ESTHEL RONCO,1 THIERRY NAAS,2 CATHERINE DUPORT,3YVON MICHEL-BRIAND,3 AND ROGER LABIA4
Laboratoire de Microbiologie, H6pital Raymond Poincare, Faculte6 de Medecine Paris-Ouest, 92380 Garches,Laboratoire de Bacteriologie, Faculte6 de Me6decine, 25030 Besanqon,3 and Museum d'Histoire Naturelle
URA 401 Centre National de la Recherche Scientifique, 75231 Paris 05, 4 France, and AbteilungMikrobiologie, Biozentrum der Universitat Basel, 4056 Basel, Switzerland2
Received 17 November 1992/Accepted 3 March 1993
A clinical isolate ofPseudomonas aeruginosa RNL-1 showed resistance to extended-spectrum cephalosporinswhich was inhibited by clavulanic acid. Although this strain contained three plasmids ca. 80, 20, and 4 kb long,the resistance could not be transferred by mating-out assays with P. aeruginosa or Escherichia coli. Cloning ofa 2.1-kb Sau3A fragment from P. aeruginosa RNL-1 into plasmid pACYC184 produced pPZ1, a recombinantplasmid that encodes a 1-lactamase. This f-lactamase (PER-1) had a relative molecular mass of 29 kDa anda pl of 5.4 and was biosynthesized by P. aeruginosa RNL-1 along with a likely cephalosporinase with a pl of8.7. PER-1 showed a broad substrate profile by hydrolyzing benzylpenicillin, amoxicillin, ticarcillin cepha-lothin, cefoperazone, cefuroxime, HR 221, ceftriaxone, ceftazidime, and (moderately) aztreonam but notoxacillin, imipenem, or cephamycins. Vmax values for extended-spectrum cephalosporins were uncommonlyhigh, and the affinity of the enzyme for most compounds was relatively low (i.e., high Km)* PER-1 activity wasinhibited by clavulanic acid, sulbactam, imipenem, and cephamycins but not by EDTA. A 1.1-kb SnaBIfragment from pPZ1 failed to hybridize with plasmids that encode TEM-, SHV-, OXA-, or CARB/PSE-type13-lactamase or with the ampC gene of P. aeruginosa. However, the same probe appeared to hybridize withchromosomal but not plasmid DNA from P. aeruginosa RNL-1. This study reports the properties of a novelextended-spectrum 13-lactamase in P. aeruginosa which may not be derived by point mutations from previouslyknown enzymes of this species.
More than 50 biochemically distinct P-lactamases respon-sible for resistance to ,-lactams have been reported ingram-negative bacteria. The resistance of broad-spectrumcephalosporins to these ,B-lactamases was a widely acceptedconcept in the beginning of the 1980s. However, overpro-duction of chromosomally mediated cephalosporinases hasbeen described as responsible for failure of treatment ofgram-negative bacterial infections with extended-spectrumcephalosporins (39). Since 1983, plasmid-mediated extend-ed-spectrum 3-lactamases have been reported, primarily inKiebsiella pneumoniae and then in numerous Enterobacte-raceae species (16, 34). These enzymes hydrolyze extend-ed-spectrum cephalosporins and aztreonam to various ex-tents but usually neither cephamycins (cefoxitin andmoxalactam) nor carbapenems (imipenem and meropenem).A common feature of these enzymes is inhibition of theiractivity by clavulanic acid. These enzymes are Ambler classA 1-lactamases, members of the TEM or SHV series thatdiffer by a few point mutations in their structural genes (16,34).Such enzymes from Pseudomonas aeruginosa have not
been reported. In this species, induction or derepression ofchromosomal cephalosporinases may lead to resistance toextended-spectrum cephalosporins but their activity is notinhibited by clavulanic acid (22). The other P-lactamasescommonly found in P. aeruginosa do not confer clinical
* Corresponding author.t Present address: Laboratoire de Microbiologie, H6pital Ray-
mond Poincare, 104 Bd Raymond Poincare, 92380 Garches, France.
resistance to extended-spectrum cephalosporins. In thisspecies, TEM-1 and TEM-2 1-lactamases confer additionalresistance to ureidopenicillins (26). The OXA-type (oxacil-lin-hydrolyzing) enzymes possess high-level hydrolytic ac-tivity against cloxacillin, oxacillin, and methicillin (9, 10).Their activities are inhibited by clavulanic acid but to alesser extent than TEM- or SHV-derivative P-lactamases.This group includes OXA-1 to OXA-7, as well as PSE-2, andpossesses hydrolytic activities similar to and protein se-quences related to those of oxacillin-hydrolyzing enzymes.CARB- and PSE-type j-lactamases (with the exception ofPSE-2), also called carbenicillin-hydrolyzing enzymes, arefound primarily in P. aeruginosa. They tend to hydrolyzecephalosporins much more slowly than penicillin and gener-ally have slow rates of cloxacillin hydrolysis (9, 10). Boththe oxacillin-hydrolyzing enzymes and carbenicillin-hydro-lyzing enzymes described so far do not confer clinicalresistance to extended-spectrum cephalosporins on P. aeru-ginosa.
In this report, we describe a novel, likely chromosomallymediated, extended-spectrum ,B-lactamase from a clinicalisolate of P. aeruginosa. The purpose of this work was to (i)determine the enzymatic properties of the 1-lactamase, (ii)characterize its genetic determinant, and (iii) search forDNA homology with previously described TEM- and SHV-derived 1-lactamases, as well as with oxacillin-hydrolyzingand carbenicillin-hydrolyzing enzymes. Special interest inthis P-lactamase comes from its high level of hydrolyticactivity against extended-spectrum cephalosporins and inhi-bition of its activity by clavulanic acid, sulbactam, imi-penem, and cephamycins.
962
Vol. 37, No. 5
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

EXTENDED-SPECTRUM j-LACTAMASE FROM P. AERUGINOSA 963
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Relevant genotype Source oror phenotype reference
StrainsEscherichia coliJM109
Pseudomonas aeru-
ginosa RNL-1P. aeruginosa PAO
2635P. aeruginosa NTCT
8203P. aeruginosa PouP. aeruginosa CilP. aeruginosa DaXanthomonas malto-
philia 6077T
Plasmids
pACYC184pHUC37
pKl9pPZl
pRAZl
pUC19
p124p453RlllR151Rmsl49RP4RGN238R46R57b
end4l hsdR17gyrA96A(lac proA) recABIreL4 supE44 thi F'
(laclq lacZAM15proAB+ traD36)
PER-1 P-lactamase
FP- trp-54 nf-3 fon-101
AmpC from P. aerugi-nosa
blaPSE-l
blaCARB-3blaPSE4
blaL-l blaL-2
cat tet
Recombinant plasmidcontaining 450-bpPstI-NotI internal
fragment of blaSHV-3Kanamycin resistance
2.1-kb Sau3A fragmentfrom P. aeruginosaRNL-1 cloned into
pACYC1841.1-kb SnaBI fragmentfrom pPZ1 cloned into
SmaI site from di-
gested pK19Recombinant plasmid
containing a 298-bpHincII-PstI internal
fragment for blaTEmlblaSHV-2
blasHv-lblaTEM-l
blaPSE-2blaPSE-3blaTEM-2
blaOXA-2
blaOXA-3a IPSC, Institut Pasteur strain collection.
MATERIALS AND METHODS
Bacterial strains and plasmids. The bacterial strains andplasmids used in this work are listed in Table 1. P. aerugi-nosa RNL-1 was isolated from the urinary tract of a hospi-talized patient at Raymond Poincare Hospital, Garches,France, in 1991. Antimicrobial regimens before admissionwere not documented, and the patient did not receive anyantibiotic treatment before isolation of the strain at thehospital. The strain was identified with API-20 NE (Bi-omerieux, La Balme-les-Grottes, France) and belonged toserogroup 0:2 (antisera from Diagnotics Pasteur, Mames-La-Coquette, France). On the basis of agar disc diffusionassay results, the isolate was resistant to extended-spectrumcephalosporins. A marked synergistic effect between clavu-lanic acid and extended-spectrum cephalosporins was ob-served that suggested the presence of an uncommon P-lac-
tam resistance mechanism in P. aeruginosa. Analysis ofresistance to ,B-lactams is described below. P. aeruginosaRNL-1 was also resistant to amikacin, chloramphenicol,gentamicin, kanamycin, neomycin, netilmycin, sisomycin,streptomycin, sulfonamides, and tobramycin.
Antimicrobial agents and MIC determinations. The antimi-crobial agents used in this study were obtained from stan-dard laboratory powders and were used immediately aftersolubilization. The agents and their sources were as follows:ampicillin, Bristol; amoxicillin, clavulanic acid, cloxacillin,and ticarcillin, Smith Kline French-Beecham; aztreonam,Squibb; cefamandole and moxalactam, Eli Lilly; cefpirome,ceftriaxone, and HR 221, Hoffmann-La Roche; cefuroxime,ceftazidime, and cephalothin, Glaxo; cefoxitin and imi-penem, Merck Sharp & Dohme; cefotaxime, Hoechst-Rous-sel; meropenem, ICI; benzylpenicillin, Specia; sulbactam,Pfizer; piperacillin, Lederle; rifampin, Sigma.MICs were determined by an agar dilution technique on
Mueller-Hinton agar (Diagnostics Pasteur) with a Steersmultiple inoculator and an inoculum of 104 CFU per spot. Allplates were incubated at 37°C for 18 h. MICs of 13-lactamswere determined alone or in combination with 2 ,ug ofclavulanic acid per ml or 4 ,ug of sulbactam per ml.
Plasmid content and conjugation assays. Lysates of P.aeruginosa RNL-1 were prepared by the Kieser method(17). Plasmid DNA was detected by electrophoresis in a0.7% agarose gel (95 V, 4 h) (27). The molecular sizes ofplasmid DNAs were estimated by comparison with plasmidswhose sizes are known (23, 43). Direct transfer of resistanceinto P. aeruginosa PAO 2635 and into in vitro-obtainedrifampin-resistant Escherichia coli JM109 was attempted byliquid and solid mating-out assays at 30 and 37°C as previ-ously described (36). Transconjugant selection was per-formed on Luria agar plates (Diagnostic Pasteur) containingrifampin (100 p,g/ml) and either ceftazidime (10 ,ug/ml) orticarcillin (250 ,ug/ml).
Cloning experiments and analysis of recombinant plasmids.Chromosomal DNA of P. aeruginosa RNL-1 was preparedas previously described (14). Fragments from partiallySau3A-digested genomic P. aeruginosa DNA (Boehringer,Mannheim, Germany) were ligated to the BamHI site ofpACYC184 (11). BamHI-digested pACYC184 was dephos-phorylated and electrophoresed in 0.8% low-melting-pointagarose, and then the band was excised and purified withGenclean II (Bio 101, Inc., La Jolla, Calif.). Ligation wasperformed with a 2:1 vector-insert ratio at a final concentra-tion of ca. 200 ng of DNA in a ligation mixture containing 1U of T4 DNA ligase (Boehringer) at 15°C for 18 h. Recom-binant plasmids were transformed by electroporation into E.coli JM109 (Bio-Rad, Richmond, Calif.). Antibiotic-resistantcolonies were selected on Luria agar plates supplementedwith 1 or 5 ,ug of ceftazidime per ml, 50 or 100 ,ug ofampicillin per ml, 100 or 200 ,ug of ticarcillin per ml.Recombinant DNA plasmids were obtained from 500-ml
Luria broth cultures grown overnight at 37°C and isolated inaccordance with the Quiagen protocol (Diagen). The reac-tion conditions used for restriction enzyme digestions werethose suggested by the supplier (Boehringer). RestrictedDNA fragments were separated on 0.8% agarose in 90 mMTris-90 mM boric acid-3 mM EDTA (pH 8.0). The gels werestained in a solution of ethidium bromide for 15 min andwashed for 15 min in distilled water. The standard molecularweight marker was a 1-kb DNA ladder (BRL, Basel, Swit-zerland). Estimation of fragment sizes and construction ofphysical maps were done after restriction enzyme doubledigestion.
VOL. 37, 1993
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIMICROB. AGENTS CHEMOTHER.
13-Lactamase preparation. Cultures were grown overnightat 37°C in 100 ml of Trypticase soy broth (DiagnosticsPasteur). Bacterial suspensions were disrupted by sonifica-tion (4 x 20 s at 20 Hz) and centrifuged (30 min, 20,000 x g,4°C). The supernatant contained the crude enzyme extracts.
Isoelectric focusing. Crude ,-lactamase extracts were sub-mitted to analytical isoelectric focusing on a pH 3.5 to 9.5ampholin polyacrylamide gel (Pharmacia, Uppsala, Sweden)for 36 h at 10 W of constant power on a flatbed apparatus(FBE-3000; Pharmacia). The 13-lactamases were visualizedwith an overlay of agar-iodine starch gel containing penicillin(0.01% [wt/vol]) in 0.1 M phosphate buffer (pH 7.0) (2).
13-Lactamase activity and determination of kinetic con-stants. The P-lactamase activity of crude extracts of E. coliJM109 harboring recombinant plasmid pPZ1 (see Results)and the kinetic constants of preparations were determinedby the one-line computerized microacidimetric method atpH 7.0 and 37°C as previously described (18). As assessed byisoelectric focusing, these crude extracts contained only asingle f3-lactamase activity. The Km was expressed in micro-molar concentrations, and Vma, was expressed relative tothat of benzylpenicillin (Vm. = 100). In the case of sub-strates with low or undetectable Vm. values, enzyme-substrate affinity was measured as Ki (inhibition constant)rather than Km with benzylpenicillin as the substrate.
Inhibition of 13-lactamase activity. Various concentrationsof clavulanic acid, sulbactam, imipenem, and cephamycinswere preincubated with the enzymes for 10 min at 37°Cbefore testing of the rate of benzylpenicillin hydrolysis.Inhibition by cloxacillin was determined from the rate ofbenzylpenicillin hydrolysis in the presence up to 0.1 mMinhibitor. Ki values were consequently determined. Inhibi-tion by EDTA (final concentration, 1 mM) (Sigma) or NaCl(final concentration, 100 mM) was tested by preincubationwith the enzyme for 10 min at 37°C before testing of the ratesof benzylpenicillin and ceftazidime hydrolysis. The rate ofhydrolysis was also studied when the enzyme was preincu-bated with up to 10 mM ZnCl2.
Determination of relative molecular mass. The relativemolecular mass of the 3-lactamase obtained from E. coliJM109 harboring recombinant plasmid pRAZ1 (see Results)was estimated by sodium dodecyl sulfate (SDS)-polyacryl-amide gel electrophoresis (19). Crude extracts and markerproteins (Pharmacia, Uppsala, Sweden) were treated with1% SDS-3% P-mercaptoethanol at 100°C for 2 min and thensubjected to electrophoresis in a 12.5% gel (20 mA, 5 h, roomtemperature). Renaturation of P-lactamase activity afterdenaturing electrophoresis was performed as previouslydescribed (25).
Hybridization. DNA-DNA hybridizations were performedby the method of Southern (40). DNA was extracted from P.aenrginosa RNL-1, P. aeruginosa NTCT 8203, X. malto-philia 6077T (blaL 1 blaL 2), P. aeruginosa Pou (blaPSEl), P.aeruginosa Da (blaPSE4),P. aeruginosa Cil (blaCARB-3), P.aeruginosa harboring plasmid Rmsl49 (blaPSE3) or R151(blaPSE2), E. coli harboring plasmid RGN238 (blaoxA1),and Salmonella typhimurium harboring plasmids R46(blaoxA2) and R57b (blaOXA-3). Approximately 10 ,ug ofeach of these DNAs was digested by SnaBI and run on a0.7% agarose gel along with the following DNA fragments:the 298-bp HincII-PstI internal fragment of the blaTEMl1structural gene from pUC19 (45), the 435-bp NotI-PstIinternal fragment of the blaSHV-3 structural gene frompHUC37 (30), and the 1.1-kb SnaBI fragment from pPZ1 (asa positive control). DNAs were transferred onto and immo-bilized on a Biodyne A transfer membrane (PALL, Ports-
pPZl
E Hc HcBE HcB H
I I I LI LI L1 I I I*
I I IPv Sc PVSc
E
IPvPv
1 kbFIG. 1. Restriction endonuclease map of recombinant plasmid
pPZ1, which codes for the PER-1 ,-lactamase. The thin linesindicate vector pACYC184, and the thick line represents the clonedinsert from P. aeruginosa RNL-1. Restriction sites: B, BamHI; E,EcoRI; H, HindlIl; Hc, HincII; P, PstI; Pv, PvuI; S, SnaBI; Sc,Scal. The vectorBamHI site marked with an asterisk was destroyedafter cloning of the Sau3A insert from P. aeruginosa RNL-1.
mouth, England). The probe consisted of the 1.1-kb SnaBIfragment from recombinant plasmid pPZ1. The DNA probewas labeled with [32P]dATP with a random primer DNAlabeling kit (Bio-Rad). Hybridization reactions were per-formed under high- and low-stringency conditions (24).
RESULTS
Cloning of the extended-spectrum 3-lactamase gene. TotalDNA from P. aeruginosa RNL-1 was partially digested withrestriction endonuclease Sau3A and ligated to BamHI-di-gested plasmid pACYC184. The ligated-DNA mixture wastransformed into E. coli JM109 with selection for ampicillin-,ticarcillin-, or ceftazidime-resistant colonies. Despite 12attempts, only three colonies were obtained on ampicillin (50,ug/ml)-containing plates. These colonies were grown over-night, and their recombinant plasmids were extracted. Thethree recombinant plasmids contained inserts of the samesize, i.e., 2.1 kb. One plasmid, pPZ1, was retained fordetailed restriction map analysis (Fig. 1). Subcloning ofvarious restriction digests led to isolation of a 1.1-kb SnaBIfragment from pPZ1, which was inserted into SmaI-digestedmulticopy vector pK19 (pRAZ1), which expressed the samelevel of ,B-lactam resistance as pPZ1. It was used to deter-mine the relative molecular mass of the cloned P-lactamase.
Antibiotic susceptibility. MICs of ,B-lactams showed thehigh resistance of P. aeruginosa RNL-1 to ticarcillin andextended-spectrum cephalosporins (Table 2). Susceptibilityof P. aeruginosa RNL-1 to these antibiotics was partiallyrestored with clavulanic acid. MICs of P-lactams for E. coliJM109 harboring recombinant plasmid pPZ1 confirmed theextended-spectrum cephalosporin resistance conferred bythe cloned 3-lactamase gene. In this strain, MICs of aztreo-nam and ceftazidime were markedly increased comparedwith those of cefotaxime and ceftriaxone. The MIC ofmoxalactam was slightly increased, while that of cefoxitinwas not. All of the 13-lactams tested, except imipenem andcefoxitin, had decreased MICs in the presence of clavulanicacid or sulbactam, and these were more obvious in E. coliharboring pPZ1 than in the original strain, P. aeruginosa
I
964 NORDMANN ET AL.
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
EXTENDED-SPECTRUM P-LACTAMASE FROM P. AERUGINOSA 965
TABLE 2. MICs of 3-lactams for P. aenrginosa RNL-1,E. coli JM109 harboring recombinant plasmid pPZ1,
and reference strain E. coli JM109
MIC (,ug/ml) for:Antibiotic(s) P. aeruginosa E. coli
RNL-1 JM1O(pPZlr E. coi JM109
Amoxicillin >512 >512 <2Amoxicillin-Clab >512 2 -cAmoxicillin-Sued >512 32Ticarcillin 512 >512 1Ticarcillin-Cla 256 2Ticarcillin-Sul 256 32Piperacillin 32 8 1Piperacillin-Cla 32 1Piperacillin-Sul 32 1Cephalothin >512 128 8Cephalothin-Cla >512 8Cephalothin-Sul >512 64Cefamandole >512 32 0.25Cefamandole-Cla >512 0.25Cefamandole-Sul >512 2Moxalactam 16 0.50 0.12Moxalactam-Cla 16 0.12Moxalactam-Sul 16 0.25Cefoxitin >512 8 8Cefoxitin-Cla >512 8Cefoxitin-Sul >512 8Ceftazidime 128 256 0.12Ceftazidime-Cla 4 0.12Ceftazidime-Sul 64 64Cefotaxime 64 4 0.06Cefotaxime-Cla 16 0.06Cefotaxime-Sul 32 0.25Ceftriaxone 128 4 <0.03Ceftriaxone-Cla 32 <0.03Ceftriaxone-Sul 64 0.12Imipenem 0.5 <0.03 <0.03Imipenem-Cla 0.5 <0.03Imipenem-Sul 0.5 <0.03Aztreonam 256 128 0.06Aztreonam-Cla 16 0.06Aztreonam-Sul 64 16
a E. coli JM109 harboring recombinant plasmid pPZ1 produced the PER-113-lactamase.b Cla, clavulanic acid at a fixed concentration of 2 pg/ml. The clavulanic
acid MIC for E. coli JM109 was 16 ,ug/ml.c, not determined.
d Sul, sulbactam at a fixed concentration of 4 pg/ml. The sulbactam MIC forE. coli JM109 was 64 pg/ml.
RNL-1. Both P. aeruginosa RNL-1 and E. coli harboringpPZ1 remained fully susceptible to imipenem.
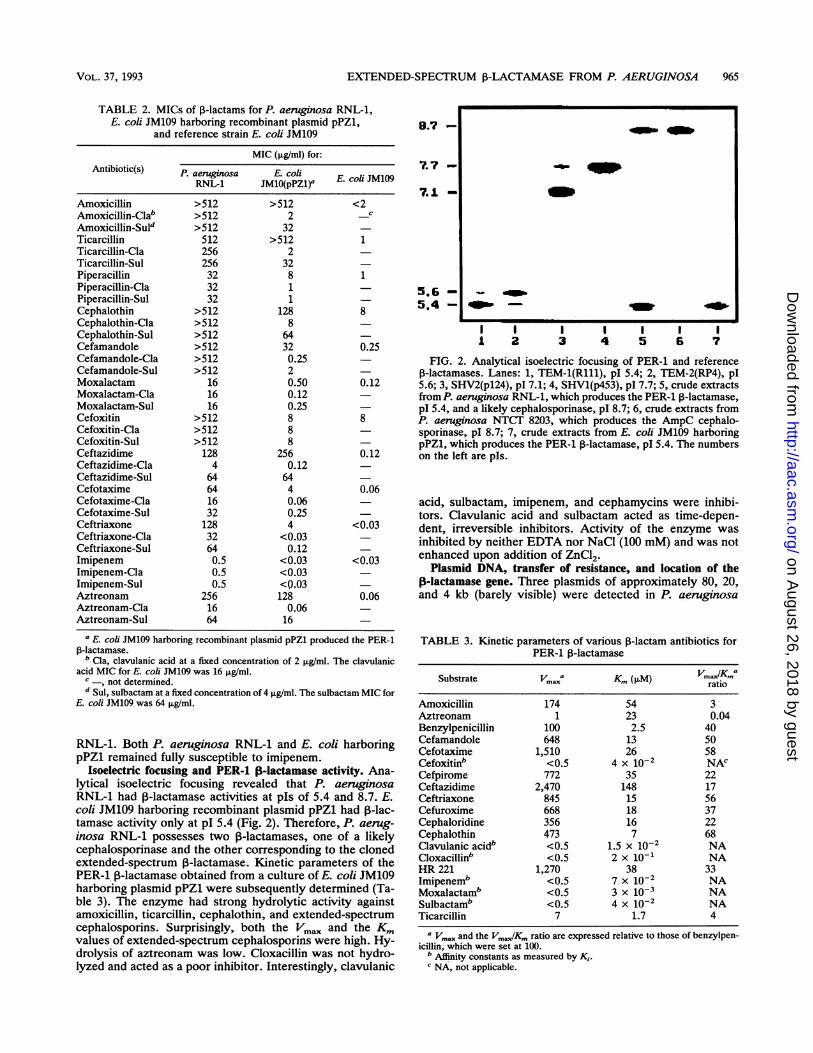
Isoelectric focusing and PER-1 I8-lactamase activity. Ana-lytical isoelectric focusing revealed that P. aeruginosaRNL-1 had ,B-lactamase activities at pIs of 5.4 and 8.7. E.coli JM109 harboring recombinant plasmid pPZ1 had P-lac-tamase activity only at pI 5.4 (Fig. 2). Therefore, P. aerug-inosa RNL-1 possesses two 3-lactamases, one of a likelycephalosporinase and the other corresponding to the clonedextended-spectrum 3-lactamase. Kinetic parameters of thePER-1 13-lactamase obtained from a culture of E. coli JM109harboring plasmid pPZ1 were subsequently determined (Ta-ble 3). The enzyme had strong hydrolytic activity againstamoxicillin, ticarcillin, cephalothin, and extended-spectrumcephalosporins. Surprisingly, both the Vm. and the Kmvalues of extended-spectrum cephalosporins were high. Hy-drolysis of aztreonam was low. Cloxacillin was not hydro-lyzed and acted as a poor inhibitor. Interestingly, clavulanic
e.w?-
7.1 -
5.6 -5.4 -
I I
1 aI I I I
3 4 5 7
FIG. 2. Analytical isoelectric focusing of PER-1 and referenceP-lactamases. Lanes: 1, TEM-1(R111), pI 5.4; 2, TEM-2(RP4), pl5.6; 3, SHV2(p124), pl 7.1; 4, SHV1(p453), pI 7.7; 5, crude extractsfrom P. aeruginosa RNL-1, which produces the PER-1 P-lactamase,pI 5.4, and a likely cephalosporinase, pI 8.7; 6, crude extracts fromP. aeruginosa NTCT 8203, which produces the AmpC cephalo-sporinase, pI 8.7; 7, crude extracts from E. coli JM109 harboringpPZ1, which produces the PER-1 1-lactamase, pI 5.4. The numberson the left are pIs.
acid, sulbactam, imipenem, and cephamycins were inhibi-tors. Clavulanic acid and sulbactam acted as time-depen-dent, irreversible inhibitors. Activity of the enzyme wasinhibited by neither EDTA nor NaCl (100 mM) and was notenhanced upon addition of ZnCl2.
Plasmid DNA, transfer of resistance, and location of the13-lactamase gene. Three plasmids of approximately 80, 20,and 4 kb (barely visible) were detected in P. aeruginosa
TABLE 3. Kinetic parameters of various 3-lactam antibiotics forPER-1 ,B-lactamase
Substrate Vma Km (pM) ratio
Amoxicillin 174 54 3Aztreonam 1 23 0.04Benzylpenicillin 100 2.5 40Cefamandole 648 13 50Cefotaxime 1,510 26 58Cefoxitinb <0.5 4 x 10-2 NACCefpirome 772 35 22Ceftazidime 2,470 148 17Ceftriaxone 845 15 56Cefuroxime 668 18 37Cephaloridine 356 16 22Cephalothin 473 7 68Clavulanic acid' <0.5 1.5 x 10-2 NACloxacillinb <0.5 2 x 10-1 NAHR 221 1,270 38 33Imipenemb <0.5 7 x 10-2 NAMoxalactamb <0.5 3 x 10-3 NASulbactamb <0.5 4 x 10-2 NATicarcillin 7 1.7 4
a Vin, and the Vin,,x/Km ratio are expressed relative to those of benzylpen-icillin, which were set at 100.
b Affinity constants as measured by Ki.c NA, not applicable.
d -Wqo i
VOL. 37, 1993
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
ANTIMICROB. AGENTS CHEMOTHER.
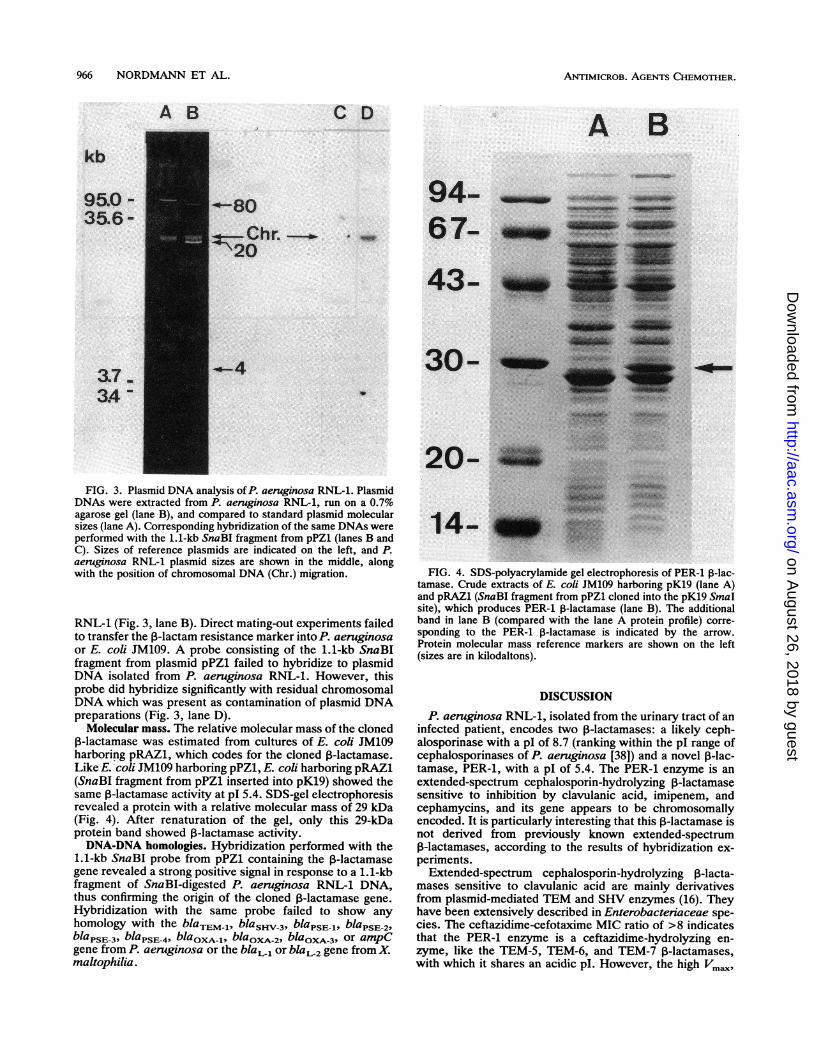
FIG. 3. Plasmid DNA analysis of P. aeruginosa RNL-1. PlasmidDNAs were extracted from P. aeruginosa RNL-1, run on a 0.7%agarose gel (lane B), and compared to standard plasmid molecularsizes (lane A). Corresponding hybridization of the same DNAs wereperformed with the 1.1-kb SnaBI fragment from pPZ1 (lanes B andC). Sizes of reference plasmids are indicated on the left, and P.aeruginosa RNL-1 plasmid sizes are shown in the middle, alongwith the position of chromosomal DNA (Chr.) migration.
RNL-1 (Fig. 3, lane B). Direct mating-out experiments failedto transfer the I-lactam resistance marker into P. aeruginosaor E. coli JM109. A probe consisting of the 1.1-kb SnaBIfragment from plasmid pPZ1 failed to hybridize to plasmidDNA isolated from P. aeruginosa RNL-1. However, thisprobe did hybridize significantly with residual chromosomalDNA which was present as contamination of plasmid DNApreparations (Fig. 3, lane D).
Molecular mass. The relative molecular mass of the cloned13-lactamase was estimated from cultures of E. coli JM109harboring pRAZi, which codes for the cloned 3-lactamase.Like E. coli JM109 harboring pPZ1, E. coli harboring pRAZi(SnaBI fragment from pPZ1 inserted into pKl9) showed thesame 1-lactamase activity at pI 5.4. SDS-gel electrophoresisrevealed a protein with a relative molecular mass of 29 kDa(Fig. 4). After renaturation of the gel, only this 29-kDaprotein band showed ,B-lactamase activity.DNA-DNA homologies. Hybridization performed with the
1.1-kb SnaBI probe from pPZ1 containing the 3-lactamasegene revealed a strong positive signal in response to a 1.1-kbfragment of SnaBI-digested P. aeruginosa RNL-1 DNA,thus confirming the origin of the cloned ,-lactamase gene.Hybridization with the same probe failed to show anyhomology with the blaTEM-l, blaSRV-3, blaPSE-l, blaPSE-2,blaPSE-3, blaPSE-4, blaOXA-1, blaOXA-2, blaOXA-3, or ampCgene from P. aeruginosa or the blaL l or blaL 2 gene from X.maltophilia.
FIG. 4. SDS-polyacrylamide gel electrophoresis of PER-i 3-lac-tamase. Crude extracts of E. coli JM109 harboring pKl9 (lane A)and pRAZi (SnaBI fragment from pPZ1 cloned into the pKl9 SmnaIsite), which produces PER-i 13-lactamase (lane B). The additionalband in lane B (compared with the lane A protein profile) corre-sponding to the PER-i P3-lactamase is indicated by the arrow.Protein molecular mass reference markers are shown on the left(sizes are in kilodaltons).
DISCUSSION
P. aeruginosa RNL-i, isolated from the urinary tract of aninfected patient, encodes two 13-lactamases: a likely ceph-alosporinase with a pI of 8.7 (ranking within the pI range ofcephalosporinases of P. aeruginosa (38]) and a novel 13-lac-tamase, PER-i, with a pI of 5.4. The PER-i enzyme is anextended-spectrum cephalosporin-hydrolyzing 13-lactamasesensitive to inhibition by clavulanic acid, imipenem, andcephamycins, and its gene appears to be chromosomallyencoded. It is particularly interesting that this 13-lactamase isnot derived from previously known extended-spectrum13-lactamases, according to the results of hybridization ex-periments.Extended-spectrum cephalosporin-hydrolyzing 13-lacta-
mases sensitive to clavulanic acid are mainly derivativesfrom plasmid-mediated TEM and SHV enzymes (16). Theyhave been extensively described in Enterobacteriaceae spe-cies. The ceftazidime-cefotaxime MIC ratio of >8 indicatesthat the PER-i enzyme is a ceftazidime-hydrolyzing en-zyme, like the TEM-5, TEM-6, and TEM-7 P-lactamases,with which it shares an acidic pI. However, the high Vm.,
966 NORDMANN ET AL.
1.-
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

EXTENDED-SPECTRUM I-LACTAMASE FROM P. AERUGINOSA 967
combined with high Km values for extended-spectrum ceph-alosporins, with PER-1 ,3-lactamase is not present in anyknown TEM or SHV derivatives. PER-1 ,B-lactamase did notresult from point mutations of TEM or SHV derivatives, ashybridization experiments failed to report any homologyunder high- or low-stringency conditions. According to theBush 3-lactamase classification, one may classify PER-1,-lactamase within group 2b', although it differs from TEMand SHV derivatives (9). Recently, plasmid-mediated non-TEM, non-SHV, extended-spectrum ,-lactamases havebeen reported in Kiebsiella oxytoca and E. coli (4, 44). Theseenzymes, named MEN-1 and KH, differ from PER-1 P-lac-tamase by their pls of 8.4 and 5.2, respectively, and theirlower hydrolytic properties regarding extended-spectrumcephalosporins. PER-1 differs particularly by its pI and itsstrong inhibition of activity by clavul4dic acid from rareplasmid-mediated extended-spectrum --lactamases whichhydrolyze cephamycins, such as CMY-1, CMY-2, andMIR-1 (6, 31). These enzymes have hydrolytic propertiesthat resemble those of chromosomal AmpC cephalospori-nase. In fact, protein sequence analysis of CMY-2 isolatedfromK pneumoniae revealed that it isi97% similar to AmpCfrom Citrobacter freundii (7). Similarly, BIL-1 may derivefrom an AmpC-type origin (33). The PER-1 enzyme sharessome similarities with the Bush; group 2e enzymes (10).These enzymes, such as FEC-1 from E. coli, Form II fromCitrobacter diversus ULA-27, and L-2 from X. maltophilia,are active predominantly against cephalosporins and areinhibited by clavulanic acid. However, relatively low ratesof hydrolysis of extended-spectrum cephalosporins and dif-ferences in pIs (alkaline pI) indicate that these enzymesdiffer from the PER-1 ,-lactamase. Moreover, in the case ofthe L-2 j-lactamase from X. maltophilia, chrdmosomalDNA of this species did not hybridize with a probe contain-ing the PER-1 13-lactamase gene.
Hybridizations with the oxacillin-hydrolyzing and carbe-nicillin-hydrolyzing 3-lactamase genes tested failed to revealany homology with the PER-1 ,-lactamase gene probe.Although genetic diversity has been reported among theoxacillin-hydrolyzing enzymes and all available oxacillin-hydrolyzing enzyme genes were not tested in our hybridiza-tion study, the absence of oxacillin hydrolysis for the PER-1P-lactamase excludes this enzyme from this group. More-over, as opposed to the strong inhibition property of clavu-lanic acid described here, oxacillin-hydrolyzing enzyme ac-tivities are usually poorly inhibited by this inhibitor. Inaddition, oxacillin-hydrolyzing enzymes are inhibited byNaCl (100 mM) (10), which was not the case for PER-11-lactamase. The published sequences of carbenicillinasesindicate that they are closely related at both the DNA andprotein levels. Therefore, negative hybridization performedwith blaPSE and blacARB genes excludes assignment of thePER-1 enzyme to this group of ,-lactamases.By in vitro mutagenesis, it has been shown that mutation
in the blaPSEA gene, as well as in the blaCARB-4 gene maylead to extended-spectrum derivatives (8). Although theextended-spectrum properties of the PER-1 1-lactamasemay suggest the occurrence of a similar event, the hybrid-ization experiments we performed do not argue in favor ofthe isolation of such a carbenicillin-hydrolyzing 3-lactamasederivative.
Recently, two novel ,B-lactamases were reported in P.aeruginosa which are also different from PER-1 3-lactamase(21, 29). The first novel chromosomally mediated ,B-lacta-mase hydrolyzes oxacillin strongly and extended-spectrumcephalosporins weakly (29). It has a pI of 8.0, and its activity
is not inhibited by clavulanic acid. Similar to the PER-1P-lactamase, its activity is inhibited by imipenem. Theproperty of imipenem acting as an inhibitor has been alreadydescribed for the Ambler class A P-lactamase of Bacilluscereus (28). In this case, imipenem reversibly reacted withthe enzyme with transient formation of an altered ,B-lacta-mase. Concomitant induction of the chromosomal cephalo-sporinase by imipenem and inhibition of the pI 8.0 enzymegave an image of simultaneous synergy and antagonismwhen a disc of imipenem was placed in front of a disc ofceftazidime in an agar diffusion assay. Such an image couldnot be found with the P. aeruginosa RNL-1 strain, since thePER-1 f-lactamase strongly hydrolyzes extended-spectrumcephalosporins. The second recently reported ,3-lactamase isa non-TEM-, non-SHV-derived 3-lactamase (21). It is plas-mid mediated, has a pI of 6.4, and like the PER-1 enzyme,confers resistance to extended-spectrum cephalosporins.However, the Vm. values of extended-spectrum cephalos-porins are much lower than that of the PER-1 P-lactamase,which hydrolyzes oxacillin and whose activity is not inhib-ited by clavulanic acid. As suggested by its biochemicalproperties, this pI 6.4 enzyme is an oxacillin-hydrolyzingenzyme. Indeed, sequence analysis revealed that it is aderivative of the blaPSE2 gene obtained by point mutationswhich confer resistance to extended-spectrum cephalospor-ins (21; oral communication).
Interestingly, two discrepancies were noted between theMICs obtained for E. coli possessing recombinant plasmidpPZ1 and the hydrolytic properties of the PER-1 enzyme. (i)Concerning aztreonam, its Vm., and VmaIjKm ratio wereparticularly low compared with the corresponding MIC.Such a high MIC may have resulted from poor penetration ofaztreonam through the outer membrane barrier. Such aproperty has also been reported for the TEM-22 f-lactamase(1). (ii) It is intriguing how moxalactam could behave as anefficient inhibitor (Ki, 3 x 10-3 PM) and had a slight butreproducible increase in its MIC as measured in E. colipossessing recombinant plasmid pPZ1, which codes forPER-1.As resistance to aminoglycosides and to sulfonamides was
also found in P. aeruginosa RNL-1, and as such antibiotic-resistant genes are present within Tn2l and Tn3 transposonderivatives of this species (20), further work will evaluate thepotential presence of a transposon containing these antibi-otic resistance genes along with the PER-1 3-lactamasegene. Sequence analysis of the PER-i-encoding gene willhelp to compare it with other extended-spectrum ,B-lacta-mases and will likely cotifirm its serine-enzymatic nature, asstrongly suggested by its biochemical properties. Absence ofinhibition of its activity by EDTA makes it different from thereported plasmid-mediated carbapenem-hydrolyzing 3-lacta-mase in P. aeruginosa, which also strongly hydrolyzesextended-spectrum cephalosporins (42). Finally, from a ther-apeutic point of view, this report emphasizes the fact thatwith P. aeruginosa, 3-lactamases other than the commonlyfound cephalosporinases may lead to failure of therapeuticregimens which include extended-spectrum cephalosporins.
ACKNOVLEDGMENTS
We thank C. Nauciel, in whose laboratory we isolated P. aerug-inosa RNL-1; W. Arber, in whose laboratory part of this work wasperformed; and M. Barthelemy, for precious advice. We are alsograteful to C. Bailly, A. Morand, and M. Thouverez for technicalassistance.
This study was supported by Merck Sharp & Dohme Chibret, the
VOL. 37, 1993
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIMICROB. AGENTS CHEMOTHER.
Fondation pour la Recherche Medicale, and the Ministere del'Education Nationale, France.
REFERENCES1. Arlet, G., M. Rouveau, G. Fournier, P. H. Lagrange, and A.
Philippon. 1992. Novel extended-spectrum 3-lactamase (TEM-22) in Klebsiella pneumoniae conferring higher resistance toaztreonam than to extended-spectrum cephalosporins. ProgramAbstr. 32nd Intersci. Conf. Antimicrob. Agents Chemother.,abstr. 1270.
2. Barthelemy, M., M. Guionie, and R. Labia. 1978. Beta-lacta-mases: determination of their isoelectric points. Antimicrob.Agents Chemother. 13:695-698.
3. Barthelemy, M., J. Peduzzi, H. Ben Yaghlane, and R. Labia.1988. Single amino-acid substitution between SHV-1 P-lacta-mase and cefotaxime-hydrolysing SHV-2 enzyme. FEBS Lett.231:217-220.
4. Barthelemy, M., J. Pduzzi, H. Bernard, C. Tancrede, and R.Labia. 1992. Close amino-acid sequence relationship betweenthe new plasmid-mediated extended-spectrum ,B-lactamaseMEN-1 and chromosomally encoded enzymes of Klebsiellaoxytoca. Biochim. Biophys. Acta 1122:15-22.
5. Barthelemy, M., J. PWduzzi, C. Verchere-Beaur, H. Ben Yagh-lane, and R. Labia. 1988. Purification and biochemical proper-ties of Pitton's 2 ,-lactamase (SHV-1). Ann. Inst. Pasteur/Microbiol. (Paris) 137B:19-27.
6. Bauernfeind, A., Y. Chong, and S. Schweinghart. 1989. Ex-tended broad spectrum 1-lactamase in Klebsiella pneumoniaeincluding resistance to cephamycins. Infection 17:316-321.
7. Bauernfeind, A., P. Mangold, S. Schweighart, G. H. Miller, K. J.Shaw, H. Giamarellou, and K. D. Dornbush. 1992. Molecularanalysis of a transferable cephamycinase in Kiebsiella pneumo-niae. Program Abstr. 32nd Intersci. Conf. Antimicrob. AgentsChemother., abstr. 1268.
8. Bejaoui, N., and R. C. Levesque. 1992. Structure-function anal-ysis of critical amino acids for extended-spectrum ceftazidimaseactivity in Pseudomonas aeruginosa carbenicillinases. ProgramAbstr. 32nd Intersci. Conf. Antimicrob. Agents Chemother.,abstr. 579.
9. Bush, K. 1989. Classification of P-lactamases: groups 1, 2a, 2b,and 2b'. Antimicrob. Agents Chemother. 33:264-270.
10. Bush, K. 1989. Classification of P-lactamases: groups 2c, 2d, 2e,3, and 4. Antimicrob. Agents Chemother. 33:271-276.
11. Chang, A. C. Y., and S. N. Cohen. 1978. Construction andcharacterization of amplifiable multicopy DNA cloning vehiclesderived from the P15A cryptic miniplasmid. J. Bacteriol. 134:1141-1156.
12. Datta, N., R. W. Hedges, E. J. Shaw, R. B. Sykes, and M. H.Richmond. 1971. Properties of an R factor from Pseudomonasaeruginosa. J. Bacteriol. 108:1244-1249.
13. Hedges, R. W., and G. A. Jacoby. 1980. Compatibility andmolecular properties of plasmid Rms 149 in Pseudomonasaeruginosa and Eschenichia coli. Plasmid 3:1-6.
14. Honore, N., M. H. Nicolas, and S. T. Cole. 1986. Induciblecephalosporinase production in clinical isolates of Enterobactercloacae is controlled by a regulatory gene that has been deletedfrom Escherichia coli. EMBO J. 5:3709-3714.
15. Jacob, A. A., J. A. Shapiro, L. Yamamoto, D. I. Smith, S. N.Cohen, and D. Berg. 1977. Plasmids studied in Escherichia coliand other enteric bacteria, p. 607-638. In A. I. Bukhari, J. A.Shapiro, and S. L. Adhya (ed.), DNA insertion elements,plasmids and episomes. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.
16. Jacoby, G. A., and A. A. Medeiros. 1991. More extended-spectrum ,-lactamases. Antimicrob. Agents Chemother. 35:1697-1704.
17. Kieser, T. 1984. Factors affecting the isolation of CCCDNAfrom Streptomyces lividans and Eschenchia coli. Plasmid 12:19-36.
18. Labia, R., M. Guionie, M. Barthelemy, and A. Philippon. 1981.Properties -of three carbenicillin hydrolysing 3-lactamases(CARB) from Pseudomonas aeruginosa: identification of a newenzyme. J. Antimicrob. Chemother. 7:49-56.
19. Laemmli, U. K., and M. Farve. 1973. Maturation of the head ofbacteriophage T4. I. DNA packaging events. J. Mol. Biol.80:575-599.
20. Levesque, R. C., and G. A. Jacoby. 1988. Molecular structureand interrelationships of multiresistance P-lactamase trans-posons. Plasmid 19:21-29.
21. Livermore, D. M., L. M. C. Hall, D. Gur, M. Akova, and H. E.Akalin. 1992. Non-TEM, non-SHV ceftazidimase, transferablefrom a P. aeruginosa isolate. Program Abstr. 32nd Intersci.Conf. Antimicrob. Agents Chemother., abstr. 585.
22. Livermore, D. M., and Y. J. Yang. 1987. 3-Lactamase labilityand inducer power of newer P-lactam antibiotics in relation totheir activity against 3-lactamase-inducibility mutants ofPseudomonas aeruginosa. J. Infect. Dis. 155:775-782.
23. Macrina, F. L., D. J. Kopecko, K. R. Jones, D. J. Ayers, andS. M. McCowen. 1978. A multiple plasmid containing Esche-richia coli strain: convenient source of size reference for plas-mid molecules. Plasmid 1:417-420.
24. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
25. Massidda, O., G. M. Rossolini, and G. Satta. 1991. The Aero-monas hydrophila cphA gene: molecular heterogeneity amongclass B metallo-p-lactamases. J. Bacteriol. 173:4611-4617.
26. Medeiros, A. A., and G. A. Jacoby. 1986. Beta-lactamasemediated resistance, p. 49-84. In S. F. Queener, J. A. Webler,and S. W. Queener (ed.), Beta-lactam antibiotics for clinicaluse. Marcel Dekker, Inc., New York.
27. Michel-Briand, Y., M. J. Dupont, I. Chardon-Loriaux, and M.Jouvenot. 1981. Isolation of an antibiotic multiresistance plas-mid from Pseudomonas aeruginosa. J. Antimicrob. Chemother.7:371-378.
28. Monks, J., and S. G. Waley. 1988. Imipenem as substrate andinhibitor of ,B-lactamases. Biochem. J. 253:323-328.
29. Mugnier, P., M. D. Kitzis, A. Philippon, L. Gutmann, and E.Collatz. 1992. Simultaneous synergy and antagonism betweenimipenem (Imi) and other P-lactam antibiotics in a clinicalisolate ofPseudomonas aeruginosa (PAe). Program Abstr. 32ndIntersci. Conf. Antimicrob. Agents Chemother., abstr. 584.
30. Nicolas, M.-H., V. Jarlier, N. Honore, A. Philippon, and S. T.Cole. 1989. Molecular characterization of the gene encodingSHV-3 P-lactamase responsible for transferable cefotaxime re-sistance in clinical isolates of Klebsiella pneumoniae. Antimi-crob. Agents Chemother. 33:2096-2100.
31. Papanicolaou, G. A., A. A. Medeiros, and G. A. Jacoby. 1990.Novel plasmid-mediated ,B-lactamase (MIR-1) conferring resis-tance to oxyimino- and a-methoxy ,-lactams in clinical isolatesof Klebsiella pneumoniae. Antimicrob. Agents Chemother. 34:2200-2209.
32. Paul, G., M. Barthelemy, A. Philippon, J. Peduzzi, L. Gilley, R.Labia, and P. Nevot. 1988. Immunological comparison of con-stitutive P-lactamases of gram-negative bacteria by neutraliza-tion in zymogram gels: properties of anti-TEM-1 and anti-TEM-2 sera. Ann. Inst. Pasteur/Microbiol. (Paris) 139:435-451.
33. Payne, D. J., N. Woodford, and S. G. B. Amyes. 1992. Charac-terization of the plasmid mediated 3-lactamase BIL-1. J. Anti-microb. Chemother. 30:119-127.
34. Philippon, A., R. Labia, and G. Jacoby. 1989. Extended-spec-trum 3-lactamases. Antimicrob. Agents Chemother. 33:1131-1136.
35. Philippon, A. M., G. Paul, and G. A. Jacoby. 1983. Properties ofPSE-2 13-lactamase and genetic basis for its production inPseudomonas aeruginosa. Antimicrob. Agents Chemother. 24:362-369.
36. Plesiat, P., B. Alkhalaf, and Y. Michel-Briand. 1988. Prevalenceand profile of plasmids in Pseudomonas aeruginosa. Eur. J.Clin. Microbiol. Infect. Dis. 7:261-264.
37. Pridmore, R. D. 1987. New and versatile cloning vectors withkanamycin resistance marker. Gene 56:309-312.
38. Sanders, C. C., M. L. Gates, and W. E. Sanders, Jr. 1988.Heterogeneity of class I r-lactamase expression in clinicalisolates of Pseudomonas aeruginosa. Antimicrob. AgentsChemother. 32:1893-1895.
968 NORDMANN ET AL.
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

EXTENDED-SPECTRUM ,-LACTAMASE FROM P. AERUGINOSA 969
39. Sanders, W. E., Jr., and C. C. Sanders. 1988. Inducible P-lac-tamases: clinical and epidemiological implications for use ofnewer cephalosporins. Rev. Infect. Dis. 10:820-838.
40. Southern, E. M. 1975. Detection of specific sequences among
DNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
41. Stanisich, V., and J. M. Ortiz. 1976. Similarities betweenplasmids of the P incompatibility group derived from differentbacterial genera. J. Gen. Microbiol. 94:281-289.
42. Watanabe, M., S. lyobe, M. Inoue, and S. Mitsuhashi. 1991.Transferable imipenem resistance in Pseudomonas aeruginosa.
Antimicrob. Agents Chemother. 35:147-151.43. Witchitz, J. L., and Y. A. Chabbert. 1971. Resistance transfer-
able a la gentamicine. Expression du caractere de resistance.Ann. Inst. Pasteur (Paris) 121:733-743.
44. Wu, S. W., K. Dornbusch, M. Norgren, and G. Kronvall. 1992.Extended-spectrum 13-lactamase from Klebsiella oxytoca, notbelonging to the TEM or SHV family. J. Antimicrob.Chemother. 30:3-16.
45. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. ImprovedM13 phage cloning vectors and host strains: nucleotide se-
quences of the M13mpl8 and pUC19 vectors. Gene 33:103-109.
VOL. 37, 1993
on August 26, 2018 by guest
http://aac.asm.org/
Dow
nloaded from





























