Embed Size (px)
Citation preview

Chapter 5 Organization and Expression of Ig Genes
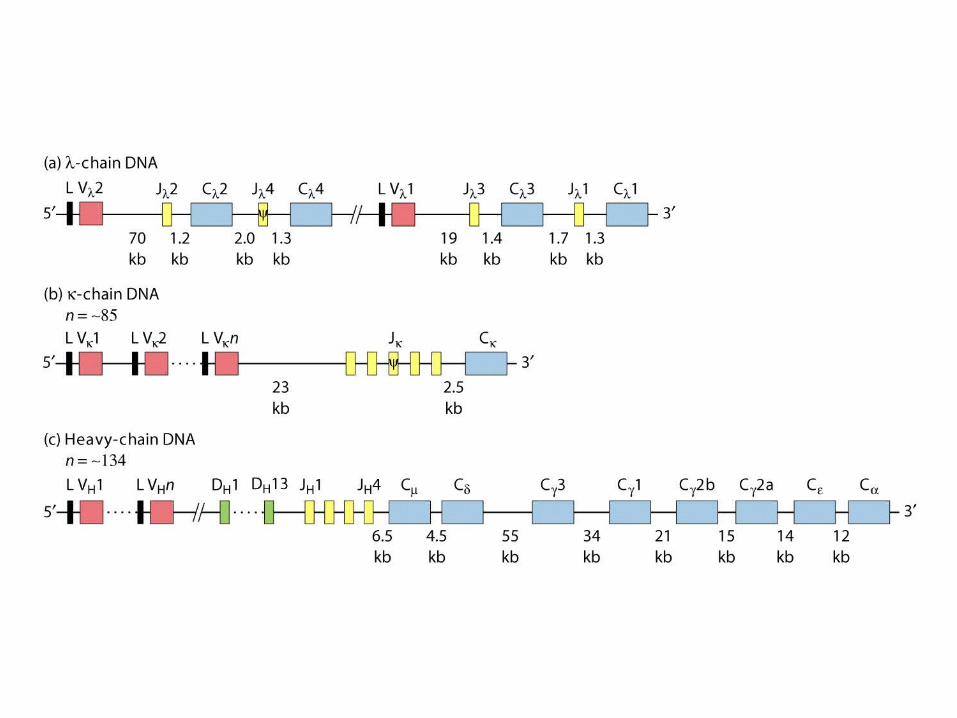
chain
chain (n= ~85)
H chain (n= ~134)

Unique features of Ig genes (1)
- Vertebrates can respond to a limitless array of foreign proteins.
- Every Ab molecule contains a unique a.a.sequence in its V region, but only one of a limited number of invariable sequences in its C region.
- Germ-line DNA contains multiple gene segments encode portions of a single Ig H or L chain.

Unique features of Ig genes (2)
- The Ig gene segments carried in the germ cells can’t be transcribed & translated into H & L chains until they are rearranged into functional genes.
- During B-cell maturation in the bone marrow, Ig gene segments are rearranged and generated into more than 1010 combinations of V region.
- Each B cell has a unique combination and is antigenically committed to a specific epitope.

Unique features of Ig genes (3)
- Mature B cells no longer contain identical chromosomal DNA to germ-line DNA.
- After antigenic stimulation, further rearrangement of C-region gene segments can generate changes in isotypes without changing the specificity of Ig. - Genomic rearrangement is an essential feature of lymphocyte differentiation, and no other vertebrate cell type has been shown to undergo this process.

本章大綱 :• Genetic model compatible with Ig structure• Multigene organization of Ig genes• V-region gene rearrangements• Mechanism of V-region DNA rearrangements• Generation of Ab diversity• Class switching among C-region genes• Expression of Ig genes• Regulation of Ig-gene transcription• Ab genes and Ab engineering

Genetic Model Compatible with Ig Structure


• The vast diversity of antibody specificities
• The presence in Ig heavy and light chains of a variable region at the amino-terminal end and a constant region at the carboxyl-terminal end
• The existence of isotypes with the same antigenic specificity.

Germ-line theory: The genome contributed by the germ cells, egg and sperm, contains a large repertoire of Ig genes.
Somatic-variation theory:
The genome contains a small number of Ig genes, from which a large number of Ab specificities are generated in the somatic cells by mutation or recombination.

- How could stability be maintained in the C region while some kind of diversifying mechanism generated the V region?
- There must be mechanisms not only for generating Ab diversity but also for maintaining constancy.
- Neither the germ-line nor the somatic variation theory could offer a reasonable explanation of the central feature of Ig structure.

The Two-gene model of Dryer and Bennett (1965)
Two separate genes encode a single Ig H or L chain, one gene for the V region and the other for the C region.

The suggestion that two genes encoded a single polypeptide contradicted the existing one gene-one polypeptide principle and was without precedent in any known biological system.

Verification of the Dryer and Bennet Hypothesis (by Tonegawa and Hozumi, 1976)
First direct evidence that separate genes encode the V and C regions of Ig and that the genes are rearranged in the course of B-cell differentiation.
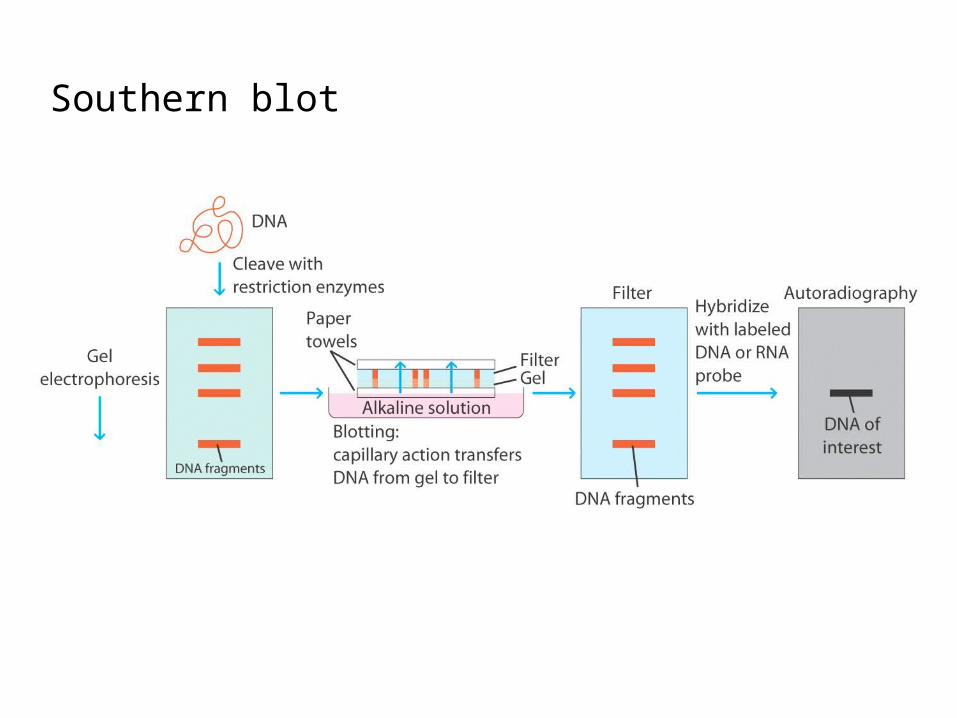
Southern blot
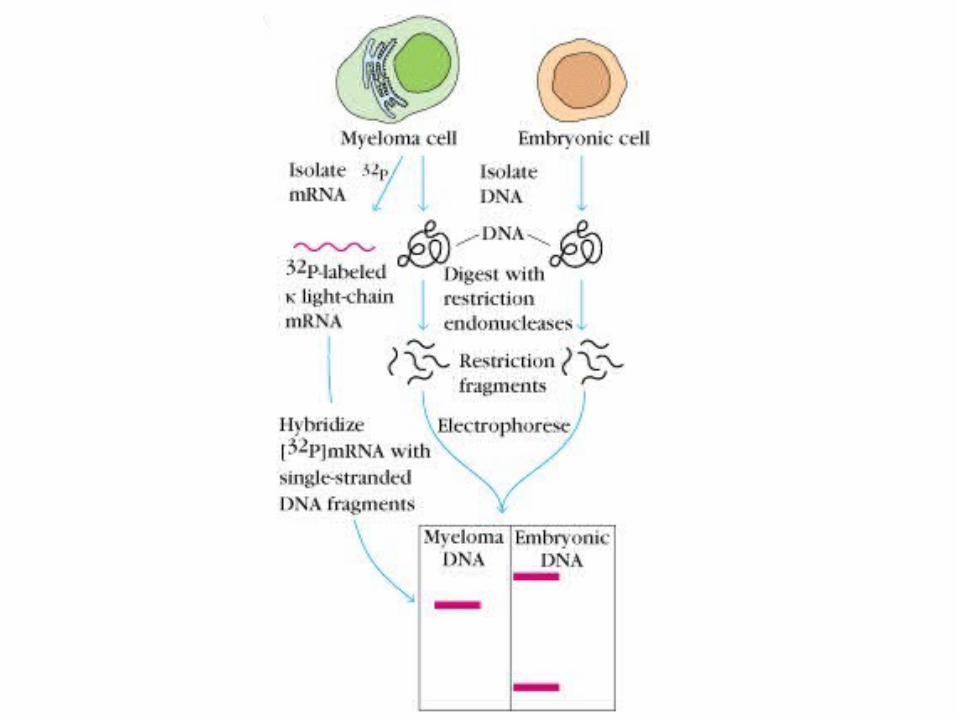


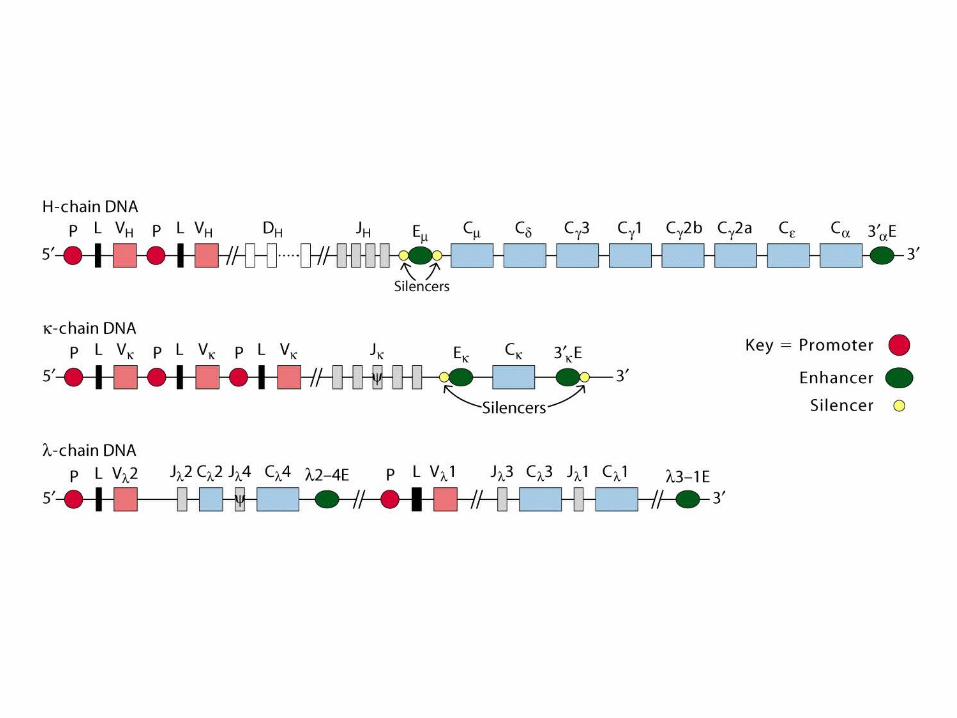
Multigene organization of Ig genes

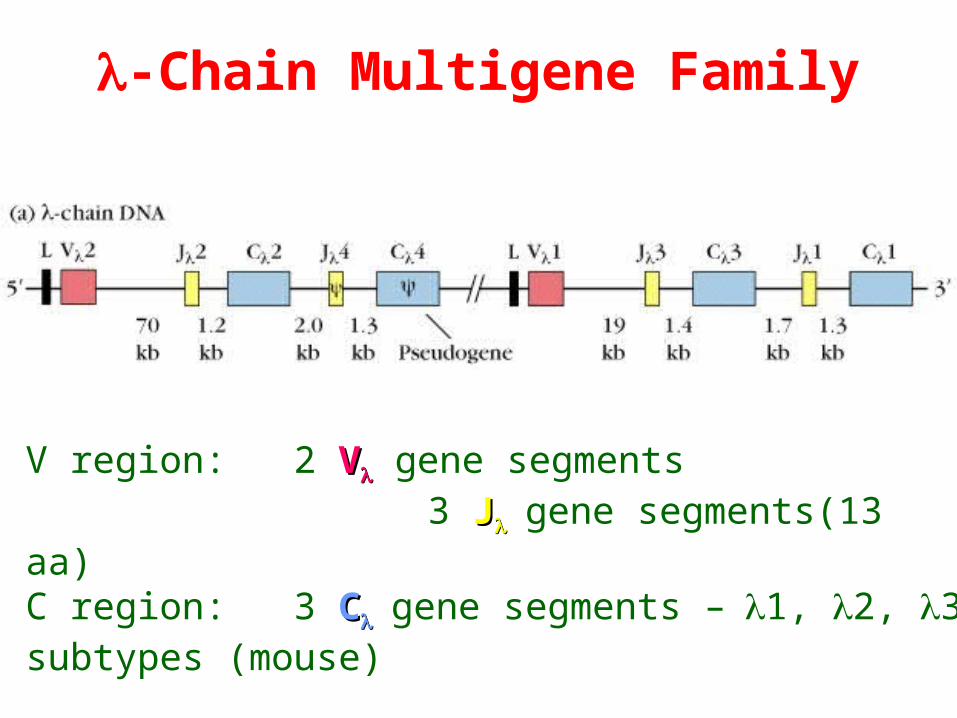
-Chain Multigene Family
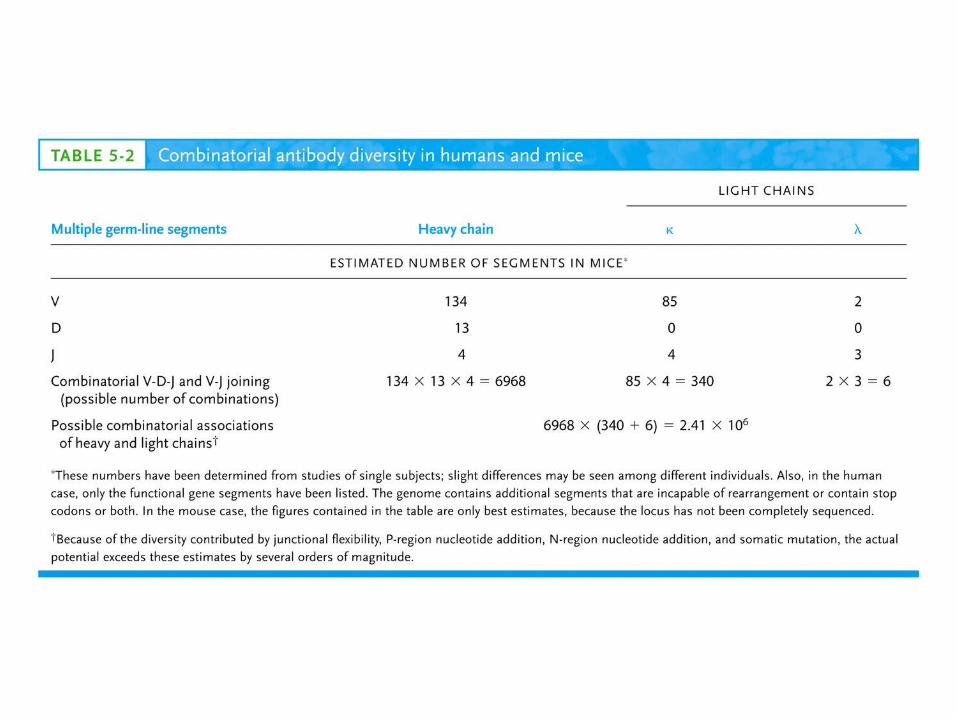
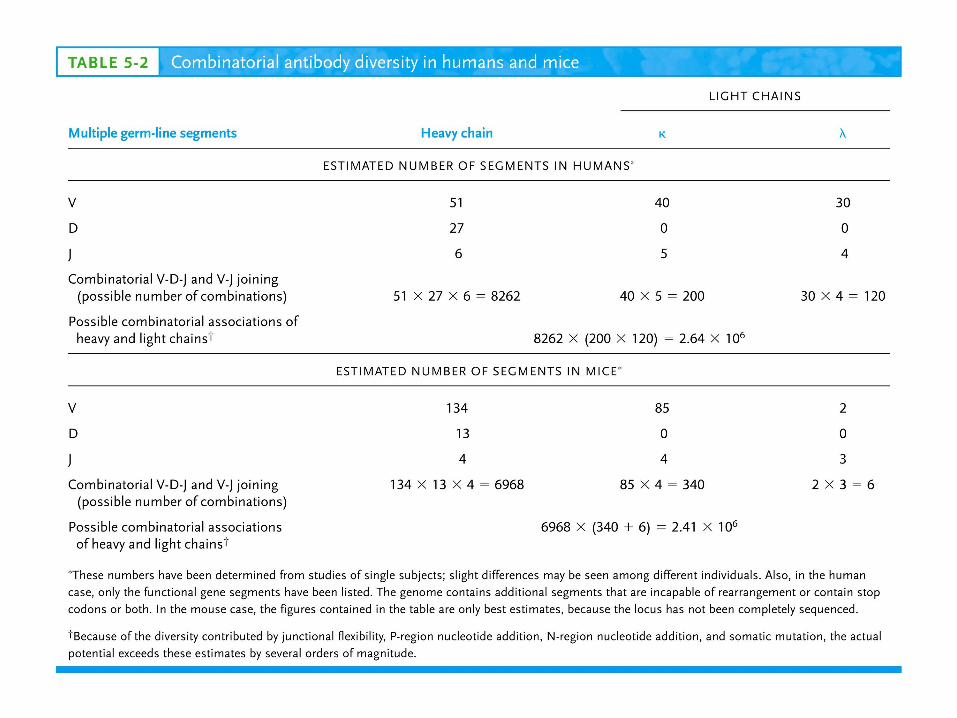
V region: 2 VV gene segments 3 JJgene segments(13 aa)C region: 3 CCgene segments –1, 2, 3 subtypes (mouse)
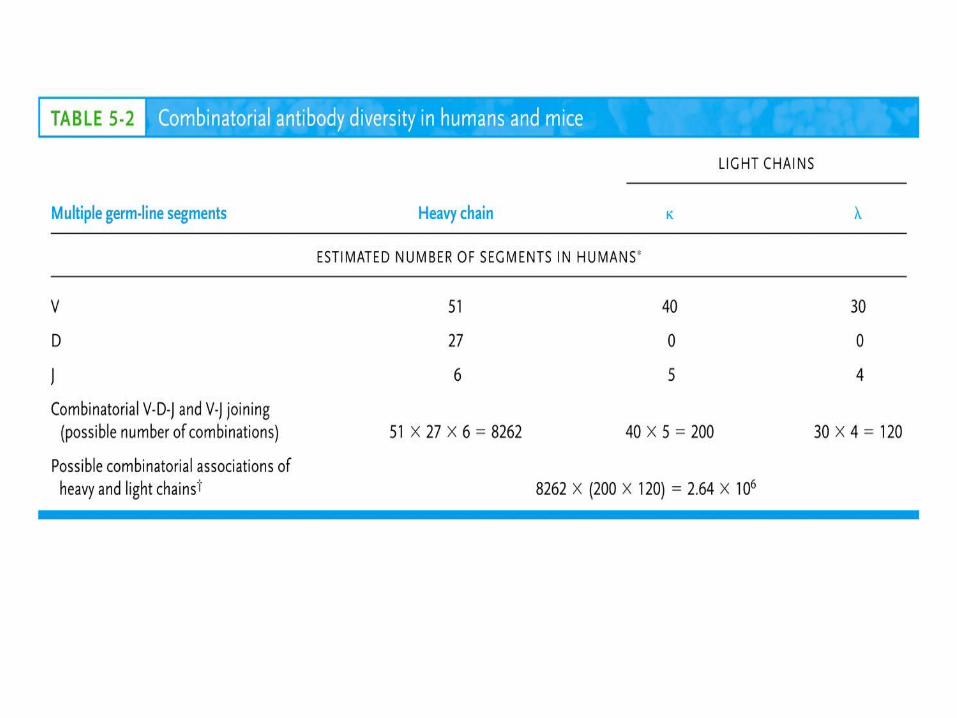
In humans: 30 VV, 4 JJ and 4 CC segments
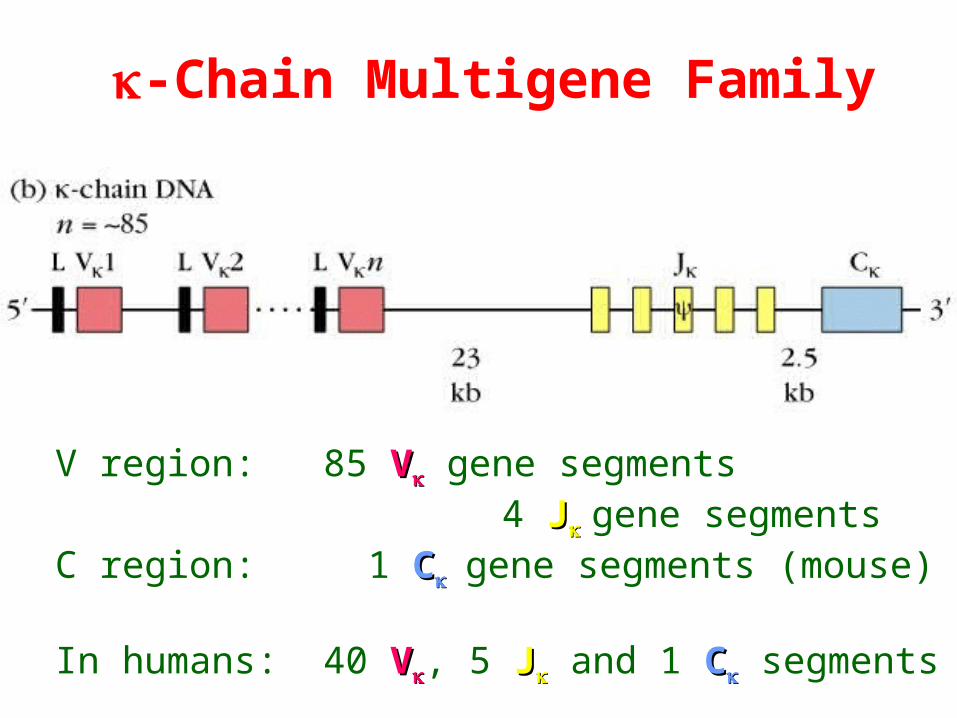
-Chain Multigene Family
V region: 85 VV gene segments 4 JJgene segmentsC region: 1 CCgene segments (mouse)
In humans: 40 VV, 5 JJ and 1 CC segments
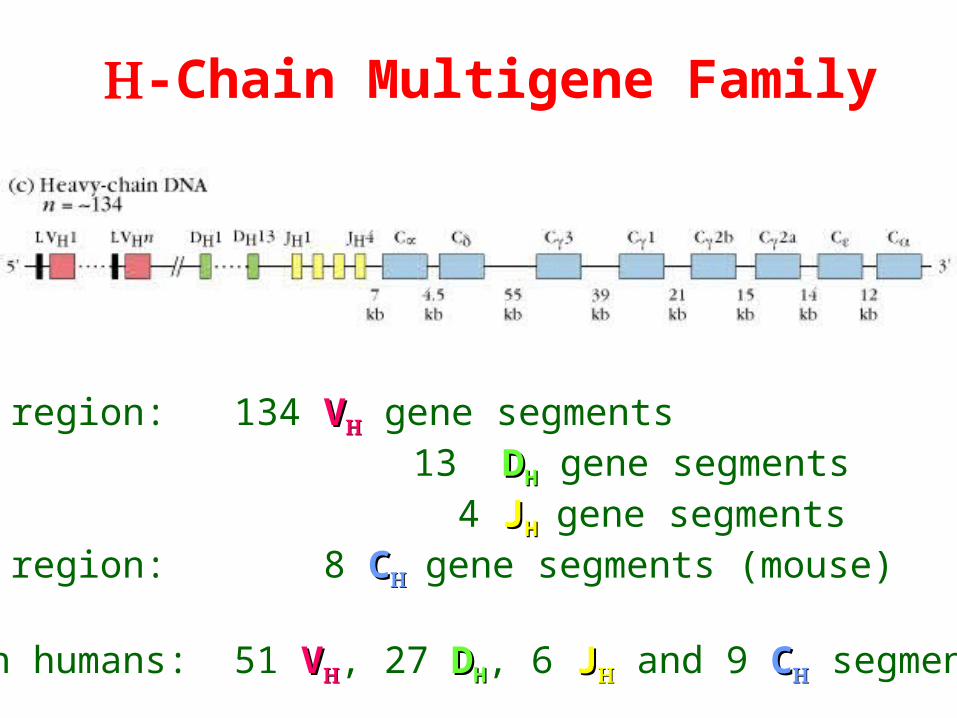
-Chain Multigene Family
V region: 134 VV gene segments 13 DDHH gene segments 4 JJHHgene segmentsC region: 8 CCgene segments (mouse)
In humans: 51 VV, 27 DDHH, 6 JJ and 9 CC segments

V-region gene rearrangements

V Region gene rearrangements
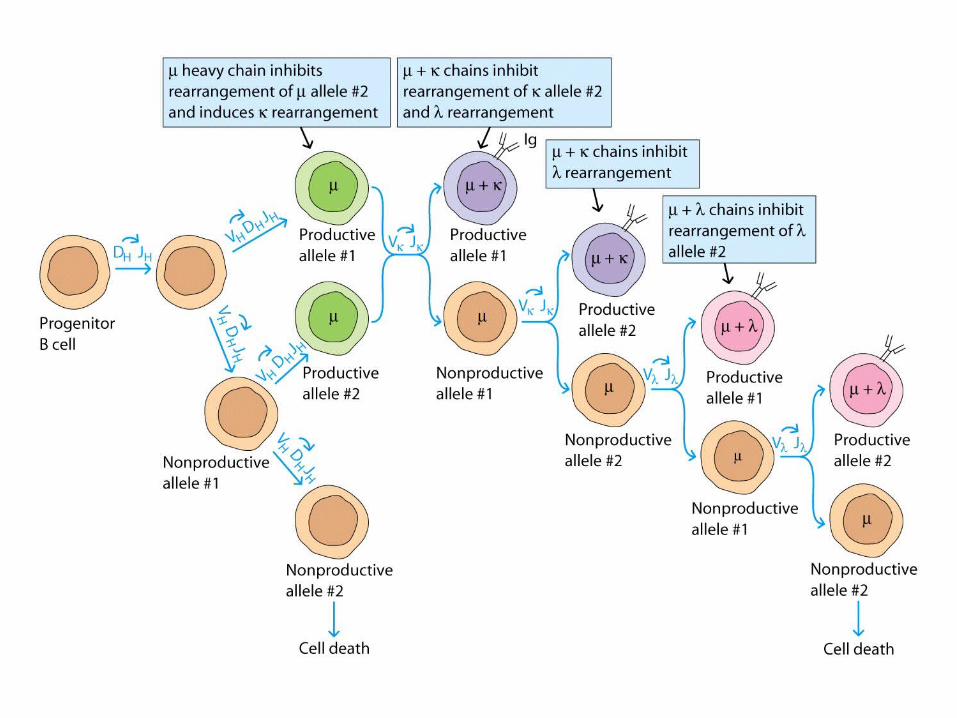
- The H-chain V-region genes rearrange first, then the L-chain V-region genes.
- The rearrangements are random events
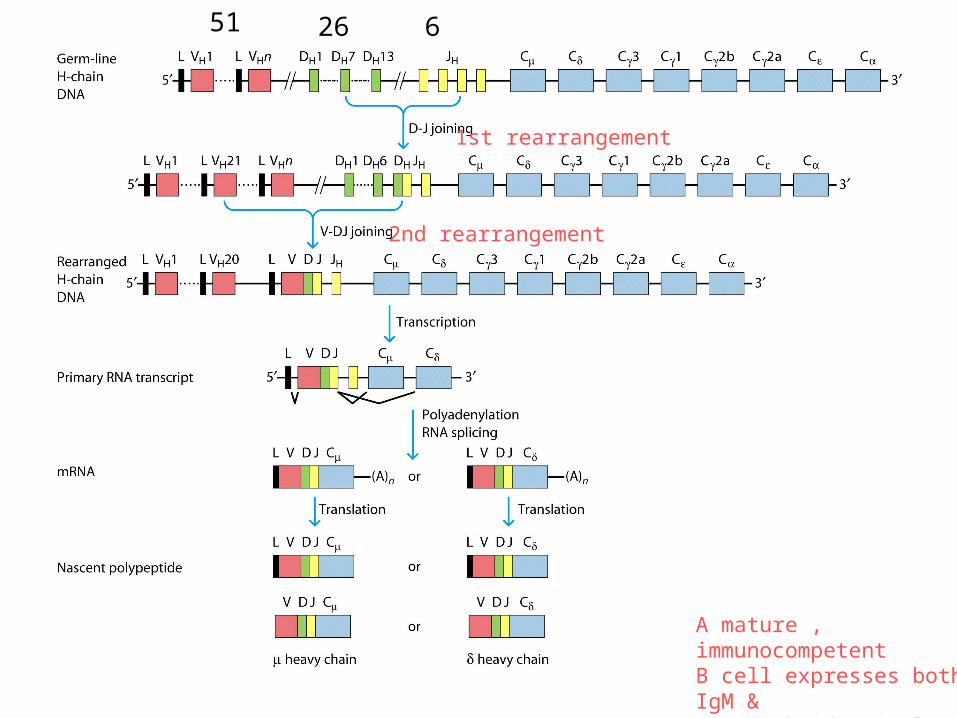
1st rearrangement
2nd rearrangement
A mature , immunocompetentB cell expresses both IgM & IgD with identical antigenic specificity on its surface.
51 26 6


Mechanism of V-region DNA rearrangements
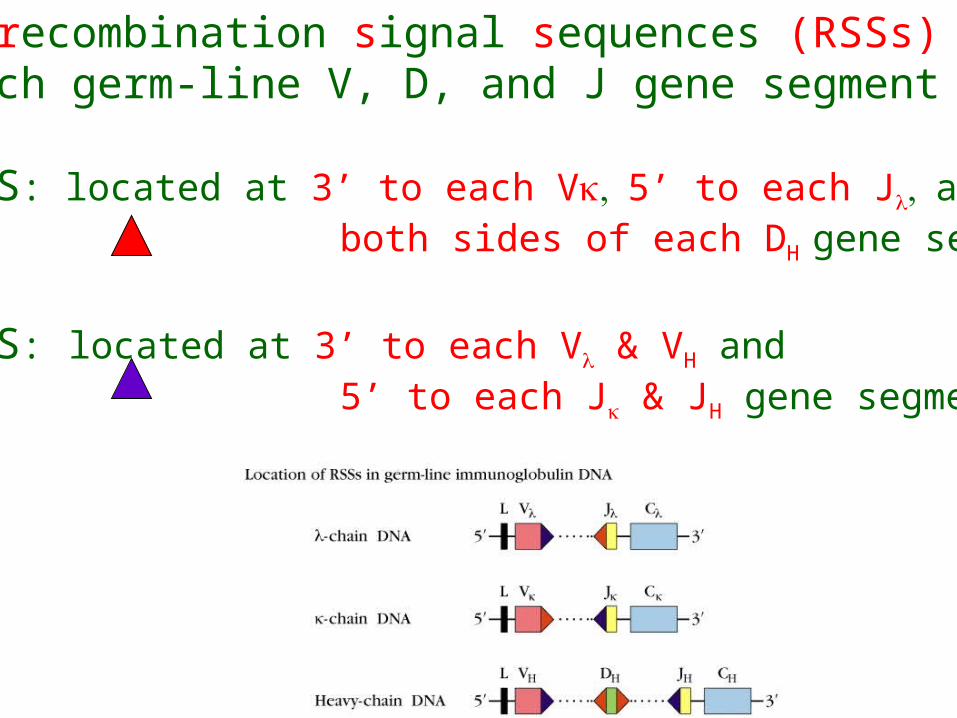
Two unique recombination signal sequences (RSSs) flanking each germ-line V, D, and J gene segment
One-turn RSS: located at 3’ to each V5’ to each Jand both sides of each DH gene segment
Two-turn RSS: located at 3’ to each V & VH and 5’ to each J & JH gene segment
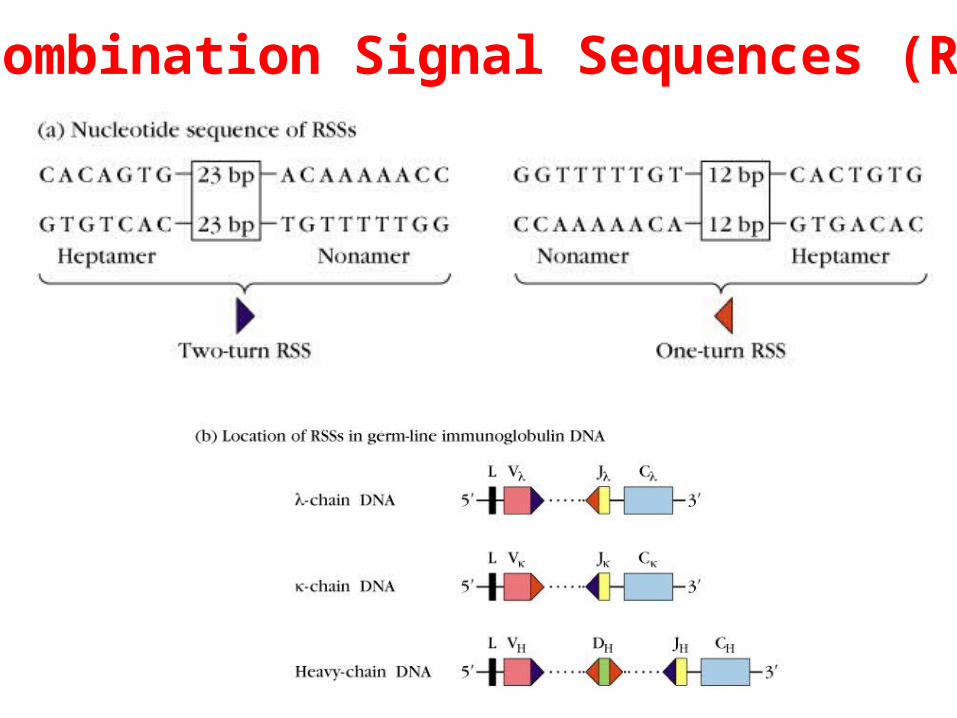
Recombination Signal Sequences (RSS)

One turn/two-turn joining rule
- Signal sequences having a one-turn spacer (12 bp) can join only with sequences having a two-turn spacer (23 bp) (one-turn/two turn joining rule).
- This joining rule ensures that a VL segment joins only to a JL segment and not to another VL segment. - The rule likewise ensures that VH, DH, and JH segments join in proper order and that segments of the same type do not join each other.

Enzymatic Joining of Gene Segments
Recombination-Activating Genes: RAG-1, RAG-2 - mediate V-(D)-J joining
TdT: terminal deoxynucleotidyl transferase
DSBR: double strand break repair enzymes
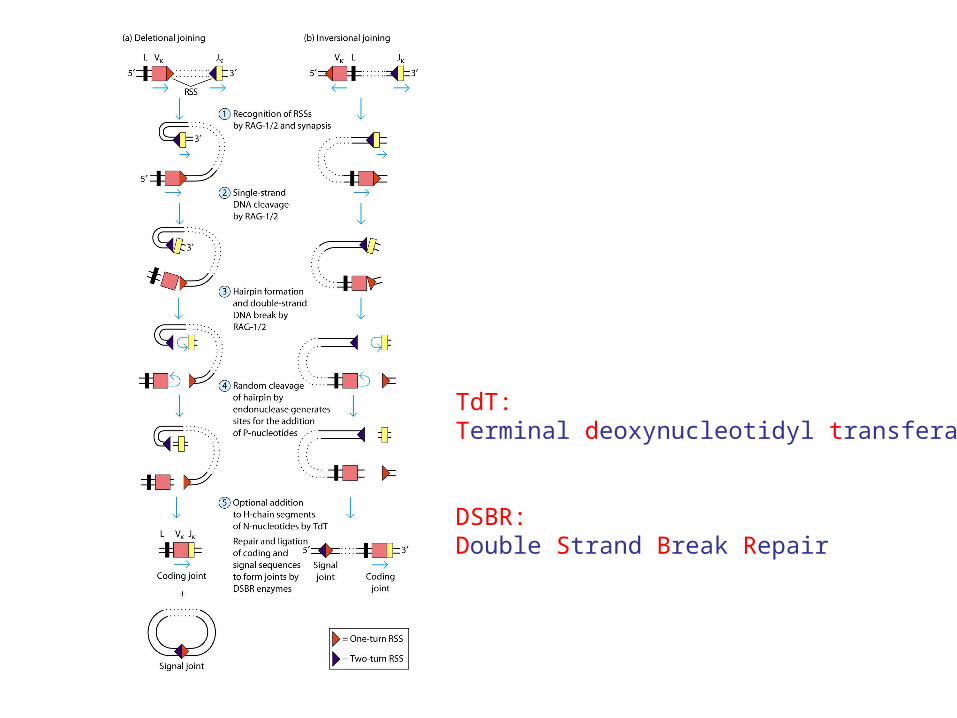
TdT:Terminal deoxynucleotidyl transferase
DSBR:Double Strand Break Repair

Deletional joining (coding joint): -two gene segments are in the same transcriptional orientation
Inversional joining (signal joint):- two gene segments have opposite orientation
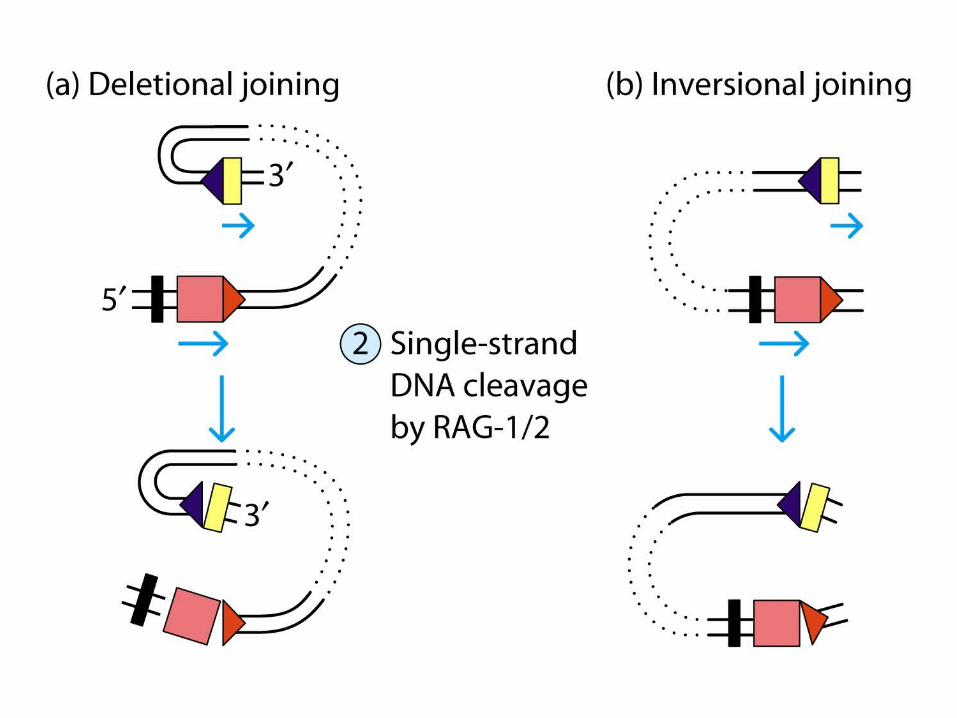
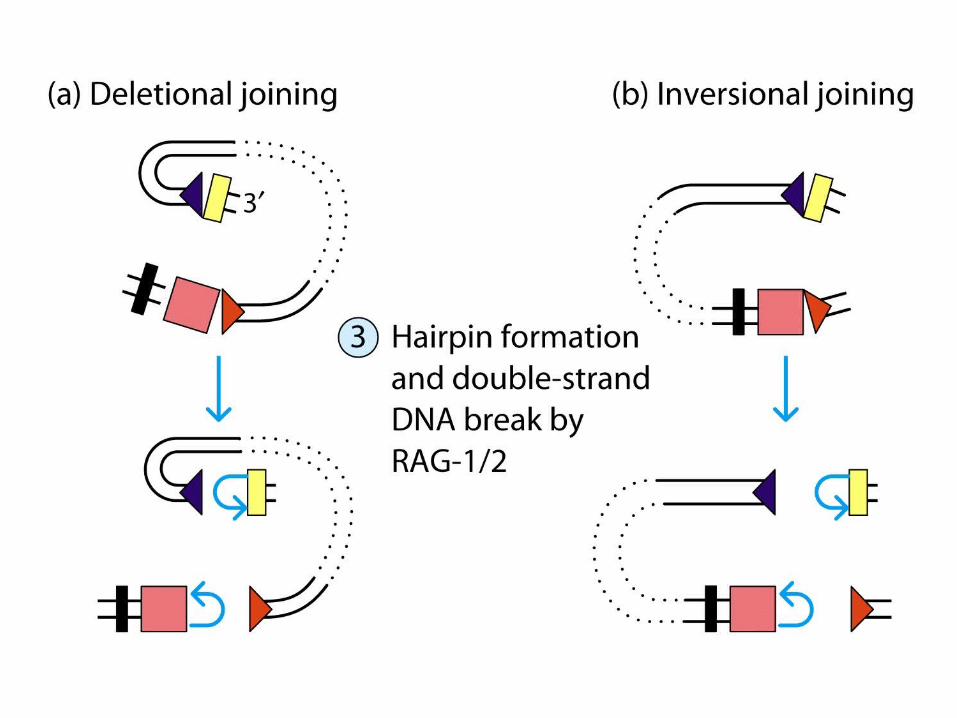
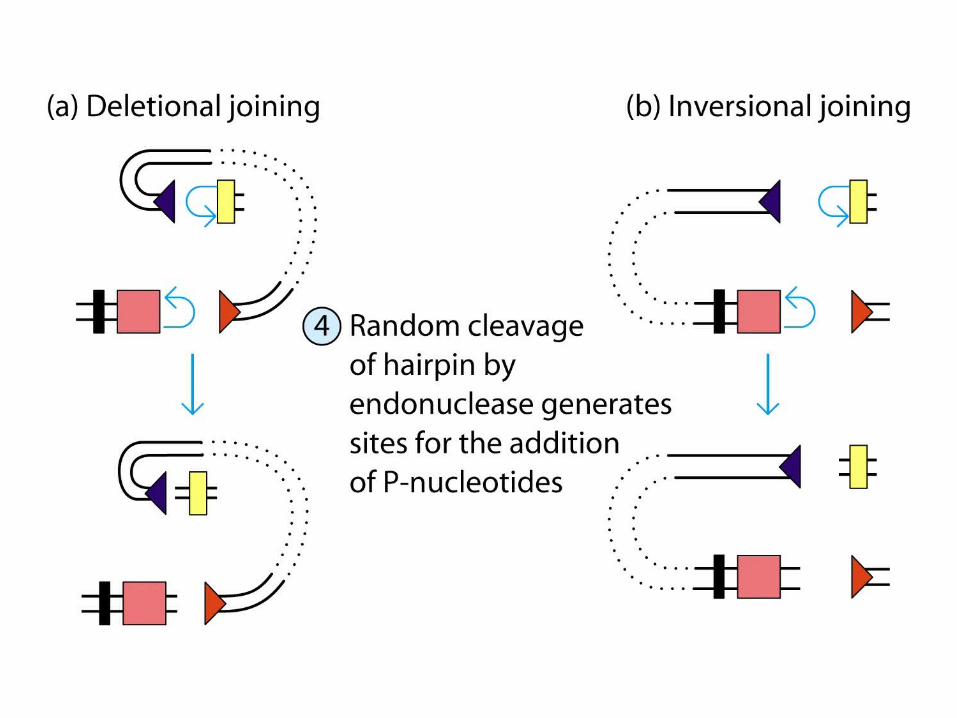
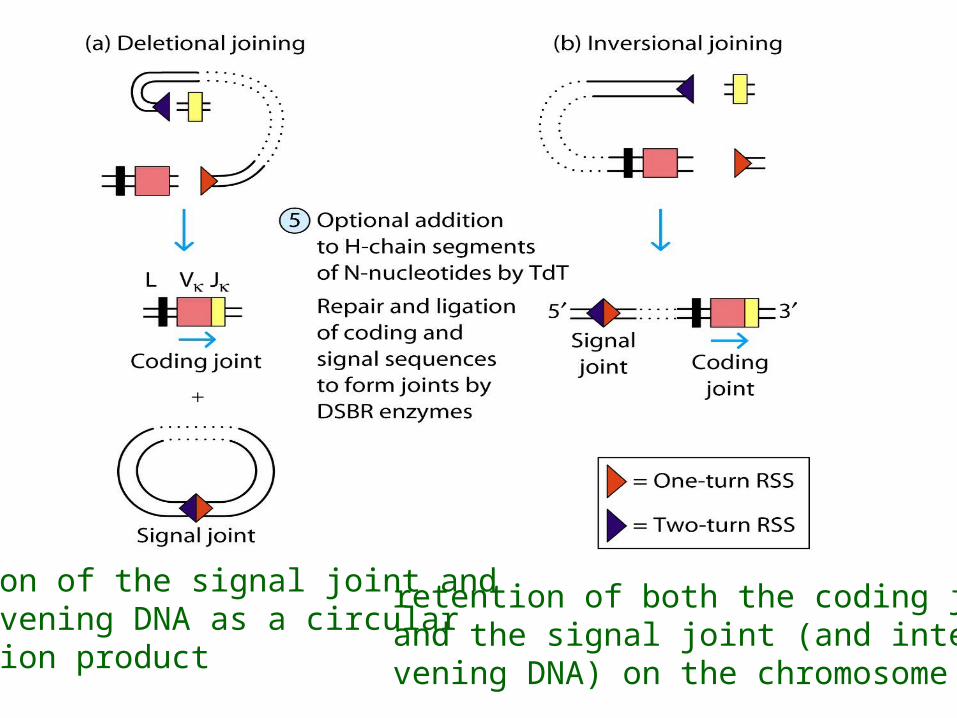
deletion of the signal joint and intervening DNA as a circular excision product
retention of both the coding joint and the signal joint (and inter-vening DNA) on the chromosome
<15 nt

Homework:
If a mouse has a defect on RAG-1 or 2 , what will happen?
If you knock out TdT or DSBR enzyme from a mouse, what will happen?
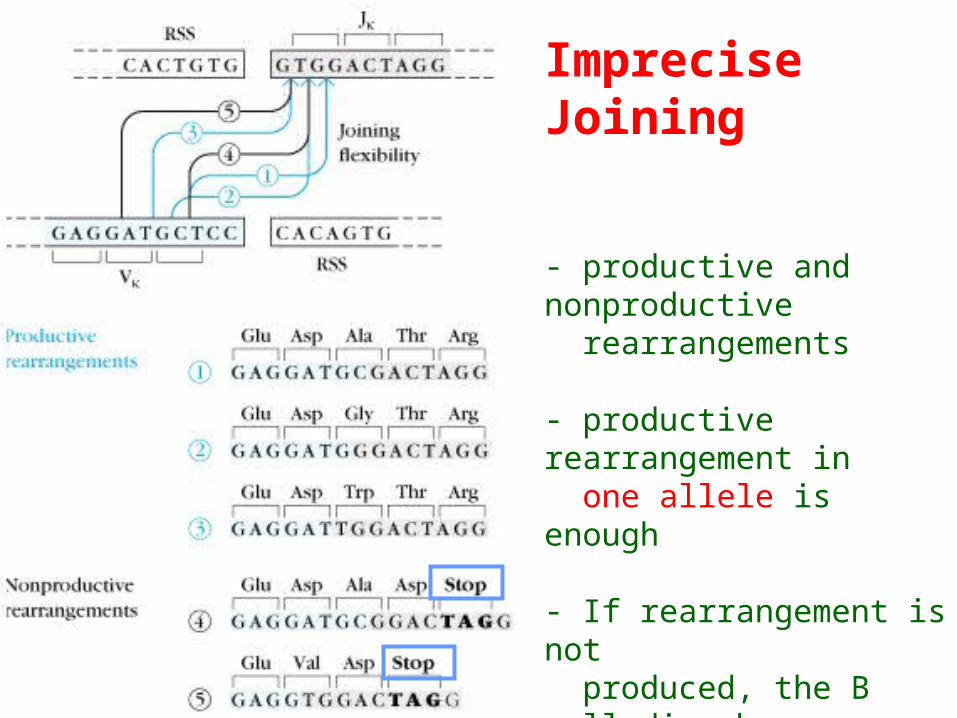
Imprecise Joining
- productive and nonproductive rearrangements
- productive rearrangement in one allele is enough
- If rearrangement is not produced, the B cell dies by apoptosis.
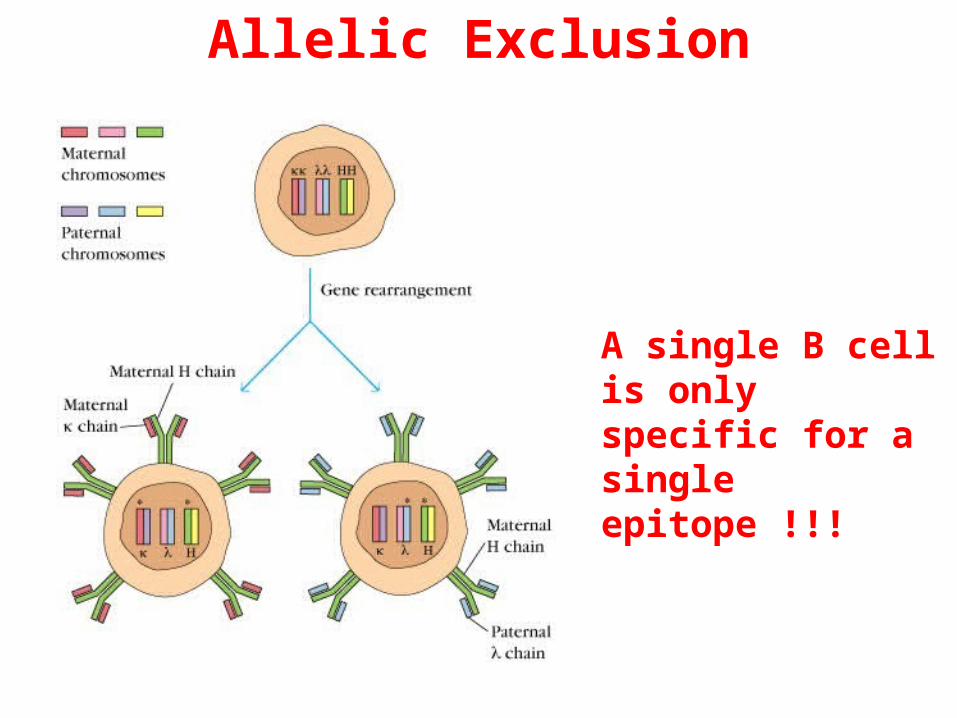
Allelic Exclusion
A single B cell is only specific for a single epitope !!!

Generation of Ab diversity

Antibody Diversity
Seven means of generation of Ab diversity:
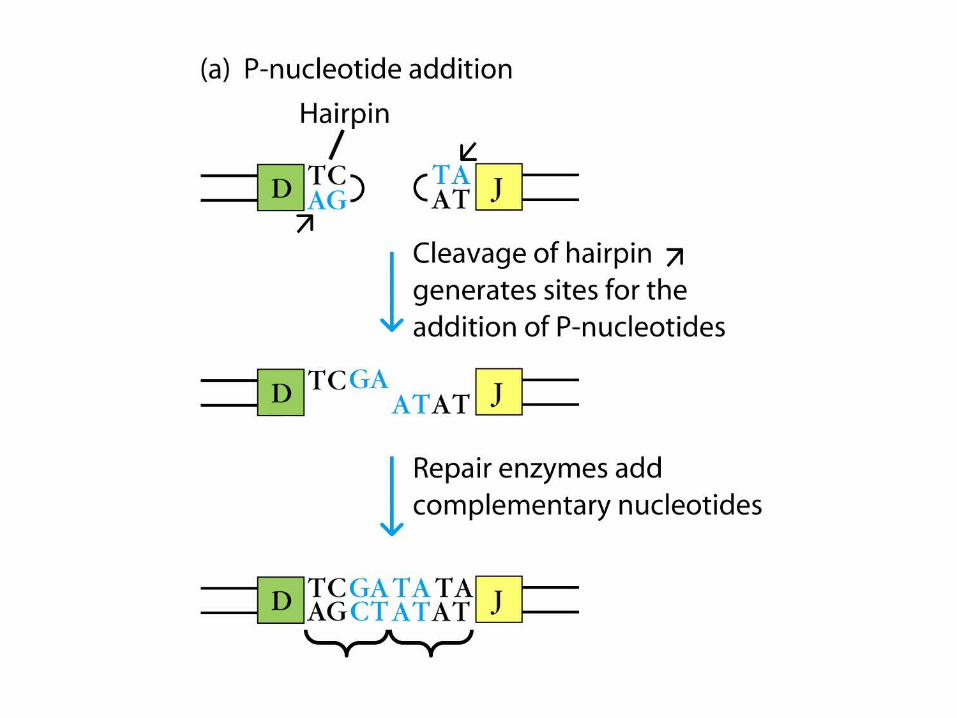
1. Multiple germ-line V, D, and J gene segments 2. Combinatorial V-(D)-J joining 3. Junctional flexibility 4. P-region nucleotide addition (P-addition) 5. N-region nucleotide addition (N-addition) 6. Somatic hypermutation 7. Combinatorial association of light and heavy chains

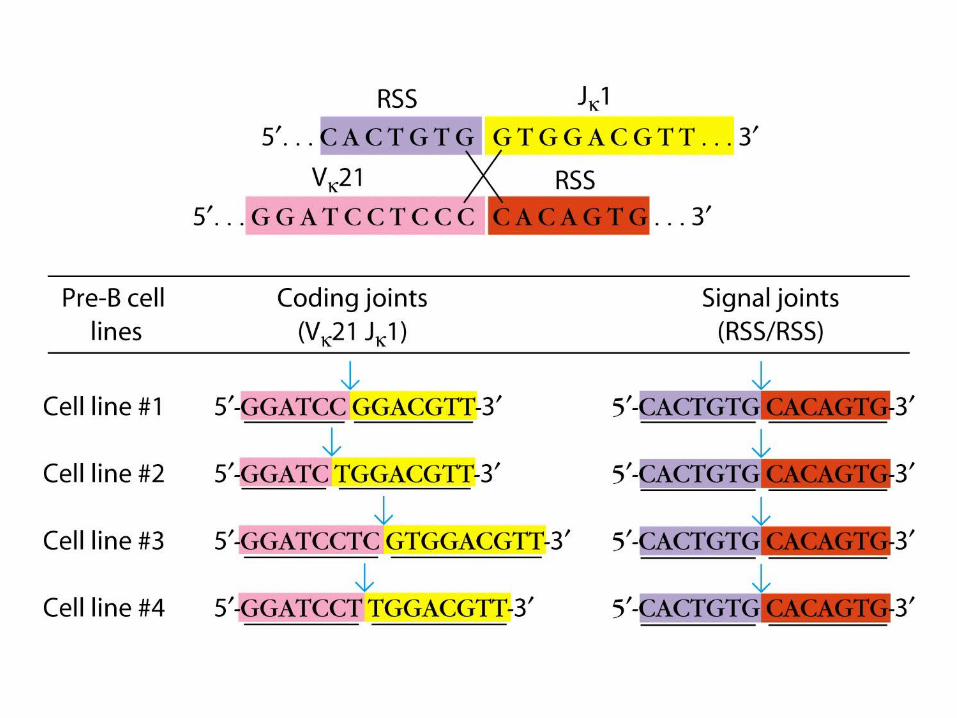
Junctional Flexibility
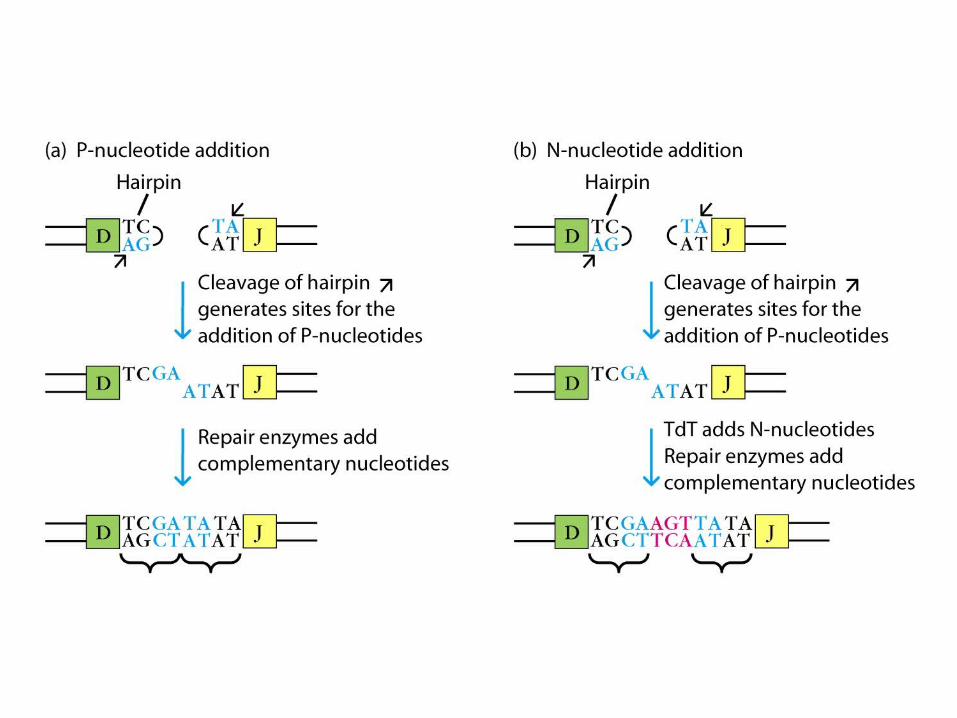

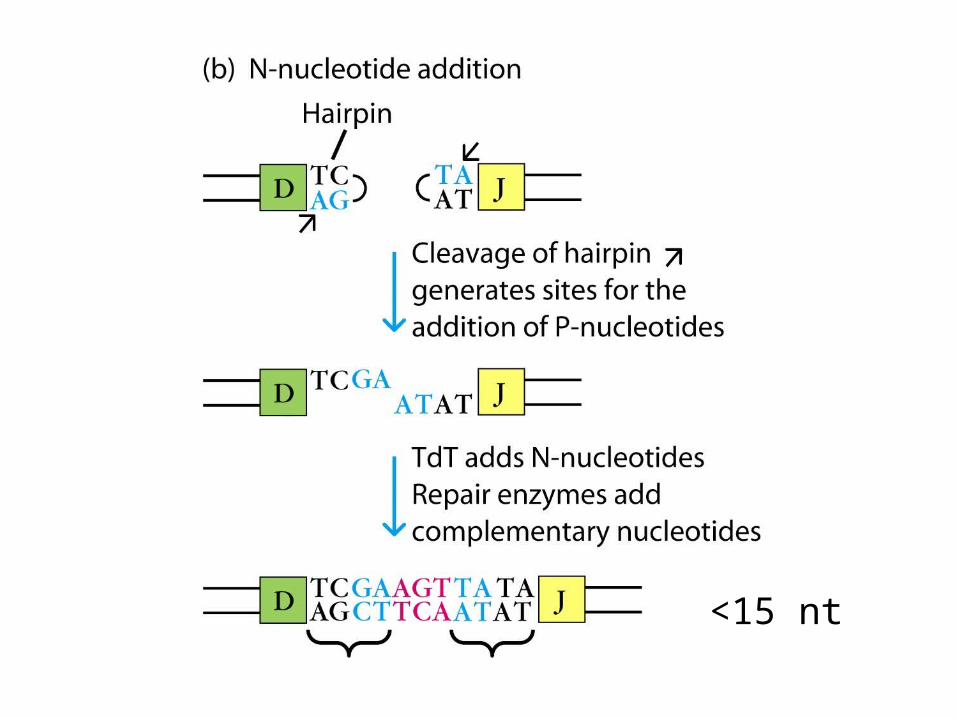
- Up to 15 N-nucleotides can be added to both the DH-JH and VH-DHJH joints.
- Thus, a complete H-chain V region is encoded by a VHNDHNJH unit.
- N regions appears to consist of wholly random sequences



Somatic Hypermutation
- Somatic hypermutation occurs only within germinal germinal centerscenters, structures that form in secondary lymphoid organs within a week or so of immunization with an Ag that activates a T-cell-dependent B-cell response.
- Somatic hypermutation occurs at a frequency approaching 10-3/bp/generation.
- This rate is at least 100,000s-fold higher than the spontaneous mutation rate, about 10-8/bp/peneration, in other genes.
- B cells with higher-affinity Ig receptors will be preferentially selected for survival because of their greater ability to bind to the Ag. ----- Affinity MaturationAffinity Maturation
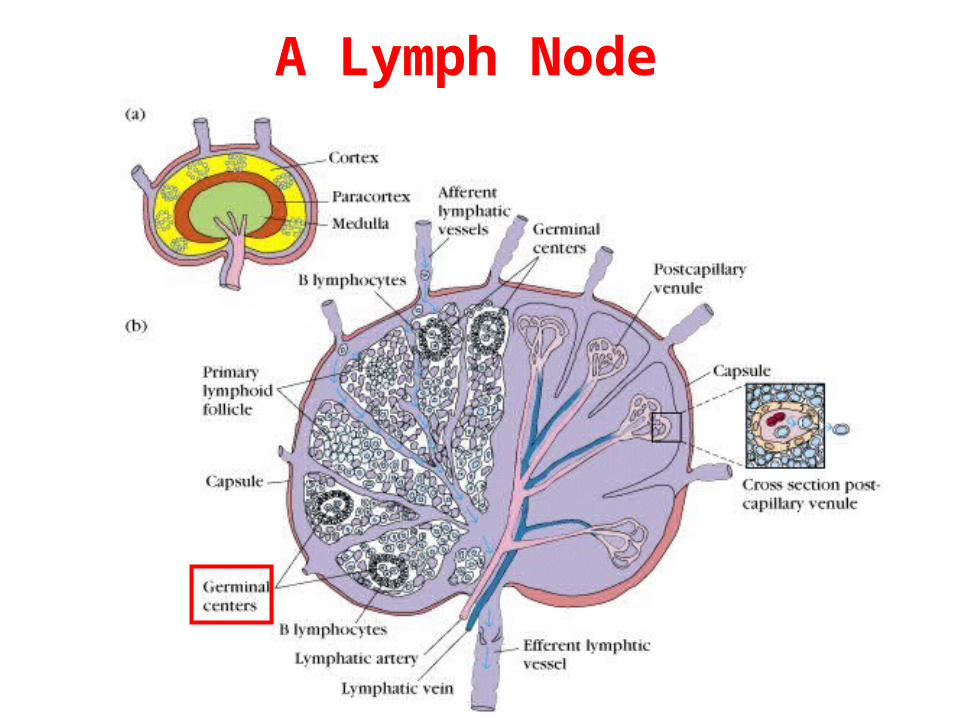
A Lymph Node

The Spleen
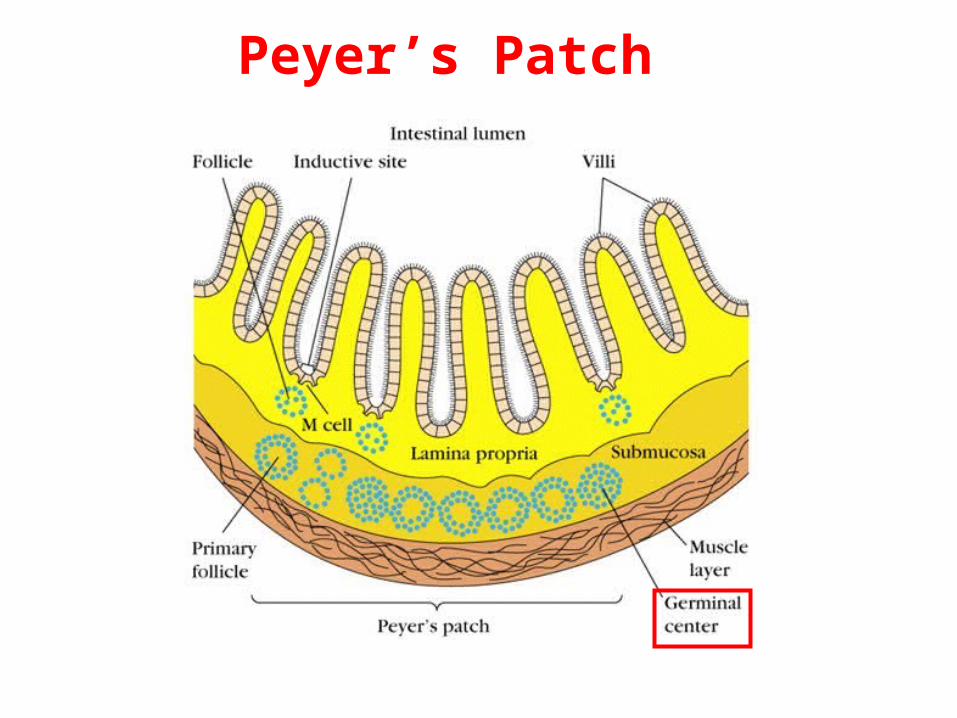
Peyer’s Patch

Antibody Diversity
Seven means of generation of Ab diversity:
1. Multiple germ-line V, D, and J gene segments 2. Combinatorial V-(D)-J joining 3. Junctional flexibility 4. P-region nucleotide addition (P-addition) 5. N-region nucleotide addition (N-addition) 6. Somatic hypermutation – after Ag stimulation 7. Combinatorial association of light and heavy chains

Class Switching Among C-Region Genes

Class (isotype) switching
- Class-specific recombinase proteins may bind to switch regionsswitch regions and facilitate DNA recombination.
- Cytokines secreted by activated TH cells have been shown to induce B cells to class switch to a particular isotype.
- IL-4 induces C to C1 or C (Chapter 11)
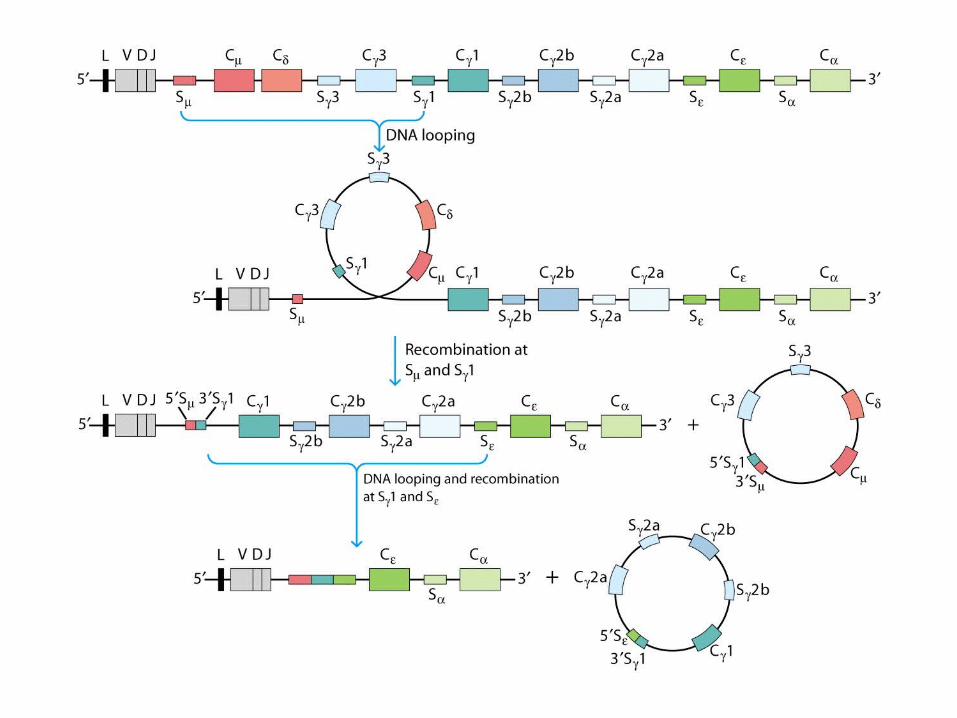
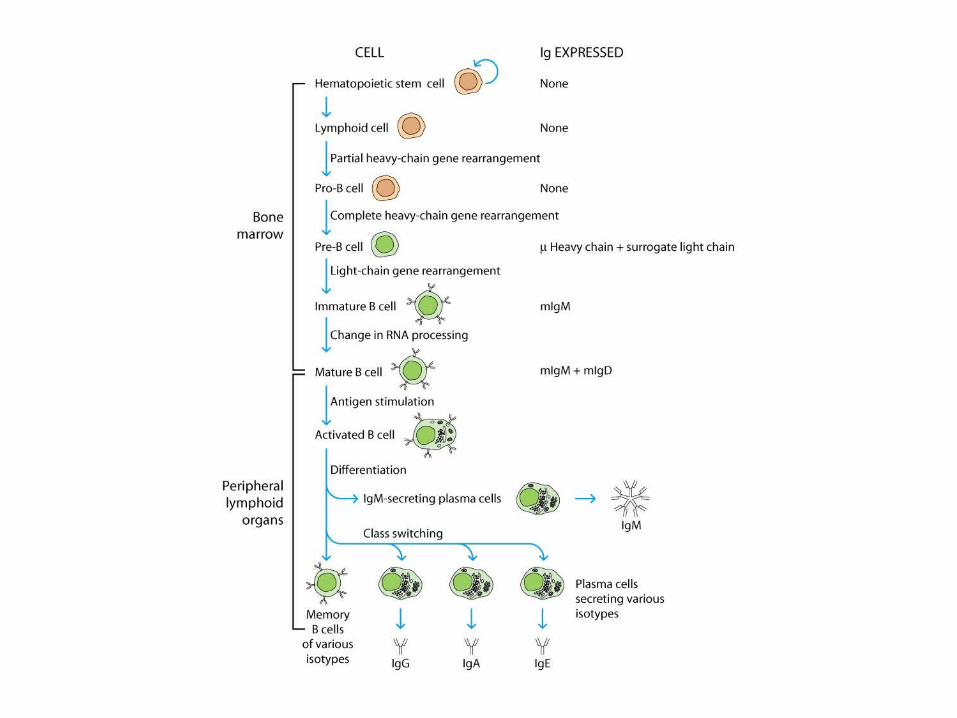

Expression of Ig Genes
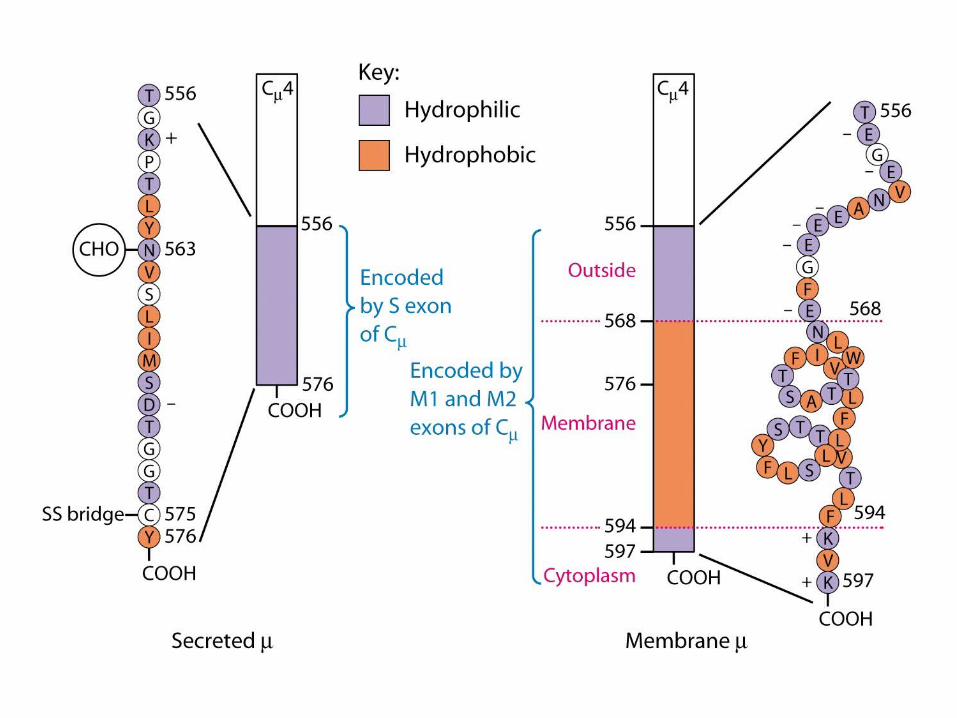
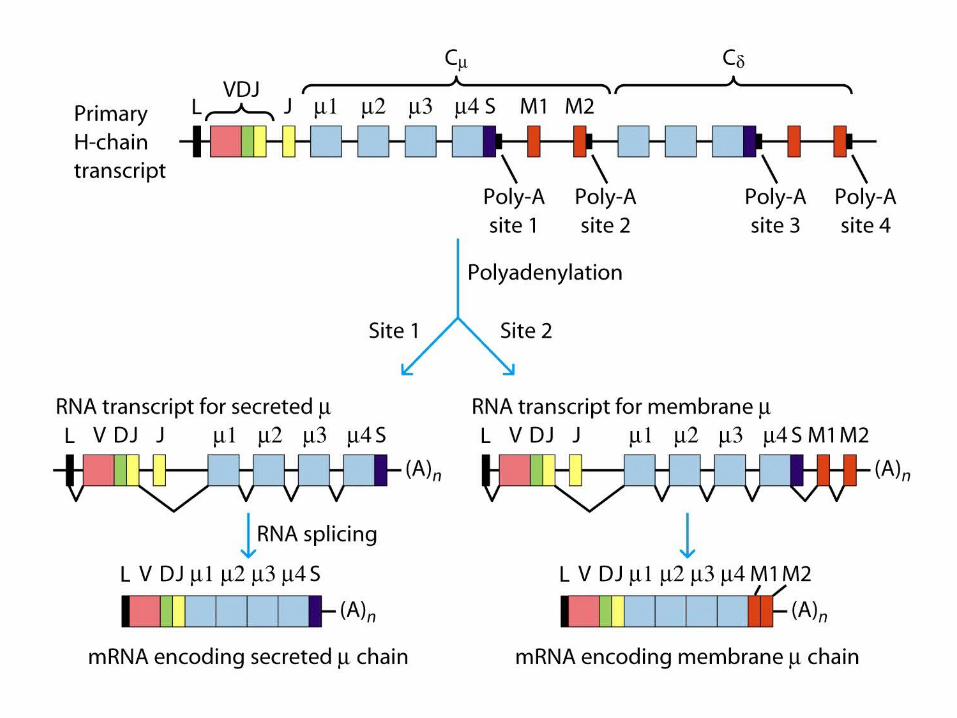
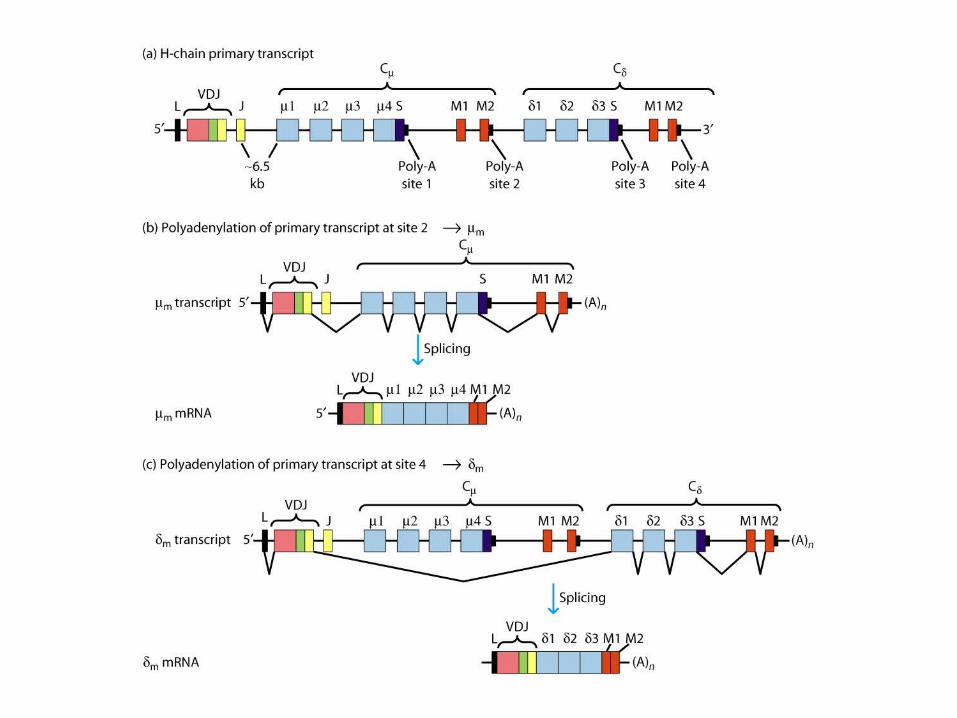
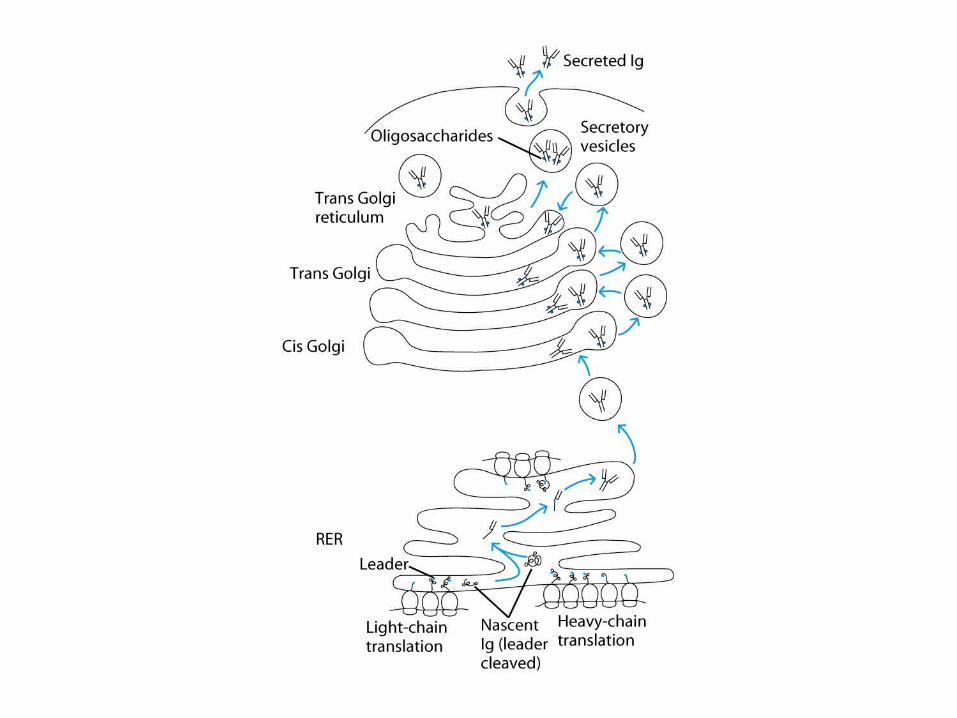
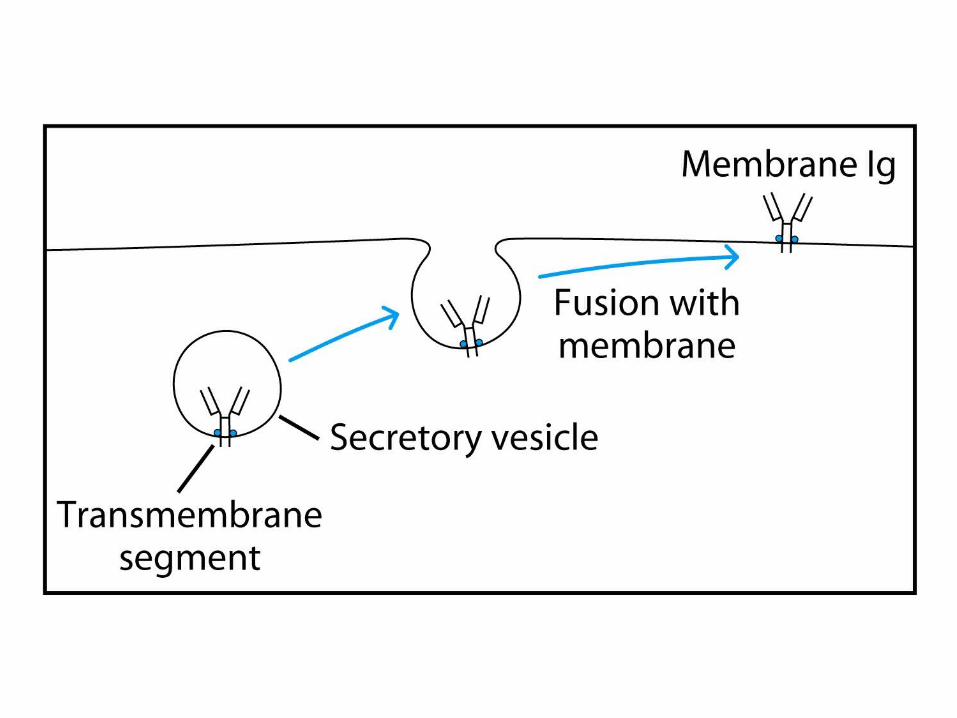

Regulation of Ig-Gene Transcription









![IG and TR single chain fragment variable (scFv) sequence … · 2017. 8. 23. · lected receptor type (for the IG: IGHV, IGKV and IGLV; for the TR: TRAV, TRBV, TRGV and TRDV) [26]](https://img.pdfslide.us/doc/110x75/60ba33a319ea9f32c25a8194/ig-and-tr-single-chain-fragment-variable-scfv-sequence-2017-8-23-lected-receptor.jpg)



















