Embed Size (px)
Citation preview

Chapter-6
Purification and
Characterization of Laccase

Purification and Characterization of Laccase
125
CHAPTER VI
PURIFICATION AND CHARACTERIZATION OFLACCASE
6.1 INTRODUCTION
The globa l marke t for industrial enzymes is estimated at $3.3 billion in 2010. This market is
expected to reach $4.4 bi l l ion by 2015, a compound annual growth rate of 6% over the 5 -
year forecast per iod [BCC, January, 2011].The chemical processes are now carried out
biologically by biocatalysts (enzymes) which are integral components of any biological system. Some
microbes are important sources of enzymes and are preferred over sources of plant and animal origin. As
a result, more than 50% of the industrial enzymes are obtained from bacteria [Prakash et al., 2013].
Laccases are widely distributed among plants, fungi [Morozova et al., 2007], insects [Hoegger et al.,
2006] and bacteria [Claus, 2003]. To date, more than a hundred laccases have been isolated and
characterized. Most of them were derived from fungi, including especially white rot basidiomycetes,
whereas laccases from plant are less studied [Mayer and Staples, 2002; Morozova et al., 2007]. In
contrast to fungal laccases, only a few bacterial laccases have been so far studied, although recent rapid
progress in whole genome analysis suggests that these enzymes are widespread in bacteria [Sharma et
al., 2007]. Bacterial laccases have advantageous properties compared to fungal laccases with respect to
industrial applications; they are highly active and much more stable at high temperature and high pH
value than fungal laccases [Sharma et al., 2007].
Down stream processing costs are extremely important factors as these constitute upto 80 % of the
overall production cost of enzymes [Sadana and Beelaram, 1994]. For the development of downstream
process for isolating and purifying an enzyme, one of the main objectives remains to bring down the cost
required strategic selection of the purification process. The selection of process largely depends as upon
the source and nature of the targeted enzyme. Reports of laccases in Actinomycetes were based on rather
non-specific substrate reactions, but have been verified for Streptomyces griseus [Freeman et al., 1993],
S. lavendulae [Suzuki et al., 2003]. The significant objective of initial stage of purification is the
removal of large solid particles and microbial cells by centrifugation or filtration. Then supernatant is
extracted into major fraction using salts, organic solvents, pH change or through surfactants. For
purifying product various chromatographic technique are also applied.
There have been a lot of efforts to purify stable and active laccase from many different sources
such as Rhus vernicifera, Rhus succedanea, Acer pseudoplatanus, Pinus taeda, Polyporus versicolor,
Neurospora crassa, Pleutrotus ostreatus, Polyporus pinsitus, Rhizoctonia solani, Myceliophthora
thermophila, Scytalidium thermophilum, Polyporus anisoporus, Phlebia radata, Podospora anserina,

Purification and Characterization of Laccase
126
Myrothecium verrucaria, etc [Solomon et al., 1996]. Actinomycetes are believed to be the potent
producer of laccases, next to fungi. Purification and characterization of laccases from Actinomycetes,
especially, different Streptomyces sp. has been accounted. The laccase-like phenol oxidase from
Streptomyces griseus has been reported to have a highly unique homotrimer structure [Endo et al., 2003]
while the small laccase (SLAC) from Streptomyces coelicolor has been drscribed as a dimer, lacking the
second domain [Machczynski et al., 2004]. Laccase from Streptomyces lavendulae has been reported as
thermostable, being stable at 70 °C [Suzuki et al., 2003]. Laccase from Streptomyces cyaneus was
capable of oxidizing non-phenolic compounds in the presence of mediators [Arias et al., 2003].
This chapter aims to develop purification strategy for laccase recovered from Streptomyces
chartreusis strain NBRC 12753 for its kinetic and molecular characterization.
6.2 MATERIALS AND METHODS
6.2.1 Chemicals
The Sephadex G-100, ABTS and Syringaldezine were purchased from Sigma Chemical Corps.
(St. Louis, U.S.A.). The ammonium sulphate, silver nitrate, sodium carbonate, acrylamide, bis-
acrylamide, acetone, formaldehyde were procured from Qualigens, (Mumbai, India). All other chemicals
used were of analytical grade and of highest purity available.
6.2.2. Filtration of Fermented Culture Broth
After the completion of submerged fermentation (as described in Chapter 4), as a preliminary
process of down stream the fermented broth was filtered through glass filter assembly equipped with
porcelain G-5 disc and vacuum pump connection.
6.2.3 Salt Precipitation of Laccase
For efficient sustainable precipitation of produced laccase in terms of recovery of enzyme
activity and purification fold of recovered enzyme, optimization of (NH4)2SO4 saturation, optimization of
pH, optimization of temperature and optimization of precipitation time were determined.
6.2.3.1 Optimization of (NH4)2SO4 Saturation Gradient and pH for Salting out of Laccase
The aliquote of 20.0 mL of prefiltered fermented broth through G-5 glass filter was separately
placed in 17 beakers. To develop the gradient of (NH4)2SO4 saturation, different amount of anhydrous
crystalline (NH4)2SO4 (in gm) was added in each beaker as follow per 20 ml of broth content like 2.14;
2.72; 3.32; 3.94; 4.58; 5.24; 5.90; 6.61; 7.32; 8.08; 8.84; 9.66; 10.46; 11.34; 12.22; 13.18 and 14.14. For
pH optimization the aliquot of 20 ml of G-5 glass filtered fermented broth with pH value of 7.6 were

Purification and Characterization of Laccase
127
separately collected into 10 small glass beakers. The desired aliquot of 0.1 N HCl or 0.1 N NaOH was
added in each beaker to develop the pH value of 6.8, 7.0, 7.2, 7.4, 7.6, 7.8, 8.0, 8.2, 8.4 and 8.6 separately.
To develop 80 % salt saturation, 10.46 gm of anhydrous crystalline (NH4)2SO4 was added into the each
beaker and vigorously shaken to dissolve the added salt. The vigorous shaking is given to dissolve the
added salt content and then transferred individually into 17 conical polycarbon 30 ml centrifugal tubes.
The tubes were incubated at 10 º C for 12 hrs, were centrifuged at 10,000 rpm for 30 minutes , the
supernatant was decanded and 20.0 mL of phosphate buffer (7.6 pH) was added for the resuspension of
sedimented protein content. After vigorous shaking, 1.0 mL was aliquoted for the determination of
laccase activity (ABTS method) [Bourbonnais and Paice, 1990] while the remaining amount was used
to determine for protein content by Bradfoed’s test method [Bradford, 1976].
6.2.3.2 Optimization of Temperature and Precipitation Time for Salting out of Laccase
The aliquote of 20 mL of G-5 glass filtered fermented broth with pH value of 7.6 were separately
collected into 5 small glass beaker. To develop 80 % salt saturation, 10.46 gm of anhydrous crystalline
(NH4)2SO4 was added into the each beaker and vigorously shaken to dissolve the added salt. The desired
aliquot of 0.1 N NaOH was added in each beaker to develop the pH value of 8.0. The content was
transferred into 30.0 mL conical polycarbon centrifugal tubes. The tubes were incubated at 4ºC, 7ºC, 10 º
C, 13º C, 16ºC for 12 hrs separately. For optimization of precipitation time the tubes were incubated at
its optimum temperature (7 º C) for 4 hrs, 6 hrs, 8 hrs, 10 hrs, 12 hrs separately. After the incubation, the
tubes were centrifuged at 10,000 rpm for 30 minutes for the sedimentation of crystallized proteinic
content. After centrifugation from each tube the supernatant was decanted and 2.0 mL of optimized (pH
8.0) phosphate buffer was added for the resuspension of sedimented protein content. After vigorous
shaking, 1.0 mL was aliquoted for the determination of laccase activity (ABTS method) [Bourbonnais
and Paice, 1990] while the remaining amount was used to determine for protein content by Bradford’s
test method [Bradford, 1976].
6.2.4 Precipitation of Laccase by Optimized Parameters of Salting out Techniques
100 mL of G-5 glass filtered fermented broth was collected in to 250 mL conical flask. The
amount of 44.20 gm of anhydrous crystalline (NH4)2SO4 was added to develop 80 % salt saturation and
dissolved by vigorous shaking. The flask was incubated at 10º C for 12 hrs. After incubation the
developed precipitates were collected through G-5 glass filter (discarded as non-laccase protein). The
supernatant were collected in a 250 mL conical flask and the salt saturation was brought to 70 % by
adding 8.1 gm of anhydrous (NH4)2SO4. By adding appropriate amount of 0.1 N NaOH the alkalinity of
the salted filtrate was brought to pH value of 7.8. Then the flask was incubated at 7 º C for 10 hrs. After
incubation the precipitated protein was collected by centrifuging content at 10,000 rpm for 30 minutes.

Purification and Characterization of Laccase
128
The sedimented protein content was redissolved into 50 mL, 8.0 pH phosphate buffer by vigorous
shaking.
6.2.5 Desalting of Laccase Salted out by (NH4)2SO4 Saturation
At redissolution the laccase salted out by (NH4)2SO4 saturation, consist more than double
concentrated enzyme with the impurities of low molecular weight proteins of various cell metabolites as
well as trace content of salt. The prepared 100 mL laccase suspension was placed in dialysis membrane
bag with 20kda cut-off and dialysis was carried out with 1000 mL of 7.6 pH buffer at 150 rpm on
magnetic stirrer for 18 hrs in air conditioned room.
6.2.6 Gel Filtration Chromatography
The 50 mL of desalted laccase enzyme sample was loaded on sephadex G-100 column (1.5 cm ×
40 cm) equilibriated with 0.05 M, 7.8 pH phosphate buffer. The enzyme was eluted with the same buffer
and 2 mL fractions were collected at a flow rate of 0.5 mL/minutes. The laccase activity and protein
content of each fraction was determined. The fraction containing highest activity that is fractionation tube
no. 7 to 11 was mixed together yielding 10.0 mL content as purified enzyme sample and used for
characterization studies of laccase.
6.3 CHARACTERIZATION OF LACCASE FOR MOLECULAR WEIGHT
DETERMINATION
6.3.1 1D Gel Electrophoresis
50 mg each of Standard and Sample were dissolved in 200µL of Sodium Acetate buffer (pH -5).
The samples were centrifuged and supernatant was taken. 10µL of each standard and sample were loaded
onto single dimensional gel (1D gel). The gel was Silver stained after the run to visualize the bands by
silver nitrate staining method.
6.3.2 Gel Activity Staining
10µL of each standard and sample were loaded onto 1D gel in native conditions. Laccase activity
staining was performed in 2, 2’- Azino - bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt
(ABTS) as the substrate. Non-Denaturing gels (Native PAGE) were allowed to stand in 100mL of
Sodium Acetate buffer (100mM, pH -5) with 10mL of 10mM ABTS in 100mM acetate buffer pH-5 for
30 minutes in dark. Laccase activity bands were indicated by the development of green coloured bands.

Purification and Characterization of Laccase
129
6.3.3 2D Gel Electrophoresis (Sodium dodecyl sulfate polyacrylamide gel electrophoresis)
10µL of the standard and sample were diluted to 125µL with rehydration buffer and loaded on the
IEF strip 3-10pH Linear, 7cm (Biorad) and kept for Iso-Electric Focusing. After IEF run, the strips were
equilibrated in equilibration buffer and the second dimension was carried out on a 10% SDS-PAGE. The
gels were Silver stained after the run to visualize the bands. SDS-PAGE was performed to determine
sample purity and approximate molecular mass of laccase. High range (6.2 - 201.2 kDa) molecular weight
markers (Sandor Proteomics Pvt.Ltd.) were used to determine the molecular mass of purified laccase. The
markers were; 201.2kD (Myosin), 114.3 KD (β-Galactosidase), 74KD (Bovine Serum Albumin), 48KD
(Ovalbumin), 34.4KD (Carbonic Anhydrase), 27.2KD (Soybean Trypsin Inhibitor), 17KD (Lysozyme),
and 6.2 KD (Aprotinin).
6.3.4 MALDI-TOF Mass Spectrometry and Protein Identification
The sample given in 1D-gel band was trypsin digested and mixed with HCCA (a-Cyano- 4-
hydroxycinnamic Acid) matrix in 1:1 ratio and the resulting 2µl was spotted onto the MALDI plate. After
air drying the sample, it was analyzed on the MALDI TOF/TOF ULTRAFLEX III instrument and further
analysis was done with FLEX ANALYSIS SOFTWARE for obtaining the PEPTIDE MASS
FINGERPRINT. The masses obtained in PMF were submitted for Mascot search database for significant
identification of the protein.
6.3.5 Determination of pH Optima for Laccase Activity
The Tris (HYDROXYMETHYL) AMINOMETHANE (Tris-HCl) Buffer gradient was prepared
for the pH range of 9.0 to 7.2. 10 tubes were prepared, containing 500µL of 50mM ABTS, 500 µl of
enzyme solution consisting 500 unit of laccase and 4000 µL of tris-HCl buffer gradient in individual tube
yielding the pH value of tube content individually at pH 7.2, 7.4, 7.6, 7.8, 8.0, 8.2, 8.4, 8.6, 8.8, 9.0. After
appropriate shaking, the tubes were incubated at 50 °C for 15 minutes. The inactivation of enzyme was
carried out by placing the tubes in boiling water for 10 minutes. The enzyme assay was carried out by
U.V/Visible photospectrometry (Make: Jasco corporation, Spectra manager version 1.53.00 [Build 1]).
6.3.6 Determination of Temperature Optima and Thermal Stability for Laccase Activity
The numbers of 8 tubes were prepared. Each tube contained 500µL of 50mM ABTS, 4000 µL of
phosphate buffer at pH value 7.8 and 500 µL of enzyme solution consisting 500 unit of laccase.
Immediately after appropriate mixing the tube was individually incubated at 30 °C, 35° C, 40° C, 45° C,
50 °C, 55° C, 60° C, 65° C for 15 minutes. The inactivation of enzyme was carried out by placing the
tubes in boiling water for 10 minutes. The thermal stability was determined under same assay conditions

Purification and Characterization of Laccase
130
after preincubation of laccase for 1,5,10,15,20,24 and upto 48 hrs at optimum temperature, pH and buffer
concentration. The enzyme assay was carried out by U.V/Visible photospectrometry.
6.3.7 Effect of Various Metal Ions on Laccase Enzymatic Reaction
3 tubes were prepared containing 500µL of 50mM ABTS, 3500 µL of phosphate buffer at pH
value 8.0, and to determine the acceleration of enzyme reaction 500 µL of 10 mM of Cobalt (Co),Copper
(Cu), Mercury (Hg)), Magnesium (Mg), Manganeze (Mn) and Zinc (Zn) in individual tubes. 500 µL of
enzyme solution consisting 500 unit of laccase was added in each tubes. After appropriate mixing the
tubes were incubated at 50 °C for 15 minutes. The inactivation of enzyme was carried out by placing the
tubes in boiling water for 10 minutes. The enzyme assay was carried out by U.V/Visible
photospectrometry.
6.3.8 Effect of Inhibitor on Laccase Enzymatic Reaction
3 tubes were prepared containing 500µL of 50mM ABTS, 3500 µL of phosphate buffer at pH
value 8.0, and to determine the deccelaration of enzyme reaction, 500 µL of 1.0 mM of Sodium azide, L-
cystine, Thiourea, EDTA and Thioglycolic acid in individual tubes. 500 µL of enzyme solution consisting
500 unit of laccase was added in each tubes. After appropriate mixing the tubes were incubated at 50º C
for 15 minutes. The inactivation of enzyme was carried out by placing the tubes in boiling water for 10
minutes. The enzyme assay was carried out by U.V/Visible photospectrometry.
6.3.9 Determination of Substrate Concentration on Laccase Activity for Kinetic Parameters
Three substrates such as ABTS, Guaiacol and Syringaldezine were used for determination of
substrate concentration on laccase activity. 10 tubes were prepared containing 500 µL of enzyme solution
containing 18.84 U (0.1 mg laccase having 184.4 specific activity), 4000 µL of phosphate buffer at pH
value 8.0 and 500 µL of substrate was added to form periodic substrate concentration gradient from 10
mM to 60 mM per 5000 µL of reaction content. After appropriate mixing the tubes were incubated at 50
°C for 15 minutes. The inactivation of enzyme was carried out by placing the tubes in boiling water for 10
minutes. The enzyme assay was carried out by U.V/Visible photospectrometry. Km and Vmax value of
pure laccase were determined by measurement of laccase activity with various concentrations of (10 mM
-60 mM) ABTS, Guaiacol and Syringaldezine. Kinetic constant Km and Vmax were calculated from the
Lineweaver bulk plot with substrate concentration. The Kcat was obtained with equation: Vmax – Kcat
[Et], where [Et] is the total enzyme concentration.

Purification and Characterization of Laccase
131
6.3.10 Absorption Spectrum of Laccase
The UV- visible spectrum of the purified laccase was determined at wavelengths between 200-
700 nm at room temperature using a UV-visible spectrophotometer (Make: Jasco corporation with spectra
manager version 1.53.00 [Build 1], at University school of sciences, Department of Chemistry, Gujarat
university, Ahmedabad, Gujarat-India.)
6.4 RESULTS AND DISCUSSION
6.4.1 Filtration of Fermented Culture Broth
The filtration through G-5 porcelin glassed sintered funnel under vacuum had yielded transparent
light yellow brownish solution. At analysis the filtrate had the potency of 400 units of laccase and protein
content of 32.25 mg/mL with specific activity of 12.4 was observed as single fold purification with 100 %
yield of enzyme content.
6.4.2 Optimization of (NH4)2SO4 Saturation Gradient and pH for Salting out of Laccase
To optimize (NH4)2SO4 concentration, 20mL filtrate with 400 U/mL, were taken in each tube and
salt saturation was developed to have gradient from 20 to 100 saturation. Under standardized down stream
condition, 75 % salt saturation had given precipitation of 176 U/mL and then at 80 % salt saturation
precipitation was observed with maximum recovery of 268 U/mL (Fig. 6.1) which remain more or less
same at higher salt saturation. At 80 % salt saturation maximum activity of laccase could be recovered
with the loss of 132 U/mL, yielding 67.0 % recovery of produced enzyme.
Fig. 6.1 Optimization of (NH4)2SO4 saturation gradient
Purification and Characterization of Laccase
132
Khammuang and Rakrudee (2009) purified laccase from fruiting bodies of Ganoderma sp. MK05 by
ammonium sulphate precipitation with 40-70 % saturation and obtained 1.34 fold from Stereum ostrea
with 80 % ammonium sulphate supported our result in the present investigation [Khammuang and
Rakrudee, 2009].
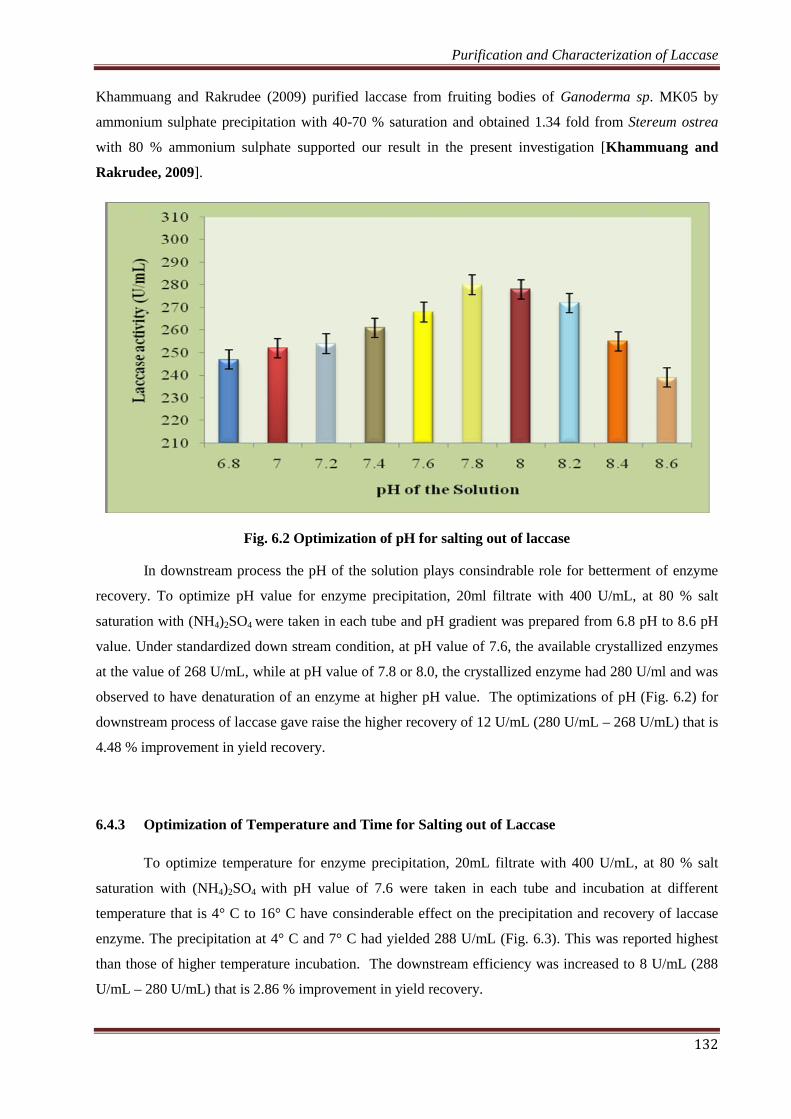
Fig. 6.2 Optimization of pH for salting out of laccase
In downstream process the pH of the solution plays consindrable role for betterment of enzyme
recovery. To optimize pH value for enzyme precipitation, 20ml filtrate with 400 U/mL, at 80 % salt
saturation with (NH4)2SO4 were taken in each tube and pH gradient was prepared from 6.8 pH to 8.6 pH
value. Under standardized down stream condition, at pH value of 7.6, the available crystallized enzymes
at the value of 268 U/mL, while at pH value of 7.8 or 8.0, the crystallized enzyme had 280 U/ml and was
observed to have denaturation of an enzyme at higher pH value. The optimizations of pH (Fig. 6.2) for
downstream process of laccase gave raise the higher recovery of 12 U/mL (280 U/mL – 268 U/mL) that is
4.48 % improvement in yield recovery.
6.4.3 Optimization of Temperature and Time for Salting out of Laccase
To optimize temperature for enzyme precipitation, 20mL filtrate with 400 U/mL, at 80 % salt
saturation with (NH4)2SO4 with pH value of 7.6 were taken in each tube and incubation at different
temperature that is 4° C to 16° C have consinderable effect on the precipitation and recovery of laccase
enzyme. The precipitation at 4° C and 7° C had yielded 288 U/mL (Fig. 6.3). This was reported highest
than those of higher temperature incubation. The downstream efficiency was increased to 8 U/mL (288
U/mL – 280 U/mL) that is 2.86 % improvement in yield recovery.

Purification and Characterization of Laccase
133
Fig. 6.3 Optimization of temperature for salting out of laccase
To optimize incubation time for enzyme precipitation, 20mL filtrate with 400 U/mL, at 80 % salt
saturation with (NH4)2SO4 with pH value of 7.6 and incubated at 7° C for different time period that is 4
hrs to 12 hrs have consinderable effect on the precipitation and recovery of laccase enzyme. The
incubation period of 10 hrs is proved sufficient for maximum recovery of 299 U/mL as compared to
insufficient recovery of product under shorter period of incubation. The prolonged further incubation had
no significant rise in recovery process. By optimization of incubation period the downstream efficiency
was increased to 11 U/mL (299 U/mL – 288 U/mL) (Fig. 6.4) that is 3.81 % improvement in yield
recovery.
Fig. 6.4 Optimization of time for salting out of laccase

Purification and Characterization of Laccase
134
6.4.4 Precipitation of Laccase by Optimized Parameters of Salting out Techniques
The G-5 filtered fermented broth [having Laccase Activity of 400 U/mL with 12.4 specific
activity] was treated as 1 fold purification with 100 % yield recovery. The Laccase salting out of 100 ml
fermented broth adjusted to 7.8 pH and 80 % salt saturation with (NH4)2SO4.incubated at 7 °C for 10 hrs
had yielded optimized precipitation of laccase. The available crystallized enzyme, at re-suspension into
50mL of 7.8 pH phosphate buffer had yielded enzyme solution with the recovery of 30460 Unit of laccase
and resulted as 609 U/mL, with 64.48 specific activities. During Salt precipitation with (NH4)2SO4 there
was a loss of 9540 unit of laccase probably due to inactivation of enzyme in salty environment. The
process was resulted efficient for removing 2752 mg of non enzymatic proteinious fraction and
improvement in specific enzyme activity to 64.48 U/mg which was resulted into 5.2 fold of enzyme
purification with 76.15 % of yield recovery (Table 6.1).
6.4.5 Desalting of Laccase Salted out by (NH4)2SO4 Saturation
The desalting of 50mL laccase solution available from fractionization by (NH4)2SO4 saturation, at
dialysis through membrane bag -20kda cut-off against 1000 mL of 7.6 pH Buffer at 150 rpm on magnetic
stirrer for 18 hrs in air conditioning room proved satisfactory for the removal of salt content as well as
law molecular weight cell metabolites and biodegraded proteinous content of fermentation medium. The
process had removed 58.0 mg of low molecular weight proteinous impurities and thus improves the
specific activity to 71.20 U/mg with 5.90 purification fold and 73.87 % of yield recovery (Table 6.1)
6.4.6 Gel Filtration Chromatography
Loading of desalted 10mL of laccase solution, at sephadex G-100 column, fractionized into 2 mL
of fraction at the rate of 0.5mL/min, resulted into maximum enzyme recovery in fraction no. 7 with 2060
U/mL and gradually declined to 1810 U/mL in fraction no. 11. There was sudden drop in enzyme activity
from test tube no.11 onward and were discarded for better quantifying of fractionized enzyme product.
The proteins in mg/ml and laccase activity in U/mL were shown in Fig. 6.5. The content of fraction no 7
to 11 were mixed togheter, yielding 10.0 mL of fractionized purified laccase content having with 1032
U/mL potency with specific activity of 184.2 U/mg (Table 6.1).
As per Table 6.1 protein content in mg/ml and loss of protein were detrmined stepwise with
measurement of laccase activity in Unit/mg. Purification fold from fermented broth to Sephadex G 100
treatment were optimized and found 14.8 fold with 51.6 % yield.

Purification and Characterization of Laccase
135
Fig. 6.5 Gel filtration fraction number v/s Laccase yield & Protein
(Where = Protein in mg/ml and for Laccase activity in U/mL)
Table 6.1 Kinetic profile of downstream process for laccase
No. Treatment Volume
in mL
Unit
/mL
Total
units
Protein
mg/mL
Total
protein
mg
Specific
activity
U/mg
Loss
unit
Loss
protein
Purifi
cation
fold
Yield
(%)
1 Fermented
broth
100 400 40,000 32.25 3225 12.4 Nil Nil 1 100
2 (NH4)2SO4
Precipitation
50 609 30460 9.46 473 64.48 9540 2752 5.2 76.15
3 Dialysis 50 591 29550 8.3 415 71.20 910 58 5.90 73.87
4 Sephadex
G-100
10 1032 10320 5.6 56 184.2 8910 303 14.8 51.6

Purification and Characterization of Laccase
136
6.4.7. Characterization of the Purified Laccase by SDS PAGE, Activity Staining and Isoelectric
Focusing
The expected laccase spot from 2D gel are marked as per Fig. 6.6A. Activity staining was
performed with ABTS resulted in a single band and the result is shown in Fig. 6.6A. The homogeneity of
the purified laccase was indicated by SDS-PAGE, where one single band could be detected,
corresponding to approximately 34 kDa (Fig.6.6B). Fig 6.6B indicates the sample fraction, with different
fractions of molecular weight markers.
The results was very much similar with some alkaline laccase molecular masses such as
Streptomyces psammoticus laccase (43 Kda) [Niladevi et al., 2008], alkaline laccase Ssl1 from
Streptomyces sviceus (33 Kda) [Gunne and Urlacher, 2012], and extracellular laccase from
Thermobifida fusca (24.7 Kda) [Chen et al., 2013]. Laccase activity was observed in native condition.
The isoelectric point (PI) of the purified laccase was found in the range between 5.5-6.5. MALDI analysis
and Mascot search tool database were used for significant identification of the protein resulting as
LAC1_THACU laccase with mass of 64679 with score of 26 and 6 query matches (Fig. 6.7).

Purification and Characterization of Laccase
137
Fig. 6.6(A) Activity staining
Laccase
Fig. 6.6B Molecular mass of purified lacasse from Streptomyces chartreusis strain NBRC 12753
74.1KD201.2KD
114.3KD
48KD
34.4KD
27.2KD
17KD
6.4KD
74.1KD
Sample (Laccase)Standard (ABTS)

Purification and Characterization of Laccase
138
MASCOT SEARCH PAGE:
Fig. 6.7 MS Spectra o laccase Mascot search page
Laccase
Purification and Characterization of Laccase
139
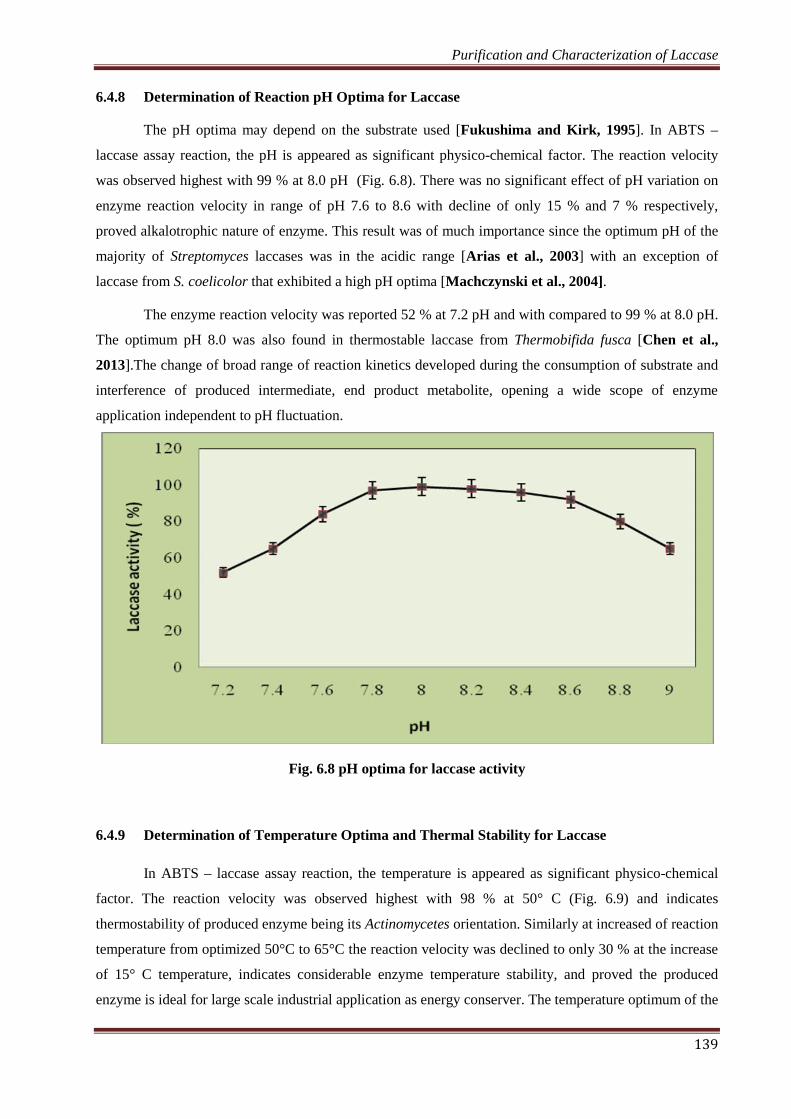
6.4.8 Determination of Reaction pH Optima for Laccase
The pH optima may depend on the substrate used [Fukushima and Kirk, 1995]. In ABTS –
laccase assay reaction, the pH is appeared as significant physico-chemical factor. The reaction velocity
was observed highest with 99 % at 8.0 pH (Fig. 6.8). There was no significant effect of pH variation on
enzyme reaction velocity in range of pH 7.6 to 8.6 with decline of only 15 % and 7 % respectively,
proved alkalotrophic nature of enzyme. This result was of much importance since the optimum pH of the
majority of Streptomyces laccases was in the acidic range [Arias et al., 2003] with an exception of
laccase from S. coelicolor that exhibited a high pH optima [Machczynski et al., 2004].
The enzyme reaction velocity was reported 52 % at 7.2 pH and with compared to 99 % at 8.0 pH.
The optimum pH 8.0 was also found in thermostable laccase from Thermobifida fusca [Chen et al.,
2013].The change of broad range of reaction kinetics developed during the consumption of substrate and
interference of produced intermediate, end product metabolite, opening a wide scope of enzyme
application independent to pH fluctuation.
Fig. 6.8 pH optima for laccase activity
6.4.9 Determination of Temperature Optima and Thermal Stability for Laccase
In ABTS – laccase assay reaction, the temperature is appeared as significant physico-chemical
factor. The reaction velocity was observed highest with 98 % at 50° C (Fig. 6.9) and indicates
thermostability of produced enzyme being its Actinomycetes orientation. Similarly at increased of reaction
temperature from optimized 50°C to 65°C the reaction velocity was declined to only 30 % at the increase
of 15° C temperature, indicates considerable enzyme temperature stability, and proved the produced
enzyme is ideal for large scale industrial application as energy conserver. The temperature optimum of the

Purification and Characterization of Laccase
140
laccase from Trametes versicolor was 50 °C with ABTS as a substrate in citrate-phosphate buffer of pH
3.0 [Han et al., 2005].
Fig. 6.9 Temperature (in °C) optima for laccase activity
Thermal stability study with the purified laccase was conducted at 30° C, 35° C, 40° C, 50° C,
60° C for 48 hrs. Laccase was found quite stable at 30° C and 35 °C to retain 78 % and 61 % activity
respectively (Fig. 6.10 A,B). While at 40 °C, 50 °C, 60 °C half life of its activity was observed upto 10
hrs, 7 hrs and 3 hrs respectively (Fig. 6.10 C, D and E).
Cont..
A B

Purification and Characterization of Laccase
141
Fig. 6.10 Thermal stability of purified laccase at 30 °C [A] , 35 °C [B], 40 °C[C], 50 °C [D]
at 60 °C [E]
It has been distinguished that stability of laccase at elevated temperature improved under nutral
pH conditions [Min et al., 2001]. Temperature ranges for high enzyme activity are narrow in many
laccases [Shin and Lee, 2000; Min et al., 2001; Iyer and Chattoo, 2003]. Laccase from Trametes
versicolor showed high relative activity and high stability over a broad temperature range [Han et al.,
2005].
C D
E

Purification and Characterization of Laccase
142
6.4.10 Effect of Metal Ions on Laccase Activity
Table 6.2 summarizes the effect of various metal ions on the activity of purified laccase. The
enzyme activity was enhanced by metal ions such as Cu and Zn (each at 10 mM and 100 mM) while the
heavy metals like Hg and Co reduced the activity considerably, even at low concentration (10 mM) in
comparision of other metal ions. The most obvious stimulating effect on laccase activity was exerted by
Cu. Significant enhancement of enzyme activity was also observed with Zn. Mg and Mn were found to
enhance the activity almost at the same level.
Table 6.2 Effect of metal ions on purified laccase from Streptomyces chartreusis strain NBRC 12753
Metal ions Concentration in mMa Relative activityb %
Control None 100
Cobalt (Co+2)
10 21.23
100 6.78
Copper (Cu+2)
10 125.67
100 102.34
Mercury (Hg+2)
10 11.89
100 5.65
Magnesium (Mg+2)
10 103.21
100 95.60
Manganeze (Mn+2)
10 102.89
100 95.00
Zinc (Zn+2)
10 120.31
100 101.11
(a- Final concentration of different metal ions were decided specifically for each of the compoundb -Values represent the means of values from three independent experiments, with a maximalsample mean deviation of + 5%)
The laccase activity was highly sensitive to Hg+2, indicating the presence of thiol group, essential for its
activity. However, the present result on the effect of Zn+2 on the purified laccase were found to be similar
to the earlier report of Streptomyces psammoticus [Niladevi et al., 2008]. The relative activity of
Cladosporium cladosporioides were stimulated by metal ions such as Cu +2 (109 %) and Mg +2 (119 %)
[Aslam et al., 2012].

Purification and Characterization of Laccase
143
6.4.11 Effect of Inhibitors on Laccase Activity
Table 6.3 represents the effects of putative inhibitors on purified laccase from Streptomyces
chartreusis strain NBRC 12753. The enzyme was inhibited by all the inhibitors that were studied.
However, the level of inhibition assorted significantly with the nature and concentration of the tested
inhibitors. Absolute inhibition of laccase activity was observed with sodium azide at the considered
concentration. Sodium azide is a common inhibitor of metalloproteins and laccase inhibition by this
compound has been well recognized.
Table 6.3 Effect of inhibitors on purified laccase from Streptomyces chartreusis strain NBRC 12753
Inhibitors Concentration in mMa Inhibitionb %
Control None 100
Sodium azide 1 100
L-cysteine 1 100
Thiourea 1 55
Thioglycollic acid 1 100
EDTA 1 100
(a Final concentration of different metal ions were decided specifically for each of the compoundb Values represent the means of values from three independent experiments, with a maximalsample mean deviation of + 5%)
Sodium azide has been statemented to prevent the substrate oxidation by laccase [Johannes and
Majcherczyk, 2000]. EDTA is an inhibitor of mettalo – enzymes including laccases due to its property of
forming inactive complexes with inorganic prosthetic groups/ cofactors of the enzyme [Sadhasivam et
al., 2008]. Similar results have also been reported from Streptomyces cyaneus [Arias et al., 2003]. The
sulfhydryl organic compound containing L-cysteine shows complete inhibition of laccase.The
thioglycollic acid also inhibits the laccase activity at same concentration while thiourea was found to be
comparatively less inhibitory to the enzyme. Similar results were found for thioglycollic acid at a same
concentration cause 100 % laccase inhibition in Chaetomium thermophilium [Chefetz et al., 1998]
6.4.12 Determination of Substrate Concentration on Laccase Activity for Kinetic Parameters
The substrate specificity studies indicated that Syringaldezine is the most suitable substrate for
laccase from Streptomyces chartreusis strain NBRC 12753 (Table 6.4). The higher specificity towards

Purification and Characterization of Laccase
144
Syringaldezine at pH 8.0 has already been reported from Streptomyces sviceus [Gunne and Urlacher,
2012]. ABTS was the next preferred substrate for the enzyme. The specificity towards ABTS was much
higher than guaiacol.This might be due to the effect of assay pH on the particular substrate. All the assays
were carried out at the same pH (7.5), which may not be suitable for all substrates.
Table 6.4 Kinetic parameters of purified laccase determined from double reciprocal plot
Substrates Km
(mM)
Vmax
(µM/mL/minute)
Kcat
(Sec-1)
Kcat/Km
(mM-1.Sec-1)
ABTS 5.55 434.78 12318.8 2219.6
Guaiacol 1.20 500 14166.7 11805.6
Syringaldezine 12.5 526.31 14912.1 1193.0
The kinetic parameter Km was determined with three different substrates ABTS, guaiacol and
syringaldazine. The enzyme showed affinity towards syringaldezine with a Km value of 12.5 mM. The
Km value with guaiacol and ABTS was 1.20 mM and 5.55 mM respectively. The high Km value obtained
for syringaldazine indicated that the enzyme had less affinity towards this substrate. The Vmax value for
syringaldazine was 526.31µM/mL/minute. The Vmax values corresponding to ABTS and guaiacol were
434.78 and 500 µM/ml/min respectively. The Km and Vmax values for different substrates of laccase
from Streptomyces chartreusis strain NBRC 12753 were determined by Lineweaver-Burk plot (Fig. 6.11
A, B and C). The affinity of the purified enzyme towards different substrates were in the order; Guaiacol
> ABTS > syringaldazine and the results of the kinetic parameters were very much in conformity with the
substrate specificity analysis.
Cont..
A

Purification and Characterization of Laccase
145
Fig. 6.11 Lineweaver-Burk plots for Guaiacol (A), ABTS (B) and Syringaldezine (C) for
determination of Km and Vmax
The ratio Kcat/Km gives an indication of each enzyme and therefore allows us to compare the
efficiencies of the different enzymes towards different substrates. Table 6.4 shows that ABTS is the most
efficiently oxidized by laccase as it exhibits the highest values for Kcat/Km ratio for all the substrates
studied. The highest value of Kcat/Km was found for ABTS in laccase from Pleorotus floride
[Satishkumar and Palvannan, 2013] and laccase from Coriolopsis rigida [Saparrat et al., 2002].
6.4.13 Spectral Studies of Laccase
The absorption spectrum of laccase was studied in the range of 200 to 800 nm (Fig. 6.12). The
maximum absorption at around 600 nm is a characteristic of type 1 copper of a typical laccase. The
spectral graph indicated that the purified laccase belongs to the group of well established blue-laccases of
fungi and bacteria. Almost same results were found in alkaline laccase from Streptomyces psammoticus
C
B

Purification and Characterization of Laccase
146
[Niladevi et al., 2008] and alkaline laccase Ssl1 from Streptomyces sviceus [Gunne and Urlacher,
2012].
Fig. 6.12 Absorbance spectra of laccase
6.5 CONCLUSION
Laccases are versatile biocatalysts for industrial applications. Much research has been done with
fungal laccases; however, recently more information has become available also for bacterial laccases. The
laccase recovered from Streptomyces chartreusis strain NBRC 12753 was purified subjecting to the
ammonium sulphate precipitation and gel filtration chrometrography using Sepadex G100 and finally
about 15 fold purification was achieved. The purified laccase from Streptomyces chartreusis strain NBRC
12753 had a low molecular weight around 34 Kda as compared to other laccases produced from bacteria.
It differed from other laccases in a few aspects such as the low specificity towards syringaldazine and
activity enhancement by Zinc ion. The inhibition study carried out with some specific inhibitors like
EDTA and Sodium azide further confirms the production of laccase. The enzyme showed very high
affinity towards ABTS and exhibited wide substrate range. The effect of different inhibitors on the
purified enzyme conformed to the general pattern of laccase inhibition. The present study has identified,
purified and characterized an alkaline laccase from an Actinomycete, Streptomyces chartreusis. The
present work has made a new attempt for the identification of similar enzymes from this group of
organisms. The purified laccase showed the presence of only type I copper atom which was charetcterized
from its absorbance around 600 nm. The relatively high activities at alkaline pH and at 50 °C make
suitable for industrial applications.





























