Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, May 1970, p. 431-439Copyright © 1970 American Society for Microbiology
Vol. 1, No. 5Printed in U.S.A.
Characteristics of Endotoxin-Altering FractionsDerived from Normal Serum
III. Isolation and Properties of Horse Serum a2-MacroglobulinMORIMASA YOSHIOKA AND SEISHI KONNO
The Kitasato Institute, Tokyo 108, and Department of Microbiology, Keio University School of Medicine,Tokyo 160, Japan
Received for publication 15 January 1970
The endotoxin-altering activity of fractions isolated from normal horse serum wasexamined by incubation of Salmonella typhosa strain 0-901 endotoxin (Boivin) in asolution of the fraction, and subsequent quantitation of any diminution in the ca-pacity of endotoxin to be precipitated by specific anti-endotoxin antiserum. The horseserum fraction isolated by precipitation with ammonium sulfate at a concentrationbetween 1.6 and 2.7 M was incubated with Pronase PA and then with trypsin. Whenthis partly digested fraction was passed twice through a Sephadex G-200 column andeluted with 0.2 M tris(hydroxymethyl)aminomethane buffer, most of the endotoxin-altering activity was found in the first protein peak designated F-la. F-la was foundto be homogeneous and corresponded to an a2-macroglobulin by the techniques ofelectrophoresis, immunodiffusion, and ultracentrifugation. Approximately 100-foldmore F-la than endotoxin was needed to reduce the antigenicity of the endotoxinby one-half. Alteration was increased when F-la was incubated with the endotoxinat acid pH or at 45 C rather than at 37 C and was lost after heating F-la at 56 C for30 min. N-ethylmaleimide increased the endotoxin-altering activity of horse serum,F-la, and human plasma fraction 1110, whereas p-chloromercuribenzoate did not.On the other hand, diazonium-l-H-tetrazole, iodoacetic acid, and benzylchloridesuppressed the activity of F-la. When the interaction of endotoxin and F-la wasexamined by immunodiffusion techniques, depolymerization of the endotoxin mole-cule was indicated. The endotoxin-altering factor of horse serum is discussed in re-lation to the mechanisms of other known reagents, such as deoxycholate and so-dium lauryl sulfate.
Human serum or plasma components, whichdecrease in vitro the antigenic and biologicalproperties of bacterial endotoxin, have beenfound in Cohn's fractions 11o and IV1 (21), andin an a1-lipoprotein and a,-globulin esterase (16).Moreover, bile salts present in the plasma havebeen suggested as major active components(12). A variety of reagents other than those ofblood origin, including mild acid hydrolysis,have also been shown to degrade the endotoxinmolecule nonspecifically, e.g., papain (7), bro-melin (22), liver extract (9), dodecyl sulfate (1),acetylation (3, 8), and ionizing radiation (11).Whether identical or diverse chemical sites of theendotoxin molecule are affected by these varioustreatments is not known.An endotoxin-depolymerizing effect of some
of these reagents has been postulated and re-lated to the decrease in serological or biologicalactivity after observations by immunodiffusion
(14, 16), centrifugation (1, 18), and electronmicroscopy (12). Inactivation of endotoxin byhuman serum was reported to be reversible (14),and serological or biological activity of endotoxinwas restored after repolymerization of dispersedsubunits (12).
In a previous publication (22), an endotoxin-altering factor (EAF) from horse serum was ob-tained by fractionation with ammonium sulfate.The activity of EAF was inhibited with the addi-tion of iodoacetic acid, benzyl chloride, 6-iodoso-benzoic acid or guanidine compounds, and apossible role of sulfhydryl groups in EAF waspostulated. To obtain more definitive informa-tion on endotoxin detoxification by EAF, furtherpurification of the active ammonium sulfatefraction by using proteolytic enzyme digestion ofEAF and gel diffusion analysis was carried out.a2-Macroglobulin was found to comprise a
431
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

432 YOSHIOKA
major part of EAF. The characteristics of thisfraction are described.
MATERIALS AND METHODSThe procedure for preparation of endotoxin from a
stock strain of Salmonella typhosa 0-901 with tri-chloroacetic acid extraction and the quantitativeprecipitin reaction, as applied to the analysis of anti-gen (endotoxin), have been described previously(21). In the latter, EAF activity was defined as a de-crease in the per cent of endotoxin precipitable by itshomologous antiserum.Normal horse serum. Sterile, defibrinated, normal
horse serum was obtained from a dealer. The absenceof S. typhosa antibody in the serum was confirmed byagglutination reaction before fractionation.Human plasma fractions. Fractions 11o and IV,
prepared by Cohn's method 10 were kindly suppliedby H. Kawanishi at the Kitasato University, Schoolof Hygienic Sciences, Tokyo.
Preparation of ammonium sulfate fractions. Allof the following procedures were conducted at ap-proximately 4 C. One volume of sterile horse serumwas added to 2 volumes of deionized water, and thesalt concentration was adjusted to 2.7 M with 3.93 Mammonium sulfate solution. The mixture was stirredgently for 2 hr at pH 6.8. After centrifugation at 7,000rev/min for 30 min, the supernatant fluid was precipi-tated by increasing the salt concentration to 3.2 Mwith ammonium sulfate powder. The protein sedi-ment was dissolved in a small amount of 0.007 Mphosphate buffer (PB), pH 7.0; dialyzed in a cellulosetubing against deionized water, 0.004 M PB, and0.007 M PB successively, each for 1 day; lyophilized;and designated fr. A1. The precipitate resulting fromaddition of 2.7 M ammonium sulfate to horse serumwas dissolved in 0.007 M PB, the volume approximat-ing twice that of the original serum, and dialyzed inthe three steps mentioned above. Salt solution wasadded to the material remaining in the dialysis sacuntil a final concentration of 1.6 M ammonium sulfateat pH 8.0 was reached. After stirring gently for 2 hr,the mixture was centrifuged at 7,000 rev/min for 30min. The precipitate was rehydrated, dialyzed, lyo-philized as described above, and designated fr. Bl.To the supernatant fluid was added ammonium sulfatepowder until the concentration reached 2.7 M at pH6.8. It was then gently stirred for 3 hr, centrifuged,dialyzed, lyophilized, and designated fr. B2. In a typ-ical experiment, 19.5 g of fr. Al, 17.0 g of fr. B1,and 21.9 g of fr. B2 were obtained from 1 liter ofhorse serum.Enzyme treatment of fr. B2. Fr. B2 was consistently
found to contain a major part of the EAF present inthe original serum. Accordingly, it was subjected topartial degradation with Pronase and trypsin beforefurther fractionation. Since the two enzymes usedwere shown to inactivate EAF (22), the conditions ofenzyme treatment were altered so as not to give anyappreciable decrease in EAF activity of fr. B2, butstill to achieve limited proteolysis. Towards this end,fr. B2 and Pronase PA (Kaken Kagaku Co., Tokyo)were dissolved in phosphate-buffered saline (PBS;
kND KONNO INFEC. IMMUN.
0.50 g of NaH2PO4, 1.30 g of Na2HPO4, and 8.77g of NaCl per liter; pH 7.0) at a weight ratio of 50to 1. The mixture was incubated in a water bath at37 C for 30 min, and disodium ethylenediamine-tetraacetate (EDTA; Kanto Chemical Co., Tokyo)was added at 0.002 M to suppress the activity of theenzyme. After standing for 10 min at 37 C, the mixturewas cooled to 20 C and adjusted to pH 7.7 with 0.5M borate buffer, pH 7.9. Trypsin, type I (crystallized2X; Sigma Chemical Co., St. Louis, Mo.), dissolvedin distilled water, was then added to the Pronase-treated fr. B2 solution at a dry-weight ratio of 100 to 1.Trypsin treatment was performed in a water bath at20 C for 4 hr, and the fraction was lyophilized imme-diately thereafter. An inhibitor of trypsin was notutilized because the trypsin-soy bean trypsin inhibitorcomplex was eluted with the active protein fractionthrough the Sephadex G-200 column in the fractiona-tion procedure described below. However, trypsinwithout the inhibitor was easily separated and elutedfrom the column.
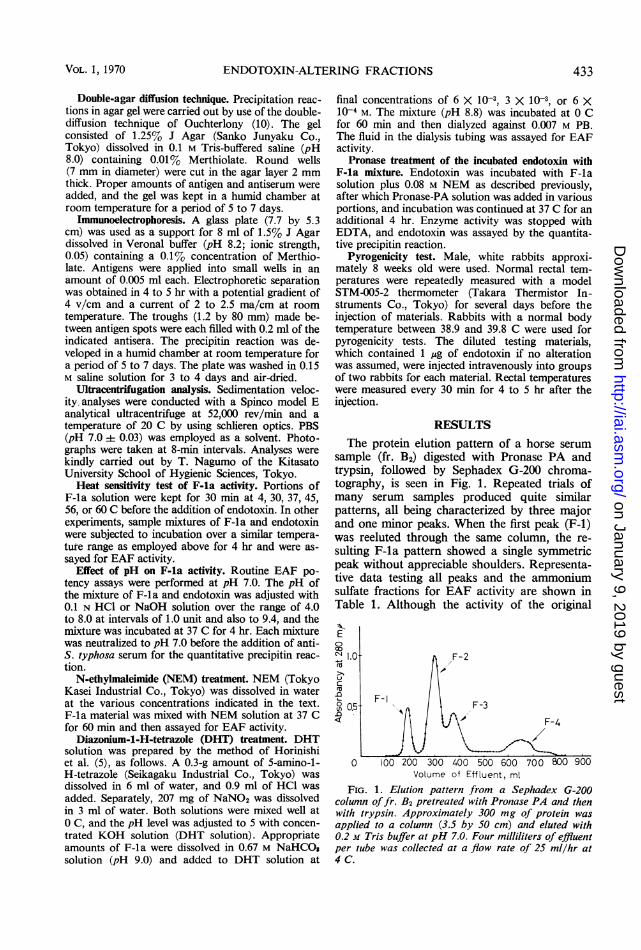
Gel filtration. Sephadex G-200 (Pharmacia, Swe-den) with a particle size of 200 to 400 mesh was used.A column (3.5 by 50 cm or 4.5 by 70 cm) was equil-ibrated with 0.2 M tris (hydroxymethyl)aminomethane(Tris) buffer (pH 7.0) for several hours and chargedwith a 2-ml portion of the enzyme-treated fr. B2,rehydrated to a protein concentration of 7.5%.The buffer was added uniformly by a peristaltic pump(LKB 4912 A, Sweden) at a flow rate of 25 to 33 mlper hr and at a temperature of 4 C. Four-milliliterportions of effluent were collected with a RadiRacfraction collector (LKB 3400 B), and the absorbanceat 280 nm was monitored by a Uvicord II ultra-violet absorptionmeter (LKB 8300 A), fitted with aChopper Bar Recorder (LKB 6520). The EAF activ-ity was found solely in the first protein peak (F-1).F-1 was reeluted through the same column, and themain protein fraction was collected and termed F-la.
Preparation of antiserum. F-la in saline solution,or the original horse serum, was mixed well with anequal amount of incomplete Freund Adjuvant(Difco) by using 20 mg of F-la or 0.5 ml of horseserum, and the mixture was injected subcutaneouslyonce into a foot pad of rabbits. All animals were bled1.5 months later. Sera were separated and frozen at-20 C after Merthiolate was added.
Polyacrylamide gel electrophoresis. A method mod-ified by Tachibana and Yamamura (19) was em-ployed. Twenty milliliters of 10% cyanogum (poly-acrylamide) 41, 0.04 ml of EMAPN (both fromSeikagaku Industrial Co., Tokyo), 25 ml of Tris-citrate buffer (4.65 g of triaminomethane and 0.5 gof citric acid were made to 1 liter with distilled water),40 mg of ammonium persulfate, and 5 ml of deio-nized water were solidified in a container (123 by108 by 3 mm). Samples were applied in small troughs(13 by 1 by 0.5 mm each). One end of the gel layer wasdipped into 4.5 ml of 0.035 M Tris buffer, andthe other into 5.5 ml of 0.05 M boric acid, pH8.4. An electric potential of 150 v and a current of 20ma were applied for 70 min. Proteins were stainedwith 1.0% Amino Black lOB in water-methanol-acetic acid (5:5:1) solution.
* wA
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
ENDOTOXIN-ALTERING FRACTIONS
Double-agar diffusion technique. Precipitation reac-tions in agar gel were carried out by use of the double-diffusion technique of Ouchterlony (10). The gelconsisted of 1.25% J Agar (Sanko Junyaku Co.,Tokyo) dissolved in 0.1 M Tris-buffered saline (pH8.0) containing 0.01% Merthiolate. Round wells(7 mm in diameter) were cut in the agar layer 2 mmthick. Proper amounts of antigen and antiserum wereadded, and the gel was kept in a humid chamber atroom temperature for a period of 5 to 7 days.
Immunoelectrophoresis. A glass plate (7.7 by 5.3cm) was used as a support for 8 ml of 1.5% J Agardissolved in Veronal buffer (pH 8.2; ionic strength,0.05) containing a 0.1% concentration of Merthio-late. Antigens were applied into small wells in anamount of 0.005 ml each. Electrophoretic separationwas obtained in 4 to 5 hr with a potential gradient of4 v/cm and a current of 2 to 2.5 ma/cm at roomtemperature. The troughs (1.2 by 80 mm) made be-tween antigen spots were each filled with 0.2 ml of theindicated antisera. The precipitin reaction was de-veloped in a humid chamber at room temperature fora period of 5 to 7 days. The plate was washed in 0.15M saline solution for 3 to 4 days and air-dried.
Ultracentrifugation analysis. Sedimentation veloc-ity. analyses were conducted with a Spinco model Eanalytical ultracentrifuge at 52,000 rev/min and atemperature of 20 C by using schlieren optics. PBS(pH 7.0 i 0.03) was employed as a solvent. Photo-graphs were taken at 8-min intervals. Analyses werekindly carried out by T. Nagumo of the KitasatoUniversity School of Hygienic Sciences, Tokyo.
Heat sensitivity test of F-la activity. Portions ofF-la solution were kept for 30 min at 4, 30, 37, 45,56, or 60 C before the addition of endotoxin. In otherexperiments, sample mixtures of F-la and endotoxinwere subjected to incubation over a similar tempera-ture range as employed above for 4 hr and were as-sayed for EAF activity.
Effect of pH on F-la activity. Routine EAF po-tency assays were performed at pH 7.0. The pH ofthe mixture of F-la and endotoxin was adjusted with0.1 N HCl or NaOH solution over the range of 4.0to 8.0 at intervals of 1.0 unit and also to 9.4, and themixture was incubated at 37 C for 4 hr. Each mixturewas neutralized to pH 7.0 before the addition of anti-S. typhosa serum for the quantitative precipitin reac-tion.
N-ethylmaleimide (NEM) treatment. NEM (TokyoKasei Industrial Co., Tokyo) was dissolved in waterat the various concentrations indicated in the text.F-la material was mixed with NEM solution at 37 Cfor 60 min and then assayed for EAF activity.
Diazonium-l-H-tetrazole (DHT) treatment. DHTsolution was prepared by the method of Horinishiet al. (5), as follows. A 0.3-g amount of 5-amino-l-H-tetrazole (Seikagaku Industrial Co., Tokyo) wasdissolved in 6 ml of water, and 0.9 ml of HCI wasadded. Separately, 207 mg of NaNO2 was dixsolvedin 3 ml of water. Both solutions were mixed well at0 C, and the pH level was adjusted to 5 with concen-trated KOH solution (DHT solution). Appropriateamounts of F-la were dissolved in 0.67 M NaHCOssolution (pH 9.0) and added to DHT solution at
final concentrations of 6 X 10-3, 3 X 10-3, or 6 X10-4 M. The mixture (pH 8.8) was incubated at 0 Cfor 60 min and then dialyzed against 0.007 M PB.The fluid in the dialysis tubing was assayed for EAFactivity.
Pronase treatment of the incubated endotoxin withF-la mixture. Endotoxin was incubated with F-lasolution plus 0.08 M NEM as described previously,after which Pronase-PA solution was added in variousportions, and incubation was continued at 37 C for anadditional 4 hr. Enzyme activity was stopped withEDTA, and endotoxin was assayed by the quantita-tive precipitin reaction.
Pyrogenicity test. Male, white rabbits approxi-mately 8 weeks old were used. Normal rectal tem-peratures were repeatedly measured with a modelSTM-005-2 thermometer (Takara Thermistor In-struments Co., Tokyo) for several days before theinjection of materials. Rabbits with a normal bodytemperature between 38.9 and 39.8 C were used forpyrogenicity tests. The diluted testing materials,which contained 1 ug of endotoxin if no alterationwas assumed, were injected intravenously into groupsof two rabbits for each material. Rectal temperatureswere measured every 30 min for 4 to 5 hr after theinjection.
RESULTSThe protein elution pattern of a horse serum
sample (fr. B2) digested with Pronase PA andtrypsin, followed by Sephadex G-200 chroma-tography, is seen in Fig. 1. Repeated trials ofmany serum samples produced quite similarpatterns, all being characterized by three majorand one minor peaks. When the first peak (F-1)was reeluted through the same column, the re-sulting F-la pattern showed a single symmetricpeak without appreciable shoulders. Representa-tive data testing all peaks and the ammoniumsulfate fractions for EAF activity are shown inTable 1. Although the activity of the original
E0co" 1.0
(I,%
n
° 0.5-.D
F-4
0 100 200 300 400 500 600 700 800 900Volume of Effluent, ml
FIG. 1. Elution pattern from a Sephadex G-200column offr. B2 pretreated with Pronase PA and thenwith trypsin. Approximately 300 mg of protein wasapplied to a column (3.5 by 50 cm) and eluted with0.2 M Tris buffer at pH 7.0. Four milliliters of effluentper tube was collected at a flow rate of 25 ml/hr at4 C.
VOL. I) 1970 433
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
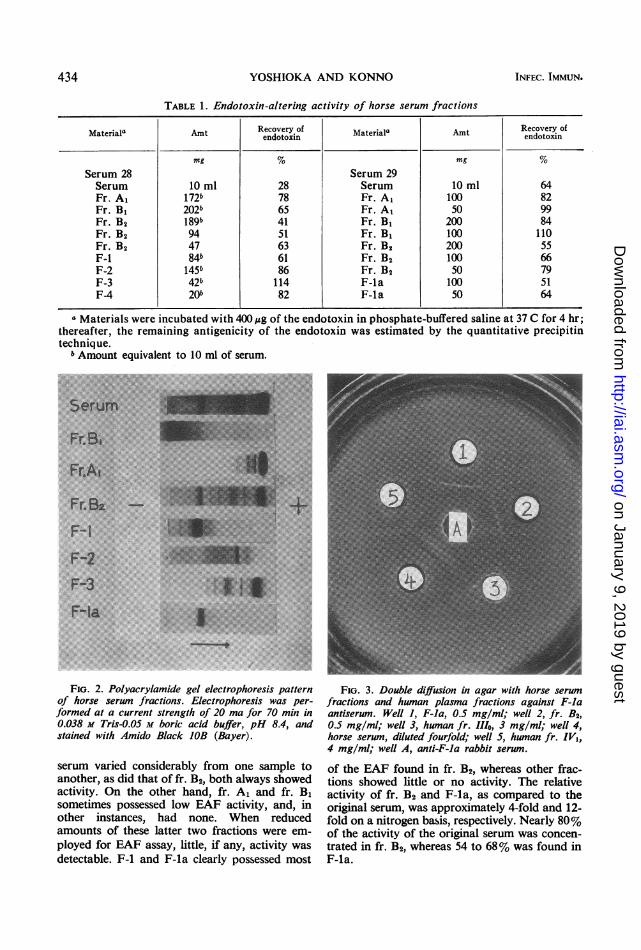
TABLE 1. Endotoxin-altering activity of horse serum fractions
Materiala Amt Recovery of Material" Amt Recovery ofendotoxin Am edovetofi
mg % mg %Serum 28 Serum 29Serum 10 ml 28 Serum 10 ml 64Fr. A1 172b 78 Fr. A, 100 82Fr. B1 202b 65 Fr. A, 50 99Fr. B2 189b 41 Fr. B1 200 84Fr. B2 94 51 Fr. B1 100 110Fr. B2 47 63 Fr. B2 200 55F-i 84b 61 Fr. B2 100 66F-2 145b 86 Fr. B2 50 79F-3 42b 114 F-la 100 51F-4 20b 82 F-la 50 64
a Materials were incubated with 400,ug of the endotoxin in phosphate-buffered saline at 37 C for 4 hr;thereafter, the remaining antigenicity of the endotoxin was estimated by the quantitative precipitintechnique.
b Amount equivalent to 10 ml of serum.~~~~~~~~~~~~~~~~~~~~~~~.......... .. ... .... .-.....FI. 2. P y gel e p pattern
.. .. .. -..... : ;..... .:;.-.. ..... .... ...si.:-;
of horse serum fr s.... .... i ap.e
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.* ...... :.
-r.2..- X i i 2 i-m><$$<ggE;~~~~~~~~~~~~~~~~~~~......
*F.r.B .
*rs : ~~~~~~~~~~~~~~~~~~~~~~~....
stained with Amido Black JOB( )
someime posse lowEA activity an in
...~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~..........
otherinstances, had none When ,reducedc
FIG. 2. Polyacrylamide gel electrophoresis patternof horse sero n fractions.Eler trophoresis was per-formed at a current strength of 20 ma for 70 min in0.038 M Tris-0.05 M boriF acidbuler, pH 8.4, andstained with Amido Black IOB (Bayer).
serum varied considerably from one sample toanother, as did that of fr. B2, both always showedactivity. On the other hand, fr. Al and fr. B,sometimes possessed low EAF activity, and, inother instances, had none. When reducedamounts of these latter two fractions were em-ployed for EAF assay, little, if any, activity wasdetectable. F-1 and F-la clearly possessed most
FIG. 3. Double diffusion in agar withl horse serumfractions and human plasma fractions against F-laantiserum. Well 1, F-la, 0.5 mg/ml; well 2, fr. B2,0.5 mg/ml; well 3, human fr. IIlo, 3 mg/ml; well 4,horse serum, diluted fourfold; well 5, human fr. IV1,4 mg/ml; well A, anti-F-la rabbit serum.
of the EAF found in fr. B2, whereas other frac-tions showed little or no activity. The relativeactivity of fr. B2 and F-la, as compared to theoriginal serum, was approximately 4-fold and 12-fold on a nitrogen basis, respectively. Nearly 80%of the activity of the original serum was concen-trated in fr. B2, whereas 54 to 68% was found inF-la.
434 YOSHIOKA AND KONNO INFEC. IMMUN.
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
ENDOTOXIN-ALTERING FRACTIONS
F-la, stored in a dried state for several monthsat -20 C, exhibited EAF potency equal to thatof freshly prepared samples; its activity began todecline somewhat after 6 months.
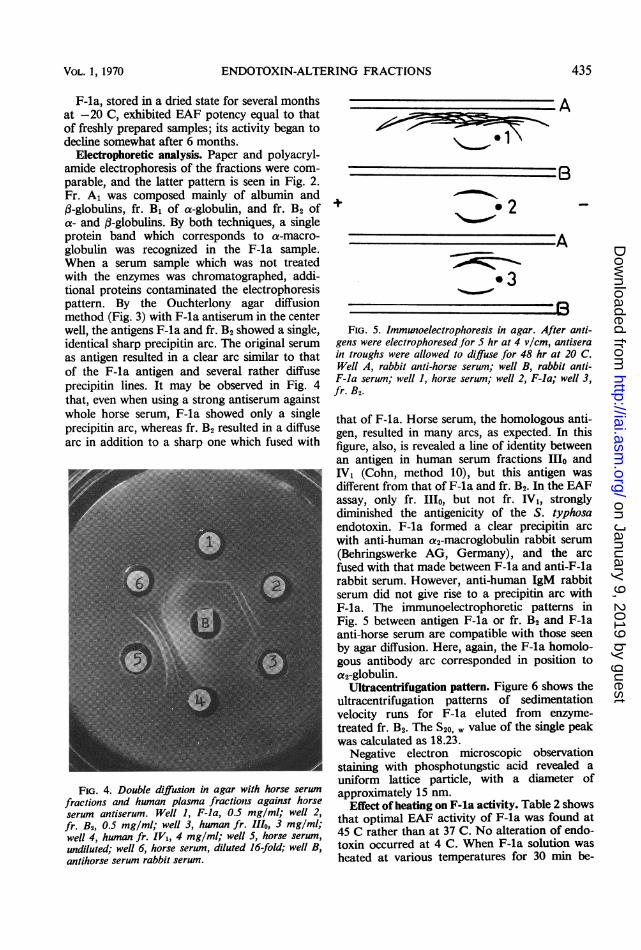
Electrophoretic analysis. Paper and polyacryl-amide electrophoresis of the fractions were com-parable, and the latter pattern is seen in Fig. 2.Fr. A1 was composed mainly of albumin and(3-globulins, fr. B1 of a-globulin, and fr. B2 ofa- and :-globulins. By both techniques, a singleprotein band which corresponds to a-macro-globulin was recognized in the F-la sample.When a serum sample which was not treatedwith the enzymes was chromatographed, addi-tional proteins contaminated the electrophoresispattern. By the Ouchterlony agar diffusionmethod (Fig. 3) with F-la antiserum in the centerwell, the antigens F-la and fr. B2 showed a single,identical sharp precipitin arc. The original serumas antigen resulted in a clear arc similar to thatof the F-la antigen and several rather diffuseprecipitin lines. It may be observed in Fig. 4that, even when using a strong antiserum againstwhole horse serum, F-la showed only a singleprecipitin arc, whereas fr. B2 resulted in a diffusearc in addition to a sharp one which fused with
FIG. 4. Double diffusion in agar with horse serum
fractions and human plasma fractions against horseserum antiserum. Well 1, F-la, 0.5 mg/ml; well 2,fr. B2, 0.5 mg/ml; well 3, human fr. 111o, 3 mg/ml;well 4, human fr. IV1, 4 mg/ml; well 5, horse serum,undiluted; well 6, horse serum, diluted 16-fold; well B,antihorse serum rabbit serum.
A
+ 2
A
*3R
FIG. 5. Immunoelectrophoresis in agar. After anti-gens were electrophoresed for 5 hr at 4 v/cm, antiserain troughs were allowed to diffuse for 48 hr at 20 C.Well A, rabbit anti-horse serum; well B, rabbit anti-F-la serum; well 1, horse serwn; well 2, F-la; well 3,fr. B2.
that of F-la. Horse serum, the homologous anti-gen, resulted in many arcs, as expected. In thisfigure, also, is revealed a line of identity betweenan antigen in human serum fractions 11o andIV, (Cohn, method 10), but this antigen wasdifferent from that of F-la and fr. B2. In the EAFassay, only fr. Illo, but not fr. IV1, stronglydiminished the antigenicity of the S. typhosaendotoxin. F-la formed a clear precipitin arcwith anti-human a2-macroglobulin rabbit serum(Behringswerke AG, Germany), and the arcfused with that made between F-la and anti-F-larabbit serum. However, anti-human IgM rabbitserum did not give rise to a precipitin arc withF-la. The immunoelectrophoretic patterns inFig. 5 between antigen F-la or fr. B2 and F-laanti-horse serum are compatible with those seenby agar diffusion. Here, again, the F-la homolo-gous antibody arc corresponded in position toa2-globulin.
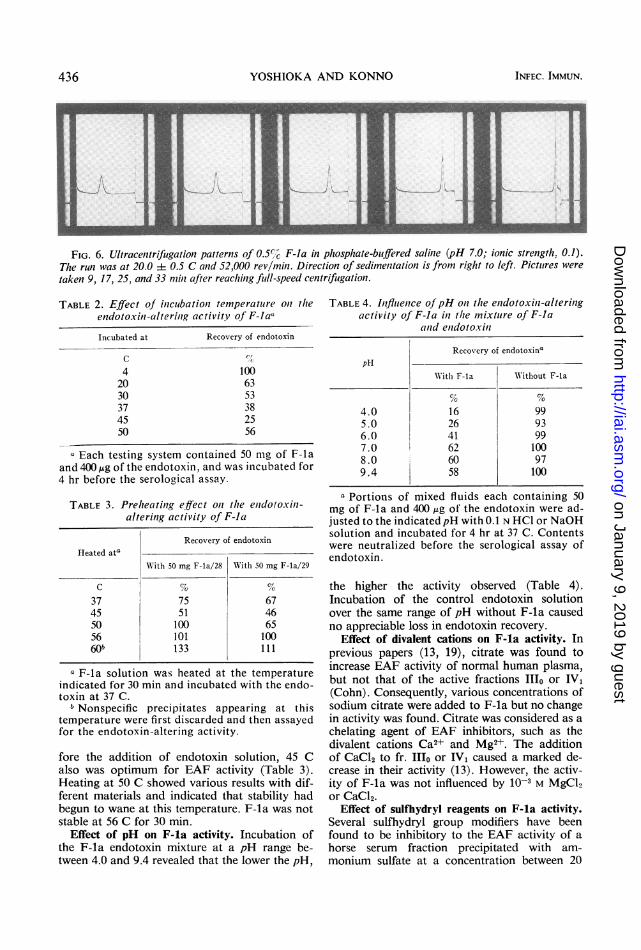
Ultracentrifugation pattern. Figure 6 shows theultracentrifugation patterns of sedimentationvelocity runs for F-la eluted from enzyme-treated fr. B2. The S20, value of the single peakwas calculated as 18.23.
Negative electron microscopic observationstaining with phosphotungstic acid revealed auniform lattice particle, with a diameter ofapproximately 15 nm.
Effect of heating on F-la activity. Table 2 showsthat optimal EAF activity of F-la was found at45 C rather than at 37 C. No alteration of endo-toxin occurred at 4 C. When F-la solution washeated at various temperatures for 30 min be-
435VOL. 1, 1970
;_m
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
YOSHIOKA AND KONNO
FIG. 6. Ultracentrifugation patterns of 0.5%7c F-la in phosphate-buiffered saline (pH 7.0; ionic strength, 0.1).The run was at 20.0 i 0.5 C and 52,000 revlmili. Direction of sedimentation is from riglht to left. Pictutres were
taken 9, 17 25, and 33 miii after reaching full-speed centrifugation.
TABLE 2. Effect of incubationz temperature ont theendotoxin -altering activity of F-laa
Incubated at Recovery of endotoxin
C (7C
4 10020 6330 5337 3845 2550 56
a Each testing system contained 50 mg of F-laand 400,ug of the endotoxin, and was incubated for4 hr before the serological assay.
TABLE 3. Preheatinig effiect onl the enidotoxini-altering activity of F-la
Recovery of endotoxinHeated ata
Wlith 50 mg F-la/28 With 50 mg F-la/29
C % N37 75 6745 51 4650 100 6556 101 10060b 133 111
a F-la solution was heated at the temperatureindicated for 30 min and incubated with the endo-toxin at 37 C.
bNonspecific precipitates appearing at thistemperature were first discarded and then assayedfor the endotoxin-altering activity.
fore the addition of endotoxin solution, 45 Calso was optimum for EAF activity (Table 3).Heating at 50 C showed various results with dif-ferent materials and indicated that stability hadbegun to wane at this temperature. F-la was notstable at 56 C for 30 min.
Effect of pH on F-la activity. Incubation ofthe F-la endotoxin mixture at a pH range be-tween 4.0 and 9.4 revealed that the lower the pH,
TABLE 4. Influence ofpH oni the enidotoxint-alteringactivity of F-la in the mixture of F-Ia
and endotoxin
Recovery of endotoxinapHI
WN'ith F-la WNithout F-la
4.0 16 995.0 26 936.0 41 997.0 62 1008.0 60 979.4 58 100
a Portions of mixed fluids each containing 50mg of F-la and 400 Ag of the endotoxin were ad-justed to the indicatedpH with O.1 N HC1 or NaOHsolution and incubated for 4 hr at 37 C. Contentswere neutralized before the serological assay ofendotoxin.
the higher the activity observed (Table 4).Incubation of the control endotoxin solutionover the same range of pH without F-la causedno appreciable loss in endotoxin recovery.
Effect of divalent cations on F-la activity. Inprevious papers (13, 19), citrate was found toincrease EAF activity of normal human plasma,but not that of the active fractions Ilo or IV,(Cohn). Consequently, various concentrations ofsodium citrate were added to F-la but no changein activity was found. Citrate was considered as achelating agent of EAF inhibitors, such as thedivalent cations Ca2+ and Mg2+. The additionof CaCl2 to fr. IIIo or IV, caused a marked de-crease in their activity (13). However, the activ-ity of F-la was not influenced by 10-3 M MgCI2or CaCl2.
Effect of sulfhydryl reagents on F-la activity.Several sulfhydryl group modifiers have beenfound to be inhibitory to the EAF activity of ahorse serum fraction precipitated with am-monium sulfate at a concentration between 20
436 INFEC. IMMUN.
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
ENDOTOXIN-ALTERING FRACTIONS
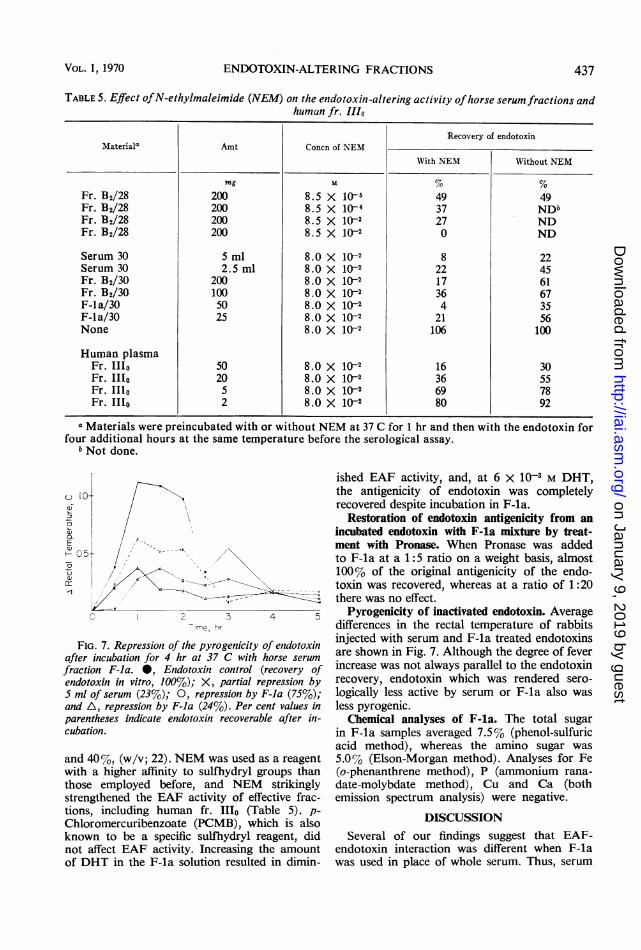
TABLE 5. Effect ofN-ethylmaleimide (NEM) on the endotoxin-altering activity ofhorse serum fractions andhuman fr. 111o
Recovery of endotoxinMateriala Amt Concn of NEM
With NEM Without NEM
mg % %Fr. B2/28 200 8.5 X 10-i 49 49Fr. B2/28 200 8.5 X 1l-4 37 NDbFr. B2/28 200 8.5 X 10-s 27 NDFr. B2/28 200 8.5 X 10-2 0 ND
Serum 30 5 ml 8.0 X 10-2 8 22Serum 30 2.5 ml 8.0 X 10-2 22 45Fr. B2/30 200 8.0 X 10-2 17 61Fr. B2/30 100 8.0 X 10-2 36 67F-la/30 50 8.0 X 10-2 4 35F-la/30 25 8.0 X 10-2 21 56None 8.0 X 10-2 106 100
Human plasmaFr. I1Io 50 8.0 X 10-2 16 30Fr. IIo 20 8.0 X 10-2 36 55Fr. 1Ilo 5 8.0 X 10-2 69 78Fr. 1I1o 2 8.0 X 1(-2 80 92
a Materials were preincubated with or without NEM at 37 C for 1 hr and then with the endotoxin forfour additional hours at the same temperature before the serological assay.
bNot done.
ished EAF activity, and, at 6 X 10-3 M DHT,the antigenicity of endotoxin was completelyrecovered despite incubation in F-la.
Restoration of endotoxin antigenicity from anincubated endotoxin with F-la mixture by treat-
-/4--.s ment with Pronase. When Pronase was addedto F-la at a 1:5 ratio on a weight basis, almost
t\\b \ 100% of the original antigenicity of the endo-toxin was recovered, whereas at a ratio of 1:20
,-';- - - - -~ there was no effect.o 2 3 4 5 Pyrogenicity of inactivated endotoxin. Average
me, hr differences in the rectal temperature of rabbits
[G. 7. Repression of the pyrogenicity of endotoxin injected with serum and F-la treated endotoxinsincubation for 4 hr at 37 C with horse serum are shown in Fig. 7. Although the degree of fever
ion F-la. 0, Endotoxin control (recovery of increase was not always parallel to the endotoxintoxin in vitro, 100%); X, partial repression by recovery, endotoxin which was rendered sero-'of serum (23%); 0, repression by F-la (75%); logically less active by serum or F-la also wasA, repression by F-la (24%). Per cent values in less pyrogenic.ntheses indicate endotoxin recoverable after in- Chemical analyses of F-la. The total sugartion. in F-la samples averaged 7.5% (phenol-sulfuric
acid method), whereas the amino sugar was40%, (w/v; 22). NEM was used as a reagent 5.0% (Elson-Morgan method). Analyses for Fea higher affinity to sulfhydryl groups than (o-phenanthrene method), P (ammonium rana-
e employed before, and NEM strikingly date-molybdate method), Cu and Ca (bothigthened the EAF activity of effective frac- emission spectrum analysis) were negative.
tions, including human fr. IIo (Table 5). p-Chloromercuribenzoate (PCMB), which is alsoknown to be a specific sulfhydryl reagent, didnot affect EAF activity. Increasing the amountof DHT in the F-la solution resulted in dimin-
DISCUSSIONSeveral of our findings suggest that EAF-
endotoxin interaction was different when F-lawas used in place of whole serum. Thus, serum
e
a)
()
H-
~1
Fiafterfractendo,S mlandpare)cubai
andwiththosstren
VOL. 1, 1970 437
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

YOSHIOKA AND KONNO
was more active at alkaline pH, whereas the ac-tivity of F-la was higher at acid pH. Ca or Mgions were highly inhibitory to the endotoxin-altering activity of serum and human fractionsIII and IV1, but they did not-affect the activityof F-la. These differences might have occurredbecause serum also contains an endotoxin-disaggregating factor (2, 20), which increases thepyrogenic or antigenic activity of the endotoxinsbut does not disrupt it into subunits. Self-aggrega-tion among endotoxin particles, however, maynot be too strong, inasmuch as a single additionof citrate or oxalate has been reported to disaggre-gate endotoxins (20).A possible existence of EAF in serum as other
than a2-macroglobulin also has not been ex-cluded. By agar diffusion techniques, F-lareacted with only a single precipitin line eitherwith F-la antiserum or with horse serum anti-serum. On the other hand, diffusion of horseserum against F-la antiserum resulted in atleast four lines. Thus, several antibodies appearto have been produced by trace amounts of serumprotein in the major component of F-la, a2-macroglobulin. The content of these minor com-ponents in F-la would have been too small toproduce visible precipitin lines, but enough waspresent in whole serum to produce clear reactionlines. Another possibility is that F-la was brokendown in vivo during immunization and elicitedseveral antibodies, and the fragments existed inserum as other than F-la and reacted with thoseantibodies. The precipitin line formed betweenhuman plasma fraction IIo or IV1 and F-la anti-serum apparently spurred to that formed by F-la.The fraction 11o here employed was highlyactive as an EAF, but no activity was found infr. IV,. If the EAF in F-la were antigenic andformed the precipitin line with the F-la anti-serum, the EAF in human fr. 11o should not havea component antigenically identical to the F-la.Although F-la-antibody precipitation formed asingle line, there is no proof that the EAF itselfwas the identical substance which produced theprecipitin reaction. Moreover, the EAF alsocould consist of two or more distinct types ofsubstances.
It is questionable whether all the protein in F-laa2-macroglobulin is required for its endotoxin-altering activity. To reduce the antigenicity of theendotoxin by one-half, 100-fold as much F-la,which was approximately 13-fold times as activeon a weight basis as that of the original serum,was needed as that of endotoxin. This ratiocould be lowered to approximately 10-fold withthe addition of NEM to F-la. This ratio shouldreflect the molecular weights of subunits of both
EAF and the endotoxin, carrying the smallestnumber of reactive sites.
Lyophilized F-la, stored at -20 C, was foundstable for approximately 6 months. The F-lasolution lost its activity when heated at 56 Cfor 30 min. Endotoxin alteration was achievedby F-la most efficiently at 45 C rather than at37 C. Our previous observation (21) using humanplasma fr. 11o and fr. IV1, showed the most effec-tive temperature to be 37 C, but Skarnes et al.(17) found that inactivation of Serratia marces-cens endotoxin with serum at 45 C was as com-plete as at 37 C, when examined for capacity tocause tumor damage in mice.
Endotoxins once inactivated with humanplasma fr. IV1 (14), papain (15), or rabbit liverproteins (9) have been reactivated. These factssuggest that the alteration of endotoxin is notbased on hydrolytic cleavage. Recently, Ribi et al.(12) presented evidence that deoxycholate (NaD),a surfactant, depolymerized the endotoxin mole-cule and reduced molecular size from a completeantigen of about 1,000,000 to a postulated in-complete antigen with a molecular weight of10,000 to 20,000. They hypothesized that, oncedepolymerized molecules of endotoxin werereleased from NaD, prompt reaggregationoccurred, as was shown by restoration of pyro-genicity. When a small amount of human plasmawas added after the NaD treatment, depoly-merized endotoxin molecules did not reaggre-gate even though NaD was removed by dialysis.If, on the other hand, the adsorbed humanplasma was removed by digestion with Pronase,aggregation reoccurred. Similar findings werereported by Oroszlan et al. (9), who used sodiumlauryl sulfate, another detergent, for the disso-ciation of S. marcescens endotoxin, and by Beeret al. (1) who used sodium dodecyl sulfate andEscherichia coli endotoxin. In the present work,approximately 100% of the endotoxin activitywas regained after digestion with Pronase of aninactivated mixture of F-la and endotoxin bythe procedure employed by Rudbach and John-son (14). This fact suggests that an analogousmechanism was functional with F-la. However,deoxycholates were not detected in F-la by thethin-layer chromatography method of Froscheand Wagener (4). Slight amounts of two uniden-tified bile salts and cholesterol were detected.However, dried samples of these three compo-nents recovered from an F-la sample appearednot to have any activity on endotoxin.
Sulfhydryl inhibitors, o-iodosobenzoic acid,iodoacetic acid, benzyl chloride, and guanidinesalts showed a marked degree of suppression ofEAF potency in the horse serum fraction ob-tained by the salting-out method (22). NEM is
438 INFEC. IMMUN.
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

ENDOTOXIN-ALTERING FRACTIONS
known to be a specific sulfhydryl inhibitor, asis PCMB, although the specificity of NEM ishigher (6). PCMB had no effect on the endotoxin-altering activity of F-la, whereas treatment ofthe original horse serum, fr. B2, F-la, or humanfr. 11o (Cohn) with NEM before the addition ofthe endotoxin uniformly and strikingly increasedthe EAF potency. Since use of cysteine hydro-chloride, a sulfhydrylamino acid, or NEM alonedid not give rise to alteration in the antigenicityof endotoxin, an interaction between free sulf-hydryl groups in the protein molecule and sulf-hydryl modifiers still remains speculative. Inaddition, the endotoxin-altering activity of F-lawas highly suppressed with DHT, indicating thatthe histidine residues of F-la were modified.Analysis of active end groups of a2-macroglobu-lin fragments may lead to a closer understandingof the mechanism of EAF-endotoxin interaction.
ACKNOWLEDGMENT
This investigation was supported by Public Health Servicegrant AI-04283 from the National Institute of Allergy and In-fectious Diseases.
LITERATURE CITED
1. Beer, H., T. Staehelin, D. Herdon, and I. B. Abraham. 1965.Relationship between particle size and biological activityof E. coli Boivin endotoxin. J. Clin. Invest. 44:592-602.
2. Cluff, L. E., and I. L. Bennet, Jr. 1957. Factors influencingthe alteration of the pyrogenic action of endotoxin by serum.
Bull. Johns Hopkins Hosp. 101:281-291.3. Freedman, H. H., and B. M. Sultzer. 1962. Dissociation of
the biological properties of bacterial endotoxin by chemicalmodification of the molecule. J. Exp. Med. 116:929-942.
4. Frosche, B., and H. Wagener. 1967. Quantitative determina-tion of conjugated bile acids in serum in acute hepatitis.Nature 213:404-405.
5. Horinishi, H., K. Kurihara, and K. Shibata. 1965. States ofamino acid residues in proteins. VI. Heme-linked histidineresidues in cytochrome C as determined with diazonium-1-H-tetrazole. Biochim. Biophys. Acta 111:520-528.
6. Katz, A. M., and W. F. H. M. Mommaertes. 1962. The sulf-
hydryl groups of actin. Biochim. Biophys. Acta 65:82-92.
7. Kim, Y. B., and D. W. Watson. 1964. Inactivation of gram-negative bacterial endotoxins by papain. Proc. Soc. Exp.Biol. Med. 115:140-142.
439
8. Martin, W. J., and S. Marcus. 1966. Detoxified bacterial
endotoxin. I. Preparation and biological properties of an
acetylated crude endotoxin from Salmonella typhimuriumJ. Bacteriol. 91:1453-1459.
9. Oroszlan, S. I., P. T. Mora, and M. J. Shear. 1963. Reversibleinactivation of an endotoxin by intracellular protein. Bio-chem. Pharmacol. 12:1131-1146.
10. Ouchterlony, 0. 1948. In vitro method for testing the toxin-producing capacity of diphtheria bacteria. Acta Pathol.Microbiol. Scand. 25:186-191.
11. Previte, J. J., Y. Chang, and H. M. El-Bisi. 1967. Detoxifi-cation of Salmonella typhimurium lipopolysaccharide byionizing radiation. J. Bacteriol. 93:1607-1614.
12. Ribi, E., R. L. Anacker, R. Brown, W. T. Haskins, B. Malm-gren, K. C. Milner, and J. A. Rudbach. 1966. Reaction ofendotoxin and surfactants. I. Physical and biological prop-
erties of endotoxin treated with sodium deoxycholate. J.Bacteriol. 92:1493-1509.
13. Rosen, F. S., R. C. Skarnes, M. Landy, and M. J. Shear.1958. Inactivation of endotoxin by a humoral component.
III. Role of divalent cation and a dialyzable component.
J. Exp. Med. 108:701-711.14. Rudbach, J. A., and A. G. Johnson. 1966. Alteration and
restoration of endotoxin activity after complexing withplasma proteins. J. Bacteriol. 92:892-898.
15. Rudbach, J. A., E. Ribi, and K. C. Milner. 1965. Reactiva-tion of papain-treated endotoxin. Proc. Soc. Exp. Biol.Med. 119:115-118.
16. Skarnes, R. C. 1966. The inactivation of endotoxin after in-teraction with certain proteins of normal serum. Ann.N.Y. Acad. Sci. 133:644-662.
17. Skarnes, R. C., F. S. Rosen, M. J. Shear, and M. Landy.1958. Inactivation of endotoxin by a humoral component.II. Interaction of endotoxin with serum and plasma. J.Exp. Med. 108:685-699.
18. Stauch, J. E., and A. G. Johnson. 1959. The alteration of bac-terial endotoxins by human and rabbit serum. J. Immunol.82:252-263.
19. Tachibana, T., and K. Yamamura. 1965. The thin layer elec-trophoresis. The use of synthetic polyacrylamide gel as a
supporting medium (in Japanese). SABCO J. 1:15-20.20. Yoshioka, M., M. Hamaska, Y. Saito, and S. Konno. 1963.
Characteristics of endotoxin altering fractions derived fromnormal serum. I. Separation of beta lipoproteins and lipidfrom serum fractions. Kitasato Arch. Exp. Med. 36:27-36.
21. Yoshioka, M., and A. G. Johnson. 1962. Characteristics ofendotoxin altering fractions derived from normal human
serum. J. Immunol. 89:326-335.22. Yoshioka, M., S. Konno, and Y. Saito. 1965. Characteristics
of endotoxin altering fraction derived from normal serum.
II. Preliminary examination of horse serum fractions.
Kitasato Arch. Exp. Med. 38:1-5.
VOL. 1, 1970
on January 9, 2019 by guesthttp://iai.asm
.org/D
ownloaded from





























