Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 2011, p. 7772–7778 Vol. 77, No. 210099-2240/11/$12.00 doi:10.1128/AEM.05761-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Characterization of a Feedback-Resistant Mevalonate Kinase from theArchaeon Methanosarcina mazei�
Yuliya A. Primak, Mai Du, Michael C. Miller, Derek H. Wells, Alex T. Nielsen,Walter Weyler, and Zachary Q. Beck*
Genencor, 925 Page Mill Road, Palo Alto, California 94304
Received 20 June 2011/Accepted 26 August 2011
The mevalonate pathway is utilized for the biosynthesis of isoprenoids in many bacterial, eukaryotic, andarchaeal organisms. Based on previous reports of its feedback inhibition, mevalonate kinase (MVK) may playan important regulatory role in the biosynthesis of mevalonate pathway-derived compounds. Here we reportthe purification, kinetic characterization, and inhibition analysis of the MVK from the archaeon Methanosar-cina mazei. The inhibition of the M. mazei MVK by the following metabolites derived from the mevalonatepathway was explored: dimethylallyl diphosphate (DMAPP), geranyl pyrophosphate (GPP), farnesyl pyrophos-phate (FPP), isopentenyl monophosphate (IP), and diphosphomevalonate. M. mazei MVK was not inhibited byDMAPP, GPP, FPP, diphosphomevalonate, or IP, a proposed intermediate in an alternative isoprenoidpathway present in archaea. Our findings suggest that the M. mazei MVK represents a distinct class ofmevalonate kinases that can be differentiated from previously characterized MVKs based on its inhibitionprofile.
Isoprenoids are a large and diverse class of compounds con-taining more than 40,000 naturally occurring terpenes and ter-penoids (33). They encompass many classes of bioactive mol-ecules, including carotenoids, steroid hormones, phytols, redoxcarriers, secondary metabolites, and pheromones, that makethem commercially attractive for the production of compoundsvarying from pharmaceuticals to biofuels (5, 15, 21, 23, 24).Currently, a number of groups are working on increasing theproduction of terpenoid compounds for a variety of medicinal,agricultural, sustainable biofuel, and biomaterial applications(21, 23, 24, 33). All isoprenoids are biosynthesized from thefive carbon precursors, isopentenyl diphosphate (IPP), and itsisomer, dimethylallyl diphosphate (DMAPP). Two pathwaysfor the biosynthesis of these central metabolites have beendescribed, the mevalonate pathway (28) and the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway (25). The meval-onate pathway typically is found in animals, plants, and inmany Gram-positive bacteria, including Streptococcus pneu-moniae (17, 31, 32). Some enzymes of the mevalonate pathwayalso have been identified in archaea; however, the completepathway has not been elucidated (27). The mevalonate path-way catalyzes the conversion of three molecules of acetyl co-enzyme A (CoA) to IPP and DMAPP. Briefly, two moleculesof acetyl-CoA undergo a Claisen condensation to form aceto-acetyl-CoA, which is catalyzed by acetoacetyl-CoA thiolase.3-Hydroxy-3-methylglutaryl-CoA (HMG-CoA) synthase thencatalyzes an aldol reaction between acetoacetyl-CoA and athird molecule of acetyl-CoA. The conversion of HMG-CoA tomevalonate subsequently is catalyzed by HMG-CoA reductase.Mevalonate kinase (MVK) and phosphomevalonate kinase
(PMK) catalyze the phosphorylation of the primary alcohol ofmevalonate and the phosphate of phosphomevalonate, respec-tively, to form diphosphomevalonate. The penultimate reac-tion in the pathway is the phosphorylative decarboxylation ofdiphosphomevalonate catalyzed by the diphosphomevalonatedecarboxylase to yield IPP (10, 32). IPP isomerase (IDI) cat-alyzes the conversion of IPP to DMAPP (Fig. 1).
A distinguishing characteristic of archaeal organisms is thatisoprenoids make up the major component of their membranelipids. In contrast, the lipids of eukaryotic and bacterial organ-isms are composed primarily of fatty acids (6, 17, 20, 27).Studies of isoprenoid biosynthesis in archaea have demon-strated that both acetate and mevalonate are precursors forIPP formation, indicating that the mevalonate pathway is in-volved in their biosynthesis (11, 17). Putative homologues of allmevalonate pathway genes, excluding the diphosphomeval-onate decarboxylase, have been identified in archaea bygenomic analysis (3, 7, 17, 19). In addition, putative isopente-nyl monophosphate kinases have been identified and charac-terized from archaea, suggesting the possible utilization of amodified mevalonate pathway for the production of iso-prenoids in archaea (8, 17) (Fig. 1).
Eukaryotic, bacterial, and archaeal organisms must ensurethe sufficient production of a variety of isoprenoid compoundsthat are essential for the proper growth, signaling, transport,and life cycle controls as well as the prevention of the overac-cumulation of potentially toxic products, such as cholesterol(15, 27). Organisms manage these tasks through the intricateregulation of isoprenoid-producing pathways (15). MVK wasdemonstrated to be an important regulatory point in the me-valonate pathway in both bacteria (1, 2, 31) and eukaryotes (4,9, 13, 16, 18). Previously the small-molecule regulation ofMVKs could be divided into two classes. The first class isinhibited by metabolites downstream of the diphosphomeval-onate decarboxylase reaction (IPP, DMAPP, GPP, FPP, and
* Corresponding author. Mailing address: Genencor, 925 Page MillRoad, Palo Alto, CA 94304. Phone: (650) 846-4003. Fax: (650) 845-6500. E-mail: [email protected].
� Published ahead of print on 9 September 2011.
7772
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

longer chain isoprenoids) (9, 16, 18, 31). The regulation of aeukaryotic MVK isolated from pig liver was first reported byDorsey and Porter in 1968 (9). Their detailed kinetic analysisrevealed the significant feedback regulation of this enzyme byGPP and FPP and, to a lesser degree, by DMAPP, IPP, and PPi
(9). Human MVK subsequently was characterized and foundto be inhibited by FPP, GPP, IPP, DMAPP, and geranylgeranylpyrophosphate (18, 22). The characterization of four plantMVKs and S. cerevisiae MVK by Gray and Kekwick in 1972revealed that they all are inhibited by GPP, FPP, geranylgera-nyl pyrophosphate, and phytyl pyrophosphate (16). In addi-tion, two MVKs from Gram-positive cocci, Staphylococcus au-
reus and Enterococcus faecalis, were found to be competitivelyinhibited by FPP with respect to ATP, with a Ki of 45 �M (31).
The second class of MVKs is inhibited by diphosphomeval-onate but not by metabolites downstream of the diphosphom-evalonate decarboxylase. Interestingly, DMAPP, IPP, GPP,and FPP were not feedback inhibitors of the Gram-positivebacterium S. pneumoniae MVK at concentrations of up to 12�M; however, diphosphomevalonate inhibited S. pneumoniaeMVK at nanomolar concentrations (2).
We report the overexpression, purification, kinetic analysis,and inhibition studies of the mvk gene product from the ar-chaeon Methanosarcina mazei. The S. cerevisiae and S. pneu-
FIG. 1. Mevalonate pathway. The proposed modified pathway for the production of isoprenoids in archaea organisms is illustrated in thebox (17).
VOL. 77, 2011 CHARACTERIZATION OF A FEEDBACK-RESISTANT M. MAZEI MVK 7773
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from
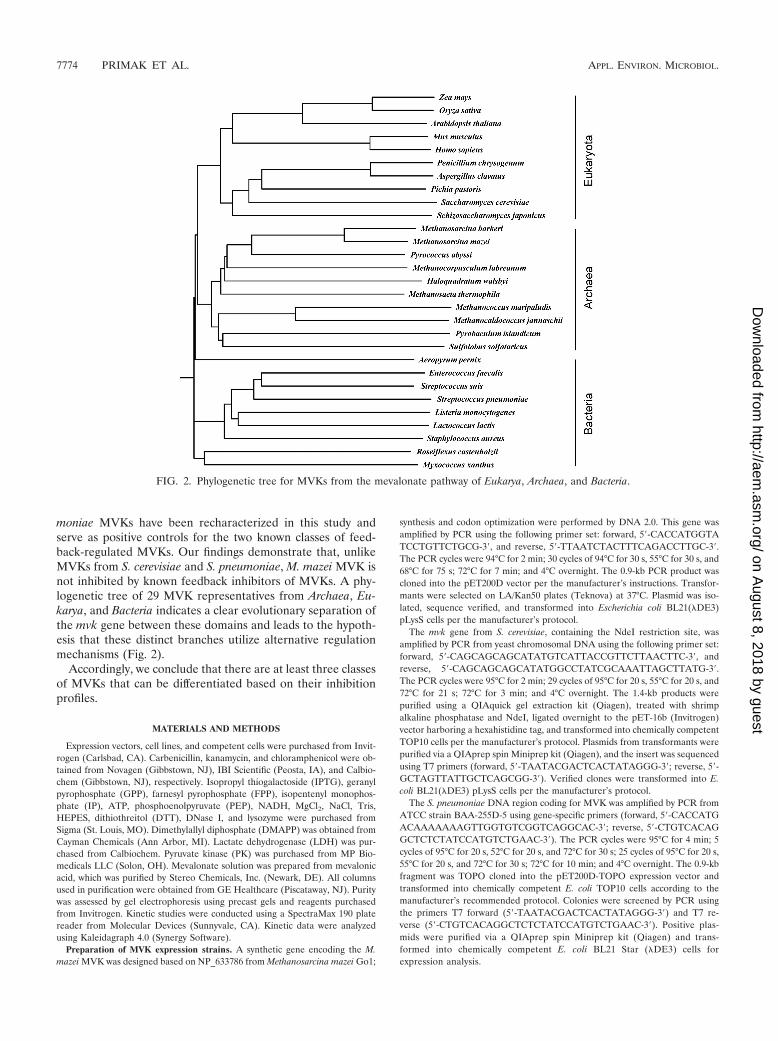
moniae MVKs have been recharacterized in this study andserve as positive controls for the two known classes of feed-back-regulated MVKs. Our findings demonstrate that, unlikeMVKs from S. cerevisiae and S. pneumoniae, M. mazei MVK isnot inhibited by known feedback inhibitors of MVKs. A phy-logenetic tree of 29 MVK representatives from Archaea, Eu-karya, and Bacteria indicates a clear evolutionary separation ofthe mvk gene between these domains and leads to the hypoth-esis that these distinct branches utilize alternative regulationmechanisms (Fig. 2).
Accordingly, we conclude that there are at least three classesof MVKs that can be differentiated based on their inhibitionprofiles.
MATERIALS AND METHODS
Expression vectors, cell lines, and competent cells were purchased from Invit-rogen (Carlsbad, CA). Carbenicillin, kanamycin, and chloramphenicol were ob-tained from Novagen (Gibbstown, NJ), IBI Scientific (Peosta, IA), and Calbio-chem (Gibbstown, NJ), respectively. Isopropyl thiogalactoside (IPTG), geranylpyrophosphate (GPP), farnesyl pyrophosphate (FPP), isopentenyl monophos-phate (IP), ATP, phosphoenolpyruvate (PEP), NADH, MgCl2, NaCl, Tris,HEPES, dithiothreitol (DTT), DNase I, and lysozyme were purchased fromSigma (St. Louis, MO). Dimethylallyl diphosphate (DMAPP) was obtained fromCayman Chemicals (Ann Arbor, MI). Lactate dehydrogenase (LDH) was pur-chased from Calbiochem. Pyruvate kinase (PK) was purchased from MP Bio-medicals LLC (Solon, OH). Mevalonate solution was prepared from mevalonicacid, which was purified by Stereo Chemicals, Inc. (Newark, DE). All columnsused in purification were obtained from GE Healthcare (Piscataway, NJ). Puritywas assessed by gel electrophoresis using precast gels and reagents purchasedfrom Invitrogen. Kinetic studies were conducted using a SpectraMax 190 platereader from Molecular Devices (Sunnyvale, CA). Kinetic data were analyzedusing Kaleidagraph 4.0 (Synergy Software).
Preparation of MVK expression strains. A synthetic gene encoding the M.mazei MVK was designed based on NP_633786 from Methanosarcina mazei Go1;
synthesis and codon optimization were performed by DNA 2.0. This gene wasamplified by PCR using the following primer set: forward, 5�-CACCATGGTATCCTGTTCTGCG-3�, and reverse, 5�-TTAATCTACTTTCAGACCTTGC-3�.The PCR cycles were 94°C for 2 min; 30 cycles of 94°C for 30 s, 55°C for 30 s, and68°C for 75 s; 72°C for 7 min; and 4°C overnight. The 0.9-kb PCR product wascloned into the pET200D vector per the manufacturer’s instructions. Transfor-mants were selected on LA/Kan50 plates (Teknova) at 37°C. Plasmid was iso-lated, sequence verified, and transformed into Escherichia coli BL21(�DE3)pLysS cells per the manufacturer’s protocol.
The mvk gene from S. cerevisiae, containing the NdeI restriction site, wasamplified by PCR from yeast chromosomal DNA using the following primer set:forward, 5�-CAGCAGCAGCATATGTCATTACCGTTCTTAACTTC-3�, andreverse, 5�-CAGCAGCAGCATATGGCCTATCGCAAATTAGCTTATG-3�.The PCR cycles were 95°C for 2 min; 29 cycles of 95°C for 20 s, 55°C for 20 s, and72°C for 21 s; 72°C for 3 min; and 4°C overnight. The 1.4-kb products werepurified using a QIAquick gel extraction kit (Qiagen), treated with shrimpalkaline phosphatase and NdeI, ligated overnight to the pET-16b (Invitrogen)vector harboring a hexahistidine tag, and transformed into chemically competentTOP10 cells per the manufacturer’s protocol. Plasmids from transformants werepurified via a QIAprep spin Miniprep kit (Qiagen), and the insert was sequencedusing T7 primers (forward, 5�-TAATACGACTCACTATAGGG-3�; reverse, 5�-GCTAGTTATTGCTCAGCGG-3�). Verified clones were transformed into E.coli BL21(�DE3) pLysS cells per the manufacturer’s protocol.
The S. pneumoniae DNA region coding for MVK was amplified by PCR fromATCC strain BAA-255D-5 using gene-specific primers (forward, 5�-CACCATGACAAAAAAAGTTGGTGTCGGTCAGGCAC-3�; reverse, 5�-CTGTCACAGGCTCTCTATCCATGTCTGAAC-3�). The PCR cycles were 95°C for 4 min; 5cycles of 95°C for 20 s, 52°C for 20 s, and 72°C for 30 s; 25 cycles of 95°C for 20 s,55°C for 20 s, and 72°C for 30 s; 72°C for 10 min; and 4°C overnight. The 0.9-kbfragment was TOPO cloned into the pET200D-TOPO expression vector andtransformed into chemically competent E. coli TOP10 cells according to themanufacturer’s recommended protocol. Colonies were screened by PCR usingthe primers T7 forward (5�-TAATACGACTCACTATAGGG-3�) and T7 re-verse (5�-CTGTCACAGGCTCTCTATCCATGTCTGAAC-3�). Positive plas-mids were purified via a QIAprep spin Miniprep kit (Qiagen) and trans-formed into chemically competent E. coli BL21 Star (�DE3) cells forexpression analysis.
FIG. 2. Phylogenetic tree for MVKs from the mevalonate pathway of Eukarya, Archaea, and Bacteria.
7774 PRIMAK ET AL. APPL. ENVIRON. MICROBIOL.
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from
Expression and purification of recombinant MVKs from M. mazei, S. cerevi-siae, and S. pneumoniae. Cells containing the M. mazei MVK expression plasmidwere grown in Terrific broth (26) supplemented with 50 mg/liter kanamycin and30 mg/liter chloramphenicol and were induced overnight with the addition of 0.5mM IPTG. Cells containing the MVK expression plasmids were grown in Luria-Bertani broth (26) supplemented with 50 mg/liter carbenicillin and 30 mg/literchloramphenicol for the expression of S. cerevisiae MVK or 50 mg/liter kanamy-cin for the expression of S. pneumoniae MVK, and they were induced overnightwith the addition of 0.2 mM IPTG at an optical density at 600 nm (OD600) of�0.4 to 0.6. All cells were harvested by centrifugation at 10,000 � g for 10 minand resuspended in 0.05 M sodium phosphate, 0.3 M sodium chloride, 0.02 Mimidazole (pH 8.0) buffer containing lysozyme and DNase I. Resuspended cellswere lysed by repeated passes through a French pressure cell at 20,000 lb/in2.Cell lysates were clarified by ultracentrifugation at 229,000 � g for 1 h. Thesupernatants were loaded onto a HiTrap IMAC HP column charged with nickelsulfate and equilibrated with 0.05 M sodium phosphate, 0.3 M sodium chloride,0.02 M imidazole (pH 8.0). Enzymes were isolated using a linear gradient from0.02 to 0.5 M imidazole. Fractions containing MVK were identified using SDS-PAGE (Invitrogen), pooled, and desalted into 0.05 M HEPES, 0.05 M sodiumchloride (pH 7.4) with 1 mM DTT using a Hi Prep 26/10 desalting column. MVKfrom S. cerevisiae was further purified on an anion-exchange HiTrap Q HPcolumn. The column was washed with 0.05 M Tris, 0.05 M sodium chloride (pH7.6) with 1 mM DTT and eluted with a 0.05 to 1.0 M sodium chloride gradient.Fractions containing MVK were desalted into 0.05 M HEPES, 0.05 M sodiumchloride (pH 7.4) containing 1 mM DTT. The purity of all three enzymes wasgreater than 95% as judged by SDS-PAGE and Coomassie staining. The proteinswere optically quantitated at 280 nm using the following conversion factors: 0.343OD/mg/ml for M. mazei MVK, 0.597 OD/mg/ml for S. cerevisiae MVK, and 0.516OD/mg/ml for S. pneumoniae MVK. These values were obtained using theExPASy ProtParam tool (14).
Expression and purification of recombinant PMK enzyme from S. cerevisiae.The S. cerevisiae DNA region coding for the PMK protein was amplified by PCRusing gene specific primers (forward, 5�-CACCTCAGAGTTGAGAGCCTTCAGTGC-3�; reverse, 5�-GAATTCTGCATGCAGCTACCTTAAG-3�), TOPOcloned into the pET200D-TOPO expression vector (Invitrogen), and trans-formed into chemically competent E. coli TOP10 cells according to the manu-facturer’s recommended protocol. Colonies were screened by PCR using T7forward (5�-TAATACGACTCACTATAGGG-3�) and a gene-specific reverse(5�-GAATTCTGCATGCAGCTACCTTAAG-3�) primer. Positive plasmidswere purified via a QIAprep spin Miniprep kit (Qiagen) and transformed intochemically competent E. coli BL21(�DE3) cells for expression analysis. Cellscontaining the PMK expression plasmid were grown in Terrific broth supple-mented with 50 mg/liter kanamycin. The culture was induced with 0.2 mM IPTGat an OD600 of 0.9 and harvested by centrifugation after 6 h at 30°C. Thepurification of PMK involved nickel affinity and anion-exchange chromatographyand followed the same protocol as that described above for S. cerevisiae MVK.The purity was greater than 95% as judged by SDS-PAGE and Coomassiestaining. The protein was optically quantitated at 280 nm using a conversionfactor of 1.099 OD/mg/ml.
Native molecular mass determination. The native molecular masses of theMVKs and PMK were determined by size-exclusion chromatography using aSuperdex 200 10/300 GL column. The column was equilibrated using the follow-ing seven molecular standards with masses ranging from 6.5 to 669 kDa: apro-
tinin (6.5 kDa), RNase A (13.7 kDa), carbonic anhydrase (29 kDa), ovalbumin(43 kDa), conalbumin (75 kDa), aldolase (158 kDa), and thyroglobulin (669kDa). The column void volume was calculated using the elution volume of bluedextran 2000. Column equilibration and sample runs were performed in 50 mMHEPES, 150 mM NaCl (pH 7.4) buffer containing 1 mM DTT at room temper-ature. The masses of M. mazei, S. cerevisiae, and S. pneumoniae MVKs, as well asthat of S. cerevisiae PMK, were calculated using the linear fit to the plot of logmasses versus the elution volume obtained for the molecular standards.
Enzyme activity and inhibition by DMAPP, GPP, and FPP. The catalyticactivities of the MVKs were measured using a modified spectrophotometric assaythat couples ADP formation to pyruvate synthesis and reduction to lactate (13).The initial rate of disappearance of NADH serves as a measure of the phos-phorylation of mevalonate by MVK. The assays were performed in triplicate ina 96-well plate (Costar catalog number 9017) format at 30°C. Each 100-�lreaction mixture contained 0.4 mM PEP, 0.05 mM DTT, 0.32 mM NADH, 10mM MgCl2, 2 U of LDH, and 2 U of PK in 50 mM Tris, 50 mM NaCl (pH 7.6).
The Michaelis constants for MVK (designated Km-Mev) from M. mazei and S.pneumoniae were determined at a saturating concentration of ATP (5 mM) andwith mevalonate concentrations ranging from 0.005 to 5 mM. The reaction wasinitiated with the addition of 80 nM (0.25 �g) purified M. mazei or 60 nM (0.21�g) S. pneumoniae MVK. The Km-ATP for these MVKs was determined similarly,using saturating concentrations of mevalonate (1.25 mM) and ATP concentra-tions ranging from 0.005 to 5 mM. Km values for S. cerevisiae MVK weredetermined using the same procedure with the following exceptions: substrateconcentrations ranged from 0.039 to 5 mM, and the reaction was initiated byadding 10 nM (50.1 ng) of purified S. cerevisiae MVK. Absorbance changesassociated with the amount of NADH oxidized to NAD� were monitored con-tinuously at 340 nm and plotted against time to determine the rate of theMVK-coupled reactions. Protein inhibition studies were performed in quadru-plicate by adding terpenyl diphosphates (DMAPP, GPP, FPP, and diphospho-mevalonate) at various concentrations to the reaction mix. The inhibition studiesof M. mazei MVK also included studies with isopentenyl monophosphate.
Inhibition of MVKs by diphosphomevalonate. The inhibition of three MVKsby diphosphomevalonate was investigated using a spectrophotometric pyruvatekinase- and lactate dehydrogenase-coupled assay, as previously described (2).This approach couples two reactions of the mevalonate pathway, the initialphosphorylation of mevalonate by MVK and the subsequent conversion of phos-phomevalonate to diphosphomevalonate by PMK. Reaction mixtures contained100 mM Tris-HCl, 100 mM NaCl, 1 mM DTT, 10 mM MgCl2, 5 mM ATP, 2.5mM NADH, 4 mM PEP, 10 U of LDH, 10 U of PK, and 1 mM mevalonate.Initially, MVK was added to the reaction mixture and the depletion of NADHwas monitored at 386 nm. After all of the mevalonate was converted to phos-phomevalonate, S. cerevisiae PMK was added to the mixture to catalyze thereaction from phosphomevalonate to diphosphomevalonate. To test for thefeedback inhibition of the MVK by diphosphomevalonate, both PMK and MVKwere added simultaneously to the reaction mixture. The inhibition of M. mazeiMVK by diphosphomevalonate was evaluated using 1.7 �M MVK and 2 �MPMK. Inhibition studies of S. cerevisiae and S. pneumoniae MVKs by diphospho-mevalonate utilized 0.1 �M MVK and 1 �M PMK as well as 1 �M MVK and 2�M PMK, respectively.
Phylogenetic analysis of MVK. Sequences of MVK from a range of differentorganisms were retrieved and aligned using ClustalW multiple sequence align-
TABLE 1. Kinetic characterization of MVKs
MVK origin
Kinetic constant
Kmapp-Mev
a
(�M)
Kmapp-ATP
a
(�M)kcat
a (s�1) Ki (DPM)b
Ki-Mev
(DMAPP)b
(�M)
Ki-ATP(DMAPP)
b
(�M)
Ki-Mev(GPP)
b
(�M)
Ki-ATP (GPP)b
(�M)
Ki-Mev(FPP)
b
(�M)
Ki-ATP (FPP)b
(�M) Ki-(IP)b
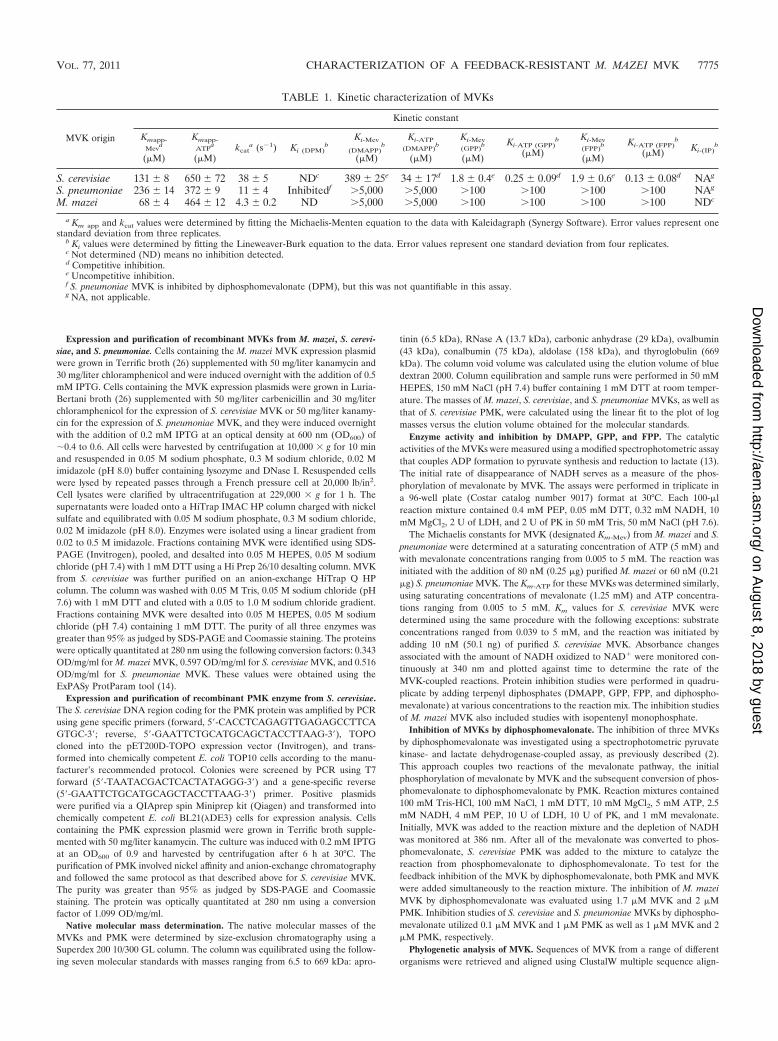
S. cerevisiae 131 � 8 650 � 72 38 � 5 NDc 389 � 25e 34 � 17d 1.8 � 0.4e 0.25 � 0.09d 1.9 � 0.6e 0.13 � 0.08d NAg
S. pneumoniae 236 � 14 372 � 9 11 � 4 Inhibitedf 5,000 5,000 100 100 100 100 NAg
M. mazei 68 � 4 464 � 12 4.3 � 0.2 ND 5,000 5,000 100 100 100 100 NDc
a Km app and kcat values were determined by fitting the Michaelis-Menten equation to the data with Kaleidagraph (Synergy Software). Error values represent onestandard deviation from three replicates.
b Ki values were determined by fitting the Lineweaver-Burk equation to the data. Error values represent one standard deviation from four replicates.c Not determined (ND) means no inhibition detected.d Competitive inhibition.e Uncompetitive inhibition.f S. pneumoniae MVK is inhibited by diphosphomevalonate (DPM), but this was not quantifiable in this assay.g NA, not applicable.
VOL. 77, 2011 CHARACTERIZATION OF A FEEDBACK-RESISTANT M. MAZEI MVK 7775
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

ment (30). A rooted phylogenetic tree (phenogram) was derived using the pro-gram DrawGram (12).
RESULTS
Characterization of MVKs. Three MVKs, from M. mazei, S.cerevisiae, and S. pneumoniae, as well as PMK from S. cerevi-siae, were expressed in E. coli, extracted, and purified usingaffinity chromatography to 95% apparent homogeneity. Theapparent masses of the MVKs and PMK were determined bygel filtration to be 78 kDa for M. mazei MVK, 97 kDa for S.cerevisiae MVK, 72 kDa for S. pneumoniae MVK, and 47 kDafor PMK. The calculated molecular masses using the aminoacid sequence are 35.5, 51, 39, and 53 kDa for M. mazei, S.cerevisiae, and S. pneumoniae MVKs and S. cerevisiae PMK,respectively. This suggests that the MVKs tested form dimersand that S. cerevisiae PMK is a monomer in solution.
The rates of mevalonate phosphorylation by the archaeal,eukaryotic, and bacterial MVKs were monitored (2). The ap-parent Km (Km app) values were evaluated for each enzyme withrespect to ATP (Km app-ATP) and mevalonate (Km app-Mev)using the Michaelis-Menten equation (Table 1). Of the threeenzymes assayed, M. mazei MVK had the slowest turnover(kcat) at 30°C of 4.3 s�1. S. cerevisiae MVK had a kcat nearlyfour times faster than that of M. mazei MVK. However, M.mazei MVK had the lowest apparent Km of 68 �M; S. cerevisiaeand S. pneumoniae MVKs had apparent Km values of 131 and236 �M, respectively.
M. mazei MVK is not inhibited by DMAPP, GPP, FPP, or IP.The potential inhibition of the M. mazei MVK by the down-stream products (DMAPP, GPP, and FPP) of the mevalonatepathway was evaluated. The catalytic activity of M. mazei MVKwas not inhibited by 5 mM DMAPP, 100 �M GPP, or 100 �MFPP. The archaeal mevalonate pathway has been postulated tocontain an IP kinase that catalyzes the formation of IPP, there-fore we examined the inhibition of M. mazei MVK by isopen-tenyl monophosphate (IP) (17). Our experiments demon-strated that M. mazei MVK is not inhibited by concentrationsof IP of up to 100 �M.
S. cerevisiae MVK is inhibited by DMAPP, GPP, and FPP,products of the mevalonate pathway. The MVK from yeast wasreported to be inhibited by GPP, FPP, geranylgeranyl pyro-phosphate, and phytyl pyrophosphate (16, 29) and serves as apositive control for a class of MVKs that are inhibited byintermediates downstream of diphosphomevalonate decarbox-ylase. In this study, the inhibition of S. cerevisiae MVK wasprobed with the isoprenoid precursors DMAPP, GPP, andFPP. Our results demonstrate that DMAPP, GPP, and FPP arecompetitive inhibitors of S. cerevisiae MVK with respect toATP and uncompetitive inhibitors with respect to mevalonate.The inhibition constants (Kis) of DMAPP, GPP, and FPP forthe S. cerevisiae MVK with respect to ATP were 34 � 17
FIG. 3. Conversion of mevalonate to phosphomevalonate cata-lyzed by S. cerevisiae (A), M. mazei (B), and S. pneumoniae (C) MVKswas monitored in the presence and absence of S. cerevisiae PMK. Therate of conversion of mevalonate to phosphomevalonate and subse-quently to diphosphomevalonate was detected indirectly by the oxida-tion of NADH at 386 nm. Reactions that were initiated by the simul-taneous addition of MVK and PMK are indicated by line i on eachgraph. Reactions that were initiated with MVK in the absence of PMKare indicated by line ii in each graph. Reactions were allowed toproceed until mevalonate was completely converted to phosphomeval-onate. PMK then was added to the reaction mixture at line iii tocomplete conversion to diphosphomevalonate. Reaction mixtures con-
tained the following components: 100 mM Tris-HCl, 100 mM NaCl, 1mM DTT, 10 mM MgCl2, 5 mM ATP, 2.5 mM NADH, 4 mM PEP, 10U of LDH, 10 U of PK, and 1 mM mevalonate. To this mixture wasadded 0.1 �M S. cerevisiae MVK and 1 �M S. cerevisiae PMK (A), 1.7�M M. mazei MVK and 2 �M S. cerevisiae PMK (B), and 1 �M S.pneumoniae MVK and 2 �M S. cerevisiae PMK (C).
7776 PRIMAK ET AL. APPL. ENVIRON. MICROBIOL.
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

(DMAPP), 0.25 � 0.09 (GPP), and 0.13 � 0.08 �M (FPP). TheKis of DMAPP, GPP, and FPP for S. cerevisiae MVK withrespect to mevalonate were 389 � 25 (DMAPP), 1.8 � 0.4(GPP), and 1.9 � 0.6 �M (FPP). The inhibition constants aresummarized in Table 1.
Similarly to M. mazei MVK, the S. pneumoniae MVK waspreviously demonstrated to be uninhibited by DMAPP, GPP,and FPP at concentrations of up to 12 �M (2). Significantlygreater concentrations of these metabolites may be encoun-tered during the metabolic engineering of terpenoid pathways;therefore, we assayed the inhibition of S. pneumoniae MVKusing 5 mM DMAPP, 100 �M GPP, and 100 �M FPP andconfirmed that the S. pneumoniae MVK is not inhibited atthese concentrations.
Inhibition of MVK by diphosphomevalonate. The effect ofdiphosphomevalonate on the rate of MVK reactions was ana-lyzed using a pyruvate kinase and lactate dehydrogenase cou-pling system, as previously described (2). Briefly, the additionof mevalonate to the reaction mixture containing MVK re-sulted in the quantitative conversion of the substrate to phos-phomevalonate. The subsequent addition of PMK resulted inthe conversion of phosphomevalonate to diphosphomeval-onate. To demonstrate feedback inhibition, both MVK andPMK were added at the initiation of the assay. The inhibitionof MVK was indicated if the rates of mevalonate conversion tophosphomevalonate and diphosphomevalonate were signifi-cantly decreased compared to those from the assays performedby the sequential addition of MVK and PMK. In our studies,when S. cerevisiae PMK and M. mazei MVK were present at theinitiation of the reaction, the mevalonate was completely con-verted into diphosphomevalonate (Fig. 3B). The same resultwas obtained when S. cerevisiae MVK was assayed with PMK(Fig. 3A), demonstrating that neither M. mazei nor S. cerevisiaeMVK is inhibited by diphosphomevalonate. However, when S.pneumoniae MVK and PMK were present at the initiation ofthe reaction, the velocity of mevalonate conversion signifi-cantly decreased (Fig. 3C), verifying the inhibition of S. pneu-moniae MVK by diphosphomevalonate (2).
DISCUSSION
These studies demonstrate that at least three classes ofMVKs can be distinguished based on their inhibition profiles(Fig. 4). Unlike previously reported MVKs, the MVK of thearchaeon M. mazei was not inhibited by DMAPP, GPP, FPP,diphosphomevalonate, or IP, a proposed intermediate of themevalonate pathway in archaea (17). The MVK of S. cerevisiaealso was not inhibited by diphosphomevalonate accumulationin our studies, but it was inhibited by DMAPP, GPP, and FPP,similarly to the human enzyme. Furthermore, the inhibition ofS. pneumoniae MVK was probed using 100 �M GPP and FPPas well as 5 mM DMAPP, but no significant inhibition ofenzyme activity was observed.
We hypothesize that MVKs have evolved with different reg-ulation mechanisms to accommodate their specific utilizationof isoprenoids (17, 20, 27). A phylogenetic tree of the MVKsfrom Bacteria, Archaea, and Eukarya was constructed to assessthe similarity between MVKs from the three domains of life(Fig. 2). Interestingly, the 29 MVKs that were surveyed clearlyseparated into three classes, suggesting the vertical transfer of
the mvk gene. It should be noted that the MVK of the ther-mostable archaeon Methanocaldococcus jannaschii has beenstudied by Huang et al. and was found to be inhibited bymicromolar concentrations of GPP, FPP, and IPP metabolites(19). However, the MVKs from M. mazei and M. jannaschii aredistantly related, with 32% amino acid sequence identity, andoccupy different branches of the archaeal dendrogram (Fig. 2).The specific activity of the M. jannaschii MVK at an optimumtemperature of 70 to 75°C was reported to be 387 �mol/min/mg. Approximately 25% of the maximal activity was observedat 30°C, the temperature at which our studies were conducted(19). The specific activity of M. mazei MVK we report is morethan 20 times less than the specific activity of M. jannaschiiMVK at 30°C. The regulation of these enzymes seems wide-spread and, therefore, likely is important for maintaining prop-erly functioning cells. Further studies are necessary to deter-mine if MVKs that are not inhibited by metabolites areregulated at the transcriptional, translational, or posttransla-tional level, or if the low catalytic efficiency of this enzyme isimportant for regulation.
REFERENCES
1. Andreassi, J. L., P. W. Bilder, M. W. Vetting, S. L. Roderick, and T. S. Leyh.2007. Crystal structure of the Streptococcus pneumoniae mevalonate kinase incomplex with diphosphomevalonate. Protein Sci. 16:983–989.
2. Andreassi, J. L., K. Dabovic, and T. S. Leyh. 2004. Streptococcus pneumoniaeisoprenoid biosynthesis is downregulated by diphosphomevalonate: an anti-microbial target. Biochemistry 43:16461–16466.
3. Barkley, S. J., S. B. Desai, and C. D. Poulter. 2004. Type II isopentenyldiphosphate isomerase from Synechocystis sp. strain PCC 6803. J. Bacteriol.186:8156–8158.
4. Beytia, E., J. K. Dorsey, J. Marr, W. W. Cleland, and J. W. Porter. 1970.Purification and mechanism of action of hog liver mevalonic kinase. J. Biol.Chem. 245:5450–5458.
FIG. 4. Diagram of the regulation of MVKs from S. pneumoniae, S.cerevisiae, and M. mazei by the intermediates of the mevalonate path-way. Shown is a schematic of the mevalonate pathway in S. cerevisiae,S. pneumoniae, and M. mazei with enzymes MVK, PMK, diphospho-mevalonate decarboxylase (MVD; unidentified in archaea), IDI, andfarnesyl diphosphate synthase (FPPS), as well as their correspondingintermediates, phosphomevalonate (PM), diphosphomevalonate(DPM), IPP, DMAPP, GPP, and FPP. Inhibition studies were per-formed with DMAPP, GPP, FPP, DPM, and IP. S. pneumoniae MVKis inhibited by DPM, whereas S. cerevisiae MVK is inhibited byDMAPP, GPP, and FPP. M. mazei MVK is not inhibited by DMAPP,GPP, FPP, or DPM. The inhibition of M. mazei MVK also was testedwith IP, the proposed intermediate of an alternative archaeal meval-onate pathway involving a putative phosphomevalonate decarboxylaseand isopentenyl monophosphate kinase (IPK). IP did not inhibit M.mazei MVK in our studies.
VOL. 77, 2011 CHARACTERIZATION OF A FEEDBACK-RESISTANT M. MAZEI MVK 7777
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5. Bohlmann, J., and C. I. Keeling. 2008. Terpenoid biomaterials. Plant J.54:656–669.
6. Boucher, Y., M. Kamekura, and W. F. Doolittle. 2004. Origins and evolutionof isoprenoid lipid biosynthesis in archaea. Mol. Microbiol. 52:515–527.
7. Bult, C. J., et al. 1996. Complete genome sequence of the methanogenicarchaeon Methanococcus jannaschii. Science 273:1058–1073.
8. Chen, M., and C. D. Poulter. 2010. Characterization of thermophilic archaealisopentenyl phosphate kinases. Biochemistry 49:207–217.
9. Dorsey, J. K., and J. W. Porter. 1968. The inhibition of mevalonic kinase bygeranyl and farnesyl pyrophosphates. J. Biol. Chem. 243:4667–4670.
10. Doun, S. S., I. I. Burgner, J. W. S. D. Briggs, and V. W. Rodwell. 2005.Enterococcus faecalis phosphomevalonate kinase. Protein Sci. 14:1134–1139.
11. Ekiel, I., I. C. P. Smith, and G. D. Sprott. 1983. Biosynthetic pathway inMethanospirillum hungatei as determined by 13C nuclear magnetic resonance.J. Bacteriol. 156:316–326.
12. Felsenstein, J. 1989. PHYLIP–phylogeny inference package (version 3.2).Cladistics 5:164–166.
13. Fu, Z., N. E. Voynova, T. J. Herdendorf, H. M. Miziorko, and J. P. Kim.2008. Biochemical and structural basis for feedback inhibition of mevalonatekinase and isoprenoid metabolism. Biochemistry 47:3715–3724.
14. Gasteiger, E., et al. 2005. Protein identification and analysis tools on theExPASy server, p. 571–607 In J. M. Walker (ed.), The proteomics protocolshandbook. Humana Press, Totowa, NJ.
15. Goldstein, J. L., and M. S. Brown. 1990. Regulation of the mevalonatepathway. Nature 343:425–430.
16. Gray, J. C., and R. G. O. Kekwick. 1972. The inhibition of plant mevalonatekinase preparations by prenyl pyrophosphates. Biochim. Biophys. Acta 279:290–296.
17. Grochowski, L. L., H. Xu, and R. H. White. 2006. Methanocaldococcusjannaschii uses a modified mevalonate pathway for biosynthesis of isopen-tenyl diphosphate. J. Bacteriol. 188:3192–3198.
18. Hinson, D. D., K. L. Chambliss, M. J. Toth, R. D. Tanaka, and K. M. Gibson.1997. Post-translational regulation of mevalonate kinase by intermediates ofthe cholesterol and nonsterol isoprene biosynthetic pathway. J. Lipid Res.38:2216–2223.
19. Huang, K., A. I. Scott, and G. N. Bennett. 1999. Overexpression, purification,and characterization of the thermostable mevalonate kinase from Methano-coccus jannaschii. Protein Expression Purif. 17:33–40.
20. Koga, Y., and H. Morii. 2007. Biosynthesis of ether-type polar lipids inarchaea and evolutionary considerations. Microbiol. Mol. Biol. Rev. 71:97–120.
21. Martin, V. J. J., D. J. Pitera, S. T. Withers, J. D. Newman, and J. D.Keasling. 2003. Engineering a mevalonate pathway in Escherichia coli forproduction of terpenoids. Nat. Biotechnol. 21:796–802.
22. Potter, D., J. M. Wojnar, C. Narasimhan, and H. M. Miziorko. 1997. Iden-tification and functional characterization of an active-site lysine in meval-onate kinase. J. Biol. Chem. 272:5741–5746.
23. Roberts, S. C. 2007. Production and engineering of terpenoids in plant cellculture. Nat. Chem. Biol. 3:387–395.
24. Rohdich, F., A. Bacher, and W. Eisenreich. 2005. Isoprenoid biosyntheticpathways as anti-infective drug targets. Biochem. Soc. Trans. 33:785–791.
25. Rohmer, M., M. Knani, P. Simonin, B. Sutter, and H. Sahm. 1993. Iso-prenoid biosynthesis in bacteria: a novel pathway for the early steps leadingto isopentenyl diphosphate. Biochemistry 295:517–524.
26. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed., vol. 3. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY.
27. Smit, A., and A. Mushegian. 2000. Biosynthesis of isoprenoids via meval-onate in Archaea: the lost pathway. Genome Res. 10:1468–1484.
28. Spurgeon, S. L., and J. W. Porter. 1981. Biosynthesis of isoprenoid com-pounds, vol. 1. John Wiley and Sons, New York, NY.
29. Tchen, T. T. 1958. Mevalonic kinase: purification and properties. J. Biol.Chem. 233:1100–1103.
30. Thompson, J. D., D. G. Higgins, and T. J. Gibson. 1994. CLUSTAL W:improving the sensitivity of progressive multiple sequence alignment throughsequence weighting, position-specific gap penalties and weight matrix choice.Nucleic Acids Res. 22:4673–4680.
31. Voynova, N. E., S. E. Rios, and H. M. Miziorko. 2004. Staphylococcus aureusmevalonate kinase: isolation and characterization of an enzyme of the iso-prenoid biosynthetic pathway. J. Bacteriol. 186:61–67.
32. Wilding, E. I., et al. 2000. Identification, evolution, and essentiality of themevalonate pathway for isopentenyl diphosphate biosynthesis in gram-pos-itive cocci. J. Bacteriol. 182:4319–4327.
33. Withers, A. T., and J. D. Keasling. 2007. Biosynthesis and engineering ofisoprenoid small molecules. Appl. Microbiol. Biotechnol. 73:980–990.
7778 PRIMAK ET AL. APPL. ENVIRON. MICROBIOL.
on August 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from





























