Embed Size (px)
Citation preview

RESEARCH ARTICLE
Characterization and taxonomical note about ThaiErianthus germplasm collection: the morphology, floweringphenology and biogeography among E. procerus and threetypes of E. arundinaceus
Shuichiro Tagane • Werapon Ponragdee •
Taksina Sansayawichai • Akira Sugimoto •
Yoshifumi Terajima
Received: 18 December 2010 / Accepted: 10 June 2011 / Published online: 4 August 2011
� Springer Science+Business Media B.V. 2011
Abstract Erianthus, one of the genus in Saccharum
complex, is important genetic resources for sugarcane
improvement. The morphology and flowering phenol-
ogy of 108 accessions belonging to Erianthus procerus
and three types of E. arundinaceus collected from
throughout Thailand were compared. PCA analysis
based on 22 characteristics clearly supported the
separation of Type II and Type III of E. arundinaceus
from E. procerus and Type I of E. arundinaceus
according to their morphological characteristics, par-
ticularly their bud size, and the development of root
primordia, which greatly affected axis I of the PCA
analysis. E. procerus and Type I showed overlapping
in many of their characteristics including their flow-
ering periods. Flower characteristics such as rachis
joint length, which were used for previous taxonomic
classifications, were not available for the classification
of the Thai samples because of the wide variation and
overlapping among them. Most of these phenotype
similarities and differences are considered to have
developed convergently as a result of niche adaptation.
Type II and III inhabit riverbanks and streambeds
where floods occur frequently, while E. procerus and
Type I mainly grow in non-flooding areas such as
mountainous grassland, the edge of forests, and beside
fields. All Thai Erianthus show unique geographic
distributions in Thailand. In particular, the biogeo-
graphic boundary between Type II and Type III
appeared to be located at the Isthmus of Kra. Although
some types showed morphological similarities, repro-
ductive isolation among the four groups seemed to be
maintained by differentiation in geographic distribu-
tion, habitat preference, and flowering timing.
Keywords Biogeography � Erianthus
arundinaceus � Erianthus procerus � Reproductive
isolation � Taxonomy
Introduction
Knowledge of the wild germplasm is important for
more efficient breeding programs because it provides
the basis for the development of desirable plants. In
sugarcane breeding, interspecific and intergeneric
hybridization using Saccharum complex, consisting
of 5 genera of Saccharum, Erianthus, Miscanthus,
Narenga, Sclerostachya, have been conducted in
many countries. In particular, Erianthus spp. is
increasingly being used because of its desirable traits
S. Tagane � A. Sugimoto � Y. Terajima
Japan International Research Center for Agricultural
Sciences, Ishigaki 907-0002, Japan
Present Address:S. Tagane (&)
Department of Biology, Faculty of Science, Kyushu
University, 6-10-1, Hakozaki, Fukuoka 812-8581, Japan
e-mail: [email protected]
W. Ponragdee � T. Sansayawichai
Khon Kaen Field Crops Research Center, DOA,
Khon Kaen 40000, Thailand
123
Genet Resour Crop Evol (2012) 59:769–781
DOI 10.1007/s10722-011-9717-2

such as high biomass, drought tolerance, and resis-
tance to pests and diseases (D’Hont et al. 1995; Rott
et al. 1997; Piperidis et al. 2000; Ram et al. 2001;
Sugimoto et al. 2002; Cai et al. 2005).
The Genus Erianthus was established by Michaux
in Flora Boreali-Americana in 1803 based on the New
World species E. saccharoides Michaux. Old World
species display distinct differences from New World
species in many of their morphological traits; for
example, Old World species have three anthers
(Grassl 1972), and flavonoid di-C-glycoside is absent
from New World species (Williams et al. 1974).
These species are generally placed in the section
Ripidium, which is important for sugarcane breeding.
Erianthus species were sometimes classified under the
genus Saccharum (Chen and Phillips 2006; Hatusima
1975; Suvatti 1978) and were considered to be
members of the Saccharum complex. However, recent
molecular analysis based on rbcL (Zhang et al. 2002)
and ITS (Hodkinson et al. 2002) revealed that Old
World species (section Ripidium) are genetically
distinct from other species belonging to the Saccha-
rum complex.
Erianthus sect. Ripidium consists of 7 species,
which are distributed from the Mediterranean to
Southeast Asia (Grassl 1972; Daniels and Roach
1987). Two species, E. procerus (Roxb.) Raizada and
E. arundinaceus (Retz.) Jeswiet, have been paid
particular attention in sugarcane breeding programs
due to their desirable traits, such as their disease
resistance, improved ratooning and vigor, and stress
tolerance. E. procerus is distributed in NE India,
Myanmar, Thailand, and China, whereas E. arundin-
aceus has a wider distribution including India, China,
Myanmar, Thailand, Philippines, Japan, Malaysia,
Indonesia, and New Guinea. Since the basic chro-
mosome number of Erianthus was reported to be
x = 10, both species are known to polyploid, and
chromosome numbers of 2n = 40 in E. procerus and
2n = 30, 40, and 60 in E. arundinaceus have been
demonstrated (Daniels and Roach 1987; Amalraj and
Balasundaram 2006).
However, there has also been great confusion
regarding the two species, and they are sometimes
considered to be synonymous (Mukherjee 1958; Hole
1911). E. procerus was first proposed to be a species by
Roxburgh in Flora of India in 1820, but Hole (1911)
and Hooker (1896) accepted this reduction. Bor (1940)
reinstated this species as Saccharum procerum, since it
flowers after the flowering period of S. arundinaceum
is over, and possesses a ‘‘much looser’’ panicle with
longer rachis joints and pedicels. In addition to these
points, Amalraj and Balasundaram (2006) indicated
that E. procerus lacks vegetative cane; i.e., their buds
have difficulty in germination due to not prominent
buds and root primordia.
Recently, various molecular markers, including
RAPD (Nair and Mary 2006), 5SrDNA (Besse et al.
1996), the ITS (Hodkinson et al. 2002), RFLP (Besse
et al. 1997; Coto et al. 2002), AFLP (Arro et al. 2006;
Cai et al. 2005; Selvi et al. 2006), and SSR (Cai et al.
2005; Selvi et al. 2003; Swarup et al. 2009), have been
applied to determine the levels of genetic variation and
phylogenetic relationships among Erianthus species
and its relatives including sugarcane. These results
clarified that (1) the Erianthus Sect. Ripidium genera is
a far relative of sugarcane and other species belonging
to the Saccharum complex, and (2) E. arundinaceus
contains two genetically divergent groups, Indian
E. arundinaceus, which has a chromosome number
of 40 and is closely genetically related to Indian
E. procerus, and Indonesian E. arundinaceus, which
has a chromosome number of 60 and is a far genetic
relative of Indian E. arundinaceus and E. procerus.
Despite their well-resolved phylogenetic positions, the
taxonomy of E. procerus and E. arundinaceus still
remains controversial because of the wide morpholog-
ical variation and similar floral traits between the
species (Mukherjee 1958) and the limited amount of
plant materials available for these species, mainly from
India and Indonesia, where they had been paid attention
for a long time in sugarcane breeding program. In
addition, Besse et al. (1997) noted that E. arundinaceus
collected from Indonesia, New Guinea, and the Philip-
pines showed low levels of RFLP polymorphism.
In Thailand, which is geographically located
between the two regions, E. procerus and E. arundin-
aceus are found abundantly in nature. Therefore,
materials that would be useful for further taxonomic
investigations are expected to be available in Thailand.
In fact, cytological studies have revealed that Thai
E. arundinaceus displays two cytotypes, 2n = 40 and
60, while E. procerus shows 2n = 40 (Tagane,
unpublished). Further, 2n = 60 cytotypes of E. arund-
inaceus were appeared to include two divergent types
in their morphology and geographic distribution. That
is, totally three type of E. arundinaceus are considered
to be distributed in Thailand. Here, we examined the
770 Genet Resour Crop Evol (2012) 59:769–781
123

morphology, flowering phenology, and geographic
distribution among E. procerus and three types of E.
arundinaceus, in order to classify Thai Erianthus and
then discussed their geographic distribution and flow-
ering phenology, which contribute to their reproduc-
tive isolation.
Materials and methods
The taxonomic concept of E. procerus, proposed in
Amalraj and Balasundaram (2006), was accepted in
the present study. For E. arundinaceus, three types,
Types I, II and III, were recognized from our field,
phenotypic, and chromosomal observations as men-
tioned in Introduction. Type II and Type III were
considered to be Indian E. arundinaceus and Indo-
nesian E. arundinaceus, respectively.
Several field trips were conducted from 1997 to
gather samples of E. procerus and the three types of
E. arundinaceus from areas throughout their Thai
ranges (Sugimoto et al. 2002). The collected germ-
plasms were preserved at the Tha Phra site of the Khon
Kaen Field Crops Research Center (N16�200,E102�490, 150 m above sea level). The area has a
mean annual rainfall of around 1,215 mm
(1997–2007), 90% of which falls between April and
October. The accessions were propagated vegetatively
from stalk cuttings and planted in 1.5 m 9 4.5 m
plots, using 3 cuttings per plot with 1.5 m spaces within
rows and 1.5 m spaces between rows in 2003. The
plants were irrigated from November to December, and
no fertilizer was applied. So far, 148 clones have been
collected, but 108 fully-grown clones collected from
1997 to 2003 were examined in this study.
Each plant was scored on 21 morphological and
one ecological traits, as shown in Table 1. Three to
five stalks were randomly selected and measured. To
assess floret traits, three spikelets were randomly
selected from each plant and measured under stereo
microscope (Nikon). The mean and standard devia-
tion of each trait were estimated for each group.
Table 1 Description of 21
morphological and one
ecological characters used
in this study
Part No. Characters Description
Leaf 1 Leaf length Length from ligule to tip of leaf (cm)
2 Leaf width Width of leaf at the widest point (cm)
3 Leaf shape index Ratio: leaf length/leaf width
4 Midrib width Width of midrib of leaf blade at widest point (cm)
Stalk 5 Plant height From the base of the clum to flag leaf
6 Leaf sheath length Length of leaf sheath of the longest leaf blade
7 Leaf sheath hairiness An estimate of the average hairiness on leaf sheath;
(0) none, (1) sparse, (2) much, (3) dense
8 Wax on leaf sheath An estimate of the average wax on leaf sheath;
(0) none, (3) high
9 Length of hair on ligule Length of hair around ligule
10 Stem thickness Diameter of stalk at 1 m height (mm)
11 No. of root primordia No. of root primordia on stalk at around 1 m height
12 No of root eye rings No. of root eye ring on stalk at around 1 m height
13 Bud length Length of bud on stalk at around 1 m height (mm)
14 Bud width Width of bud on stalk at around 1 m height (mm)
15 Bud shape index Ratio: length/width
Flower 16 Length of inflorescence Length of inflorescence (cm)
17 Width of inflorescence Width of inflorescence at the widest point (cm)
18 No. of racemes Number of raceme joint
19 Rachis joint length Length of rachis between spikelets (mm)
20 Petiole length Length from the base of the callus to the tip (mm)
21 Glume length From the base of glume to the tip (mm)
22 Flowering date Days to coming anther after Oct. 27
Genet Resour Crop Evol (2012) 59:769–781 771
123

The flowering period of Erianthus normally starts
in October and lasts until January, which places it
within the dry season. The flowering date; i.e., the
day on which the first anther appears, was recorded
for 2 years, 2008/2009 and 2009/2010. Since the
results of the 2 years showed a similar pattern, the
flowering date collected in 2009/2010 was trans-
formed to days after the 27th of October and used in
the principal component analysis (PCA).
The characteristics were standardized across types,
and PCA was performed with 22 characters for 108
Erianthus clones to assess the phenotypic variation
among and within the four groups. Canonical
Discriminant Analysis (CDA) was performed to
detect how and which of 22 characteristics were
important for the separation of the four groups. Both
multivariate analyses were conducted using R soft-
ware (R Development Core Team 2007).
Results
Type I had much longer leaf blades and leaf sheathes
than Type II and Type III, but the leaf width of
E. procerus was much narrower than those of Type I
and II (Table 2). Consequently, E. procerus had the
narrowest leaves. The plant height of Type I varied
from 267.0 to 631.0 cm, and that of type III was
significantly smaller than those of E. procerus and
Type I. Concerning leaf sheath traits, only the leaf
sheath of Type I had hairs on its surface, while the
others only had hairs on the edge of their leaf sheath
and around the ligule. There was no wax on the leaf
sheathes of Type III, while the other three groups did
display wax on their leaf sheathes. Type II had the
significant biggest stalk in the other plants. All stalks
examined in Type II and Type III showed more root
primordia than those examined in E. procerus and
Type I. The mean bud sizes of the Type II and Type
III were 1.2 9 1.0 cm and 1.2 9 0.8 cm (length 9
width). Type II had the biggest buds, whereas those
of E. procerus were smallest (mean length 9 width:
0.5 9 0.4 cm), although the differences among
E. procerus, Type I and Type II were not significant.
A slightly larger inflorescence size was shown in
E. procerus and Type I, and rachis length also
showed a wide variation.
The same pattern of flowering phenology was
confirmed between 2008/2009 and 2009/2010, although
flowering occurred slightly later in 2009/2010 (Fig. 1).
The flowering period of Type III starts at the end of
October and finishes at the end of November; i.e.,
significantly earlier than those of the other three
groups. E. procerus tended to bloom latest but over-
lapped with that of Type I, and there was no significant
difference between them. The flowering date of Type II
was intermediate between those of Type III and
E. procerus. The mean initial flowering dates of
E. procerus, Type I, Type II, and Type III in
2009/2010 were January 11, January 1, December 17,
and November 10, respectively.
Principal component analysis of the four groups
graphically summarized the phenotypic similarities
and differences among Erianthus individuals (Fig. 2).
The first three principal component axes accounted
for 53.0% of the variation, with component I
accounting for 23.1%, component II accounting for
20.7%, and component III accounting for 9.2%.
Principal component axis I was composed of the
degree of development of buds and root primordia on
the stalk, with the plants that were easily propagated
from stalks being grouped on the left of the axis and
individuals that were difficult to vegetatively propa-
gate grouped on the right of the axis (Table 3). Axis
II was affected by plant size, plant height, inflores-
cence length, leaf width, and stalk diameter and
reflected morphological characteristics. For example,
plant height ranged from 460.7 ± 83.2 cm in Type I
and 398.4 ± 91.4 in Type III, and inflorescence
length ranged from 86.4 ± 14.6 in Type I and
68.6 ± 12.8 in Type III (Table 2). Component III
mainly consisted of rachis joint length and petiole
length but offered no further separation of the
grouping. E. procerus were generally located on the
right in the dimensional scatter diagram (Fig. 2). The
Type III individuals were scattered in the lower left
quadrant, and Type II were located in the upper left
quadrant. Type I was scattered in the region inter-
mediate between these groups and partially over-
lapped with E. procerus.
Canonical discriminant analysis (CDA) and the
plot of 108 clones of E. procerus and three types of E.
arundinaceus on the plane of CAN1 and CAN2
(Fig. 3) revealed significant different among these
four groups (P \ 0.001, wilks’ K). Canonical dis-
criminant analysis (CDA) showed that 92.79% of the
variation was explained by the first two canonical
variables, 50.4 and 41.4%, respectively. In CDA, the
772 Genet Resour Crop Evol (2012) 59:769–781
123
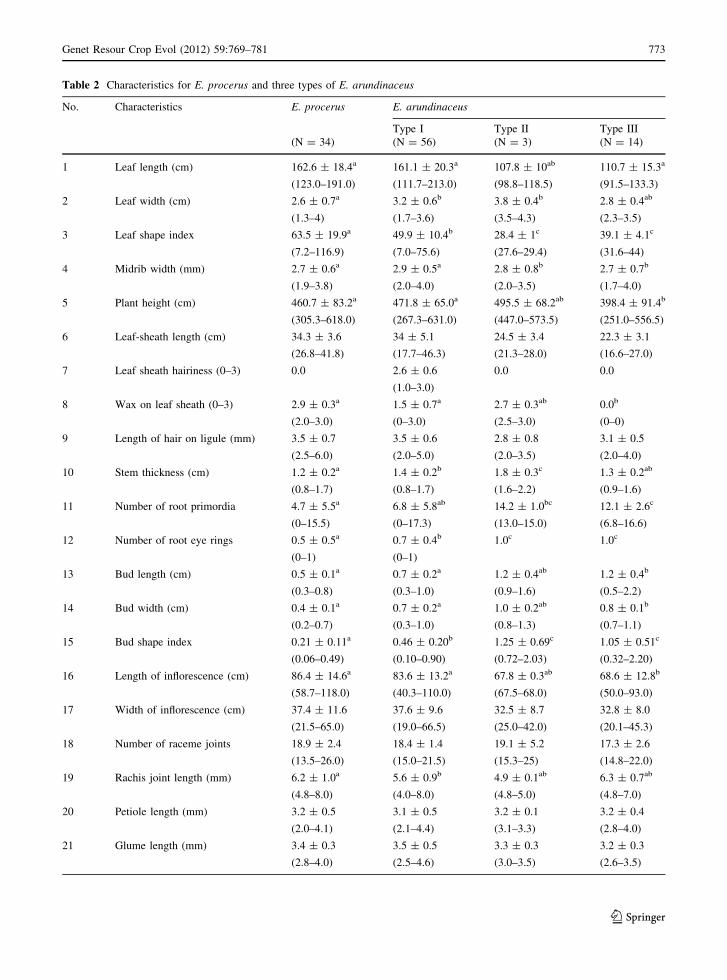
Table 2 Characteristics for E. procerus and three types of E. arundinaceus
No. Characteristics E. procerus E. arundinaceus
Type I Type II Type III
(N = 34) (N = 56) (N = 3) (N = 14)
1 Leaf length (cm) 162.6 ± 18.4a
(123.0–191.0)
161.1 ± 20.3a
(111.7–213.0)
107.8 ± 10ab
(98.8–118.5)
110.7 ± 15.3a
(91.5–133.3)
2 Leaf width (cm) 2.6 ± 0.7a
(1.3–4)
3.2 ± 0.6b
(1.7–3.6)
3.8 ± 0.4b
(3.5–4.3)
2.8 ± 0.4ab
(2.3–3.5)
3 Leaf shape index 63.5 ± 19.9a
(7.2–116.9)
49.9 ± 10.4b
(7.0–75.6)
28.4 ± 1c
(27.6–29.4)
39.1 ± 4.1c
(31.6–44)
4 Midrib width (mm) 2.7 ± 0.6a
(1.9–3.8)
2.9 ± 0.5a
(2.0–4.0)
2.8 ± 0.8b
(2.0–3.5)
2.7 ± 0.7b
(1.7–4.0)
5 Plant height (cm) 460.7 ± 83.2a
(305.3–618.0)
471.8 ± 65.0a
(267.3–631.0)
495.5 ± 68.2ab
(447.0–573.5)
398.4 ± 91.4b
(251.0–556.5)
6 Leaf-sheath length (cm) 34.3 ± 3.6
(26.8–41.8)
34 ± 5.1
(17.7–46.3)
24.5 ± 3.4
(21.3–28.0)
22.3 ± 3.1
(16.6–27.0)
7 Leaf sheath hairiness (0–3) 0.0 2.6 ± 0.6
(1.0–3.0)
0.0 0.0
8 Wax on leaf sheath (0–3) 2.9 ± 0.3a
(2.0–3.0)
1.5 ± 0.7a
(0–3.0)
2.7 ± 0.3ab
(2.5–3.0)
0.0b
(0–0)
9 Length of hair on ligule (mm) 3.5 ± 0.7
(2.5–6.0)
3.5 ± 0.6
(2.0–5.0)
2.8 ± 0.8
(2.0–3.5)
3.1 ± 0.5
(2.0–4.0)
10 Stem thickness (cm) 1.2 ± 0.2a
(0.8–1.7)
1.4 ± 0.2b
(0.8–1.7)
1.8 ± 0.3c
(1.6–2.2)
1.3 ± 0.2ab
(0.9–1.6)
11 Number of root primordia 4.7 ± 5.5a
(0–15.5)
6.8 ± 5.8ab
(0–17.3)
14.2 ± 1.0bc
(13.0–15.0)
12.1 ± 2.6c
(6.8–16.6)
12 Number of root eye rings 0.5 ± 0.5a
(0–1)
0.7 ± 0.4b
(0–1)
1.0c 1.0c
13 Bud length (cm) 0.5 ± 0.1a
(0.3–0.8)
0.7 ± 0.2a
(0.3–1.0)
1.2 ± 0.4ab
(0.9–1.6)
1.2 ± 0.4b
(0.5–2.2)
14 Bud width (cm) 0.4 ± 0.1a
(0.2–0.7)
0.7 ± 0.2a
(0.3–1.0)
1.0 ± 0.2ab
(0.8–1.3)
0.8 ± 0.1b
(0.7–1.1)
15 Bud shape index 0.21 ± 0.11a
(0.06–0.49)
0.46 ± 0.20b
(0.10–0.90)
1.25 ± 0.69c
(0.72–2.03)
1.05 ± 0.51c
(0.32–2.20)
16 Length of inflorescence (cm) 86.4 ± 14.6a
(58.7–118.0)
83.6 ± 13.2a
(40.3–110.0)
67.8 ± 0.3ab
(67.5–68.0)
68.6 ± 12.8b
(50.0–93.0)
17 Width of inflorescence (cm) 37.4 ± 11.6
(21.5–65.0)
37.6 ± 9.6
(19.0–66.5)
32.5 ± 8.7
(25.0–42.0)
32.8 ± 8.0
(20.1–45.3)
18 Number of raceme joints 18.9 ± 2.4
(13.5–26.0)
18.4 ± 1.4
(15.0–21.5)
19.1 ± 5.2
(15.3–25)
17.3 ± 2.6
(14.8–22.0)
19 Rachis joint length (mm) 6.2 ± 1.0a
(4.8–8.0)
5.6 ± 0.9b
(4.0–8.0)
4.9 ± 0.1ab
(4.8–5.0)
6.3 ± 0.7ab
(4.8–7.0)
20 Petiole length (mm) 3.2 ± 0.5
(2.0–4.1)
3.1 ± 0.5
(2.1–4.4)
3.2 ± 0.1
(3.1–3.3)
3.2 ± 0.4
(2.8–4.0)
21 Glume length (mm) 3.4 ± 0.3
(2.8–4.0)
3.5 ± 0.5
(2.5–4.6)
3.3 ± 0.3
(3.0–3.5)
3.2 ± 0.3
(2.6–3.5)
Genet Resour Crop Evol (2012) 59:769–781 773
123

Fig. 1 Flowering
phenology of E. procerusand three types of
E. arundinaceus cultivated
at KKFCRC, Thailand.
Black and white bar show
the collected year, black in
2008/2009 and white in
2009/2010
Table 2 continued
No. Characteristics E. procerus E. arundinaceus
Type I Type II Type III
(N = 34) (N = 56) (N = 3) (N = 14)
22 Flowering days in 09/10 82.9 ± 12.8a
(36–103)
71 ± 12.9a
(34–97)
53.6 ± 4.2b
(49–57)
17.2 ± 9.2c
(5–40)
A one-way ANOVA followed by a post-hoc Turkey test was performed to test for differences among four groups. Different letters
indicate significant differences at P \ 0.05
774 Genet Resour Crop Evol (2012) 59:769–781
123

contribution of each characteristic to a reduced
variable is expressed by its standardized canonical
coefficient (Table 3). In CAN1, the largest absolute
value of a canonical coefficient was leaf sheath
hairiness, indicating that this character contributes
most to CAN1 (Table 3). Its contribution was in a
negative direction. The second largest was flowering
date, bud length, leaf sheath length and glume length,
all of which except bud length contribute in a
negative direction. On the other hand, positive
contributions in CAN2 were by leaf sheath hairiness,
bud width and bud length, and negative contributions
by no. of root eye rings (Table 3). Considering glume
length is not significant among four groups (ANOVA,
P [ 0.052), leaf sheath hairiness, bud length, bud
width and flowering date are considered to be the
most important to identify the four groups. A plot of
the first two canonical variables clearly identified
differences among E. procerus and three types of
E. arundinaceus, with two assumed hybrid somewhat
intermediate regions (Fig. 3).
Discussion
E. procerus versus three types of E. arundinaceus
It is proper to classify Thai Erianthus into four
morphological and ecological distinguishable groups
Fig. 2 PCA scattered plot of E. procerus and three types of
E. arundinaceus based on 22 characteristics (Table 1).
E. procerus (filled square), Type I (open circle), Type II
(filled circle), and Type III (open triangle). Symbols indicated
with an arrow point out assumed hybrids: (multi symbol:ThE02-86, plus symbol: ThE02-91)
Table 3 Eigenvalues of the first three principal component
based on 21 morphological and one ecological characters of
E. procerus and three types of E. arundinaceus
No. Character PC1 PC2 PC3
1 Leaf length 0.306 0.224 -0.047
2 Leaf width -0.118 0.391 -0.004
3 Leaf shape index 0.313 -0.197 -0.090
4 Midrib width -0.033 0.322 -0.090
5 Plant height 0.043 0.358 -0.138
6 Leaf sheath length 0.282 0.196 0.011
7 Leaf sheath hairiness -0.007 0.220 0.234
8 Wax on leaf sheath 0.306 0.030 -0.124
9 Length of hair on ligule 0.070 0.085 -0.071
10 Stem thickness -0.169 0.355 -0.110
11 No. of root primordia -0.274 0.123 -0.203
12 No of root eye rings -0.266 0.097 -0.099
13 Bud length -0.356 -0.008 -0.192
14 Bud width -0.332 0.180 -0.025
15 Bud shape index -0.098 -0.259 -0.276
16 Length of Inflorescence 0.197 0.277 -0.195
17 Width of Inflorescence 0.109 0.143 -0.169
18 No. of raceme joints 0.096 0.184 -0.108
19 Rachis joint length 0.076 -0.175 -0.514
20 Petiole length 0.019 -0.062 -0.552
21 Glume length 0.082 -0.005 -0.245
22 Flowering date 0.346 0.126 0.055
Fig. 3 CDA scattered plot of E. procerus and three types of
E. arundinaceus based on 22 characteristics (Table 1).
E. procerus (filled square), Type I (open circle), Type II
(filled circle), and Type III (open triangle). Symbols indicated
with an arrow point out assumed hybrids: (multi symbol:ThE02-86, plus symbol: ThE02-91)
Genet Resour Crop Evol (2012) 59:769–781 775
123

as used in this study, because the CDA analysis
shows the separation of these four groups (Fig. 3).
The PCA analysis clearly supported the separation
of E. procerus from Type II and Type III of
E. arundinaceus according to their morphological
characteristics particularly bud size, root primordia,
and stalk size, which had a large influence on axis I
(Fig. 2). However, some overlapping was confirmed
between E. procerus and Type I E. arundinaceus in
many of the characteristics that are not particularly
useful for delineating taxa, suggesting a high degree
of overall morphological similarity between E. pro-
cerus and Type I. To recognize the taxonomic
differences between E. procerus and the three
different types of E. arundinaceus, we discuss (1)
the degree of development of bud and root primordia,
(2) rachis joint length, and (3) flowering season, all of
which have previously been used for their classifica-
tion (Mukherjee 1958; Daniels and Roach 1987;
Amalraj and Balasundaram 2006; Chen and Phillips
2006).
First, regarding the degree of development of buds
and root primordia, which Amalraj and Balasundaram
(2006) indicated to be useful taxonomical differenti-
ators, they were useful for discriminating E. procerus
from Type II and Type III of E. arundinaceus. The
former had the smallest mean of bud size and few or
no root primordia on its stalk at a height of one meter
(Table 1), while the latter two displayed larger buds
and greater numbers of root primordia. These traits
were considered to reflect adaptation to their natural
habitats. Type II and III inhabit riverbanks and
streambeds, where flooding occurs frequently in the
rainy season, and can survive during flooding, while
E. procerus is mainly found in areas were floods are
rare, such as sunny hill slopes and forest edges. These
results correspond well to the clear differences
between E. procerus and E. arundinaceus reported
by Amalraj and Balasundaram (2006). However, no
significant differences in these traits were confirmed
between Type I and E. procerus (Table 1), as shown
by the overlapping along axis I on the PCA scatter
plot, indicating that these traits are not available for
distinguishing between the two groups.
Second, concerning rachis joint length, which
many researchers have used to classify E. procerus
and E. arundinaceus (Daniels and Roach 1987; Chen
and Phillips 2006), it could not be effectively applied
to discriminate Thai E. procerus from the other
species since a high degree of overlapping between
E. procerus and the three types of E. arundinaceus
was detected. In fact, standardized canonical coeffi-
cient values of rachis joint length are both low in
CAN1 and CAN2 in CDA analysis (Tables 3, 4). The
wide variation in the flower traits of these species is
one of the problems making their classification
more complex, and it is difficult to identify these
species using flower specimens alone (Mukherjee
1958).
Third, the flowering period of E. procerus was
clearly separated from those of Type II and III
E. arundinaceus, but largely overlapped with that of
Type I E. arundinaceus (Fig. 1). This later flowering
period of E. procerus corresponded to the previous
observation that the flowering of E. arundinaceus
occurs earlier than that of E. procerus, which was
described as a taxonomic differentiators between the
two species. Therefore, this characteristic is useful for
Table 4 Standardized canonical coefficients of the first and
second canonical variables (CAN1 and CAN2, respectively)
for 22 morphological and ecological characters
No. Character CAN1 CAN2
1 Leaf length -0.054 -0.157
2 Leaf width -0.092 -0.147
3 Leaf shape index 0.118 -0.040
4 Midrib width -0.095 -0.281
5 Plant height -0.084 -0.141
6 Leaf sheath length -0.476 0.009
7 Leaf sheath hairiness -2.159 1.259
8 Wax on leaf sheath -0.230 -0.170
9 Length of hair on ligule 0.184 -0.083
10 Stem thickness 0.168 0.051
11 No. of root primordia -0.156 -0.059
12 No of root eye rings -0.229 -1.447
13 Bud length 0.554 0.393
14 Bud width -0.131 0.665
15 Bud shape index -0.038 -0.161
16 Length of Inflorescence -0.081 0.095
17 Width of Inflorescence -0.087 0.094
18 No. of raceme joints 0.083 -0.092
19 Rachis joint length 0.108 0.046
20 Petiole length 0.208 0.006
21 Glume length -0.377 0.029
22 Flowering date -0.959 -0.207
776 Genet Resour Crop Evol (2012) 59:769–781
123

the separation of E. procerus from Type II and III
E. arundinaceus in Thailand, but it was not applica-
ble to the separation of E. procerus and Type I as the
flowering period of E. procerus was slightly later
than that of Type I but most accessions showed
overlapping flowering periods.
Erianthus procerus and Type I E. arundinaceus
showed a high degree of similarity, although they
have different chromosome numbers. It is likely that
introgression between the two groups occurs com-
monly in nature because they occasionally occur
sympatrically. However, in fact, there is difference in
the flowering timing of the two groups, which
contributes to the maintenance of the species divide.
Amalraj et al. (2005) showed that the anthesis of
E. procerus generally occurred in the morning and
that of E. arundinaceus occurred in both the morning
and evening. According to our observations of our
Thai Erianthus collection (unpublished), E. procerus
started blooming at night, 10 p.m., and its pollen
remained viable until the next morning, 8:00 a.m.,
whereas the three types of E. arundinaceus started
blooming soon after sunset, and their pollen was
unable to germinate the next morning. As the
viability of pollen is less than a few hours at RT, in
most cases, it seemed to act as a reproductive barrier
and make natural hybridization difficult, although we
can not completely rule it out.
The differences between E. procerus and
E. arundinaceus have been well studied and have
mainly been described using Indian Erianthus arund-
inaceus (Type III in this study) and Indonesian
E. arundinaceus (Type II in this study), both of which
have received attention as materials that are useful for
sugarcane improvement programs. In a previous study,
which mainly involved samples occurring in India,
China, Indonesia, Japan, which are located at the edge
of the distribution of E. arundinaceus, we found that
the samples showed less variation. Therefore, we
concluded that the taxonomic differentiators men-
tioned in previous studies as being useful for distin-
guishing E. procerus from Type II and Type III are not
effective for differentiating it from Type I because of
the large degree of overlapping in many traits between
them. To discriminate between these two, we suggest
the presence of hairs on the leaf sheath is available
(glabrous in E. procerus vs. pubescent in Type I
Erianthus arundinaceus; Table 2).
Intraspecific variation in E. arundinaceus
In this study, based on their cytological, ecological,
and morphological traits, we defined and examined
three types of E. arundinaceus, Type II and III of
which are considered to represent Indian Erianthus
arundinaceus (the Indian-subcontinent type) and the
Indonesian E. arundinaceus (the Indonesian-New
Guinea type), respectively. The chromosome num-
bers of Types I, II, and III are 2n = 60, 60, and 40,
respectively (Tagane unpublished).
Among the three types, Type II and Type III showed a
high degree of similarity in their morphology, as
mentioned above, and individuals of both types were
located on the left side of the PCA scatter plot (Table 2;
Fig. 2). However, molecular analysis based on rDNA
(Besse et al. 1996) and RFLP (Besse et al. 1997) clearly
supported the separation of E. arundinaceus from
Indonesia versus Indian E. arundinaceus and E. proce-
rus. A possible explanation for this discrepancy with
morphological and genetic similarity is that, although
Type II diverged from Type III in ancient times, they
have evolved convergently to adapt to common niches
such as riverbanks and streambeds. In contrast, although
E. procerus and Type III might have diverged in not so
ancient age, as reflected by molecular markers, they
have evolved differently due to their different habitats,
as reflected by their morphological differences. From
the point of view of biogeography, a biogeographic
boundary is known to exist at around 10 degrees north
latitude, the Isthmus of Kra, where various kinds of flora
and fauna show transitions, for example, species of
Dinochloa (Gramineae), Orania (Palmae), Spatbolobus
(Leguminosae), and Antidesma (Euphorbiaceae) (Baker
et al. 1998; Ridder-Numan 1998). For Thai Erianthus,
this boundary appeared to apply to Type II and Type III.
Geographic isolation is considered to be a major cause
of population differentiation and speciation (Grant
1981; Liao et al. 2007).
Type III was characterized by early flowering, from
the end of October to the end of November, which was
1 month sooner than those of the other three groups
and did not overlap from that of any other group
(Fig. 1). Considering Type III (2n = 40) is the Indian-
subcontinent type, this early flowering corresponds
with the previously described taxonomic differences
between E. arundinaceus and E. procerus in India
(Mukherjee 1958). This difference in phenology is
Genet Resour Crop Evol (2012) 59:769–781 777
123

expected to act as a reproductive isolation mechanism
from other groups when Type III and other types grow
sympatrically. In fact, flowering period differentiation
is known to contribute to genetic differentiation in
grass species (Mizuguti et al. 2004). Thus, Type III is
considered to be genetically divergent from E. proce-
rus and the other two types of E. arundinaceus. PCA
analysis showed that it shares morphological similar-
ities with Type II, but in the Thai samples, Type III was
easily distinguishable from Type II through its chro-
mosome number and the presence of wax on its leaf
sheath (present in Type II vs. absent in Type III).
Type I shows the most phenotypic variation in
many characteristics, but we suggest that the most
useful morphological characteristic for taxonomically
differentiating Type I is the presence of hair on its
leaf sheath (Table 1) and its distribution, which did
not include the Malay peninsula but did include
Indochina (Fig. 4). Hair is sometime easy to drop out
naturally, and some variation in the degree of hair
growth was observed in our study, but we confirmed
that the chromosome number of the less hairy clones
was 2n = 60. Regarding hair, only Type I displayed
hair in this study. However, in our field collection trip
to Nakhon Sri Thammarat province in the south of
Thailand in 2010, where Type II was abundantly
distributed, we found that most Type II individuals
had no hair on their leaf sheathes, as characterized in
this study, but some of them did. Thus, this charac-
teristic is not appropriate for discriminating between
Type I and II, and their distribution should be used
instead. This kind of variation in Type II implies that
high genetic variation is preserved within the region
in which these samples were collected.
Two apparently intermediate clones that were
thought to be hybrids, ThE02-86 and ThE02-91, were
found in Surat Thani and Ranong provinces, which are
near to the Isthmus of Kra. ThE02-86 resembles Type
III; i.e., it has no wax and no hair on its leaf sheath, but
its chromosome number is 2n = 60. ThE02-91,
2n = 60, appeared to be Type I, as it had hair on its
leaf sheath (No. 2 in our criteria), but it was also
characterized by well-developed root primordia and a
large bud size similar to type III. This phenotypic
variation might have been caused by historical
hybridization. It is plausible that once hybrids are
formed, variation is maintained through successive
hybridization among the hybrids and backcrossing of
the hybrids with sympatric parent species. Molecular
analysis based on RAPD markers clarified that
E. arundinaceus (2n = 60) from Andaman–Nicobar
Island, India, where the flora is closely related to that
found on the Malay peninsula, is genetically interme-
diate between the Indian-subcontinent type (Type III)
and Indonesian-New Guinea type (Type II) (Nair and
Mary 2006). Further analysis at population level using
molecular markers will provide us with more detailed
information on this topic.
We classified and examined the morphological
traits and flowering period of E. procerus and three
Fig. 4 Collection site of E. procerus and three types of E. arundinaceus preserved at Khon Kaen Field Crops Research Center,
Thailand
778 Genet Resour Crop Evol (2012) 59:769–781
123

types of E. arundinaceus in Thailand. Since these
groups are polyploid, variation seems to accumulate
easily. Their large chromosome numbers and wide
variation enable them to adapt to various environ-
ments, which is thought to be a good strategy for
E. procerus and E. arundinaceus. Even though some
intermediate types have been found in natural habitats,
we suggest that these four groups were distinctly
supported by various kinds of reproductive isolation
based on habitat preference, geographic distribution,
unsynchronized flowering periods, and time. More
information on populations from other areas covering
a wide range is necessary to adequately support the
taxonomy of the three types of E. arundinaceus.
Based on our morphological, cytological, and
ecological study of Thai Erianthus, we propose that
the following characteristics are key for distinguish-
ing the E. procerus and three types of E. arundin-
aceus. Figure 4 shows a plot of the geographic
locations where the Erianthus germplasms preserved
at KKFCRC were collected, which indicates that all
four groups have somewhat unique geographic dis-
tributions in Thailand.
Key to classifying E. procerus and the three types
of E. arundinaceus
Descriptions of Thai Erianthus procerus and three
types of Erianthus arundinaceus
Erianthus procerus Chromosome number 2n = 40.
Characterized by no hair and much wax on leaf
sheath, and a lack of vegetative clumps; i.e., small
buds and few or no root primordial. Mainly distributed
on hills and mountain areas (collected from relatively
high altitudes, Min–Max and mean were 111–1,617
and 532.3 m above sea level, respectively) in the
north to west of Thailand, but a small population is
known along the Mekong River in Nong Khai and
Bung Karn provinces, northeast Thailand. This pop-
ulation appeared to have arrived from the Lao
mountains, which are located nearby and peak at
around 2,000 m above sea level. In northern part of
Lao P. D. R., where mainly consisted of mountain
ranges, E. procerus is abundantly distributed with
Erianthus longesetosus Andersson and Miscanthus
floridulus (Labill.) Warb. ex K. Schum. et Lauterb.
Type I Chromosome number 2n = 60. Character-
ized by an obviously pubescent leaf sheath. This type
showed a wide variation in its characteristics in
accordance with its adaptation to various environ-
ments, such as open hill slopes, forest edges, and
streambeds, throughout a wide distribution range and
various altitudes (Min–Max and mean were 4–1,760
and 319 m above sea level, respectively). However, it
was not found on the Malay Peninsula. Small
individuals were collected at high altitude in Chiang
Mai province, north Thailand.
Type II Chromosome number 2n = 60. Character-
ized by the presence of vegetative cane; i.e., large
buds capable of germinating and prominent root
primordia, and wax on the leaf sheath. In Thailand, its
distribution only includes the south of Thailand.
Mainly inhabits riverbanks and swamps. Most indi-
viduals do not have hair on their leaf sheathes, but
some do. It is considered to be the Indonesian-New
Guinea type mentioned by Nair and Praneetha
(2006). The Indonesian-New Guinea type is known
to display little morphological or genetic variation in
Indonesia, New Guinea and Philippines. In Thailand,
they appeared to show some variation in their
morphological characteristics.
Type III Chromosome number 2n = 40. Character-
ized by early flowering and the presence of vegetative
1. Leaf sheath pubescence 2
2. Distributed in Indo-China Type I
2*. Distributed in Malay Peninsula, below
the Isthmus of Kra
Type II
1*. Leaf sheath glabrous except at the edge
and around the ligule
3
3. Difficult to propagate vegetatively
because of small buds and few or no
root primordial, grows mainly on in
mountain slopes, at the edge of forests,
and at the side of fields
E.procerus
3*. Easy to propagate vegetatively due to
its well developed buds and root
primordia and mainly occurs in
wetlands such as riverbanks, swamps,
and streambeds
4
4. Distributed in the south of Thailand
below the Isthmus of Kra, wax on the
surface of its leaf sheath
Type II
4*. Distributed in the north, west, and
northeast of Thailand and north of the
Isthmus of Kra and lacks wax on its
leaf sheath
Type III
Genet Resour Crop Evol (2012) 59:769–781 779
123

cane similar to Type II but lacks wax on its leaf
sheath. It is considered to be Indian Erianthus
arundinaceus (Besse et al. 1996) and the Indian
subcontinent type (Nair and Praneetha 2006). In
Thailand, it mainly inhabits the land along rivers and
streams, such as the Mekong River, the tributaries of
the Pin River, and the Mae Klong River etc., but in
the north of the Malay peninsula, we found it along
the roadside, probably due to the short dry season and
high humidity. In Cambodia, this type is common at
lowland area along the Mekong River.
Conclusion
We examined four types of Thai Erianthus clones,
which are consist of two species, E. procerus and E.
arundinaceus in the present taxonomic classification.
The results showed they have wide range of variation
but clearly divergent in their morphology, ploidy,
flowering period and geographic distribution, indi-
cating that Thai Erianthus gemrplasm collection
preserved at KKFCRC provides good materials for
sugarcane breeding. Further evaluation focused on
agronomic traits, such as yield and drought tolerance,
and intergeneric hybridization between sugarcane
commercial varieties and our Erianthus clones, have
been conducted since 2004. So far, 49 hybrids, which
are confirmed by 5SrDNA marker, have been
obtained. They are expected to be interesting breed-
ing materials for sugarcane improvement.
Acknowledgments The authors wish to thank Dr. Suchirat
Sakuanrungsirikul of the Khon Kaen Field Crops Research
Center, Thailand, for providing useful comments on the
manuscript.
References
Amalraj VA, Balasundaram N (2006) On the taxonomy of the
members of ‘Saccharum complex’. Genet Resour Crop
Evol 53:35–41
Amalraj VA, Suganya A, Jebadhas AW (2005) Flowering and
floral biological studies in Erianthus Michx. Sugar Cane
Inter 23:24–27
Arro JA, Veremis JC, Kimbeng CA, Botanga C (2006) Genetic
diversity and relationships revealed by AFLP markers
among Saccharum spontaneum and related species and
Genera. J Am Soc Sugar Cane Technol 26:101–115
Baker WJ, Coode MJE, Dransfield J, Dransfield S, Harley MM,
Hoffmann P, Johns RH (1998) Patterns of distribution of
Malesian vascular plants. In: Hall R, Holloway JD (eds)
Biogeography and geological evolution of SE Asia.
Backbuys Publishers, Leiden, pp 243–258
Besse P, McIntyre CL, Berding N (1996) Ribosomal DNA
variations in Erianthus, a wild sugarcane relative
(Andropogoneae-Saccharinae). Theor Appl Genet 92:
733–743
Besse P, McIntyre CL, Berding N (1997) Characterisation of
Erianthus sect. Ripidium and Saccharum germplasm
(Andropogoneae-Saccharinae) using RFLP markers.
Euphytica 93:283–292
Bor NL (1940) Flora of Assam V. Government of Assam,
Delhi
Cai Q, Aitken KS, Fan YH, Piperidis G, Jackson P, McIntyre
CL (2005) A preliminary assessment of the genetic rela-
tionship between Erianthus rockii and the ‘‘Saccharumcomplex’’ using microsatellite (SSR) and AFLP markers.
Plant Sci 169:976–984
Chen S, Phillips SM (2006) Saccharum Linnaeus. In: Wu Z,
Raven PH (eds) Flora of China. Science Press/Missuri
Botanical Garden Press, Beijing/St. Louis, pp 576–581
Coto O, Cornide MT, Calvo D, Canales E, D’Hont A, de Prada
F (2002) Genetic diversity among wild sugarcane germ-
plasm from Laos revealed with markers. Euphytica
123:121–130
Daniels J, Roach BT (1987) Taxonomy and evolution. In:
Heinz DJ (ed) Sugarcane improvement through breeding.
Elsevier, Amsterdam, pp 7–84
D’Hont A, Rao PS, Feldmann P, Grivet L, Islam-Faridi N,
Taylor P, Glaszmann JC (1995) Identification and char-
acterisation of sugarcane intergeneric hybrids, Saccharumofficinarum 9 Erianthus arundinaceus, with molecular
markers and DNA in situ hybridisation. Theor Appl Genet
91:320–326
Grant V (1981) Plant speciation. Columbia University Press,
New York
Grassl CO (1972) Taxonomy of Saccharum relatives: Sclero-stachya, Narenga and Erianthus. Proc Int Sugar Cane
Technol 14:240–248
Hatusima S (1975) Flora of the Ryukyus. Okinawa Biological
Education and Research Society, Okinawa
Hodkinson TR, Chase MW, Lledo DM, Salamin N, Renvoize
SA (2002) Phylogenetics of Miscanthus, Saccharum and
related genera (Saccharinae, Andropogoneae, Poaceae)
based on DNA sequences from ITS nuclear ribosomal
DNA and plastid trnL intron and trnL-F intergeneric
spacers. J Plant Res 115:381–392
Hole RS (1911) On some Indian forest grasses and their oe-
cology. Indian For Mem For Bot 1:1–127
Hooker JD (1896) Flora British India, vol 7, pp 121–126
Liao PC, Havanond S, Huang S (2007) Phylogeography of
Ceriops tagal (Rhizophoraceae) in Southeast Asia: the
land barrier of the Malay Peninsula has caused population
differentiation between the Indian Ocean and South China
Sea. Conserv Genet 8:89–98
Mizuguti A, Nishiwaki A, Sugimoto Y, Oyamada M (2004)
Genetic difference between two types of Imperata cyl-indrica (L.) Beauv. Characterized by flowering phenol-
ogy. Grassland Sci 50:9–14
Mukherjee SK (1958) Revision of the genus Erianthus. Lloydia
21:157–188
780 Genet Resour Crop Evol (2012) 59:769–781
123

Nair NV, Mary S (2006) RAPD analysis reveals the presence
of mainland Indian and Indonesian forms of Erianthusarundinaceus (Retz.) Jeswiet in the Andaman–Nicobar
Islands. Curr Sci 90:1118–1122
Nair NV, Praneetha M (2006) Cyto-morphological studies on
three Erianthus arundinaceus (Retz.) Jeswiet accessions
from the Andaman-Nicobar Island, India. Cytologia
71:107–109
Piperidis G, Christopher MJ, Carroll BJ, Berding N, D’Hont A
(2000) Molecular contribution to selection of intergeneric
hybrids between sugarcane and the wild species Erianthusarundinaceus. Genome 43:1033–1037
R Development Core Team (2007) A language and environ-
ment for statistical computing. R foundation for statistical
computing, Vienna
Ram B, Sreenivasan TV, Sahi BK, Singh N (2001) Introgres-
sion of low temperature tolerance and red rot resistance
from Erianthus in sugarcane. Euphytica 122:145–153
Ridder-Numan JWA (1998) Historical biogeography of Spa-tholobus (Leguminosae-Papilionoideae) and allies in SE
Asia. In: Hall R, Holloway JD (eds) Biogeography and
geological evolution of SE Asia. Backbuys Publishers,
Leiden, pp 259–277
Rott P, Mohamed IS, Klett P, Soupa D, de Saint-Albin A,
Feldmann P, Letourmy P (1997) Resistance to leaf scald
disease is associated with limited colonization of sugar-
cane and wild relatives by Xanthomonas albilineans.
Phytophatology 87:1203–1213
Selvi A, Nair NV, Balasundaram N, Mohapatra T (2003)
Evaluation of maize microsatellite markers for genetic
diversity analysis and fingerprinting in sugarcane. Gen-
ome 46:394–403
Selvi A, Nair NV, Noyer JL, Singh NK, Balasundaram N,
Bansal KC, Koundal KR, Mohapatra T (2006) AFLP
analysis of the phenetic organization and genetic diversity
in the sugarcane complex, Saccharum and Erianthus.
Genet Resour Crop Evol 53:831–842
Sugimoto A, Ponragdee W, Sansayawichai T, Kawashima T,
Thippayarugs S, Suriyaphan P, Matsuoka M, Lerdpra-
sertrat K, Pramanee P (2002) Collecting and evaluating of
wild relatives of sugarcane as breeding materials of new
type sugarcane cultivars of cattle feed in Northeast
Thailand. JIRCAS Working Rep 30:55–60
Suvatti C (1978) Flora of Thailand. Royal Institute Thailand,
Bangkok
Swarup KP, Kalia SK, Kaul S, Dalal V, Hemaprabha G, Selvi
A, Pandit A, Singh A, Gaikwad K, Sharma TR, Srivastava
PS, Singh NK, Mohapatra T (2009) Informative genomic
microsatellite markers for efficient genotyping applica-
tions in sugarcane. Theor Appl Genet 118:327–338
Williams CA, Harborne JB, Smith P (1974) The taxonomic
significance of leaf flavonoids in Saccharum and related
genera. Phytochemestry 13:1141–1149
Zhang YW, Long HS, Fang YH, Yao YG, Cai Q, Zhang YP
(2002) Sequence variation of rbcL gene and evolution of
Saccharum and relative species. Acta Botanica Yunnanica
24:29–36
Genet Resour Crop Evol (2012) 59:769–781 781
123





























