Embed Size (px)
Citation preview

THE ~JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 251, No. 3, Issue of February 10, pp. 889-892. 1976
Printed in U.S.A.
Regulation of Carbohydrate Uptake and Adenylate Cyclase Activity Mediated by the Enzymes II of the Phosphoenolpyruvate:Sugar Phosphotransferase System in Escherichia coli*
(Received fhr publication, July 21, 1975)
MILTON H. SAIER, JR.,* BRIGITTE U. FEUCHT, AND LAURA J. HOFSTADTER
From the Department of Biology, The John Muir College, The University of California at San Diego, La Jolla, California 92037
The uptake of various carbohydrates and the synthesis of adenosine 3’:5’-monophosphate (cyclic AMP) are subject to inhibition by sugar substrates of the phosphoenolpyruvate:sugar phosphotransferase system in Escherichia coli. The induced synthesis of the sugar-specific components of the phosphotrans- ferase system was studied and correlated with the induction of regulatory interactions controlling glycerol uptake and net cyclic AMP synthesis. Activities of the Enzyme II complexes specific for glucose, fructose, and mannitol were measured both in uivo and in vitro. These activities were induced 8- to 40-fold by growth in the presence of the appropriate substrate-inducers. Cross inducer specificities were noted. Maximal inhibition of glycerol uptake and cyclic AMP synthesis by a sugar substrate of the phosphotransferase system required induction of the Enzyme II complex specific for that sugar and was abolished by mutations which destroyed Enzyme II activity. The inducer specificities of the regulatory systems and of the Enzymes II were found to be the same. A mutation which depressed the cellular activity of Enzyme I of the phosphotransferase system did not relieve sensitivity to inhibition. The results suggest that adenylate cyclase and several carbohydrate permeases are subject to coordinate regulation by a mechanism which depends on the catalytic activities of the protein components of the
phosphotransferase system.
Sugars appear to enter bacterial cells by several distinct mechanisms. Glucose, mannose, fructose, and mannitol, for example, cross the Escherichia coli membrane by a mechanism catalyzed by the phosphoenolpyruvate:sugar phosphotransfer- ase system (1). These sugars are taken up from the medium and released as sugar phosphates in the cytoplasm. The reactions catalyzed by the PTS’ are as follows:
phosphoenolpyruvate + HPr , Enzyme 1 p L - HPr + pyruvate
Enzyme II, P - HPr + wwoutl Y Comp,ex Sugar-P,,, , + HPr
Enzyme I and HPr are the general proteins, common to all sugars phosphorylated by the system, while each of the Enzyme II complexes, present in the membrane, binds and transports only one or a few sugars. Disaccharides such as lactose, melibiose, and maltose enter E. coli cells via active transport systems which can accumulate the unaltered sub-
* This study was supported by Grant GB37167 from the National Science Foundation, Grant IN-93C from the American Cancer Society, and a generous gift to M.H.S. from the Eli Lilly Pharmaceutical Corp.
$ Research Career Development Awardee, 1 K04 CA00138-01. ‘The abbreviations used are: PTS, phosphoenolpyruvate:sugar
phosphotransf’erase system; cyclic AMP, adenosine 3’:5’-monophos- phate.
strates within the cell (2, 3). Still another carbohydrate, glycerol, appears to enter the E. coli cell by facilitated diffusion, a carrier-mediated process which does not involve energy expenditure (3).
Work from several laboratories has provided evidence for specific permease interactions (4-9). Winkler and Wilson (6) reported that glucosides indirectly inhibited the activity of the lactose permease, although a reciprocal effect of galactosides on glucose permease function was not observed. These interac- tions were studied in detail by Koch and his co-workers (7), who showed that glucose permease substrates inhibited entry and accelerated exit of galactosides, provided that the per-
meases for both substrates were induced to high levels. The concentration of methyl a-glucoside which half-inhibited ga- lactoside uptake was lower than the concentration which supported half-maximal uptake of the glucoside. These results
led to the suggestion that glucose interacted with a cell constituent on the external side of the membrane surface to alter lactose permease function.
In a recent study we described the regulation of permease activities in mutant strains of Salmonella typhimurium defi- cient for components of the PTS (10, 11).2 The melibiose,
*M. H. Saier and S. Roseman, manuscripts in preparation.
883
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

884 Regulation of Carbohydrate Metabolism in E. coli
maltose, and glycerol transport systems were shown to be sensitive to inhibition by glucose and other sugar substrates of the PTS, provided that the cells were deficient for one of the energy coupling proteins of the PTS, Enzyme I or HPr. Several classes of mutations abolished sensitivity of the permeases to regulation.
1. Loss of a specific Enzyme II of the PTS rendered all transport systems resistant to regulation by substrates of that Enzyme II, but not by other sugars.
2. Mutation in a gene which mapped adjacent to the pts operon, the err gene, rendered all transport systems resistant to regulation by all sugars.
3. A third class of mutations abolished regulation of a specific permease by all inhibiting sugars. Recently we have found that this last class of mutations maps within, or very near, the genes coding for the affected permeases. Thus, the permease systems may be allosteric proteins, and the last class
of mutations may alter the transport proteins so that they are uncoupled from allosteric regulation.
The present report extends these observations to E. coli. In this organism, regulation of the activities of several permeases by the PTS can be observed, although in contrast to Salmonella, regulation appears to require the induction of the Enzyme II specific for the inhibiting sugar. It is also noted that when two non-PTS permeases are present, regulation of the activity of one by a substrate of the other may occur.
Cellular levels of cyclic AMP appear to regulate several bacterial physiological processes such as catabolic enzyme synthesis, cellular motility, plasmid replication, and growth (12215). In E. coli, cyclic AMP is synthesized by a membrane- associated adenylate cyclase (EC 4.6.1.1) (16), degraded by a cyclic AMP phosphodiesterase (EC 3.1.4.17) (17), and trans- ported across the cell membrane (18, 19). The rates at which these three processes occur determine the intracellular concen-
tration of the nucleotide. There is evidence that each of these processes is subject to regulation by sugars. Glucose stimulates efflux of cyclic AMP from the cell (18, 19) and inhibits net
production of cyclic AMP (18, 20, 21) while metabolites of glucose have been reported to enhance the activity of the phosphodiesterase in cell-free extracts (22). The mechanisms by which regulation is effected are not known.
In the present communication the relationships between the proteins which catalyze sugar transport and the regulatory systems controlling adenylate cyclase and carbohydrate per- mease activities are studied in uiuo. We show that the cellular concentrations of the protein components of the PTS deter- mine the sensitivities of these enzyme systems to regulation. Maximal inhibition by a sugar substrate of the PTS requires
the presence of high levels of the corresponding Enzyme II complex as shown by experiments in which Enzyme II levels
are manipulated by induction and mutation. Preliminary accounts of these results have appeared (23-25).
EXPERIMENTAL PROCEDURES
Materials-Radioactive sugars were purchased from New England Nuclear Corp. with the following exceptions. I-[“H]Melibiitol was synthesized by reduction of melibiose (Calbiochem) with [3H]sodium borohydride followed by paper chromatographic purification employ- mg butanol/pyridine/water (10/3/3) as solvent (26). 1. [“C]Maltose was from Calatomic and [“C]methyl wglucoside was from Amersham. Nonradioactive sugars, of the n-configuration, were purchased from Calbiochem. Methyl a-glucoside was purified free of glucose by passage through a column of AG2-X8 (OH-) resin in carbonate-free water (27). The product was subsequently crystallized from aqueous ethanol. Cyclic AMP was obtained from Sigma Chemical Corp. and
cyclic [‘HIAMP was from New England Nuclear Corp. Other chemi- cals were of the highest purity available commercially.
Growth of Bacteria-The minimal salts medium employed, a modified Medium 63 (ll), consisted of 0.05 M potassium phosphate, 2 g of ammonium sulfate per liter, and 0.2 g of MgSO,.7H,O per liter. This medium was supplemented with a carbon source and methionine when required. Bacto-agar at 1.5% was included for solid media. The complex medium used was bacto-nutrient broth. Fermentation was estimated employing eosin methylene blue agar medium without lactose (Baltimore Biological Corp.) supplemented with the sugar to be tested at 1%.
Bacterial Strains-Escherichia coli strains used in the present study are listed in Table I. Strain AB257ec-‘, a cyclic AMP phosphodiester- ase-deficient mutant, and the parental E. coli K 1Z strain, AB257, were obtained from Dr. H. V. Rickenberg (17). Both strains required methionine for growth. They were used in the present study so that the regulation of permease function could be correlated with the regulation of adenylate cyclase. Other strains listed in Table I were derived from AB257pc-‘.
The Enzyme I-deficient mutants, ptsIl09 and pts1302, were isolated as mannitol fermentation-negative strains after mutagenesis with 2.aminopurine as described previously (11). The former mutant grew slowly with mannitol as sole source of carbon and therefore presumably possessed low residual Enzyme I activity in uiuo; the latter mutant appeared to lack Enzyme I by the same criterion. Enzyme assays revealed that both strains contained less than 1% of wild type Enzyme I activity, but normal activities of the HPr and Enzyme II’;ic proteins (1). While ptsll09 could grow with glycerol, lactose, or maltose as the sole source of carbon, ptsI302 could utilize none of these compounds for growth because the corresponding catabolic enzymes were not induced. These two strains therefore resemble “leaky” and “tight” Enzyme I mutants of Salmonella typhimurium respectively in their growth and enzyme induction properties (10, 11).
The manA mutant and three other independently isolated mutant strains of the same phenotype were isolated as 2.deoxyglucose-resist- ant strains as follows: 3 drops of a nutrient broth culture of AB257~‘.’ were spread on solid media containing in Medium 63: 0.2% glycerol, 0.1% 2.deoxyglucose, 20 pg of methionine per ml, and 1.5% Bacto-agar. Strain AB257”‘-’ and other E. coli and S. typhimurium strains tested could not grow on this medium. When the plates had dried, a few crystals of N-methyl-l\i’-nitro-N-nitrosoguanidine were placed on the agar surface. After 3 days at 37” mutants capable of growth on this medium appeared. These were clonally isolated and tested for their ability to ferment mannose on eosin methylene blue (EMB) agar media containing the sugar at 1%. Four fermentation-negative strains were independently isolated, all of which had the properties described below for manA.51. They lost the ability to ferment and utilize mannose and glucosamine, but could utilize other sugars for growth at normal rates (Table II). Slow utilization of mannose and glucosamine by the parental strain when not previously grown in glucose-containing media (Table II) reflected the fact that glucose, but not mannose or glucosamine, was an effective inducer of the Enzyme II complex which phosphorylates these three sugars.s Transport experiments showed that the manA mutation prevented the intracellular accumulation of radioactive metabolites from [“Clmannose and [“Clglucosamine, and in vitro enzyme analyses (28) revealed a 90% reduction in the Enzyme II activity responsible for mannose and glucosamine phosphorylation. Enzyme I and HPr activities were normal as were the Enzyme II activities for fructose and mannitol. Similar mutants have been isolated by alternative procedures (29).2
Strains which were deficient for lactose permease or j3-galactosidase were isolated as lactose fermentation-negative strains after 2.aminopu- rine mutagenesis and penicillin screening in the presence of 0.2% lactose and 20 rg/ml of methionine (10). The strains were character- ized as follows: all strains were specifically defective for lactose fermentation. They were grown in Medium 63 containing 0.5% glycerol, 20 pg/ml of methionine, and 0.5 rn~ isopropyl a-thiogalacto- side for 4 hours to induce expression of the lactose operon. With the cell density at 10 klett units, hydrolysis of 0-nitrophenyl /3-galactoside was measured in the absence or presence of sufficient toluene to disrupt the cell membrane (30). Toluene treatment was as follows: the cell suspensions (1 ml) were shaken vigorously with 2 drops of toluene and incubated at 37” for 15 min before addition of 0-nitrophenyl P-galacto- side to a final concentration of 2.5 mu. After an appropriate incubation period at 37”, 2 ml of a solution containing 0.2 M Na,CO, and 10 rn~
’ Unpublished observations.
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

Regulation of Carbohydrate Metabolism in E. coli 885
TABLE I
Bacterial strains
All strains listed were derived from strain AB257~‘-’ (17) and are auxotrophic for methionine. Genetic designations are in accord with recommendations of Lin (3).
Strain No. Strain Designation Defect Parent
SB2252 AB257~‘-’
SB2260 ptsIl09 SB2876 ptsll09 err-27
LJ43 p tsI302 LJ34 manA L<J37 lacZ3001
L<J46 lac I’3002
L<J39 lac YZ3003
Cyclic AMP phosphodies- terase
Enzyme I of PTS (leaky) Carbohydrate repression-
resistant Enzyme I ofPTS (tight) Enzyme II”“” of PTS &Galactosidase Lactose permease p-Galactosidase and Lac-
tose permease
AB257
AB257P’-’ ptsIl09
AB257”‘-’ AB25?‘“-’ AB35?‘-’ AB2,57P’-’ AB25?‘-’
TABLE II
Growth properties of Escher&& coli strains
Cells were grown in Medium 63 supplemented with 20 pg of methionine/ml and either 1% m-lactate or 0.5Y glucose as indicated below. These were harvested, washed with Medium 6~3, and aliquots were used to inoculate 20 ml of Medium 63 containing 20 /~g of methionine/ml plus 0.5% of the carbon source indicated in 1%ml Erlenmeyer flasks fitted with side arms for measurement of turbidity. Growth was conducted at 37” with flasks rotating at 250 rpm (II). 2.Deoxyglucose, when present, was at a concentration of O.l’/o.
Generation time
Glycerol + 2-deoxyglucose
Glucose Mannose Glucosamine Mannitol Fructose
-0
1.2 2.4
3.0 1.3 1.4
hr
3c
1.0 1.2 1.5
1.3
1.1 zc z 1.3 1.3
’ 3c, generation time in excess of 10 hours.
EDTA were added, and the yellow color, due to 0-nitrophenol, was measured at 410 nm. None of the lactose-negative mutants hydrolyzed the galactoside in the absence of toluene, and only LJ46 and the parental strain released 0-nitrophenol in the presence of toluene. AB257p’-’ and LJ37, but not LJ39 or LJ46, could accumulate [l’C]iso- propyl 8.thiogalactoside against a concentration gradient. These results showed that LJ37 was defective for @-galactosidase, LJ46 was defective for the lactose permease, while LJ39 appeared to lack both proteins.
Uptake of Radioactive Sugars and Enzyme Assays-Uptake of radioactive sugars was followed as a function of time at 32” or 28” as described previously (10). Specific conditions are given in the legends to the tables and figures. Assays for the protein components of the phosphotransferase system were also as described (11) except for the experiments reported in Table IV which were conducted as described therein.
Measurement of Cyclic AMP Production-Cyclic AMP was mea- sured in the total cell suspensions (cells plus medium) as follows. Following appropriate incubations at 37”, test tubes containing 1 or 2 ml of the cell suspensions were placed in a boiling water bath for 5 min. Subsequently, the suspensions were cooled, cells were removed by centrifugation, and the clear supernatant was then adjusted to pH 4 with 4 N HCl. Cyclic AMP was routinely determined by the method of Gilman with the cyclic AMP binding protein purified from fresh bovine
skeletal muscle through the DEAE-cellulose column procedure (31). All extracts were assayed at two concentrations. The two values were averaged and compared with those for standard cyclic AMP. Several controls in addition to those described by other investigators (31) provided evidence that the material measured was, in fact, cyclic AMP.
1. The barium-zinc reagents, which precipitated most nucleotides but not cyclic AMP (32), left about 95% of the active material in the supernatant.
2. The active compound was lost by dialysis or by treatment with cyclic AMP phosphodiesterase.
3. Little or no active material was produced by E. coli 01‘ S. typhimurium adenylate cyclase mutants (strains 5336 or SBl786, respectively), while increased amounts were extractable from cyclic AMP phosphodiesterase mutants and cyclic AMP receptor protein mutants relative to the parental strains (19).
4. Measurement of cyclic AMP by three independent assay proce- dures (the Gilman procedure (30), the radioimmunoassay (331, and the protein kinase method (34)) gave comparable values (see Table VIII).
Other control experiments ruled out artifactual degradation, non- specific absorption and uptake of cyclic AMP by cells as possible sources of error: a suspension of washed AB257~“-’ cells was exposed to cyclic [‘HIAMP (5 x lo-’ M, 4 x 10’ cpm/ml) for 30 min at 37” or 0” and then brought to 100” for 5 min or left at 0”. Subsequently the cells were removed by centrifugation and the radioactivity remaining in the supernatant was measured. The concentration of cyclic [3H]- AMP was unaltered +2%. The radioactive material was subjected to electrophoresis in 0.05 M sodium borate buffer, by which cyclic AMP was clearly separated from adenosine, AMP, ADP, and ATP. Essen- tially all of the radioactivity migrated with cyclic AMP. In another ex- periment glucose-grown AB257 pee’ cells were exposed to cyclic [3H]- AMP for up to 1 hour, either in the presence or absence of 5 rnM glu- cose. The cells were filtered and washed and associated radioactivity was measured. Only background radioactivity was associated with the washed cells.
RESULTS
Induced Synthesis of Enzyme II Complexes of PTS in
Escherichia coli-In order to determine the inducibility of the sugar-specific protein components of the PTS, AB257*“-’ cells were grown in minimal medium containing one of several sugars. The capacity of the cells to accumulate radioactive metabolites from appropriate radioactive sugars was then determined in uiuo, and the specific activities of the different Enzyme II complexes were estimated in uitro. Uptake of a particular sugar substrate of the PTS was greatly enhanced by prior growth of the cells in the presence of the same substrate (Table III). For example, growth in the presence of glucose in- duced glucose and methyl ol-glucoside uptake activity, while fructose and mannitol respectively enhanced uptake rates of each of these two carbohydrates. Maximal induction was demonstrable only if low concentrations of the substrate sugars were used in the assay, presumably because the different En- zyme II complexes exhibit cross-substrate specificities.
Corresponding results were obtained when the activities of the membrane-associated Enzyme II complexes were assayed in vitro in the presence of excess quantities of the soluble components of the PTS (Table IV). The activities of the Enzyme II complexes were from 8- to 40-fold inducible. In each case, growth in the presence of the normal substrate of an Enzyme II complex maximized the activity of that complex, although other sugars exhibited some inductive capacity. Since chloramphenicol (50 pg/ml) blocked induction of the Enzyme II complexes, it appeared that synthesis, rather than the ac- tivities of the complexes were the targets of induction.
Regulation of Glycerol Uptake by PTS in Escherichia coli’-Our previous studies with Salmonella strains showed
‘Uptake studies reported here were generally conducted with “‘C-labeled substrates which could be metabolized by induced wild
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

886 Regulation of Carbohydrate Metabolism in E. coli
TABLE III TABLE IV
Phosphorylation of [“C]sugars by cell-free extracts ofEscherichia coli strain AB25P’-’ cells
Uptake of sugars by Escherichia coli strain AB25P’-’ cells
Medium 63, containing 20 &ml of methionine and the carbon sources indicated below at 0.5% (total volume 1 liter), was inoculated with 25 ml of an overnight culture of lactate-grown AB257Dc-’ cells. The cells were grown aerobically at 37” for 4 hours, harvested in the exponential phase of growth, washed four times with cold Medium 63, and resuspended in Medium 63 to the desired cell density for uptake measurements. The cell suspensions were shaken at 28” for 15 min before initiation of the transport experiment by addition of a radioac- tive sugar (final concentration, 8 PM; specific activities between 5 and 10 x lo6 cpm/pmol). Uptake of metabolizable sugars was linear with time for at least 8 min (the duration of the experiment) and with cell density within the range studied (10 to 40 pg dry weight/ml). Higher cell densities were used to estimate methyl a-glucoside accumulation (50 to 200 fig dry weight/ml). Other procedures were as described previously (10, 11).
AB257DC-’ cells were grown, harvested, and washed as described in Table III. A thick cell suspension in buffer containing 10 rnM Tris-HCl, pH 7.4, M&O, (5 mM), and dithiothreitol (0.5 mM) was passed through a French pressure cell at 10,000 p,s.i., centrifuged to remove cell debris, and then centrifuged at 200,000 x g for 1.5 hours to separate membranes from soluble proteins. The pellet was resuspended in the same buffer. For Enzyme II assays this buffer was supplemented with phosphoenolpyruvate (5 Fmol), potassium fluoride (10 rmol). a soluble extract derived from glucose-grown AB25SPC-’ cells (1 mg of protein, free of Enzyme II activities), and the membrane preparation to be assayed (0.05 to 0.5 mg of protein) in a final volume of 1 ml. The reaction was terminated after 20 min at Z37” and sugar phosphate was determined as described previously (28).
Uptake of [“C]sugar after growth
Radioactive sugar (8 WI
Glucose” Methyl n-glucos’aeb Fructose” Mannitol”
Lactate
0.9 0.7
0.5 0.8
in medium containing-
L.actate + Lactate 1 Lactate + glUCl,Se fructose mannltrll
6.3 2.8 3.4 L5.9 1.5 5.2 0.9 6.4 0.6 1.0 2.0 9.4
Phosphorylation of [“C Isugar” after growth in medium containing
Radioactive sugar (lo!4 Lactate Lactate - Lactate + Lactate
glucose fructose mannitol
Glucose 0.65 7 3.9 3.0
Methyl a-glucoside 0.06 1.6 0.3 0.65
Fructose 0.2 0.55 4.2 0.45
Mannitol 0.76 0.85 1.7 22.<5
u Micromoles of sugar phosphorylated per g of membrane protein per
“Micromoles of [“Cjsugar taken up per min per g dry weight of cells.
min at 37”
*Micromoles of [‘“C]methyl n-glucoside accumulated per g dry weight of cells after 12 min.
altered by a mutation which depressed cyclic AMP phosphodi- esterase activity.
that the uptake of several carbohydrates by the wild type organism was subject to inhibition by high concentrations of’ methyl cu-glucoside and other sugar substrates of the PTS (10). Initial attempts to extend these observations to E. coli strains were only partially successful. Uptake of isopropyl p-thi- ogalactoside via the lactose permease was subject to inhibi-
tion by methyl a-glucoside, but little or no inhibition of glyc- erol or maltose uptake was observed. The data summarized in Table V show that growth of E. coli cells in the presence of glucose greatly enhanced their sensitivity to inhibition of
glycerol uptake by methyl oc-glucoside. Sensitivity was not
type cells. Few nonmetabolizable transport substrates are available. Thus, the uptake studies represent a combination of transport and metabolism, and actual transport rates were not determined. For the purpose of the present discussion the term “transport” will be used exclusively when it is known that the transport process per se is being measured. The term “uptake” will be employed when the substrate is metabolized. Published accounts of glycerol uptake in Escherichia coli cells provide evidence that intracellular glycerol phosphorylation, rather than transmembrane glycerol transport is rate limiting for glyc- erol utilization (35, 36). In spite of these observations it is considered likely that the activity of the glycerol permease, rather than that of glycerokinase is subject to PTS-mediated regulation for the following reasons: (a) In both Salmonella typhimurium and E. coli cells, glycerol, maltose, and melibiose uptakes are regulated by PTS-sugars in a similar fashion. Transport of nonmetabolizable substrates of the maltose and melibiose permease systems has been shown to be subject to PTS-mediated regulation (10). (b) A mutation in E. coli which abolished sensitivity of glycerokinase to feedback inhibition by fruc- tose 1,6-diphosphate (glpK), and another mutation which resulted in the constitutive synthesis of the glycerol catabolic enzyme system (glpR) did not influence PTS-mediated regulation of glycerol uptake (37). (c) A mutation in S. typhimurium which specifically abolished the inhibitory effects of PTS sugars on glycerol uptake had no effect on the in vitro regulation of glycerokinase activity or on the induced expres- sion of the glycerol regulon in the absence of a repressing sugar (38). Detailed accounts of these experiments will be published elsewhere.
Several experiments suggested that growth in the presence of glucose enhanced sensitivity to inhibition by inducing the synthesis of a cell constituent (or constituents) which was
involved in regulation. First, inhibition of glycerol uptake by methyl cu-glucoside could be enhanced either by initially inducing glycerol transport and then by growing the cells in the presence of glucose (as described in Table V) or by first growing the cells in glucose-containing medium, followed by transfer of the washed cells to glycerol medium for induction of the glycerol catabolic enzyme system. Strong inhibition of glycerol uptake by methyl a-glucoside was observed by either proce- dure. Second, the inductive effect of glucose was blocked by chloramphenicol (Table V), suggesting that glucose exerted its inductive effect, only if protein synthesis could occur. Since chloramphenicol-treated cells took up and metabolized glucose normally, this result also showed that glucose metabolism per se did not promote regulation. Third, the results summarized in Table V show that induction of methyl Lu-glucoside inhibi- tion also occurred in E. coli strain ptsZ109. This strain, derived
from AB257~‘-‘, is deficient for Enzyme I of the PTS and therefore cannot transport or metabolize glucose rapidly. Thus, rapid glucose transport or metabolism was neither necessary nor sufficient to induce the regulatory interactions responsible for inhibition of glycerol uptake by methyl a-gluco- side.
Sugars other than methyl ol-glucoside were found to inhibit glycerol uptake. In each case, however, induction of the corresponding catabolic enzyme system was required. As can be seen from the data in Table VI, growth of AB257Pc-’ cells in the presence of glucose specifically enhanced inhibition of glycerol uptake by glucose and methyl oc-glucoside, but not by fructose or mannitol. By contrast, growth in the presence of fructose or mannitol respectively enhanced sensitivity to inhibition by the inducing sugar. The observation that growth in the presence of mannitol enhanced sensitivity to inhibition
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

Regulation of Carbohydrate Metabolism in E. coli 887
by glucose and methyl cY-glucoside as well as mannitol agrees with the inducer specificities of the Enzyme II complexes (Tables III and IV).
If induction of the Enzyme II complex specific for a given sugar explains the induction of the capacity of that sugar to inhibit glycerol uptake, genetic loss of a specific Enzyme II should render the strain resistant to regulation by the sugar (or sugars) for which that Enzyme II is specific, provided that no other Enzyme II recognizes the same sugar. In order to test this prediction, mutants lacking the Enzyme II specific for man- nose and glucosamine were isolated (see “Experimental Proce-
TABLE V
Inhibition of glycerol uptake by methyl a-glucoside into Escherichia coli strains
Cells were grown overnight at 37” with shaking in Medium 63
supplemented with 0.5% glycerol and 20 pg/ml ofmethionine. One-mil-
liliter aliquots of these cultures were used to inoculate 25 ml of fresh
medium of the same composition in 125.ml Erlenmeyer flasks. The
flasks were rotated at 250 rpm at 37” for 2 hours. Aliquots of 15 ml were removed from each flask (glycerol-grown cells). Subsequently glucose
was added to the remaining media to a concentration of0.5%, and after
continued growth for 2 hours, cells were harvested in the exponential
growth phase (glycerol + glucose-grown cells). Cells were washed three
times with Medium 63 and resuspended to a cell density of 50 klett units (0.2 mg of dry cells/ml) for [‘“Clglycerol uptake measurements at
32” (11). The concentration of [“Clglycerol was 0.5 mM; that of methyl
cu-glucoside was 5 mM. In one experiment (AB257eC-‘, glycerol +
glucose + chloramphenicol) 50 pg/ml of chloramphenicol was added to the growth medium together with glucose 2 hours before the cells were
harvested.
Strain
Per cent
Carbon sources for growth inhibition IJninhibited by methyl uptake rate
a-glucoside
AB257
AB257 AB257~“-’
AB257P'-'
AB257""-'
ptsJlO9
ptsIl09 ptsIl09 err-27
ptsIl09 err-27
Glycerol
Glycerol + glucose
Glycerol
Glycerol + glucose Glycerol + glucose i
chloramphenicol
Glycerol
Glycerol + glucose
Glycerol Glycerol + glucose
11
73
14
83 19
15
83
5 2
~mollminlg dry weight
15
12 12
7
6
13 10
15
20
TABLE VI
Inhibition of glycerol uptake by sugars into Escherichia coli strain AB25Pc-’ cells
Growth was conducted as described in the legend to Table V except
that the sugar indicated below was added to the culture growing
exponentially in glycerol minimal medium three hours before cells
were harvested. Uptake of [“Clglycerol (0.5 mM) in the presence and
absence of inhibiting sugar was measured as described in Table V.
Inhibiting sugar (5 mh4)
Per cent inhibition of [“C]glycerol uptake after growth in medium containing
Glycerol Glycerd Glycerol Glycerol + + +
glucose fructose mannitol
Glucose 15 90 30 80
Methyl ol-glucoside 5 95 6 90 Fructose 5 0 45 0
Mannitol 0 0 0 80
dure”). The mutants lost the ability to grow with mannose or glucosamine as sole source of carbon (Table II) although they
retained the ability to utilize glucose. Several investigations have shown that there are two Enzyme II complexes which act
on glucose as substrate (28, 29).’ One phosphorylates glucose and methyl cx-glucoside efficiently, but not mannose or gluco- samine, while the other phosphorylates glucose, mannose and glucosamine efficiently but not methyl a-glucoside. The latter complex is presumably defective in the manA mutant.
All of the glucose analogues tested strongly inhibited glycerol uptake by strain AB257 PC-l if cells were grown in the presence of glucose, but not if mannose or glucosamine served as the inducing sugar (Table VII). The latter two sugars were found to be poor inducers of Enzyme II activity in agreement with the data in Table II. Moreover, the manA mutation abolished sensitivity of glycerol uptake to inhibition by mannose and glucosamine and decreased sensitivity to inhibition by 2-deox- yglucose. It had no effect on regulation by methyl a-glucoside or glucose.
In S. typhimurium a mutation (the err mutation) was shown to render synthesis of several catabolic enzyme systems resist- ant to repression and the corresponding non-PTS transport systems resistant to inhibition by the sugar substrates of the PTS (10, 11).
The same appears to be true in E. coli. Regardless of whether
or not cells possessing a err mutation were grown in the presence of glucose, no inhibition of glycerol uptake by methyl
or-glucoside or other sugar substrates of the PTS was observed (Table V).
Detailed Characteristics of Glycerol Uptake Inhibition-Fig. 1 shows a time course for glycerol uptake by strain AB257P”.’ cells grown in the presence of glycerol and glucose. Glycerol uptake, both in the presence and absence of the glucose analogue, was linear with time, the rate in the presence of methyl cu-glucoside being only a small fraction of that observed in its absence. Moreover, inhibition was observed as soon as the glucoside was added to the cell suspension. The latter observa- tion provided evidence that extensive intracellular accumula- tion of the sugar or sugar phosphate was not required for inhibition.
Inhibition of glycerol uptake as a function of methyl cu-gluco-
side concentration is shown in Fig. 2. The ptsZl09 mutant appeared to be somewhat more sensitive to low concentrations of the glucose analogue than the parental strain. This increased sensitivity to inhibition is particularly worthy of note, since the
mutant accumulated far less sugar intracellularly than the parent. Enhanced sensitivity of leaky Enzyme I mutants of
TABLE VII
Inhibition of [‘“Cjglycerol uptake by heroses: dependence on Enzyme II”“” of PTS
The experimental design was as in Table V.
Per cent inhlbitmn of [“C]glycerol uptake
Inhi~~ine;ugar .m
Glucose
Methyl wglucoside
2.deoxyglucose Mannose
Glucosamine
jJQjT”C.’ AB2jyle-l manA (glycerol + (glycerol + (glycerul + mannOSe- glUCOSe- glucose-
gKWn) grown ) grown)
35 90 90
20 95 95
25 95 90
25 80 0
25 70 0
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from
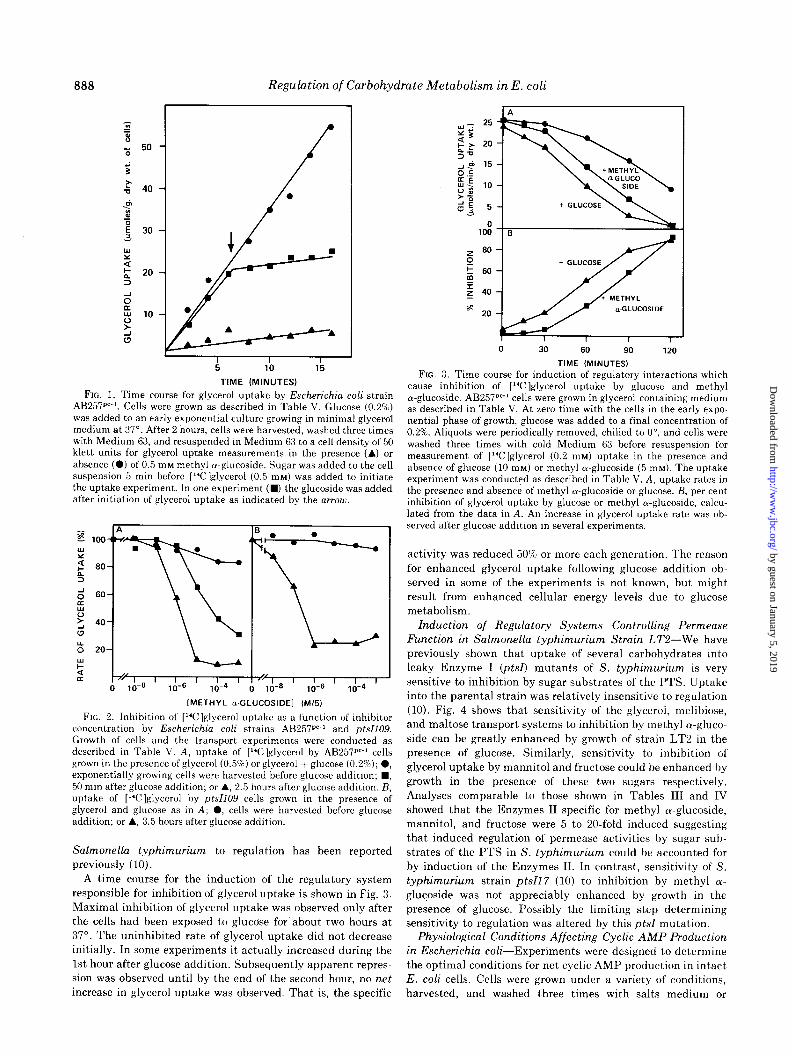
888 Regulation of Carbohydrate Metabolism in E. coli
I I I I
5 10 15
TIME (MINUTES)
FIG. 1. Time course for glycerol uptake by Escherichia coli strain A13257pc-‘. Cells were grown as described in Table V. Glucose (0.2%) was added to an early exponential culture growing in minimal glycerol medium at 37”. After 2 hours, cells were harvested, washed three times with Medium 63, and resuspended in Medium 63 to a cell density of 50 klett units for glycerol uptake measurements in the presence (A.) or absence (0) of 0.5 mM methyl cu-glucoside. Sugar was added to the cell suspension 5 min before [“C]glycerol (0.5 mM) was added to initiate the uptake experiment. In one experiment (H) the glucoside was added after initiation of glycerol uptake as indicated by the arrow.
A B,. 0 l
Y . l
2 80 t 3
d 60
E
s ‘oo!;:x ‘\,L _
y 40 6
k 20
2
d 0 10-S 10-6 10-4 0 10-S 10-S 10-4
[METHYL @GLUCOSIDE] (M/5)
FIG. 2. Inhibition of [“Clglycerol uptake as a function of inhibitor concentration by Escherichia coli strains AB257P’-’ and ptslZ09. Growth of cells and the transport experiments were conducted as described in Table V. A, uptake of [“C]glycerol by AB257ec-’ cells grown in the presence of glycerol (0.5%) or glycerol + glucose (0.2%); 0, exponentially growing cells were harvested before glucose addition; W, 50 min after glucose addition; or A, 2.5 hours after glucose addition. B, uptake of [“Clglycerol by ptsIlO9 cells grown in the presence of glycerol and glucose as in A; 0, cells were harvested before glucose addition; or A, 3.5 hours after glucose addition.
Salmonella typhimurium to regulation has been reported previously (10).
A time course for the induction of the regulatory system responsible for inhibition of glycerol uptake is shown in Fig. 3. Maximal inhibition of glycerol uptake was observed only after the cells had been exposed to glucose for’about two hours at 37”. The uninhibited rate of glycerol uptake did not decrease initially. In some experiments it actually increased during the 1st hour after glucose addition. Subsequently apparent repres- sion was observed until by the end of the second hour, no net increase in glycerol uptake was observed. That is, the specific
a-GLUCOSIDE
0 30 60 90 120
TIME (MINUTES) FIG. 3. Time course for induction of regulatory interactions which
cause inhibition of [“Clglycerol uptake by glucose and methyl cx-glucoside. AB257e’-’ cells were grown in glycerol-containing medium as described in Table V. At zero time with the cells in the early expo- nential phase of growth, glucose was added to a final concentration of 0.2%. Aliquots were periodically removed, chilled to O”, and cells were washed three times with cold Medium 63 before resuspension for measurement of [“C]glycerol (0.2 mM) uptake in the presence and absence of glucose (10 mM) or methyl ol-glucoside (5 mM). The uptake experiment was conducted as described in Table V. A, uptake rates in the presence and absence of methyl u-glucoside or glucose. B, per cent inhibition of glycerol uptake by glucose or methyl cu.glucoside, calcu- lated from the data in A. An increase in glycerol uptake rate was ob- served after glucose addition in several experiments.
activity was reduced 5OY0 or more each generation. The reason for enhanced glycerol uptake following glucose addition ob- served in some of the experiments is not known, but might result from enhanced cellular energy levels due to glucose metabolism.
Induction of Regulatory Systems Controlling Permease Function in Salmonella typhimurium Strain LT2-We have previously shown that uptake of several carbohydrates into leaky Enzyme I (pt.sI) mutants of S. typhimurium is very sensitive to inhibition by sugar substrates of the PTS. Uptake
into the parental strain was relatively insensitive to regulation (10). Fig. 4 shows that sensitivity of the glycerol, melibiose, and maltose transport systems to inhibition by methyl a-gluco-
side can be greatly enhanced by growth of strain LT2 in the presence of glucose. Similarly, sensitivity to inhibition of glycerol uptake by mannitol and fructose could be enhanced by growth in the presence of these two sugars respectively. Analyses comparable to those shown in Tables III and IV showed that the Enzymes II specific for methyl a-glucoside, mannitol, and fructose were 5 to 20.fold induced suggesting that induced regulation of permease activities by sugar sub- strates of the PTS in S. typhimurium could be accounted for by induction of the Enzymes II. In contrast, sensitivity of S.
typhimurium strain ptsIl7 (10) to inhibition by methyl a- glucoside was not appreciably enhanced by growth in the presence of glucose. Possibly the limiting step determining sensitivity to regulation was altered by this ptsl mutation.
Physiological Conditions Affecting Cyclic AMP Production in Escherichia coli-Experiments were designed to determine the optimal conditions for net cyclic AMP production in intact E. coli cells. Cells were grown under a variety of conditions, harvested, and washed three times with salts medium or
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

Regulation of Carbohydrate Metabolism in E. coli
1 A. GLYCEROL
g 100
Y 2 80
5 80
g 40
li 2 20
0
0 10-7 104 10-3
B. MELIBIITOL MALTOSE
[METHYL a-GLUCOSIDE] (~15)
FIG. 4. Regulation of permease function in Salmonella typhimu- rium. S. typhimurium strain LT.2 was grown with shaking at 37” for 4 hours in Medium 63 containing 0.5% glycerol (A), melibiose (I?), or maltose (C). Aliquots were removed and glucose was added to the remaining cell suspension to a final concentration of 0.2%~. After 2 additional hours at 37”, the cells were harvested in the exponential
buffer, and resuspended in the same solution. The cell suspen- sion was then incubated at 37”, and the cyclic AMP produced
in a short time interval was determined. Cyclic AMP produc- tion was maximal when cells were harvested during exponen- tial growth with any of a variety of sugars as carbon sources, provided that the sugar supported rapid growth. The sugars tested at 0.2%, included glucose, galactose, fructose, mannitol, and lactose. Lower rates of cyclic AMP production were observed after growth with nL-lactate or glycerol as carbon source. In each case, the rate of cyclic AMP production was proportional to cell density and linear with time for up to 30 min.
Employing galactose-grown cells, the dependence of cyclic AMP production on pH was studied. With Medium 63 ad- justed to different pH values, in uiuo cyclic AMP production
was found to have a broad neutral pH optimum with half-max- imal activities at about pH 5.5 and pH 8. The rate of cyclic AMP production was the same in Medium 63 and in potassium phosphate buffer of the same pH. All subsequent experiments were conducted with cells in Medium 63, pH 7.2.
A time course for cyclic AMP production is reproduced in Fig. 5. Net cyclic AMP synthesis was linear with time for up to 30 min. Subsequently the rate gradually decreased, approaching zero after 2 hours. Fig. 5 also shows that chloramphenicol, at a concentration that inhibited protein synthesis more than 98%, had no inhibitory effect during the initial phase of cyclic AMP production.
The decrease in the rate of cyclic AMP production with time
shown in Fig. 5 appears to be due to a decrease in the true rate of cyclic AMP synthesis. No net degradation of cyclic AMP occurred during an additional 4-hour incubation either in the
cell suspension or after removal of cells by filtration. Further- more, after a 3-hour incubation at 37”, cells removed from the medium by centrifugation and resuspended in fresh Medium 63 no longer made cyclic AMP at an appreciable rate.
Regulation of Cyclic AMP Metabolism by PTS-Previous reports have shown that glucose can regulate cyclic AMP metabolism in E. coli. The specificity of this effect was examined employing different glucose analogues (Table VIII). In the presence of either n-glucose or the nonmetabolizable
glucose analogue, methyl a-glucoside, the net synthesis of cyclic AMP was completely inhibited. Both of these sugars are substrates of the glucose phosphotransferase, being trans-
889
growth phase, washed three times with cold Medium 63, and resus- pended to 0.2 tug/ml, dry weight for transport studies. Uptake of the sugar used as a carbon source for growth was measured in the presence and absence of methyl oc-glucoside as indicated on the abcissa. 0, cells grown in the absence of glucose; W, cells exposed to glucose for 2 hours during growth
0 40 80 120 160
MINUTES FIG. 5. Time course for cyclic AMP production by Escherichia coli
cells. AB257~‘-* cells were grown overnight in 25 ml of Medium 63 containing 0.2% galactose and 20 pg/ml of methionine in a 125-1111 Erlenmeyer flask. A lo-ml aliquot of this stationary phase culture was used to inoculate 250 ml of fresh medium of the same composition. The cell suspension was rotated in a 2.liter Erlenmeyer flask at 250 rpm for 3 hours at 37” during which the cells grew 2.2 generations. Cells were harvested by centrifugation, washed three times with Medium 63, and resuspended in the same medium to 80 klett units (0.32 mg dry cells/ml). Two tubes, each containing 10 ml of the cell suspension, one with 50 @g/ml of chloramphenicol (A), and one without chlorampheni- co1 (O), were incubated at 37”. One milliliter aliquots were periodically removed and boiled for 5 min. Cells were removed by centrifugation and the cyclic AMP in the supernatant was measured by the Gilman assay procedure.
ported and phosphorylated by this enzyme system (28). By contrast, L-glucose and methyl P-6-deoxy-n-glucoside, which are not substrates of the PTS, had little influence on cyclic AMP metabolism. These results suggested that only those
glucose analogues which interact with the PTS as substrates can regulate cyclic AMP metabolism.
Fig. 6A shows net production of cyclic AMP as a function of methyl a-glucoside concentration for glucose-grown and galac- tose-grown AB257Pc-’ cells. When the cyclic AMP phos- phodiesterase-deficient strain was grown in minimal medium with glucose as carbon source, low concentrations of methyl a-glucoside (50 pM) maximally inhibited cyclic AMP produc- tion. These results are to be contrasted with those for galac- tose-grown cells. In the latter case, even high concentrations of methyl cu-glucoside exerted little regulatory effect on net cyclic
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

Regulation of Carbohydrate Metabolism in E. coli
TABLE VIII
Regulation of cyclic AMP synthesis by glucose analogues, and comparison of different assay procedures for cyclic AMP
Experiment I-Cells were grown as described in the legend to Fig. 5
except that the carbon source used for growth was 0.24 glucose.
Exponentially growing cells were washed three times with Medium 63
and were resuspended in the same medium to a cell density of 80 klett units. One-milliliter aliquots of the cell suspensions were incubated at
37” for 15 min in the presence or absence of the indicated sugar (5
mM). Subsequently, the cell suspensions were boiled for 5 min to extract the total cyclic AMP. A negligible amount of cyclic AMP
(<0.02 nmol) was present in the cell suspension at zero time. Cyclic
AMP was measured both by the Gilman assay (31) and by the protein
kinase assay procedure (34). Values are expressed as nanomoles of
cyclic AMP per ml of culture medium.
Experiment 2-AB257ec-’ cells were grown overnight in 10 ml of
Medium 63 containing 0.5% glucose and 20 &ml of methionine. One-milliliter of this cell suspension was used to inoculate 25 ml of
fresh growth medium of the same composition. The cell suspension was
rotated in a 125.ml Erlenmeyer flask at 250 rpm and 37” until the exponentially growing culture reached a cell density of 80 klett units.
Subsequently, cells were harvested by centrifugation, washed three
times with Medium 63, and resuspended to 100 klett units. Test tubes containing one-milliliter aliquots of the washed cell suspension were
incubated for 20 min at 37” in the presence or absence of the indicated
sugar (5 mM). Total cyclic AMP was extracted as described under
“Experiment 1,” and the cyclic AMP was estimated by the Gilman
and radioimmunoassay procedures. In this experiment, the cell sus- pension contained 0.40 nmol of cyclic AMP per ml at zero time.
Cyclic AMP (nmolhnl)
Sugar added during incubation Protein kinase assay
Experiment 1
None
n-glucose Methyl a-n-glucoside
L-glucose
B-Methyl 6.deoxyglucose
Experiment 2
None
n-glucose Methyl wglucoside
L-glucose
0.58 0.43
0.02 0.02 0.03 0.02
0.54 0.49 0.70
Ghan assay Kadi~limmuno Assam
1.5 1.1 0.44 0.34
0.40 0.28
1.5 1.1
AMP synthesis. It therefore appeared that glucose induced the
synthesis of a component of the regulatory system. This suggestion was strengthened by the observation that exposure of galactose-grown AB257~“-’ cells to 10 mM glucose for 1 hour at 37’ enhanced their sensitivity to regulation by methyl a-glucoside, but not if chloramphenicol was present, or if the temperature was maintained at 0”.
Corresponding data for an Enzyme I-deficient strain are plotted in Fig. 6B. Two observations are particularly worthy of
note. First, the mutant was hypersensitive to regulation by methyl a-glucoside as compared with the parental strain. Low concentrations of the sugar strongly inhibited cyclic AMP production, even with galactose-grown cells. Second, growth in the presence of glucose enhanced sensitivity to regulation. These results suggested that sensitivity of cyclic AMP metabo- lism to regulation by methyl cu-glucoside was determined by the cellular levels of Enzyme I and another protein (or proteins) specifically induced by glucose.
The behavior noted above for strain ptsZZ09 was quite
2 2 0.8
E = 0.6
: 0.4 0 ; > 0.2 0
A, AB257”’ 8. pts 1109
LrL’!cdd 0 10-7 10-5 10-3 0 10-J 10-s 10-S
[METHYL di~ucosIDE1 (M/51
FIG. 6. Cyclic AMP production by AB257~‘~’ and ptsIl09 cells as a function of methyl a-glucoside concentration. Cells were grown and prepared as described in the legend to Fig. 5 with 0.5% glucose (0) or 0.5% galactose (ml as the carbon source. Aliquots (1 ml) of the washed cell suspensions at a cell density of 100 klett units (0.4 mg dry weight per ml) were incubated at 37” for 20 min in the presence of the methyl a-glucoside concentration indicated on the abscissa. Subsequently total cyclic AMP was estimated as described under “Experimental Proce- dure.” The amounts of cyclic AMP present in the different cell prep- arations at zero time were as follows. Glucose-grown AB257ec-’ cells, 0.16 nmol/ml; galactose-grown AB257P”-’ cells, 0.13 nmol/ml; glu- cose-grown ptsIl09 cells, 0.05 nmol/ml; galactose-grown ptsIZO9 cells, 0.09 nmol/ml.
different from that observed for a mutant which appeared to completely lack functional Enzyme I (ptsZs02, Table I). Employing the conditions described in Fig. 6Z3, cyclic AMP production by this strain amounted to less than 0.05 nmol/ml during a 20-min incubation at 37” in the absence of sugar (data not shown). The behavior of this “tight” Enzyme I mutant in
the absence of methyl a-glucoside therefore resembled that of the “leaky” Enzyme I mutant in the presence of sugar.
If the glucose-inducible constituent of the system regulating
cyclic AMP synthesis is the Enzyme II complex of the PTS, induction of an Enzyme II complex should permit regulation of cyclic AMP metabolism by the corresponding sugar substrate. Table IX summarizes data for sugar substrates of four different Enzyme II complexes, those specific for (a) glucose and methyl ol-glucoside, (b) mannose, glucosamine, and glucose, (c) man- nitol, and (d) fructose. It can be seen that regulation of cyclic AMP production by substrates of either the glucose or the mannose Enzyme II complex was maximal after growth of the cells in the presence of glucose. Regulation by either mannitol or fructose was induced by the corresponding sugar, but not by glucose. Thus it appeared that regulation of cyclic AMP synthesis by a sugar substrate of the PTS generally depended
on induction of the corresponding Enzyme II (see Tables III and IV).
Table X summarizes inhibition data for the manA. mutant which lacks a functional mannose Enzyme II complex but
retains the glucose (methyl n-glucoside).specific complex. Corresponding data are included for the parental strain grown under inducing and noninducing conditions. Cyclic AMP production in the mutant strain was fully sensitive to inhibi- tion by glucose and methyl a-glucoside, partially resistant to inhibition by 2-deoxyglucose, and completely resistant to inhibition by mannose and glucosamine. These results corre- spond with the known substrate specificities of the glucose and mannose Enzyme II complexes noted above and confirm the suggestion that the Enzyme II complex, or one of its constitu- ent proteins, is a component of the system which mediates the regulation of cyclic AMP metabolism by sugar substrates of the
PTS.
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

Regulation of Carbohydrate Metabolism in E. coli 891
TABLE IX
Inhibition of net cyclic AMP synthesis by sugar substrates of PTS in Escherichia cd strainAB257pc-’ cells
Cells were grown as described in Table VIII with the carbon sources indicated below. Washed cells, resuspended to a cell density of 100 klett units (0.4 mg dry wt/ml) were incubated for 30 min at 37” in the presence or absence of an inhibiting sugar. Subsequently total cyclic AMP (medium plus cells) was extracted and measured as described under “Experimental Procedures.” The amounts of cyclic AMP produced by the various cell suspensions (in micromoles of cyclic AMP/g, dry wt of cells/30 min) follows: glucose-grown cells, 6; mannitol-grown cells, 4.4; galactose-grown cells, 3.6; fructose-grown cells, 2.1.
Inhibition sugar (5 InM)
Glucose Methyl a-glucoside Fructose Mannitol Mannose
Per cent inhibition of net cyclic AMP synthesis after growth in medium containing
Galactose GlUCM2 Fructose Mannitol
82 100 83 100 27 100 34 90 16 4 100 4
-6” - 12” -4” 100
42 100 -b -
a Mannitol stimulated net cyclic AMP synthesis slightly. b -, not determined.
TABLE X
Dependence ofsugarpromoted inhibition of cyclic AMPproduction on Enzyme II of phosphotransferase system
Cells were grown and prepared as described in Fig, 5 except that the carbon source for growth was either 0.5%) galactose (Column I, strain AB257pc-*) or 0.5% glucose (Columns 2 and 3, strains AB257e”.’ and manA51, respectively). Strain manA5l is a mutant derived from AB257~‘~’ which lacks the Enzyme II complex which catalyzes the transport and phosphorylation of mannose and glucosamine. Washed cells, resuspended to a cell density of 100 klett units, were incubated for 20 min at 37” in the presence of the sugar indicated below at a concentration of 5 mu. In the absence of sugar, galactose-grown AB257”‘.’ cells produced 0.74 nmol of cyclic AMP/ml; glucose-grown AB257p’.’ cells produced 0.58 nmol of cyclic AMP/ml; and glucose- grown manA cells produced 0.67 nmol of cyclic AMP/ml. In manA5I cells, mannose and glucosamine stimulated cyclic AMP production 10 to 30’%
Per cent Inhibition of cychc AMP production
Inhibiting sugar
Glucose Methyl cr.glucoside 2.deoxyglucose Mannose Glucosamine
AB2:j7”~~1 AR25V 1 IlUXlASl (galactose- (glUCOSe- (glWW-
grown) gWVn) grown)
85 100 100 25 100 100 95 100 90 40 100 0
60 90 0
DISCUSSION
Koch et al. and Winkler and Wilson showed that regulation of lactose permease activity by methyl a-glucoside in E. cob was enhanced by growth in the presence of glucose and that maltose transport was subject to a kinetically similar type of regulation (6-8). The involvement of the glucose permease was
inferred. Our studies on glycerol uptake permit us to correlate induction of regulatory interactions with that of the Enzyme II of the PTS. In addition, loss of a specific Enzyme II was shown to render glycerol uptake resistant to regulation by the corre-
sponding sugar substrates, confirming the importance of this enzyme complex in the regulatory process. The fact that the err
mutation abolished regulation of the glycerol, lactose, melibi- ose, and maltose uptake systems by sugar substrates of the PTS indicated that a single mechanism for the control of these permeases must be operative. Since .a similar mutation desen- sitized several permeases to regulation in S. typhimurium, it appears that the regulatory mechanism is the same in these two organisms (10, 39).
Extensive studies, particularly by Perlman, Pastan, and their collaborators, have established many details of the mechanism by which cyclic AMP regulates the induction of enzyme synthesis (12, 13). Mutants which lack adenylate cyclase, and therefore cannot synthesize cyclic AMP, lack the ability to utilize many carbon sources for growth because the requisite catabolic enzymes cannot be induced. Induction of P-galactosidase synthesis normally depends on the presence of a cyclic AMP’cyclic AMP receptor protein complex which stimulates synthesis of lac specific messenger RNA by binding to the promoter region of the lactose operon. Depressed cellular levels of cyclic AMP therefore provide a plausible explanation
for repression of enzyme synthesis (12, 13, 40-47). One mechanism by which glucose can depress intracellular
cyclic AMP levels is to inhibit net production of cyclic AMP (18-22). In this and previous communications (19, 25) this process was studied in detail. Our results can be summarized as follows.
1. Sugars inhibit the activity of adenylate cyclase in both E. coli and S. typhimurium cells.
2. The inhibitory sugar substrate need not be metabolized in order to exert its effect if it is a substrate of the phosphotrans- ferase system.
3. Cellular levels of the protein components of the PTS determine the sensitivity of adenylate cyclase to inhibition by
sugar substrates of this enzyme system. For example, there appears to exist an inverse relationship between the cellular Enzyme I or HPr concentration and sensitivity of adenylate cyclase to inhibition, while a functional Enzyme II complex
appears to be required.5 4. Other sugars, which enter the bacterial cell via alterna-
tive transport systems, can also inhibit adenylate cyclase activity, provided that functional metabolic systems specific for those sugars are present in the ce11.6
The molecular mechanisms responsible for the inhibition of adenylate cyclase and of carbohydrate permeases by sugar substrates of the PTS in bacteria are not known although sev-
eral mechanisms have been proposed (7, 8, 25, 42247). The re-
‘While growth of bacterial strains in the presence of glucose enhances Enzyme II’z’c activity 5. to lo-fold (Tables III and IV), sensitivity of adenylate cyclase and of the glycerol uptake system to inhibition may be enhanced 100. to 1000.fold under appropriate conditions (Figs. 2. 4, and 6). This observation remains unexplained.
B Glycerol uptake and adenylate cyclase activities were inhibited by a variety of metabolizable non-PTS carbohydrates. For example, galactose, lactose, xylose, ribose, and a-glycerophosphate all inhibited adenylate cgclase activity in E. coli strain AB257P’.‘by more than 90%, provided that the corresponding catabolic enzyme systems had been induced to high levels. Inhibition by lactose was abolished by the genetic loss of the lactose permease and greatly diminished by loss of P-galactosidase. The ptsIlO9 mutation had no effect on the inhibition of either glycerol uptake or adenylate cyclase by lactose, and the err-27 mutation had no effect on the inhibition of’ glycerol uptake. Methyl P-thiogalactoside did not inhibit cyclic AMP synthesis and inhibited glycerol uptake less severely than did lactose. These results suggest that lactose metabolism contributes to the regulatory effects observed with this sugar and that a mechanism is operative which is distinct from that by which PTS sugars exert regulatory control over adenyl- ate cyclase and carbohydrate permease activities.
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

892 Regulation of Carbohydrate Metabolism in E. coli
suits described here and in a previous communication (25) sug- gest that the regulation of these enzyme systems is effected by mechanisms involving the same cellular constituents. Coordi- nate regulation of adenylate cyclase and carbohydrate per- mease activities provide the bacterium with a mechanism for controlling the intracellular levels of cyclic AMP and inducer in parallel. Dual control over catabolic enzyme synthesis should result (13). The establishment of the molecular details of the regulatory processes will require the development of an in vitro assay system which allows the association of each participating protein with a specific function. These studies are presently in progress.
Acknowledgments-Dr. Susan Taylor (Department of Chem- istry, University of California, San Diego) generously per- formed the protein kinase assay for cyclic AMP, and Dr. G. Sate (Department of Biology, University of California, San Diego) supplied the reagents for the radioimmunoassay for cyclic AMP.
1.
2. 3. 4.
5.
6.
7. 8. 9.
10.
11.
12. 13.
14. 15.
16.
17.
18.
REFERENCES Roseman, S. (1972) in The Molecular Basis of Biological Transport
(Woessner, J. F., Jr., and Huijing, F., eds) pp. 181-215, Academic Press, New York
Kaback, H. R. (1972) Biochim. Biophys. Acta 265, 367-416 Lin, E. C. C. (1970) Annu. Rev. Genetics 4, 225-262 Horecker, B. I,., Thomas, J., and Monod, J. (1960) J. Biol. Chem.
235, 1580-1585 Kessler, D. P., and Rickenberg, H. V. (1963) Biochem. Biophys.
Res. Commun. 10, 482-487 Winkler, H. H., and Wilson, T. H. (1967) Biochim. Biophys. Acta.
135, 1030~1051 Koch, A. L. (1971) Biochim. Biophys. Acta 249, 197-215 McKinstry, G., and Koch, A. L. (1972) J. Bacterial. 109, 455-458 Adhya, S., and Echols, H. (1966) J. Bacterial. 92, 601-608 Saier, M. H., Jr., and Roseman, S. (1972) J. Biol. Chem. 247,
972-975 Saier, M. H., Jr., Simoni, R. D., and Roseman, S. (1970) J. Biol.
Chem. 245, 5870-5873 Pastan, I., and Perlman, R. (1970) Science 169, 339-344 Magasanik, B. (1970) in The Lactose Operon (Bechwith, J., and
Zipser, D., eds) pp. 198-219, Cold Spring Harbor Laboratory, Cold Spring Harbor
Yokota, T., and Gots. J. S. (1970) J. Bncteriol. 103, 513-516 Katz, L., Kingsbury, D. T., and Helinski, D. R. (1973) J. Bacterial.
114, 577-591 Tao, M., and Lipmann, F. (1969) Proc. Natl. Acad. Sci. U. S. A.
63, 86-92 Nielsen, L. D., Monard, D., and Rickenberg, H. V. (1973) J.
Bacterial. 116, 857-866 Makman, R. S., and Sutherland, E. W. (1965) J. Biol. Chem. 240,
1309~1314
19.
20.
21.
22.
23.
24.
25.
26.
27.
“8. 29.
30.
31. 32.
33.
34.
35.
36. 37.
38.
39.
40. 41.
42.
43
44.
45.
46.
47.
Saier, M. H., Jr., Feucht, B. U., and McCaman, M. T. (1975) J. Biol. Chem. 250, 7593-7601
Peterkofsky, A., and Gazdar, C. (1971) Proc. Natl. Acad. Sci. U. S. A. 68, 2794-2798
Peterkofsky, A., and Gazdar, C. (1973’) Proc. NatLAcad. Sci. U. S. A. 70, 2149-2152
Monard. D.. Janeck. J.. and Rickenberr. H. V. (1970) in The Lacto$e bperon iBe;kwith, J., and-‘Zipser, D., eds) pp. 393-400. Cold Sarinn Harbor Laboratorv. Cold Swing Harbor
Saier, M. H., Jr.,-Vaikntine, R. C., and” Feucht, 6. ii. (1973) Abstracts of American Society of Microbiology, p. 173
Saier, M. H., Jr., Valentine, R. C., and Feucht, B. U. (1974) Abstracts of American Society of Microbiology, p. 179
Saier, M. H., Jr., and Feucht, B. U. (1975) J. Biol. Chem. 250, 7078-7080
Saier, M. H., Jr., and Ballou, C. E. (1968) J. Biol. Chem. 243, 4319-4331
Roseman, S., Abeles, R. H., and Dorfman, A. (1952) Arch. Biochem. Biophys. 36, 232-233
Kundig, W., and Roseman, S. (1971) J. Biol. Chem. 246, 1393-1406 Adler, J., and Epstein, W. (1974) Proc. N&l. Acad. Sci. U. S. A.
71, 2895-2899 Loomis, W. F., Jr., and Magasanik, R. (1964) J. Mol. Biol. 8,
417-426 Gilman, A. G. (1970) Proc. Natl. Acad. Sci. U. S. A. 67, 305-312 Krishna, G., Weiss, B.. and Brodie, B. B. (1968) J. Pharmacol.
Exp. Ther. 163, 379-385 Steiner, A. L., Parker, C. W., and Kipnis, D. M. (1972) J. Biol.
Chem. 247, 1106-1113 Wastila, W. B., Stull, J. T., Mayer, S. E., and Walsh, D. A. (1971)
J. Biol. Chem. 246, 1996-2003 Zwaig, N., Kistler, W. S., and Lin, E. C. C. (1970) J. Bacterial.
102, 753-759 Richey, D. P., and Lin, E. C. C. (1972) J. Bacterial. 112,784-790 Saier, M. H., Jr., Castro, L., Feucht, B. U., and Morse, M. L.
(1975) Fed. Proc. 34, 491 Saier, M. H., Jr., Stroud, H., and Judice, J. (1974) Abstracts of the
American Society of Microbiology, p. 179 Jones-Mortimer, i. c., and Kornberk, H. L. (1974) FEBS Lett.
48, 93-95 Pastan, I. and Perlman, R. L. (1969) J. Biol. Chem. 244,5836-5842 Varmus, H. E., Perlman, R. L., and Pastan, I. (1970) J. Biol.
Chem. 245, 2259-2267 Saier, M. H., Jr., and Stiles, C. D. (1975) Molecular Dynamics in
Biological Membranes, pp. 94-105, Springer-Verlag, New York Wayne, P. K., Fetell, J., and Rosen, 0. M. (1975) Biochem.
Biophys. Res. Commun. 64, 81-88 Abou-Sabe, M., Burday, M., and Gentsch, J. (1975) Biochim.
Biophys. Acta 385, 281-293 Abou-SabB, M., and Mento, S. (1975) Biochim. Biophys. Acta 385,
294-304 Peterkofsky, A., Harwood, J., and Gazdar, C. (1975) J. Cyclic
Nucleotide Research I, 11-20 Epstein, W., Rothman-Denes, L. B., and Hesse, .J. (1975) Proc.
Natl. Acad. Sci. U. S. A. 72, 2X00-2304
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from

M H Saier, Jr, B U Feucht and L J HofstadterEscherichia coli.
enzymes II of the phosphoenolpyruvate: sugar phosphotransferase system in Regulation of carbohydrate uptake and adenylate cyclase activity mediated by the
1976, 251:883-892.J. Biol. Chem.
http://www.jbc.org/content/251/3/883Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/251/3/883.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 5, 2019http://w
ww
.jbc.org/D
ownloaded from





























