Embed Size (px)
Citation preview

CHAPTER II
THE EFFECT OF TEMPERATURE AND CULTUREMEDIUM ON THE GROWTH AND SPORULATION OF
DRECHSLERA CATENARIA
61
\

INTRODUCTION
Sporulation and growth of fungi are affected by light, temperature,
relative humidity and nutrient source. Light and temperature have been
described to affect the sporulation of many imperfect fungi (Carlisle,
1965; Leach, 1962a; Leach, 1962b; Leach, 1967; Marsh et aI, 1959).
Leach (1967) found Drechslera catenaria on PDA to sporulate best at
temperatures below 25C, but there was no sporulation of the fungus
without light regardless of temperature. Q. catenaria sporulated most
abundantly when near-ultraviolet was followed by darkness, though it
also sporulated under continuous light (Leach, 1967). Shoemaker (1962)
and Zeiders (1976) also reported a lack of sporulation of this fungus
without light.
However, nutrition has been implicated (Hawker, 1957) as perhaps
the most important single factor in the control of growth and reproduction
of fungi. Fungi formerly included in the genus Helminthosporium, have
been shown to grow and reproduce on potato-dextrose agar (Leach, 1967),
V-8 juice agar (Dienier, 1955; Zeiders, 1976), lactose casein hydrolysate
(Malca and Ullstrup, 1962) and various other media (Braverman and
Graham, 1960). Good growth and sporulation of Drechslera catenaria have
previously been reported on potato-sucrose agar (Drechsler, 1923),
potato-dextrose agar (Leach, 1967) and V-8 juice agar (Zeiders, 1976).
However, growth and sporulation of D. 'catertaria were variable on sucrose-
asparagine agar (Braverman and Graham, 1960).62

63
Originally there was some difficulty in inducing the Drechslera
catenaria isolate from bentgrass to sporulate adequately in culture for
preparation of inoculum for pathological studies (Larsen et aI, 1980).
The objective of this study was to determine favorable nutritional and
temperature conditions for good growth and sporulation of D. catenaria
in culture.

MATERIALS AND METHODS
Media composition. Three different media were used in this study:
lactose casein hydrolysate medium (LCH) (Malca and Ullstrup, 1962),
potato-dextrose agar (PDA) (Difco Laboratories, Detroit, MI 48201),
and 20% V-8 juice agar (V-8JA) (D~ener,1975). All three media were adjusted
to pH 6.0 with either 6N sodium hydroxide or 6N hydrochloric acid before
autoclaving using a Beckman Expandomatic SS-2 pH meter (Beckman
Instruments, Inc., Fullerton, CA 92634). Twenty ml of medium were
dispensed into each disposable polystyrene petri plate (100 x 15 em).
Fungal isolates used and 'procedure for seeding~ culture media.
A monoconidial isolate (DCL-2) and the mass isolate (DCL) of Drechslera
catenaria, previously described (Chapter I),were used in this study.
Test media were seeded with 8-rom diameter agar discs taken from ten
day LCH fungal cultures incubated at 20C and exposed to 12 hr daily
incandescent and fluorescent lighting (5.5 K lux). Each agar disc was
removed from an area approximately one cm from the margin of the colony,
inverted and placed in the center of a culture plate of appropriate
medium.
Treatment conditions. Seeded plates were incubated for 4, 7, 10
or 14 days in temperature-controlled environmental chambers at 15 ! 1,
20 ! 1, or 25 ! lC with alternating dark (12 hr) and light (12 hr)
periods. The sources of direct irradiation (5.5 K lux) in this study
64

65
were unfilteredG.E. 40-W incandescent lamps and G.E. 40-W cool-white
fluorescent lamps at a distance of 20 cm from the cultures. Temperatures
were measured using a Tele-thermometer (~odel 42SC, Yellow Springs
Instrument Co., Inc., Yellow Springs, OR 45387) with a "Banjo-surface"
temperature probe (#408) and did 'not differ between the chamber air and
the air inside the agar-filled petri plates.
Data collection. Sporulation and mycelial growth were compared on
the three media at the three temperatures. :'For sporulation measurements,
fungal colonies in culture plates were flooded with 10 ml of 1% copper
sulfate-Tween 20 solution and brushed with a camel's hair brush to dis
lodge conidia. After the conidial suspension was decanted, the cultures
were rinsed twice with 5 ml of the copper sulfate-Tween 20 solution to
remove remaining conidia. All three suspensions were combined and the
conidial concentration was determined using a Sedgwick-Rafter counting
cell (VWR Scientific, Columbus, OR 43215) in conjunction with a Roward
ocular microscopic disc (American Optical Corp., Buffalo, NY 14215).
Sporulation means are the averages of conidial counts of twenty ocular
fields.
Mycelial growth was expressed as an increase in mycelial dry
weight and colony area. The average diameter of a colony, the mean
of three random diameter measurements per colony, was used to calculate
the area of the nearly circular colony. For mycelial dry weight
determinations, colonies were removed from agar plates by immersing
the culture plate contents in 50 ml of hot distilled water (93C) for
10 minutes. The extracted colony was dried in a dry-air oven at 38C

66
for 24 hr. Final pH was determined for the agar-distilled water
solution from each culture after ten day incubation at 25C.
Data were recorded for both fungal isolates at different harvest
dates after seeding. Data from the colonies included: area (cm2),
mycelial dry weight (mg/cm2
) and sporulation (conidia/mrn2
) as affected
by temperature and medium composition. The sporulation and area data
were taken from the same treatment plates. Dry weight data were from
different plates since a destructive technique was used for estimating
sporulation. This entire experiment was done three times with three
replications per treatment for the sporulation-colony area plates and
only one replication per treatment for the dry weight determination.
The treatment plates were completely randomized within the chambers
each time the experiment was done. Treatment means were analyzed
according to each harvest date alone and for all dates together for
each character recorded. Analysis was done using the Duncan's New
Multiple Range Test at the 5% level of significance.

RESULTS
Culture "characters. Colonies of both isolates at all temperatures
. and on all media were white four days after seeding the plates. The
two isolates did not differ in mycelial pigmentation under any conditions.
After seven days, the Q. catenaria colonies on LCH were olive-gray and
moderately appressed to the agar surface, whereas on PDA, colonies were
dark gray to black with very dense erumpent growth. Colonies on V-8JA
were light gray with concentric zones of light and dark growth. The
dark zones were closely appressed hyphae with abundant sporulation and
the lighter areas consisted of erumpent aerial hyphae with sparse
sporulation. The dark zones generally corresponded to regions of new
growth exposed to light. No zones were recorded until seven days after
seeding. After ten days, the hydrogen-ion concentration of the PDA
(6.3) and LCH (6.1) cultures remained nearly unchanged from the initial
pH of 6, but the pH of the V-8JA cultures increased significantly (8.3).
Final pH did not differ among isolates or environmental treatments.
The effects of temperature and culture medium on growth and
sporulation of the two isolates of D~ catenatia are presented separately
for each harvest date in tables 2.1-2.4. Means of treatments were
compared within each harvest date as well as compared with treatments
of other harvest dates. Although there were some significant main
effects, only significant interactions pertinent to the experimental
67

68
objective are presented. When there were no differences between
isolates, treatment means for the isolates were averaged. There were
significant differences in sporulation between isolates at 7 days,
but there was no isolate effect at 4, 10 or 14 days.
Colonies incubatedfot'four days (Table 2.1). Initial mycelial
growth was significantly faster, that is greater area, on PDA at 25C
2(19.1 cm ) than at any other temperature-medium combination. There
were no significant differences in colony dry weight (mg/cm2
) among
treatments. Sporulation (conidia/rnm2
) was highest at 20C on LCH
(15.5) and PDA (15.7).
Colonies incubated for seven days (Table 2~2). After seven days,
the two isolates differed significantly in dry weight and sporulation
but not in colony area. Area was greatest at 25C on PDA and V-8JA,
with the colonies completely or nearly covering the entire medium
surfaces. Dry weight of DCL was highest on PDA at l5C (2.37). However,
dry weight of DCL-2 colonies was highest on LCH and PDA at 20C, but did
not differ significantly from colonies on PDA or V-8JA at l5C.
Sporulation was also higher for DCL than DCL-2 under optimum conditions.
Sporulation of DCL was highest at 20C on LCH (33.2) and on V-8JA
(31.2). DCL-2 colonies produced more conidia on LCH at 20C (26.5)
than under other conditions.
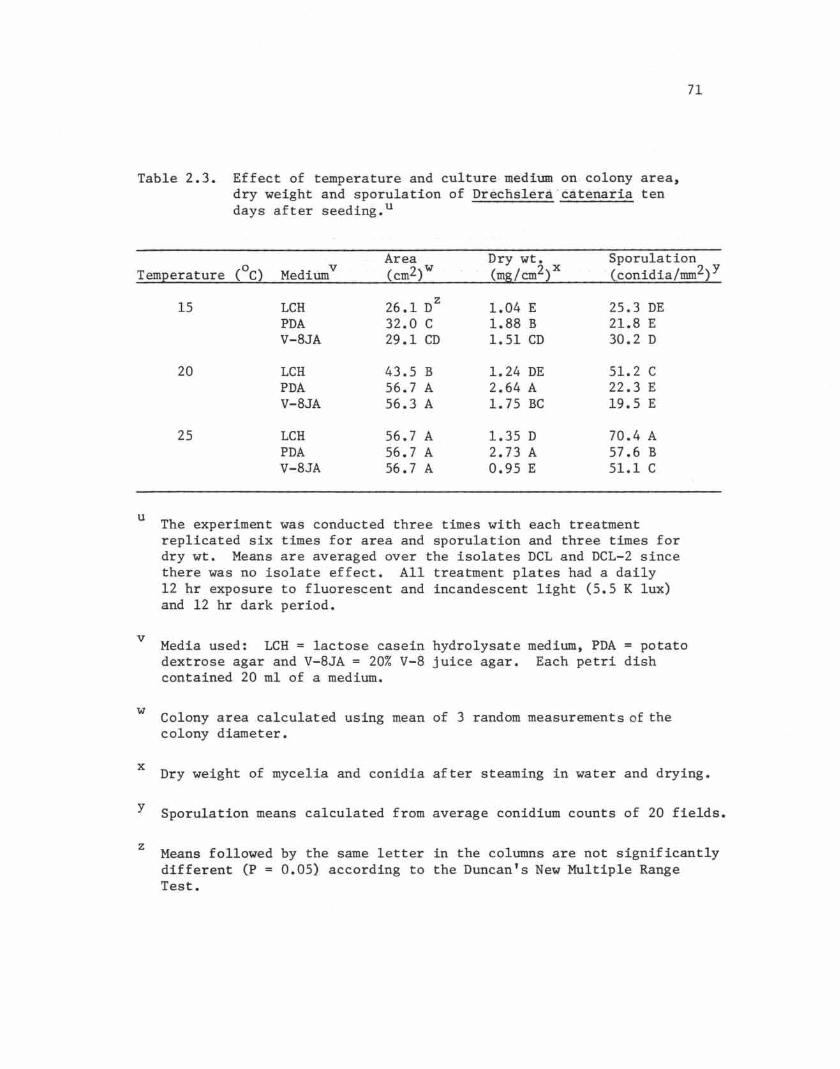
Colonies incubated for ten days (Table 2.3). Colonies had nearly
or completely covered the surfaces of PDA and V-8JA at the two highest
temperatures and on LCH at 25C. Mycelial dry weight was highest for
colonies on PDA at either 20C (2.64) or 25C (2.73). However, sporulation

69
Table 2.1. Effect of temperature and culture medium on colony area,dry weight and sporulation of'Drechsleracatenaria fourdays after seeding. u
Area Dry w~ Sporulation2Temperature (oC) MediUrilv , , , '(cm2) w (JUg/em )x (conidia/nun ) y
15 LCH 5.5 EZ0.93 A 12.5 B
PDA 7.8 D 0.88 A 6.9 CV-8JA 6.8 DE 0.92 A 7.6 C
20 LCH 3.6 F 0.96 A 15.5 APDA 7.5 D 0.71 A 15.7 AV-8JA 8.6 D 0.90 A 7.5 C
25 LCH 13.1 C 0.76 A 7.3 CPDA 19.1 A 0.82 A 8.5 CV-8JA 15.0 B 0.68 A 4.1 D
u The experiment was conducted three times with each treatmentreplicated six times for area and sporulation and three times fordry wt. Means are averaged over the isolates DCL and DCL-2 sincethere was no isolate effect. All treatment plates had a daily12 hr exposure to fluorescent and incandescent light (5.5 Klux)and 12 hr dark period.
v Media used: LCH = lactose casein hydrolysate medium, PDA = potatodextrose agar and V-8JA = 20% V-8 juice agar. Each petri dishcontained 20 ml of a medium.
w Colony area calculated using mean of 3 random measurements of thecolony diameter.
x
y
Z
Dry weight of mycelia and conidia after steaming in water and drying.
Sporulation means calculated from average conidium counts of 20 fields.
Means followed by the same letter in the columns are not significantlydifferent (P = 0.05) according to the Duncan's New Multiple RangeTest.

70
Table 2.2. Effect of temperature and culture medium on colony area, dryweight and sporulation of two isolates of Drechslera catenariaseven days ,after seeding.
u
(oC) 'MediUrilv - -Isolate'tVColony Area -Dry,Weight Sporulation
2Temperature Cem2rx ,- - 'u)'2)x (conidia/mm ) y~g em
15 LCH DCL 12.2 FZ0.93 EFGH 15.7 DEF
DCL-2 10.8 F 0.73 GH 12.6 FPDA DCL 18.1 E 2.37 A 14.7 DEF
DCL-2 15.8 EF 1.28 BCDE 13.9 EFV-8JA DCL 15.4 EF 1.54 BCD 21.2 C
DCL-2 18.1 E 1.23 CDEF 21.4 C
20 LCH DCL 19.3 E 0.76 FGH 33.2 ADCL-2 27.8 D 1.70 BC 26.5 B
PDA DCL 29.2 D 1.71 B .12.8 FDCL-2 29.9 D 1.63 BC 9.5 G
V-8JA DCL 30.9 D 1.62 BC 31.2 ADCL-2 27.4 D 1.15 DEFG 15.7 DEF
25 LCH DCL 43.8 B 0.89 EFGH 17.0 DDCL-2 36.7 C 0.52 H 16.1 DE
PDA DCL 56.7 A 1.73 B 14.2 DEFDCL-2 56.7 A 0.58 H 12.8 F
V-8JA DCL 52.4 A 0.92 EFGH 13.3 EFDCL-2 54.1 A 0.76 FGH 13.8 EF
u The experiment was conducted three times with each treatment replicatedthree times for area and sporulation and once for dry weight. Alltreatment plates had a daily 12 hr exposure to fluorescent andincandescent light (5.5 K lux) and 12 hr dark period.
v Media used: LCH = lactose casein hydrolysate medium, PDA = potatodextrose agar and V-8JA = 20% V-8 juice agar. Each petri dishcontained 20 ml of a medium.
w D. catenaria isolates: DCL = mass isolate, DCL-2 = monoconidialisolate.
x Colony area calculated using mean of 3 random measurements,of colonydiameter. Dry weight of mycelia and conidia after steaming and drying.
y
Z
Sporulation means calculated from average conidium counts of 20 fields.
Means followed by the same letter in columns are not significantlydifferent (P = 0.05) according to the Duncan's New Multiple RangeTest.
71
Table 2.3. Effect of temperature and culture-medium on- colony area,dry weight and sporulation of'Drecnslera'catenaria tendays after seeding. u
Area Dry wt. SporulationTemperature COC) Mediumv Ccm2)w CmS/cm2)x -Cconidia/mm2)y
15 LCH 26.1 DZ
1.04 E 25.3 DEPDA 32.0 C 1.88 B 21.8 EV-8JA 29.1 CD 1.51 CD 30.2 D
20 LCH 43.5 B 1.24 DE 51.2 CPDA 56.7 A 2.64 A 22.3 EV-8JA 56.3 A 1.75 BC 19.5 E
25 LCH 56.7 A 1.35 D 70.4 APDA 56.7 A 2.73 A 57.6 BV-8JA 56.7 A 0.95 E 51.1 C
u The experiment was conducted three times with each treatmentreplicated six times for area and sporulation and three times fordry wt. Means are averaged over the isolates DCL and DCL-2 sincethere was no isolate effect. All treatment plates had a daily12 hr exposure to fluorescent and incandescent light C5.5 Klux)and 12 hr dark period.
v Media used: LCH = lactose casein hydrolysate mediurn t PDA = potatodextrose agar and V-8JA = 20% V-8 juice agar. Each petri dishcontained 20 ml of a medium.
w Colony area calculated using mean of 3 random measurements of thecolony diameter.
x Dry weight of mycelia and conidia after steaming in water and drying.
y
Z
Sporulation means calculated from average conidium counts of 20 fields.
Means followed by the same letter in the columns are not significantlydifferent (P = 0.05) according to the Duncan's New Multiple RangeTest.

72
was significantly higher at 25C on LCH (70.4) than on PDA (57.6) at
the same temperature. Sporulation was also very high on V-8JA at
25C (51.1) and on LCH at 20C (51.2).
Colonies "incubated for fourteertdays (Table"2~4). Colony area
had reached maximum dimensions at all temperatures on all culture media
after two weeks, except on LCH and V-8JA at l5C. Dry weight of the
colonies was significantly higher on PDA at 20C (4.84) than on PDA
at 25C (3.65), with both differing significantly from each other and
all other treatments. Sporulation of the fungus was highest (200.1) on
LCH incubated at 20C. This treatment combination was more conducive
to conidial production than any other treatment, with the next best
sporulation on LCH at l5C (131.2).
Since mycelial dry weight and sporulation both increased with
time, greatest sporulation and maximum growth were recorded at 14 days.
When data from each harvest date were compared to one another, dry
weight of the fungus was highest on PDA incubated at 20C after 14 days.
When comparing all harvest dates, sporulation was highest for both
DCL (241.6) and DCL-2 (158.6) on LCH at 20C after 14 days, the values
differing significantly.

73
Table 2.4. Rffect of temperature and culture medium on colony area, dryweight and sporulation of Drechslera catenaria fourteen daysafter seeding. u
Area Dry.wt. SporulationTemperature (oC) Mediumv . Ccm2)w· . (mg/ cm2)X (coriidia/mm2) y
15 LCH 52.4 BZ 0.69 F 131.2 BPDA 56.7 A 2.21 C 111.1 CV-8JA 49.9 C 1.02 EF 88.3 D
20 LCH 56.7 A 1.75 CD 200.1 APDA 56.7 A 4.84 A 104.1 CV-8JA 56.7 A 1.43 DE 89.2 D
25 LCH 56.7 A 2.14 C 81.8 DPDA 56.7 A 3.65 B 56.4 EV-8JA 56.7 A 1.12 EF 46.2 E
u
v
w
x
y
Z
The experiment was conducted three times with each treatmentreplica.ted six times for area and sporulation and three times fordry wt. Means are averaged over the isolates DCL and DCL-2 sincethere was no isolate effect. All treatment plates had a daily12 hr exposure to fluorescent and incandescent light (5.5 Klux)and 12 hr dark period.
Media used: LCH = lactose casein hydrolysate medium, PDA = potatodextrose agar and V-8JA = 20% V-8 juice agar. Each petri dishcontained 20 ml of a medium.
Colony area calculated using mean of 3 random measurements of thecolony diameter.
Dry weight of mycelia and conidia after steaming in water and dry~ng.
SP?rulation means calculated from average conidium counts of 20 fields.
Means followed by the same letter in the columns are not significantlydifferent (P = 0.051 according to the Duncan's New Multiple RangeTest.

DISCUSSION
Vegetative growth and production of conidia of both isolates of
Drechslera catenaria were significantly affected by culture medium
and incubation temperature. Generally there was more rapid growth in
area at 25C and on potato-dextrose agar. Increase in mycelial dry
weight was greatest on PDA, but sporulation was generally highest on
lactose casein hydrolysate media. Both isolates of D. catenaria
sporulated by four days and continued to sporulate on all three media
at all three temperatures throughout the study. When the four harvest
times were averaged, isolates differed, with DCL generally having
higher dry weight than the monoconidial isolate (DCL-2).
Sporulation did not increase with increasing growth. Sporulation
was actually higher for those colonies having less growth. Linear
growth of fungi on agar is the least laborious method of estimating
growth, but a more accurate measurement of growth is determination of\
mycelial dry weight (Cochrane, 1958). Dry weight was higher on PDA
and V-8JA than on the LCH medium. Reproduction of fungi is thought to
be favored by depriving an established mycelium of one or more essential
nutrients or essential ones in inadequate concentrations (Cochrane,
1958). That is, fungal reproduction is most likely to occur when
vigorous mycelia exhaust their nutrients or are transferred to a medium
low in nutrients. This at times may be overridden by other factors.
74

75
It is possible that the poor growth of Q~ catertatia, especially on LCH
medium, may be the result of inadequate nutrition causing higher
sporulation in these cultures, than those on the less defined PDA and
V-8JA media.
Most plant pathogens grow best on media at an initial pH of
5.0 to 6.5 (Cochrane, 1958). All three media used in this study were
adjusted to an initial pH of 6. However by the termination of treatments,
the pH of the V-8JA cultures had become quite alkaline (ca 8.0). This
change in pH was not observed in either the LCH or PDA cultures.
Growth and sporulation of Helminthosporium and Bipolaris species have
been shown to be better at neutrality or slightly lower (Tarr and
Kafi, 1968). Conidia produced at extreme hydrogen-ion concentrations
have also been reported to vary in dimensions (Tarr and Kafi, 1968;
Harding, 1975). Growth and sporulation of D. catenaria on V-8JA may
have been affected by this change in pH. Growth, sporulation and
conidial dimensions of D. catenaria should be reinvestigated using
V-8JA with a buffer system.
Light was necessary for sporulation of both isolates of
D. catenaria, regardless of temperature or culture medium. These
results are similar to those found by Leach (1967), Shoemaker (1962)
and Zeiders (1976). The quality of light necessary for sporulation,
however, was not determined. Sporulation of H. catenarium and other
fungi has been shown to be stimulated by near ultraviolet radiation
(Leach, 1967). Leach (1962b) suggested that since fluorescent lamps
emit a small but significant amount of near-UV, much of the literature

76
reporting successful reproduction in visible light actually may have
resulted from stimulation by near-UV radiation. Bergquist et al (1972)
showed that incandescent and fluorescent lamps together give almost an
entire spectrum of light except the extremely low ranges of near-UV.
These wavelengths of light are not known to be excluded by any petri
djsh materials (Leach, 1971). Therefore, no wavelength of light could
be excluded from those thought to induce sporulation of D. catenaria.
The growth zones of Q. catenaria that formed on V-8JA could possibly
reflect an inductive reaction that is necessary for conidiogenesis,
as suggested by Leach (1967). Light is usually the primary environmental
factor causing zonation, that is, the formation of fruiting structures
in concentric rings on agar surfaces (Cochrane, 1958). Temperature
fluctuations may also induce those zonations, but in our study
temperature did not fluctuate. Regardless, Leach (1967) found
D. catenaria to be a "Constant Temperature Sporulator" that did not
need fluctuating temperatures for conidial production. The nutritional
and light requirements of the bentgrass isolate of Drechslera catenaria
in culture need to be further examined.
Although sporulation was highest after two weeks, a large number
of these conidia had already germinated or produced secondary
conidiophores from both apical and some intercalary cells. Therefore,
although harvesting from 14-day-old cultures results in higher numbers
of conidia, adequate numbers of conidia can be obtained from cultures
after only ten days. From this study, it appears that large numbers

of D. catenaria conidia can be induced to form on lactose casein
hydrolysate media by incubating cultures at 25C with exposure to
alternating light for ten days.
77





























