Embed Size (px)
Citation preview
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
153
Chapter – 4
DISCUSSION

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
154
According to the International Union for Conservation of Nature
and Natural Resources, biodiversity encompasses all life forms,
ecosystems, and ecological processes, and acknowledges the hierarchy at
genetic, taxon, and ecosystem levels (McNeely et al., 1990). Importance of
biodiversity in this era of global climate change is recognized and hence
United Nations declared 2010 as the International year of biodiversity.
India is recognized as one of the 12 mega diversity regions of the world
for its richness in overall species diversity and its contribution to the
global biodiversity is around 8% (Kapur and Jain 2004). Diversity
inventorisation is the first step towards conservation. Present thesis
focuses on inventorisation of a special group of bacteria, Anoxygenic
phototrophic Bacteria (APB) with particular reference to purple bacteria
in the classes Alphaproteobacteria, Betaproteobacteria (purple nonsulfur
bacteria) and Gammaproteobacteria (purple sulfur bacteria) from diverse
habitats of India, including a few habitats and regions hitherto
unexplored by the earlier workers. Apart from satisfying the intellectual
curiosity, studies on cultured diversity can provide raw material for
possible exploitation for human welfare and it is a source of innovation
in biotechnology (Bull et al., 1992).
Most of the new species (and other taxa) of bacteria described from
India in the past ten years belongs to the three phyla, Proteobacteria,
Actinobacteria and Firmicutes. Though new ptoteobacterial species were
extensively described, they were all restricted to chemotrophs. There
were no descriptions of new species of phototrophic Proteobacteria till
2004, though sporadic reports of their distribution in India are available.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
155
Purple non sulfur bacteria are reported from industrial effluents near
Hyderabad (Renuka et al., 1987; Sasikala et al., 1995), paddy soils of
south India (Sasikala et al., 2002), and reverine ecosystems (Sarkar and
Banerjee, 1979), while purple sulfur bacteria were isolated from tropical
waters of Pichavaram mangroves near Port Novo and other coastal
ecosystems (Loka Bharathi and Chandramohan, 1986; Krishnamurthy et
al., 1986). Though description of novel species of Anoxygenic
Phototrophic Bacteria from India started relatively recently (Ramana et
al., 2005), there are 40 species names and 2 genera names validly
described till July 2011 (http://www.bacterio.cict.fr).
Sample collection was random and arbitrary and, it was one time
sampling at a particular site (Fig. 3.1). Diverse habitats of different
regions of sampling are given in the methodology (section, 2.9). Apart
from samples such as soil, sediment, water, microbial mats and natural
purple blooms (Fig. 4.1) which were earlier analyzed for APB diversity
from India (Arunasri, 2005; Srinivas, 2008; Anil Kumar, 2008; Kalyan,
2011), dead marine invertebrate samples (jellyfish with pink, brown and
red coloured spots) (Fig. 4.1d) were also utilized for the isolation of purple
bacteria. Most of the other samples studied were from the Indian
Himalayas (20) and coast of south India (30). During sampling,
pink/purple/brown coloured ponds (Fig. 4.1a and 4.1b) and sands (Fig.
4.1c) with blooms of purple phototrophic bacteria were observed and
sampled. Apart from visual observation, habitats rich in purple bacteria
can be located (even when blooms are not present) by remote sensing
(Gitelson et al., 1999) using bacteriochlorophylls as biomarkers.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
156
For isolation of bacteria from environmental samples, two
strategies can be used: direct isolation and enrichment isolation. Since
normally purple bacteria occur in relatively low numbers in natural
habitats, they require enrichment prior to isolation. However, from
natural blooms, which are rather quite rare, they can be isolated by
direct plating or streaking on agar. Since microbial population of would
have acclimatized to respective environmental conditions of the sample,
Fig. 4.1 Few natural purple blooms and dead marine
specimen (sampling sites). (a) Purple pond near seashore, (b)
Purple bloom in the pond, (c) Purple sand near seashore at
Pamban Bridge and (d) Dead Jelly fish at seashore

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
157
mineral salts such as calcium chloride; sodium chloride and magnesium
sulfate were adjusted according to the nature of the sample
(marine/fresh water) in the enrichment media.
In the present study, different types of bacteria were enriched in
the same mineral media by varying carbon source/e- donor, nitrogen
source, salt concentration, sulfur source and pH. For example, Rba.
maris JA276T (Fig. 3.2) was enriched from moist sediment of seashore at
Cochin in a mineral medium containing 0.04% (w/v) NaCl and pyruvate
as carbon source/e- donor, whereas Mch. gracile sp. JA260 (Table 3.1)
(purple sulfur bacteria) was isolated from the same sample by replacing
pyruvate with bicarbonate as inorganic carbon source and sodium
sulfide as e- donor with 2% (w/v) NaCl. Similarly, strain JA225 of the
genus Rhodobacter could be enriched from microbial mat of hot sulfur
spring of Manikaran (Table 3.1) by using pyruvate as carbon source,
whereas Rps. faecalis JA227 (Table 3.1) was enriched from the same
sample by replacing pyruvate with sodium bicarbonate as inorganic
carbon source and hydrogen as electron donor. Few strains, those
belonging to the genera Rhodothalssium, Blastochloris and
Rhodomicrobium were isolated for the first time from India by using
different physico-chemical parameters for enrichments such as high salt
concentration (5-8% w/v)/glutarate and maltose as carbon
source/succinate as carbon source at low pH (5.5) respectively.
Incubation conditions such as light and temperature were also
varied which could allow enrichment of a wide variety of purple bacteria.
For example, strains JA349 and JA350 with two completely different

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
158
carotenoid series (spirilloxanthin and okenone respectively) of the same
genus Marichromatium were enriched from the pink coloured moist sand
at seashore at Pamban Bridge, by using fluorescent light and
incandescent light respectively (Table 3.1). Hence, it is suggested to use
multiple enrichment media that mainly vary in medium components like
carbon source, nitrogen source, NaCl concentration and pH; and
physical parameters like temperature and light (intensity and quality). In
contrast, maintaining laboratory conditions that are similar to that of the
environment may enable the growth of dominant bacteria of respective
environments.
It is suggested to isolate purple bacterial strains from the
enrichments as early as possible since algal succession was observed in
certain enrichment cultures upon prolonged incubation. However, the
incubation of the enrichment media for prolonged time (if not dominated
by undesired organisms) enables detection of slow growing organisms or
organisms that develop only after modification of the environment by
other strains.
The isolation and purification methods used here are streaking on
agar slants and plates/ agar shake cultures. Since most of the target
isolates from the study are obligate anaerobes, they were isolated by the
method of streaking on agar slants, in which anaerobic condition is
maintained by sealing with butyl rubber stoppers and flushing with inert
gas argon to replace the air in it. Isolation of facultative anaerobes of
purple nonsulfur bacteria can also be done by streaking on agar plates.
However, the same could not be followed for purple sulfur bacteria, as
they require reduced sulfur compounds like sulfide as electron donor in

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
159
their medium, which could be oxidized on exposure to air. One more
method, paraffin wax overlay (Archana et al., 2004) was also suggested
for the isolation of these bacteria which however is applicable only for
those strains which can withstand exposure to melting temperature of
wax (45-50OC).
All the 59 isolated purple bacterial strains (Table 3.2) could not be
subjected to the laborious polyphasic characterization, which includes
large numbers (about 150) of tests. Hence, for detailed polyphasic
characterization, 9 strains (JA194T, JA276T, JA296T, JA248T, JA430T,
JA310T, JA531T, JA349T, JA553T) which showed unique characters
(Table 3.2) were selected by rapid typing based on characteristics such as
habitat, morphological characters, bacteriochlorophylls and 16S RNA
gene sequence similarity (< 97%) (Stackebrandt and Goebel, 1994).
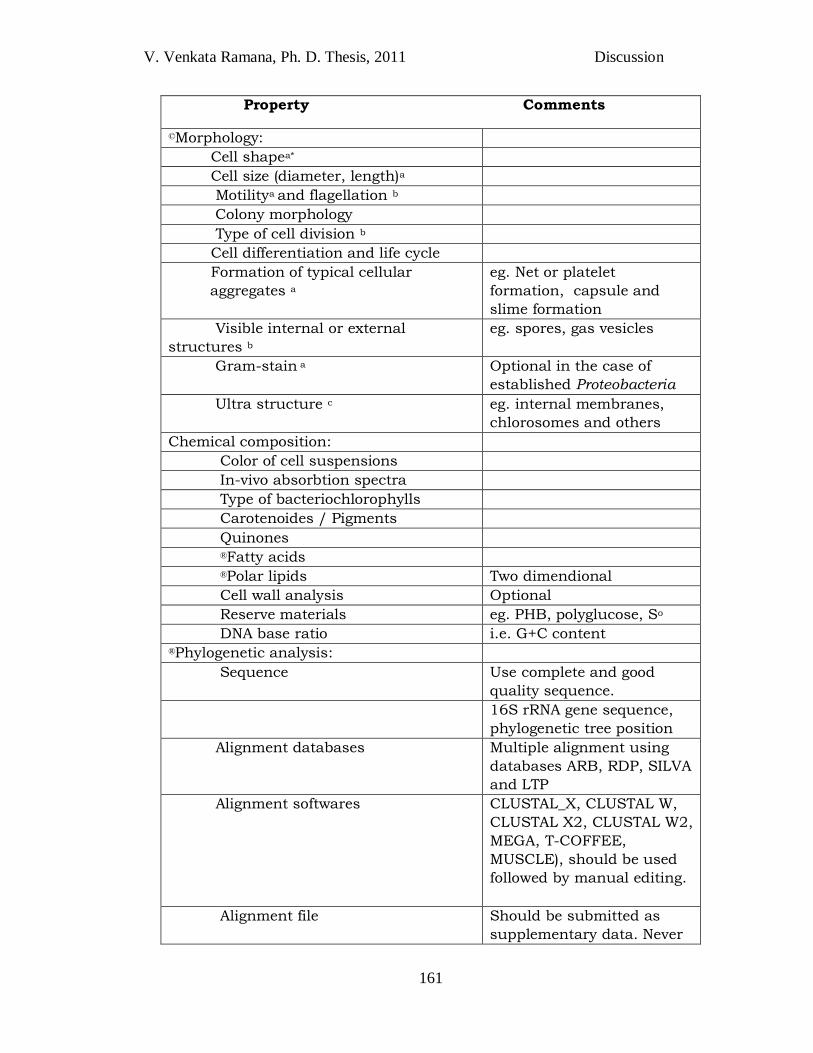
Polyphasic taxonomy includes the study of multiple parameters
with respect to the delineating taxa. As there is no single golden standard
to classify and identify the bacterial taxa, polyphasic characterization is
well accepted approach in the present bacterial taxonomic studies. The
standards and number of methods and/or parameters of polyphasic
taxonomy are not final and are subject to constant changes. Two
publications that were consulted for the description of novel taxa of
anoxygenic phototrophic bacteria are ―Recommended standards for the
description of new species of the anoxygenic phototrophic bacteria‖
(Imhoff and Caumette, 2004) and ―Notes on the characterization of
prokaryote strains for taxonomic purposes‖ (Tindall et al 2010) (Table
4.1).

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
160
Until 2010, purple bacterial taxa description has been carried out
based on recommended standards for polyphasic taxonomic study by
Imhoff and Caumette 2004 (Rdv. marinum [Srinivas et al., 2006];
Rubrivivax bezoatilyticus [Ramana et al., 2006]; Rba. vinaykumarii
[Srinivas et al., 2007]; Rba. maris [Venkata Ramana et al., 2008];
Thiohalocapsa marina [Anil Kummar et al., 2009]) with no need of
optional characters (chemotaxonomy). Since then, chemotaxonomic
characters such as FAME, polar lipids, quinones and genetic
characterization such as phylogenetic analysis based on housekeeping
genes, Multi Locus Sequence Anlaysis (MLSA) (Richter et al., 2006;
Delétoile et al., 2010) became essential (Table 4.1) and, are being used to
delineate the taxa. However, information of chemotaxonomic data are
not available for all type strains and there is an urgent need to create
libraries or databases of cellular fatty acids, quinones and polar lipids to
enable comparative analysis in taxonomic studies which may also lead to
discovery of possible biotechnological potentials. For polyphasic
characterization, the target strain has to be comparatively studied with
all nearest type strains of the respective higher taxa in all aspects at
author‘s laboratory (Tindall et al., 2010). This requirement is becoming a
bottleneck in description of novel taxa, since frequently no type strains
are available and sometimes legitimacy also is questionable (Okamura et
al., 2009).
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
161
Property Comments
©Morphology:
Cell shapea*
Cell size (diameter, length)a
Motilitya and flagellation b
Colony morphology
Type of cell division b
Cell differentiation and life cycle
Formation of typical cellular
aggregates a
eg. Net or platelet
formation, capsule and
slime formation
Visible internal or external
structures b
eg. spores, gas vesicles
Gram-stain a Optional in the case of
established Proteobacteria
Ultra structure c eg. internal membranes,
chlorosomes and others
Chemical composition:
Color of cell suspensions
In-vivo absorbtion spectra
Type of bacteriochlorophylls
Carotenoides / Pigments
Quinones
®Fatty acids
®Polar lipids Two dimendional
Cell wall analysis Optional
Reserve materials eg. PHB, polyglucose, So
DNA base ratio i.e. G+C content ®Phylogenetic analysis:
Sequence Use complete and good
quality sequence.
16S rRNA gene sequence,
phylogenetic tree position
Alignment databases Multiple alignment using
databases ARB, RDP, SILVA
and LTP
Alignment softwares CLUSTAL_X, CLUSTAL W,
CLUSTAL X2, CLUSTAL W2,
MEGA, T-COFFEE,
MUSCLE), should be used
followed by manual editing.
Alignment file Should be submitted as
supplementary data. Never
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
162
truncate an aligned file.
Pair wise similarity calculation To use ARB, PHYDIT and
jPHYDIT programs
Phylogenetic tree To use distance matrix,
maximum-parsimony,
maximum-likelihood
methods.
Genetic analyses:
DNA-DNA hybridization If applicable
®MLSA, MLST If applicable
®Sub species and strain level AFLP-PCR, PFGE, RAPD,
rep-PCR, REP-PCR, ERIC-
PCR, BOX-PCR, (GTG)5 -
PCR and ribotyping.
Physiology:
Growth medium
Temperature range and optimum
pH range and optimum
NaCl range and optimum
Vitamin requirements
Carbon, nitrogen and sulfur
Sources
Relations to sulphide Tolerance, use, oxidation
products
Relation to oxygen Anaerobic, aerobic,
microaerobic growth,
tolerance
Light responses Minimum requirement,
light sensitivity: optional
Modes of energy generation:
Phototrophy,
Potential chemotrophy,
Electron donors,
Electron acceptors
Ecology:
Natural habitat Physico-chemical
properties, pH, salinity,
temperature
Source of isolates Geographical location, GPS
positioning
Mineral salts composition of source
Abundance at habitat
Culture deposition: Minimum of 2 culture
collection certificates
Sequence deposition: To be accessible to public

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
163
EMBL Accession number
Comparitive study
Reference strains
Should be characterized at
author‘s laboratory
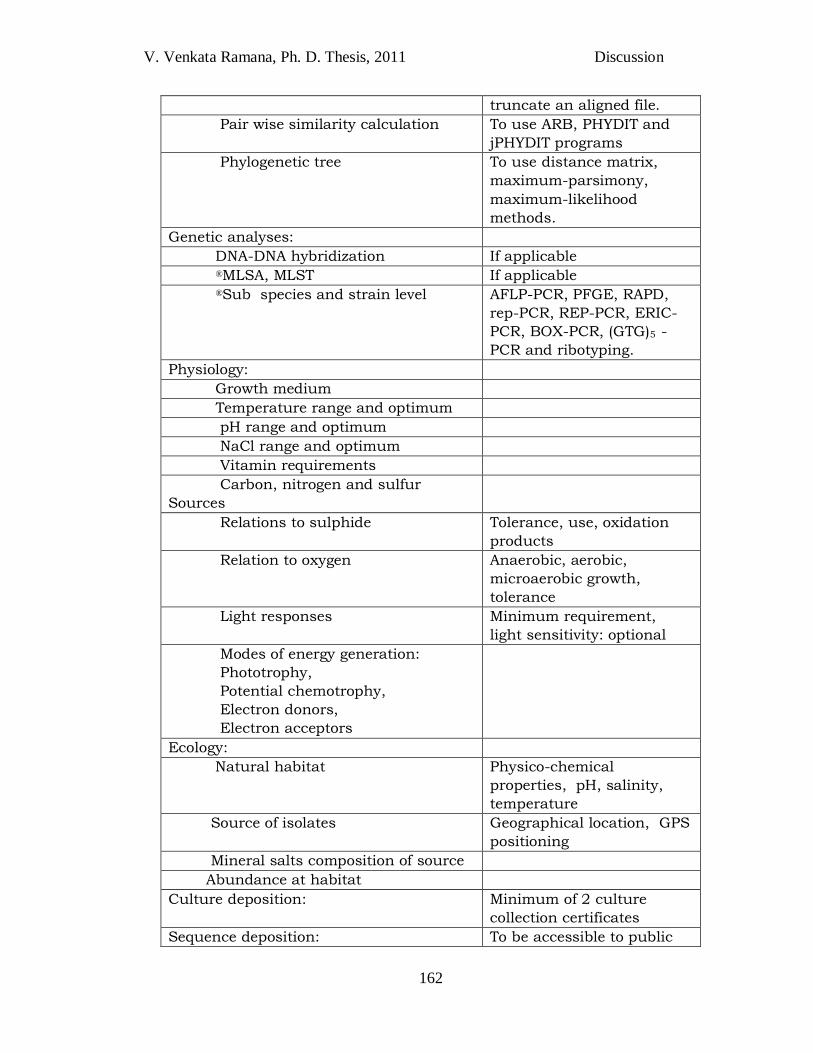
Table 4.1 Revised table of characteristics / parameters required for the description of new species of anoxygenic phototrophic bacteria. (Imhoff and Caumette 2004; modification after recommendations of ®Tindall et al., 2010). Those marked as optional provide possibly important properties of the bacterium but are not regarded as essential for the species description. *To be demonstrated by: a) a light micrograph; b) a light or electron micrograph; c) an electron micrograph.
STRAINS OF THE GENUS Rhodobacter (Rba)
The family Rhodobacteraceae of Alphaproteobacteria consists of 5
phototrophic genera; Rhodobacter, Rhodobaca, Rhodovulum,
Rubribacterium and Rhodothalassium. Rhodobacter and Rhodovulum are
the two genera each having relatively large number of species (12 each)
among APB. Rhodobacter is the type genus of the family
Rhodobacteraceae, order Rhodobacteriales, class Alphaproteobacteria.
(http://www.bacterio.cict.fr/classifgenerafamilies.html#Rhodobacteracea
e).
The genus Rhodobacter is differentiated from other genera with few
characteristic properties such as ovoid to rod shaped cell morphology,
the presence of vesicular internal membranes (Except in Rba. balsticus)
and carotenoides of spheroidene series (Imhoff, 2005). Although
Rhodobacter spp. and Rhodovulum spp. rather differ in NaCl requirement
and they share common chemotaxonomic characteristics such as large
type cytochrome C2 (Amber et al., 1979), sole quinone component Q-10
(Imhoff, 1984a), lipopolysaccharides with gulucosamine as sole amino
sugar in their lipid A moieties, phosphate and amide-linked 3-OH-14:0

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
164
and/or 3-oxo-14:0 and ester linked 3-OH-10:0 (Weckesser et al., 1995),
those enable their distinction from other genera. Rhodobacter spp are
differentiated from each other based on morphology, slime production,
sulfate assimilation, denitrification, vitamin requirement, utilization of
carbon source and fatty acid composition, 16S rRNA gene sequence
comparison and DNA-DNA hybridization (de Bont et al., 1981; Ivanova et
al., 1988; Hiraishi et al., 1996).
In enrichment cultures set up for purple nonsulfur bacteria,
members of this genus grown faster and outcompete other purple
nonsulfur bacteria. In the present study, 11 different strains of the genus
Rhodobacter; JA194T, JA247, JA260, JA276T, JA296T, JA312, JA313,
JA431, JA533, JA542 and JA555 were isolated from diverse habitats
such as soil, estuarine water, fresh water pond, water and sediment of
seashore, microbial mat, soil below the ice (high attitude) and hot sulfur
springs, whereas the existing type strains are only from fresh water,
sewage ponds and eutrophic lakes
(http://www.bacterio.cict.fr/qr/rhodobacter.html), which reflects the
identification of novel habitats of Rhodobacter strains.
Among the above 11 strains, 3 strains JA194T, JA276T,
JA296T exhibited unique characters in the rapid typing and were
subjected to detailed polyphasic study. Strains isolated from similar
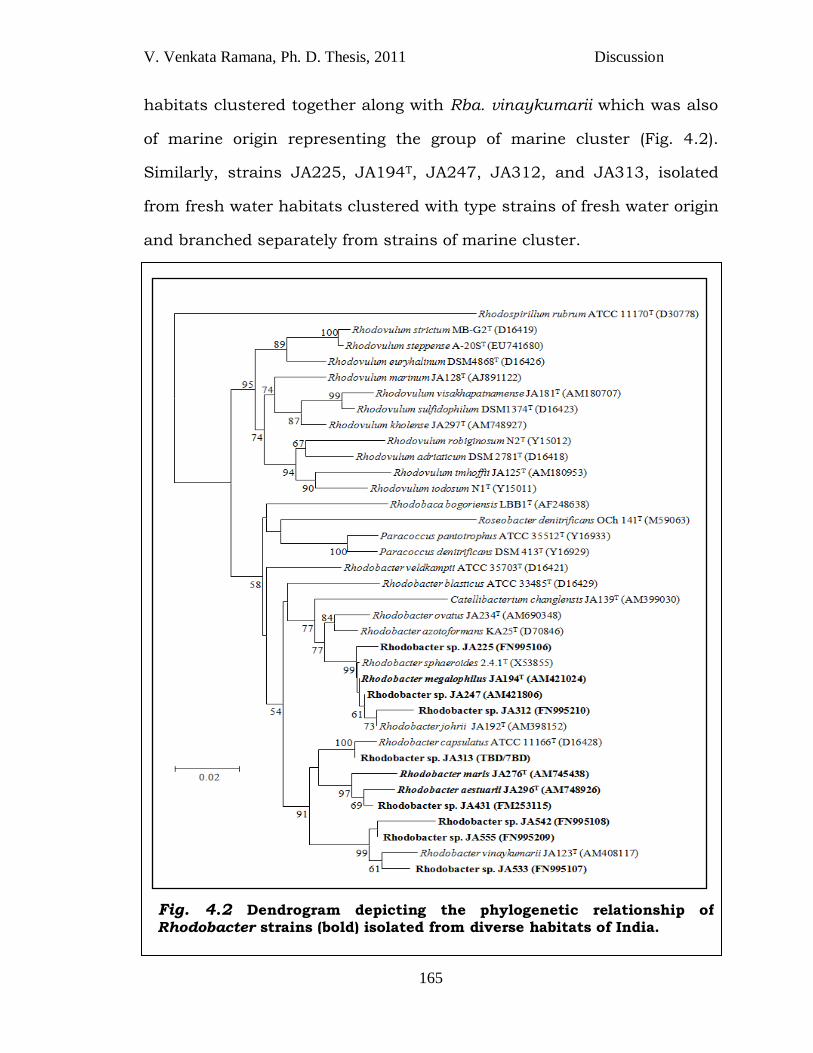
ecological niche showed close phylogenetic relationship which was
observed by the phylogenetic tree of Rhodobacter spp. from the present
study (Fig. 4.2) In the phylogenetic tree, strains JA276T, JA296T, JA431T,
JA533, JA542 and JA555 which were isolated from marine and estuarine
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
165
habitats clustered together along with Rba. vinaykumarii which was also
of marine origin representing the group of marine cluster (Fig. 4.2).
Similarly, strains JA225, JA194T, JA247, JA312, and JA313, isolated
from fresh water habitats clustered with type strains of fresh water origin
and branched separately from strains of marine cluster.
Fig. 4.2 Dendrogram depicting the phylogenetic relationship of Rhodobacter strains (bold) isolated from diverse habitats of India.
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
166
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
167

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
168
Strains JA194T and JA247
Strain JA194T is completely oval, similar to Rba. ovatus, whereas
remaining all strains are oval to rods except Rba. sphaeroides which is
spherical to rods. The strain JA194T able to grow from 5-400C (Fig. 3.9),
Strain JA194T lacked an ester peak (1735 cm-1) and differed in other
cellular components, including proteins (C–N stretch amines 1230 cm-1),
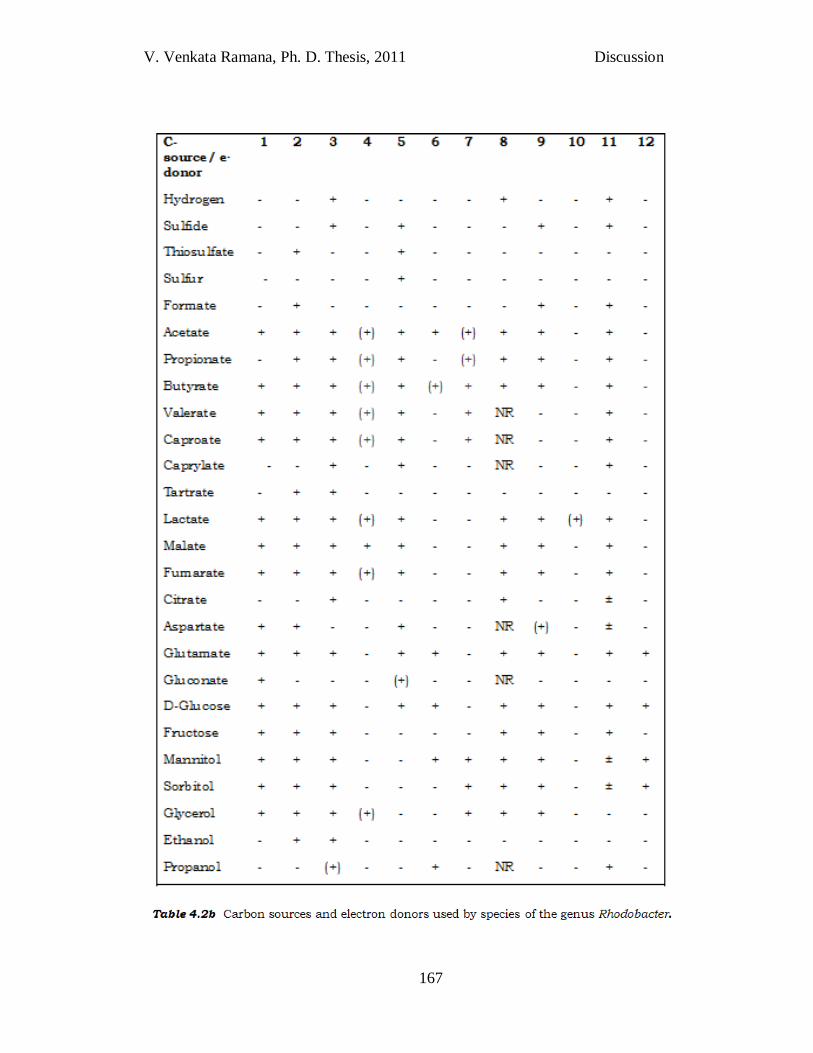
Table 4.2b. Carbon sources and electron donors used by species of
the genus Rhodobacter. Taxa: 1, Rba. johrii JA192T (Girija et al., 2010); 2, JA194T ; 3, Rba. sphaeroides (Imhoff, 2005); 4, JA276T; 5, Rba. veldkampii (Hansen & Imhoff, 1985; Imhoff, 2005); 6, Rba. vinaykumarii (Srinivas et al., 2007); 7, Rba. ovatus (Srinivas et al., 2008); 8, Rba. blasticus (Imhoff, 2005); 9, Rba. azotoformans (Hiraishi et al., 1996); 10, JA296T; 11, Rba. capsulatus (Imhoff, 2005); 12, Rba. changlensis (Anil Kumar et al., 2007a). For all taxa, organic substrate utilization was tested during photo-organoheterotrophic growth. All taxa utilized pyruvate, but not benzoate or arginine. +, substrate utilized; _, substrate not utilized or absent; ±, variable in different strains; (+), weak growth; NR, not reported.
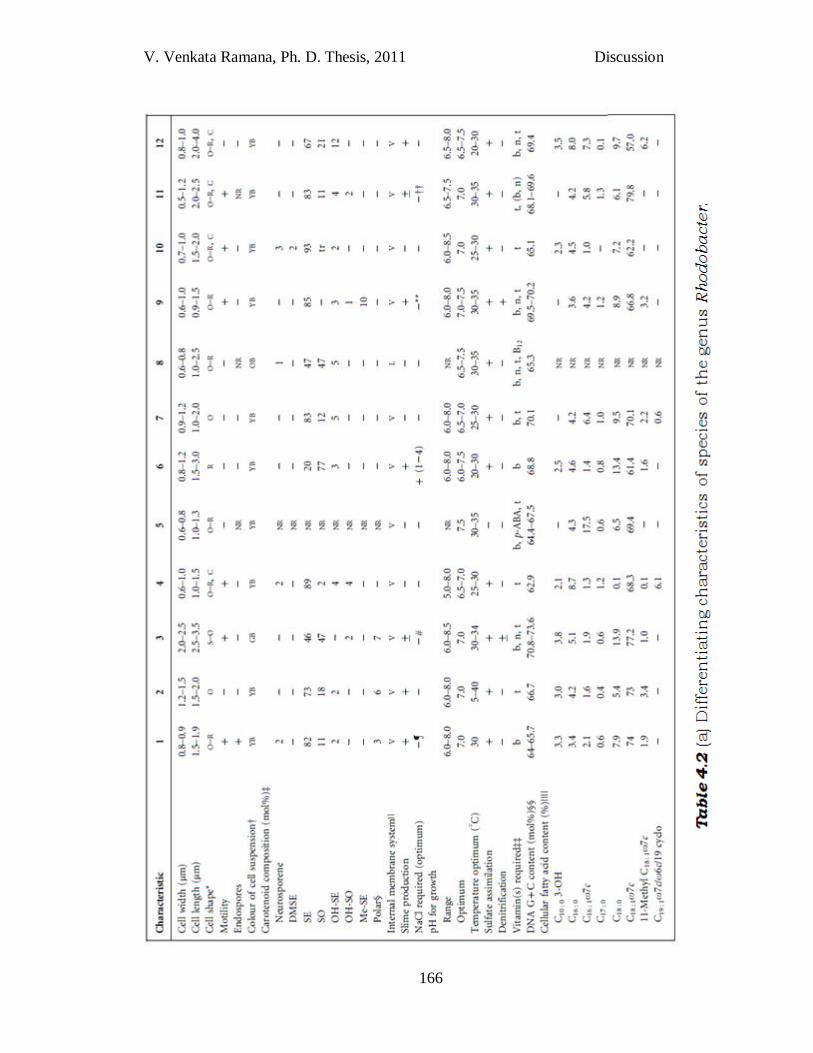
Table 4.2 (a) Differentiating characteristics of species of the genus
Rhodobacter. Taxa: 1, Rba. johrii JA192T; 2, JA194T; 3, Rba. sphaeroides DSM 158T; 4, JA276T; 5, Rba. veldkampii; 6, Rba. vinaykumarii JA123T; 7, Rba. ovatus JA234T; 8, Rba. blasticus; 9, Rba. azotoformans KA25T; 10, JA296T; 11, Rba. capsulatus; 12, Rba. changlensis JA139T. Data represent a comparative analysis done in the authors‘ laboratory unless indicated. Motile species exhibit polar flagella. +, Present; 2, absent; ±, variable in different strains; NR, not reported; *C, Chains; O, ovoid; R, rod-shaped; S, spherical; †GB, Greenish brown; OB, orange–brown; YB, yellowish brown. ‡DMSE, Demethylspheroidene; SE, spheroidene; SO, spheroidenone; OH-SE, hydroxyspheroidene; OH-SO, hydroxyspheroidenone; Me-SE, methoxyneurosporene; Polar, polar carotenoid; tr, trace; §Polar carotenoid with nine conjugated double bonds; ||L, Lamellar; V, vesicular; ¶NaCl not required, but optimal growth occurs at 3% NaCl; #Optimal growth in the absence of NaCl but growth occurs at 3% NaCl; **Optimal growth in the absence of NaCl but growth occurs at 5% NaCl; ††Data from Hiraishi et al. (1996); ‡‡b, Biotin; B12, vitamin B12; n, niacin; p-ABA, p-aminobenzoic acid; t, thiamine; (b, n), a few strains require biotin and/or niacin; §Ranges represent values for more
than one strain, including the type strain; ||||-, Not detected/not reported.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
169
polysaccharides (1183, 1131 and 1100 cm-1) and aromatic compounds
(980 cm-1), compared with Rba. sphaeroides DSM 158T. A peak in strain
JA194T at 3150 cm-1 in the lipid FT-IR fingerprint ( Fig. 3.10) was due to
N–H stretching of peptide linkages of the polypeptide chain of
lipoproteins (Lam et al., 2004) which is not observed in the Rba.
sphaeroides DSM 158T. Furthermore, the high intensities of peaks
observed in strain JA194T indicate the presence of longer acyl chains of
fatty acids (Ami et al., 2006).
Strain JA194T vary from speices of the genus Rhodobacter in
utilizing organic carbon sources/e- donors (Table 4.2b), for their
phototrophic growth. Though the strain JA194T has distinct phenotypic,
physiological and chemotaxonomic properties, its 16S RNA gene
sequence is 99% similar to the nearest type strain Rba. sphaeroides.
Furthur, genomic relatedness (67% DNA-DNA homology) and metabolite
fingerprinting of strain JA194T supported its distant phylogenetic
relatedness with the nearest type strain Rba. sphaeroides DSM 158T and
necessitated the description of strain JA194T as a novel species of the
genus Rhodobacter named as Rhodobacter megalophilus JA194T. Strain
JA247 has all characteristics similar to strain JA194T and proposed as
an additional strain of Rhodobacter megalophilus JA194T.
Description of Rhodobacter megalophilus JA194T sp. nov.
Rhodobacter megalophilus [me.ga.lo.phi‘lus. Gr. adj. megas wide; Gr. adj.
philos loving; N.L. masc. adj. megalophilus wide (temperature)-loving].
Cells are Gram-negative and oval-shaped (1.2–1.5 X 1.5– 2.0 mm).
Multiplication occurs by binary fission. Cells have vesicular

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
170
intracytoplasmic membranes and lack flagella. Phototaxis occurs. Whole-
cell absorption maxima are found at 374, 407, 446, 476, 509, 590, 800
and 854 nm. Growth occurs photo-organoheterotrophically with various
sources, including malate, fumarate, acetate, propionate, butyrate,
valerate, caproate, glutamate, aspartate, glucose, tartrate, ethanol,
mannitol, sorbitol and glycerol. Photolithoautotrophic growth is possible
in the presence of thiosulfate as electron donor and NaHCO3 as carbon
source. Nitrogen sources providing good growth are ammonium chloride
and glutamate. Nitrate and molecular nitrogen also support growth as
nitrogen sources. Thiamine is required for growth. Growth occurs from 5
to 400C. The DNA G+C content of the type strain is 66.67 mol% (by
HPLC). The type strain, JA194T (=KCTC 5602T =JCM 14598T), and an
additional strain of the species, JA247, were isolated from soils of the
Indian Himalayas.
Strain JA276T
Chain formation is observed in strain JA276T like in JA296T, Rba.
capsulatus (Molisch, 1907; Imhoff, 2005) and Rba. changlesis (Anil
Kumar et al., 2007a). Unlike, Rba. vinaykumarii (Sriniva et al., 2007)
isolated from marine water, which had obligate requirement for NaCl,
strain JA276T do not require NaCl though isolated from similar habitat,
but could tolerate upto 3% NaCl (w/v). In the genus Rhodobacter, the
fatty acids C19:1ω7c / ω 6c / 19 cyclo were observed only in strain
JA276T (Table 4.2a). Strain JA276T also varies in utilizing organic carbon
sources/e- donors (Table 4.2b) during phototrophic growth and its 16S
rRNA gene sequence is 96.2% similar to Rba. capsulatus. Morphological,

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
171
physiological and genomic (16S rRNA gene sequence) differences of strain
JA276T (Table 4.2a and 4.2b) from other species of the genus
Rhodobacter, enable strain JA276T to be described as novel species
named Rhodobacter maris JA276T.
Description of Rhodobacter maris JA276T sp. nov.
Rhodobacter maris (ma'ris. L. gen. n. maris, of the sea, pertaining to the
habitat from where the type strain was isolated).
Cells are ovoid to rods, 0.6-1.0 m wide and 1.0-1.5 m long, non-
motile, divide by binary fission and can form chains of 4-6 cells. Growth
occurs under anaerobic conditions in the light (photoorganoheterotrophy)
or under aerobic conditions in the dark (chemoorganoheterotrophy).
Internal photosynthetic membranes are of the vesicular type. The color of
phototrophic cultures is yellowish brown. The in vivo absorption
spectrum of intact cells in sucrose exhibits maxima at 377, 476, 509,
590, 803, and 860 nm confirming the presence of bacteriochlorophyll a.
Carotenoids include spheroidene and spheroidenone. Substrates that
were utilized by strain JA276T as carbon sources and electron donors
under photoorganoheterotrophic conditions include; acetate, fumarate,
pyruvate, malate, glycerol, valerate, lactate, caproate, propionate and
butyrate. Strain JA276T could not utilize formate, caprylate, gluconate,
succinate, thiosulfate, aspartate, ascorbate, benzoate, glutamate, sulfur,
proponol, glucose, fructose, mannitol, peptone, sucrose, casaminoacids,
sorbitol, ethanol, tartrate, sulfite, citrate, oxaloacetate, 2-ketoglutarate,
lactose, maltose, starch, sulfide, bicarbonate, pelarganate, arginine,
yeast extract and oleic acid. Ammonium chloride, glutamate, glutamine

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
172
and molecular nitrogen were utilized as nitrogen sources, while urea,
aspartate, nitrate and nitrite did not support growth. Magnesium sulfate,
thiosulfate, sulfite, thioglycolate and sulfide were utilized as sulfur
sources, while elemental sulfur, methionine and cysteine did not support
growth. NaCl was not obligatory for growth of strain JA276T; but
tolerates up to 3% (w/v) NaCl. Strain JA276T grew at pH 6.0–8.0
(optimum at 6.5-7.0) and at 25–35C (optimum at 30C). Thiamine is
required as growth factor. Autotrophic and fermentative growth is not
possible. The type strain, JA276T (=JCM 14794T =ATCC BAA 1549T
=CCUG 55129T) was isolated from a sediment samples collected from sea
shore of Cochin, Kerala State, India. G+C content of the type strain is
62.85 mol% (by HPLC).
Strain JA296T
The sample yielding strain JA296T was a coloured microbial mat of
brakish water, which is different from the samples that yielded other
strains of the genus Rhodobacter. Strain JA296T also has unique
characters among Rhodobacter spp such as chain formation, marine
origin, lack of NaCl requirement (with tolerance up to 2% NaCl w/v) and
different organic substrate utilization. Apart from the above, strain
JA296T is 95.5% similar to Rba. capsulatus by 16S rRNA gene sequence.
Carotenoid demethylspheroidene (DMSE) was observed only in the strain
JA296T. Therefore, based on morphological, physiological,
chemotaxonomical (Table 4.2a and 4.2b) and genetic differences, strain
JA296T was described as novel species named Rhodobacter aestuarii.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
173
Description of Rhodobacter aestuarii JA296T sp. nov.
Rhodobacter aestuarii (aes.tu.a‘ rii. L. gen. n. aestuarii of the estuary, the
habitat from which the type strain was isolated).
Cells are ovoid to rod-shaped, 0.7-1.0 m wide and 1.5-2.0 m
long, motile, divide by binary fission and form chains of 4-8 cells. Growth
occurs under anaerobic conditions in the light (photo-
organoheterotrophy) or under aerobic conditions in the dark (chemo-
organoheterotrophy). Internal photosynthetic membranes are of the
vesicular type. The color of phototrophic cultures is yellowish green.
Bacteriochlorophyll a and the carotenoids, sphaeroidene and
spheroidenone are present. Substrates that are utilized by strain JA296T
as carbon sources and electron donors under photoorganoheterotrophic
conditions include; pyruvate and lactate. NaCl was not obligatory for the
growth of strain JA296T; but tolerates up to 2% (w/v) NaCl. Strain
JA296T grew at pH 6.0–8.5 (optimum at 7.0) and at 25–35 C (optimum
at 30 C). Thiamine is required as growth factor. Photolithoautotrophic,
chemolithoautotrophic and fermentative growth could not be
demonstrated. The DNA G + C content of the type strain is 65.1 mol% (by
HPLC).
The type strain, JA296T (=ATCC BAA-1550T = CCUG 55130T) was
isolated from a brown colored microbial mat of brackish water from the
Bhitarkanika mangrove forest at Dangmal, Orissa, India.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
174
Objectionable reclassification of Rhodobacter changlensis to the
new genus Catellibacterium
Rba. changlensis (Anil Kumar et al., 2007a) is now reclassified into the
new genus Catellibacterium (Zheng et al., 2011), based on phylogenetic
dendrograms constructed using 16S rRNA gene sequences. Since,
taxonomy of phototrophic bacteria is interspersed with chemotrophs, it
might be possible in clustering Rba. changlensis with the members of the
genus Catellibacterium. However, phototrophy is a genus specific
character of Rhodobacter, Rba. changlesis should not be reclassified into
a genus Catellibacteriuim (non phototrophic members) as none of the
species of the genus Catellibacterium have phototrophy. Therefore,
Rhodobacter changlensis to be further studied carefully to delineate as
novel phototrophic genera.
STRAINS OF THE GENUS Rhodopseudomonas (Rps)
Rhodopseudomonas and Rhodoblastus are the only phototrophic
genera of the family Bradyrhizobiaceae of order Rhizobiales. Genus
Rhodopseudomonas differentiated from other genera of the family
principally based on 16S rRNA gene sequence and phototrophy. Ever
since the description of the genus Rhodopseudomonas (Czurda &
Maresch 1937), the members of this genus were subject to significant
changes including reclassification of species (Imhoff et al., 1984b;
Hiraishi & Ueda, 1994; Hiraishi, 1997) and currently, there are only
three species names remaining with this
genus(http://www.bacterio.cict.fr/qr/Rhodopseudomonas.html);Rhodpse

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
175
udomonas palustris, Rps. rhenobacensis and Rps. faecalis. Since, all the
three type strains are differentiated among themselves in the species
level based on DNA-DNA hybridization, they are called as
Genomospecies.
Species of the genus Rhodopseudomonas belong to the class
Alphaproteobacteria and share common features such as Gram-negative
rod-shaped cells, motile cells by means of polar or subpolar flagella,
polar growth, budding and asymmetric cell division. Primary
characteristic feature is the formation of rosette like aggregates.
Photosynthetic pigments are bacteriochlorophyll a and carotenoides of
the spirilloxanthin series. Preferred mode of growth is photoheterotrophic
under anoxic conditions in the light.
During the study, 11 strains of genus Rhodopseudomonas JA226,
JA227, JA228, JA229, JA251, JA253, JA310T, JA311, JA531T, JA640
and JA641 were isolated from diverse habitats (Table 3.1) of different
geographical regions of India. All the above strains share genus specific
characteristics of Rhodopseudomonas such as Gram-negative rod-shaped
cells, cells are motile by means of polar or subpolar flagella, budding,
asymmetric cell division, formation of rosette like aggregates of the genus
Rhodopseudomonas.
Based on 16S rRNA gene sequence similarity, 9 among 11 isolated
strains belongs to Rps. faecalis (Table 3.2) probably indicating their
predominance. The percentage of relatedness between strains obtained
by 16S rRNA gene sequence alignment tool did not concur with that of
phylogenetic tree. This may probably be because all three type strains

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
176
being Genomospecies, not much variation in the 16S rRNA gene
sequences was observed among them. Though most of the strains JA226,
JA228, JA229, JA227, JA251, JA640, JA641, JA310T and JA311 showed
highest 16S rRNA gene sequence similarity with Rps.faecalis, they have
clustered together and formed a single clade with Rps. palustris. Strains
JA253 and JA531T have separately claded from the node of two clusters
of Rps. faecalis and Rps. rhenobacensis (Fig. 4.3).
Strains yielding samples include fresh water of lake and stagnant
water pool, sediment of saltpan, sewage water, soil, microbial mats and
hot sulfur spring waters, whereas type strains of the genus
Rhodopseudomonas were isolated from soil/sludge/pond water (Molisch
1907), sediment of fresh water lake (Hougardy et al., 2000) and chicken
feces (Zhang et al., 2002).
Like strains of the genus Rhodobacter (Table 3.1), strains of genus
Rhodopseudomonas are also frequently isolated from most of the
samples. A mineral medium prescribed by Biebl and Pfennig 1981 for the
isolation of PNSB or the members of Rhodospirillaceae, can be applied for
isolation and cultivation of Rhodopseudomonas species (Imhoff, 1998a).
In the present study, strains of the genus Rhodopseudomonas were
widely isolated from the enrichment media (Table 2.1) formulated with
pyruvate as a carbon source with different NaCl concentrations. Based
on rapid typing, 2 strains JA310T and JA531T were selected for detailed
polyphasic characterization.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
177
Fig. 4.3 Dendrogram depicting the phylogenetic relationships strains
JA531T, JA253, DSM 123T, JA251, JA229, JA640, JA641, JA311, JA226, JA228, JA310T and JA227 with the members of the genus Rhodopseudomonas along with interspersing chemotrophs as determined using 16S rRNA gene sequence. Tree was constructed by the neibour joining method using the MEGA4 software. Bar; two nucleotide substitutions per 100 nucleotides.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
Results
178
Strain JA531T
Strain JA531T is the only strain of Rhodopseudomonas so far
isolated from the hyper saline habitat (saltpan 10% NaCl w/v). However,
this strain has no obligate requirement for NaCl but can tolerate up to
4% NaCl. Strain JA531T could grow under a wide range of pH (5-9) (Table
4.3), which was not observed in any of the other species of the genus
Rhodopseudomonas. Utilization of benzoate (up to 30 mM) was observed
by strain JA531T, which is a characteristic feature of Rps. palustris.
These unique characters made us to go for polyphasic characterization to
ascertain if it represents a new species. Strain JA531T has interspecies
variability through their habitat, cell size, type of budding, colour of the
culture, temperature, pH range, growth factor requirement, polar lipid
composition, fatty acid composition, carbon source utilization, DNA-DNA
relatedness and G+C content (Table 4.3). Since 16S rRNA gene sequence
similarity of strain JA531T is 97-98% (i.e, >97%) with Rps. palustris, Rps.
faecalis and Rps. rhenobacensis, DNA-DNA relatedness studies were
carried out. A genome relatedness of 48-62% observed further confirmed
that strain JA531T represents a novel species which was named
Rhodopseudomonas harwoodiae JA531T.
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
179
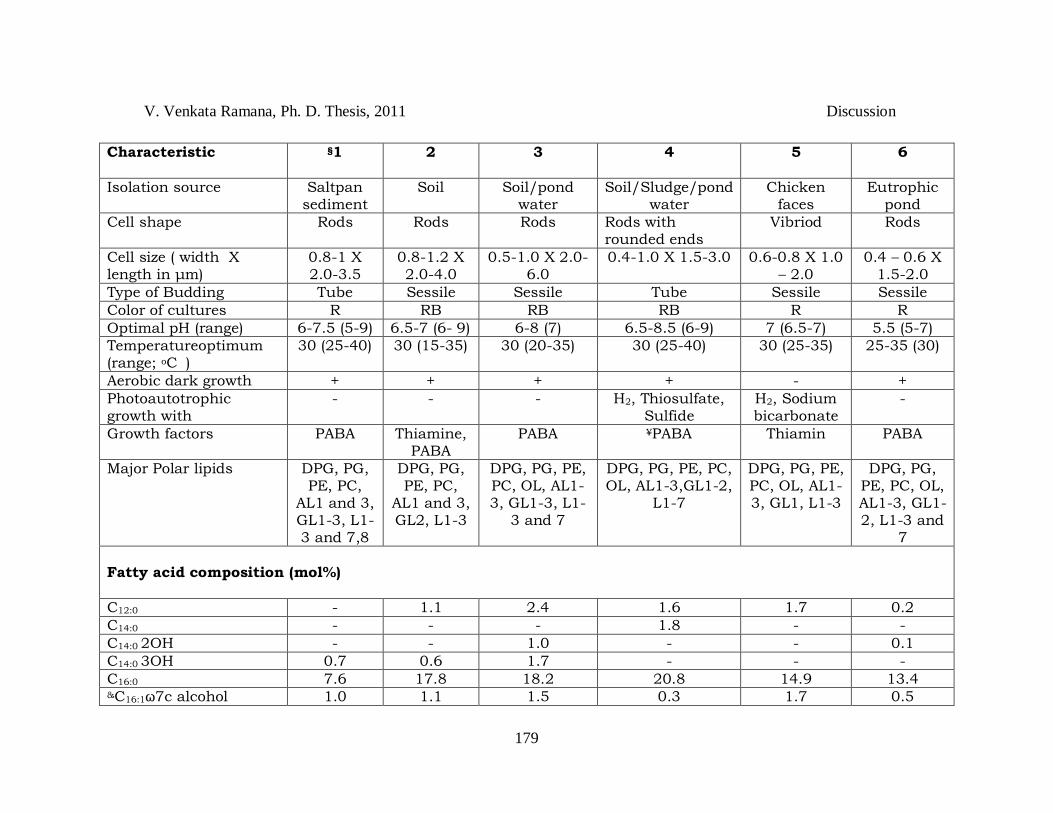
Characteristic §1 2 3 4 5 6
Isolation source Saltpan sediment
Soil Soil/pond water
Soil/Sludge/pond water
Chicken faces
Eutrophic pond
Cell shape Rods Rods Rods Rods with rounded ends
Vibriod Rods
Cell size ( width X length in μm)
0.8-1 X 2.0-3.5
0.8-1.2 X 2.0-4.0
0.5-1.0 X 2.0-6.0
0.4-1.0 X 1.5-3.0 0.6-0.8 X 1.0 – 2.0
0.4 – 0.6 X 1.5-2.0
Type of Budding Tube Sessile Sessile Tube Sessile Sessile
Color of cultures R RB RB RB R R
Optimal pH (range) 6-7.5 (5-9) 6.5-7 (6- 9) 6-8 (7) 6.5-8.5 (6-9) 7 (6.5-7) 5.5 (5-7)
Temperatureoptimum (range; oC
) 30 (25-40) 30 (15-35) 30 (20-35) 30 (25-40) 30 (25-35) 25-35 (30)
Aerobic dark growth + + + + - +
Photoautotrophic growth with
- - - H2, Thiosulfate, Sulfide
H2, Sodium bicarbonate
-
Growth factors PABA
Thiamine, PABA
PABA ¥PABA Thiamin PABA
Major Polar lipids DPG, PG, PE, PC,
AL1 and 3, GL1-3, L1-3 and 7,8
DPG, PG, PE, PC,
AL1 and 3, GL2, L1-3
DPG, PG, PE, PC, OL, AL1-3, GL1-3, L1-
3 and 7
DPG, PG, PE, PC, OL, AL1-3,GL1-2,
L1-7
DPG, PG, PE, PC, OL, AL1-3, GL1, L1-3
DPG, PG, PE, PC, OL, AL1-3, GL1-2, L1-3 and
7
Fatty acid composition (mol%)
C12:0 - 1.1 2.4 1.6 1.7 0.2
C14:0 - - - 1.8 - -
C14:0 2OH - - 1.0 - - 0.1
C14:0 3OH 0.7 0.6 1.7 - - -
C16:0 7.6 17.8 18.2 20.8 14.9 13.4 &C16:1ω7c alcohol 1.0 1.1 1.5 0.3 1.7 0.5
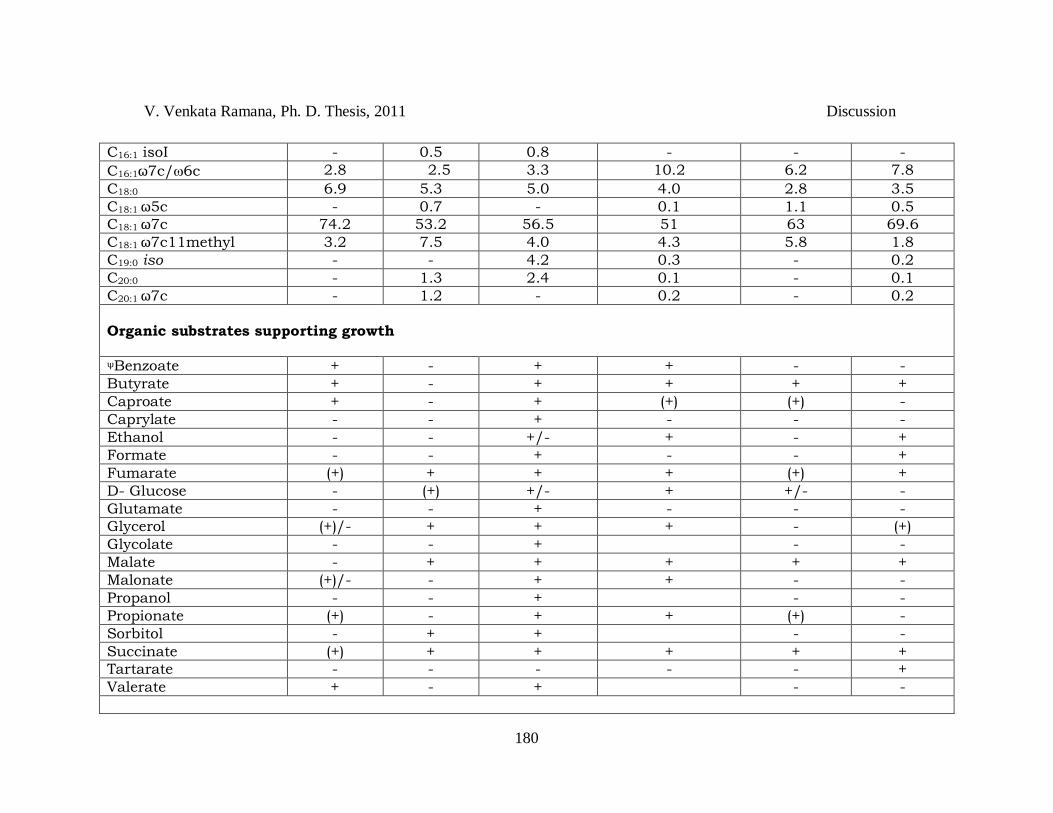
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
180
C16:1 isoI - 0.5 0.8 - - -
C16:1ω7c/6c 2.8 2.5 3.3 10.2 6.2 7.8
C18:0 6.9 5.3 5.0 4.0 2.8 3.5
C18:1 ω5c - 0.7 - 0.1 1.1 0.5
C18:1 ω7c 74.2 53.2 56.5 51 63 69.6
C18:1 ω7c11methyl 3.2 7.5 4.0 4.3 5.8 1.8
C19:0 iso - - 4.2 0.3 - 0.2
C20:0 - 1.3 2.4 0.1 - 0.1
C20:1 ω7c - 1.2 - 0.2 - 0.2
Organic substrates supporting growth ψBenzoate + - + + - -
Butyrate + - + + + +
Caproate + - + (+) (+) -
Caprylate - - + - - -
Ethanol - - +/- + - +
Formate - - + - - +
Fumarate (+) + + + (+) +
D- Glucose - (+) +/- + +/- -
Glutamate - - + - - -
Glycerol (+)/- + + + - (+)
Glycolate - - + - -
Malate - + + + + +
Malonate (+)/- - + + - -
Propanol - - + - -
Propionate (+) - + + (+) -
Sorbitol - + + - -
Succinate (+) + + + + +
Tartarate - - - - - +
Valerate + - + - -
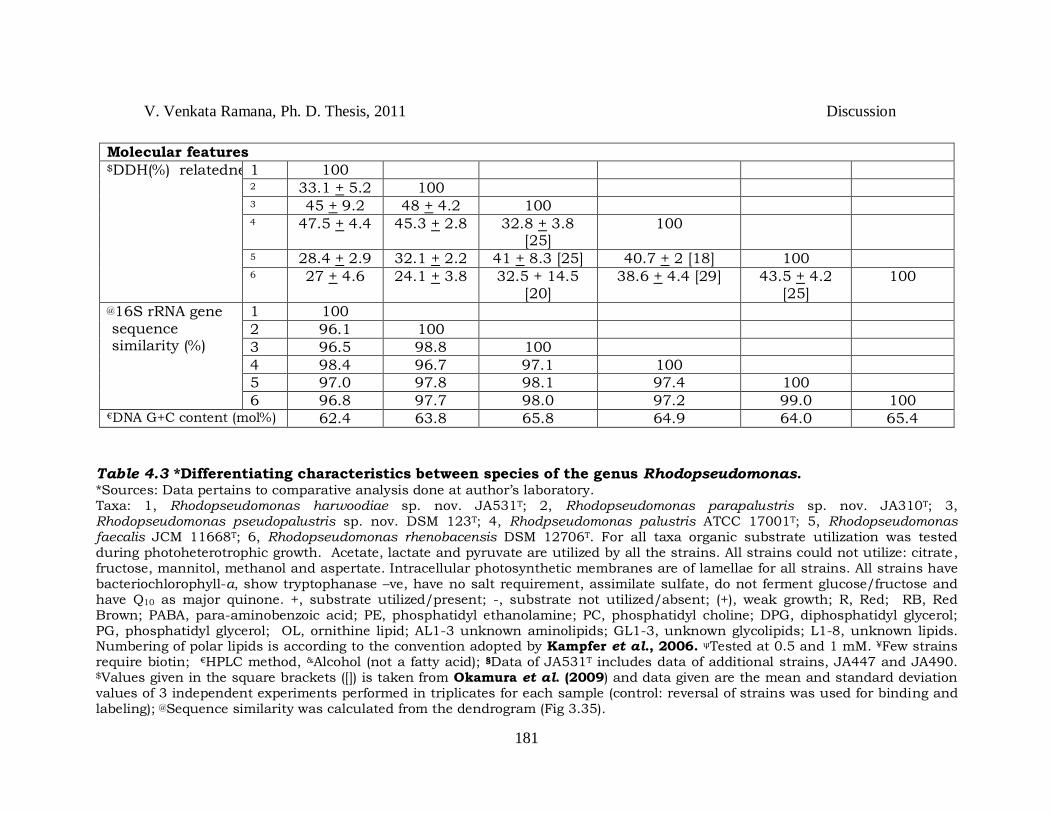
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
181
Molecular features $DDH(%) relatedness 1 100
2 33.1 + 5.2 100 3 45 + 9.2 48 + 4.2 100 4 47.5 + 4.4 45.3 + 2.8 32.8 + 3.8
[25] 100
5 28.4 + 2.9 32.1 + 2.2 41 + 8.3 [25] 40.7 + 2 [18] 100 6 27 + 4.6 24.1 + 3.8 32.5 + 14.5
[20] 38.6 + 4.4 [29] 43.5 + 4.2
[25] 100
@16S rRNA gene sequence similarity (%)
1 100
2 96.1 100
3 96.5 98.8 100
4 98.4 96.7 97.1 100
5 97.0 97.8 98.1 97.4 100
6 96.8 97.7 98.0 97.2 99.0 100 €DNA G+C content (mol%) 62.4 63.8 65.8 64.9 64.0 65.4
Table 4.3 *Differentiating characteristics between species of the genus Rhodopseudomonas. *Sources: Data pertains to comparative analysis done at author‘s laboratory. Taxa: 1, Rhodopseudomonas harwoodiae sp. nov. JA531T; 2, Rhodopseudomonas parapalustris sp. nov. JA310T; 3, Rhodopseudomonas pseudopalustris sp. nov. DSM 123T; 4, Rhodpseudomonas palustris ATCC 17001T; 5, Rhodopseudomonas faecalis JCM 11668T; 6, Rhodopseudomonas rhenobacensis DSM 12706T. For all taxa organic substrate utilization was tested during photoheterotrophic growth. Acetate, lactate and pyruvate are utilized by all the strains. All strains could not utilize: citrate, fructose, mannitol, methanol and aspertate. Intracellular photosynthetic membranes are of lamellae for all strains. All strains have bacteriochlorophyll-a, show tryptophanase –ve, have no salt requirement, assimilate sulfate, do not ferment glucose/fructose and have Q10 as major quinone. +, substrate utilized/present; -, substrate not utilized/absent; (+), weak growth; R, Red; RB, Red Brown; PABA, para-aminobenzoic acid; PE, phosphatidyl ethanolamine; PC, phosphatidyl choline; DPG, diphosphatidyl glycerol; PG, phosphatidyl glycerol; OL, ornithine lipid; AL1-3 unknown aminolipids; GL1-3, unknown glycolipids; L1-8, unknown lipids. Numbering of polar lipids is according to the convention adopted by Kampfer et al., 2006. ψTested at 0.5 and 1 mM. ¥Few strains require biotin; €HPLC method, &Alcohol (not a fatty acid); §Data of JA531T includes data of additional strains, JA447 and JA490. $Values given in the square brackets ([]) is taken from Okamura et al. (2009) and data given are the mean and standard deviation values of 3 independent experiments performed in triplicates for each sample (control: reversal of strains was used for binding and labeling); @Sequence similarity was calculated from the dendrogram (Fig 3.35).

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
182
Description of Rhodopseudomonas harwoodiae JA531T sp. nov.
Rhodopseudomonas harwoodiae (har.wo.o.di'a.e. N.L. fem. gen. n.
harwoodiae of Harwood, named after Professor Dr. Caroline S. Harwood,
a microbiologist who has significantly contributed to the aromatic
hydrocarbon metabolism of Rps. palustris)
Cells are rod shaped, 0.8-1m wide and 2.0-3.5 m long, budding
division, motile with lamellar ICM structures. Phototrophic cultures are
red. In vivo absorption spectra of intact cells exhibit maxima at 497, 590,
605, 863 nm. Bacteriochlorophyll-a and carotenoids of spirilloxanthin
series are present. No NaCl requirement (tolerates up to 4% [w/v]) The
type strain is mesophilic (range; 25-40 oC) with pH optimum at 6.0-7.5
(range; 5-9 pH). The preferred mode of growth is
photoorganoheterotrophy with a few organic compounds. Good growth
occurs on benzoate, butyrate, caproate, and valerate. Feeble growth
occurs with fumarate, glycerol, malonate, propionate and succinate as
carbon sources. Photoautotrophic and fermentative growth is absent.
Para-aminobenzoic acid is required as growth factor. C18:1 ω7c is the
major fatty acid with sufficient amount of C16:0, C18:0, C18:1ω7c11methyl,
C16:1ω7c/ω6c. Ubiquinone-10 is the major quinone. Cardiolipin,
phosphatidylethanolamine, phosphatidylglycerol, phosphatidylcholine,
two unidentified aminophospholipids and an unidentified aminolipid are
the major polar lipids. The DNA G+C content of the type strain is 62.4
mol% (by HPLC). Natural habitats are salterns of coastal ecosystem. The
type strain, JA531T (=NBRC 107575T =KCTC 5841T), is isolated from a
sediment of a saltpan at Kalambakam, Tamil Nadu, India.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
183
Strain JA310T
The enrichment and growth of strain JA310T occurred between 15-
350C with an optimum of 300C. Though the strain JA310T is
phylogenetically close to Rps. palustris, did not share the property of
benzoate utilization, which is a major characteristic feature of Rps.
palustris. Presence of small amounts of C14:03OH and C16:1 iso are
observed only in strains JA310T and Rps. palustris.
Though, strain JA310T showing >97% 16S rRNA gene sequence
similarity with nearest type strains (Imhoff and Caumette, 2004;
Stackebrandt and Ebers, 2006), its description as novel species is
supported by distant DNA-DNA homology (32.1 + 2.2) and polar lipids
composition (apart from phenotypic differences) (Table 4.3).
Polar lipid analysis revealed the presence of diphosphatidyl glycerol
(DPG), phosphatidyl glycerol (PG), phosphatidyl ethanolamine (PE),
phosphatidyl choline (PC) and an unidentified aminolipid (AL1) as the
major polar lipids in all the Rhodopseudomonas strains tested (Fig 4.3).
The major polar lipids composition observed in this study is in
concurrence with the earlier reports for the members of the genus
Rhodopseudomonas-Bradyrhizobium-213 Nitrobacter-Afipia (Hougary et
al., 2000; Imhoff et al., 1982). Major differences in the polar lipids among
the strains of Rhodopseudomonas were observed with respect to the
minor polar lipids. Glycolipid (GL3) was observed only with strains DSM
123T and JA531T which distinguish from other members of
Rhodopseudomonas, while presence of aminolipid (AL2) in strain DSM
123Tdelineates it from strain JA531T. Absence of a glycolipid (GL1) and

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
184
an aminolipid (AL2) differentiates strain JA310T with other members. A
few unidentified polar lipids (L4-L6) are unique to stain ATCC17001T (R.
palustris) and absence of a glycolipid (GL2) in stain JCM11668T (R.
faecalis) differentiates from the rest.
Like strain JA531T, strain JA310T also share genus specific
characters and showed interspecies variability (Table 4.3), which
necessitated the proposal of strain JA310T as a novel species named as
Rhodopseudomonas parapalustris sp, nov., (for sharing most of the
characteristics of Rps. palustris with few exception including separate
clustering with Rps. palustris).
Description of Rhodopseudomonas parapalustris JA310T sp. nov.
Rhodopseudomonas parapalustris (pa.ra.pa.lus'tris. Gr. prep. para,
alongside of, resembling; L. fem. adj. palustris, marshy, and also a
bacterial epithet; N.L. fem. adj. parapalustris, resembling to
Rhodopseudomonas palustris).
Cells are rod shaped, 0.8-1.2 m wide and 2.0-4.0 m long. Cells
are motile by means of polar flagella and multiply by budding. Cells of
old culture form rosettes. Internal photosynthetic membranes are
present as lamellar stacks parallel to the cytoplasmic membrane.
Phototrophically grown cultures are reddish brown. The in vivo
absorption spectrum of intact cells in sucrose exhibits maxima at 497,
590, 800 and 863. Bacteriochlorophyll a and carotenoids of the
spirilloxanthin series are present. The type strain is mesophilic (range;
15-35C). No NaCl requirement (tolerates up to 0.5% [w/v]) and growth
pH optima is from 6.5-7.0 (range, 6.0-9.0). The preferred mode of growth

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
185
is photoorganoheterotrophy with a few organic compounds. Malate,
succinate, pyruvate and sorbitol are good carbon sources. Phototrophic
growth also occurs on D-glucose. Benzoate, butyrate, caproate,
caprylate, ethanol, formate, glutamate, glycolate, malonate, proponol,
propionate, tartarate and valerate are not utilized as carbon sources.
Photoautotrophic and chemoautotrophic growth with thiosulfate / H2 /
sulfide / sulfur / sulfite / thioglycolate as the electron donor and
NaHCO3 as the carbon source is not observed. Fermentative growth does
not occur in the presence of glucose, fructose or pyruvate. Nitrate and
aspargine are used as nitrogen sources.
Thiamine and para-aminobenzoic acid are required as growth
factors. C18:1 ω7c is the major fatty acid with sufficient amounts of C12:0,
C16:0, C18:1 ω7c 11methyl, C18:0 and C16:1 ω7c/ω6c. Ubiquinone-10 is the
major Quinone. Cardiolipin, phosphatidylethanolamine,
phosphatidylglycerol and two unidentified aminophospholipids are the
major polar lipids. The G+C of genomic DNA is 63.8 mol% (by HPLC).
Natural habitats are surface soils near lake at high altitudes. The type
strain JA310T (=NBRC 106083T =KCTC 5839T) was isolated from a
sediment soil sample near Pangong Lake at Leh, Jammu and Kashmir,
India.
Strain DSM 123T
According to Okamura et al., (2009), ATCC17001T and DSM 123T
which are known to be type strain Rps. palustris are not the same.
Hence, both these strains were obtained from respective culture
collections and subjected to polyphasic characterization. The differences

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
186
exhibited by the strain DSM 123T with all other types strains of the
genus Rhodopseudomonas including DNA homology data (Table 4.3)
supported the statement made by Okamura et al., 2009 and necessitated
the description of DSM 123T as a novel species and named as
Rhodopseudomonas pseudopalustris (Venkata Ramana et al., MS under
revision).
Description of Rhodopseudomonas pseudopalustris DSM 123T sp.
nov.
Rhodopseudomonas pseudopalustris (pseudês, Gr. adj. false; L. fem. adj.
palustris, marshy, and also a bacterial epithet; N.L. fem. adj.
pseudopalustris, the false [Rhodopseudomonas] palustris)
Cells are rod shaped, 0.8-1μm wide and 2.0-6.0 μm long, budding
division, motile and have lamellar ICM structures. Phototrophic cultures
are reddish brown. In vivo absorption spectra of intact cells exhibit
maxima at 497, 590, 605, 865 nm. Bacteriochlorophyll-a and
carotenoids of spirilloxanthin series are present. No NaCl requirement
(tolerates up to 0.5% [w/v]) The type strain is mesophilic (range; 20-35
oC) with pH optimum at 7.0 (range; 6-8 pH). The preferred mode of
growth is photoorganoheterotrophy with a few organic compounds. Good
growth occurs on benzoate, butyrate, caproate, caprylate, fumarate,
glutamate, glycerol, glycolate, malate, malonate, proponal, propionate,
sorbitol, succinate and valerate. Feeble growth occurs with ethanol and
D-glucose. Photoautotrophic and fermentative growth is absent. Para-
aminobenzoic acid is required as growth factor. C18:1ω7c is the major
fatty acid 328 with sufficient amount of C16:0, C18:0,

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
187
C18:1ω7c11methyl and C16:1ω7c/ω6c. Ubiquinone-10 is the major
quinone. Diphosphatidyl glycerol, phosphatidyl glycerol, phosphatidyl
ethanolamine, phosphatidyl choline, ornithine lipid and an unidentified
aminolipid (AL1) are the major polar lipids. The DNA G+C content of the
type strain is 65.8 mol% (by HPLC). The type strain DSM123T
(=NBRC100419T; confirmed by Okamura et al., 2009) was isolated and
deposited at DSMZ by van Niel (1944).
GENUS Blastochloris (Blc)
Blastochloris (Blc) is a phototrophic genus of the family
Hypomicrobiaceae, order Rhizobiales and class Alphaprotebacteria.
Members of the genus Blastochloris have unique identity due to the
presence of BChl b, thus help in identification till genus level and
differentiating from rest of the purple non-sulfur bacteria which have
BChl a alone (Hiraishi, 1997). Strains of the genus Blastochloris also
found to have a little amount of BChl a (peak at 812 nm), since it is the
reaction centre chlorophyll even in those bacteria containing other
bacteriochlorophylls as predominant components. The genus
Blastochloris currently comprises of two validly published species names,
Blc. viridis and Blc. sulfoviridis. The two species of Blastochloris were
previously assigned to the genus Rhodopseudomonas (Drews and
Giesbrecht, 1966; Keppen and Gorlenko, 1975) based on significant
similarities in cell morphology and cell division to Rps. palustris.
Subsequently, significant differences in 16S rRNA gene sequences with
respect to other Purple nonsulfur bacteria was the reason for

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
188
reconsideration of the taxonomic position of these bacteria and
reevaluation of phenotypic differences from the type species of
Rhodopseudomonas, leading to their reassignment to the new genus
Blastochloris as Blc. viridis and Blc. sulfoviridis (Kawasaki et al., 1993b;
Hiraishi, 1997).
The sequence similarity between the 16S rRNA genes of Blc. viridis
and Blc. sulfoviridis was 98.7%, which suggest that the two species are
phylogenetically closely related organisms. However, their separation and
delineation as different species was confirmed based on the supportive
results of DNA-DNA hybridization values of 38% between the two species
(Hiraishi, 1997).
Their green to olive green coloration, together with the
characteristic long wavelength absorption maxima between 1015-1035
nm, which is due to bacteriochlorophylls b, clearly separates Blstochloris
species from other phototrophic Alphaproteobacteria.
The interspecies differentiating characteristics of the genus
Blastochloris include type of budding, colour of the culture, optimum pH,
sulfate assimilation, phototrophic growth modes, growth factor
requirements, utilization of carbon source, G+C content, major quinones,
cellular fatty acid composition and DNA-DNA relatedness.
Strain JA248T
In the present study, strain JA248T which was isolated from the epilithic
biofilm (Fig. 3.22) shares genus specific characters of Blastochloris such
as rod-shaped cells, asymmetric mode of growth and cell division,
formation of rosette like aggregates, presence of bacteriochlorophylls b

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
189
and the lamellar structure of internal photosynthetic membranes and,
also phylogenetically close to the members of the genus Blastochloris.
The same strain JA248T differentiate with Blc. sulfoviridis and Blc. viridis
regarding habitat of isolation, colour of the culture, range and optimum
of temperature for growth, presence of aerobic dark growth, quinone
composition, carotenoid composition, utilization of carbon source/e-
donor, cellular fatty acid composition, 16S rRNA gene sequence
dissimilarity (Fig. 3.27), G+C content, MLSA and Fatty acids (Table 4.4).
Particularly, the strain JA248T grows well at temperature range of 15-
400C with an optimum of 250C, whereas the other two species cannot
grow below 250C. The size of the cells of strain JA248T is twice the size of
other two species of Blastochloris and the colour is yellowish green
whereas the other two are green to olive green. Few characteristics of
strain JA248T were found to coincide with Blc. viridis such as sulfate
assimilation, presence of carotenoid lycopene, absence of
photolithoautotrophy and presence or absence of few minor cellular fatty
acids. Some characters of strain JA248T were found to coincide with Blc.
sulfoviridis, are sessile type of budding, maximum temperature tolerance,
vitamin requirement, quinones, presence of carotenoid 7, 8, 11, 12- tetra
hydro lycopene and lack of few minor cellular fatty acids.
Strain JA248T showed closest 16S rRNA gene sequence similarity
with the type strain Blastochloris sulfoviridis DSM 729T (98.5% sequence)
and Blastochloris viridis DSM 133T (98.1%) (Table 3.2). Though strain
JA248T is showing close 16S rRNA gene sequence similarity with Blc.
sulfoviridis, it clustered with Blc. viridis in the phylogenetic tree (Fig.
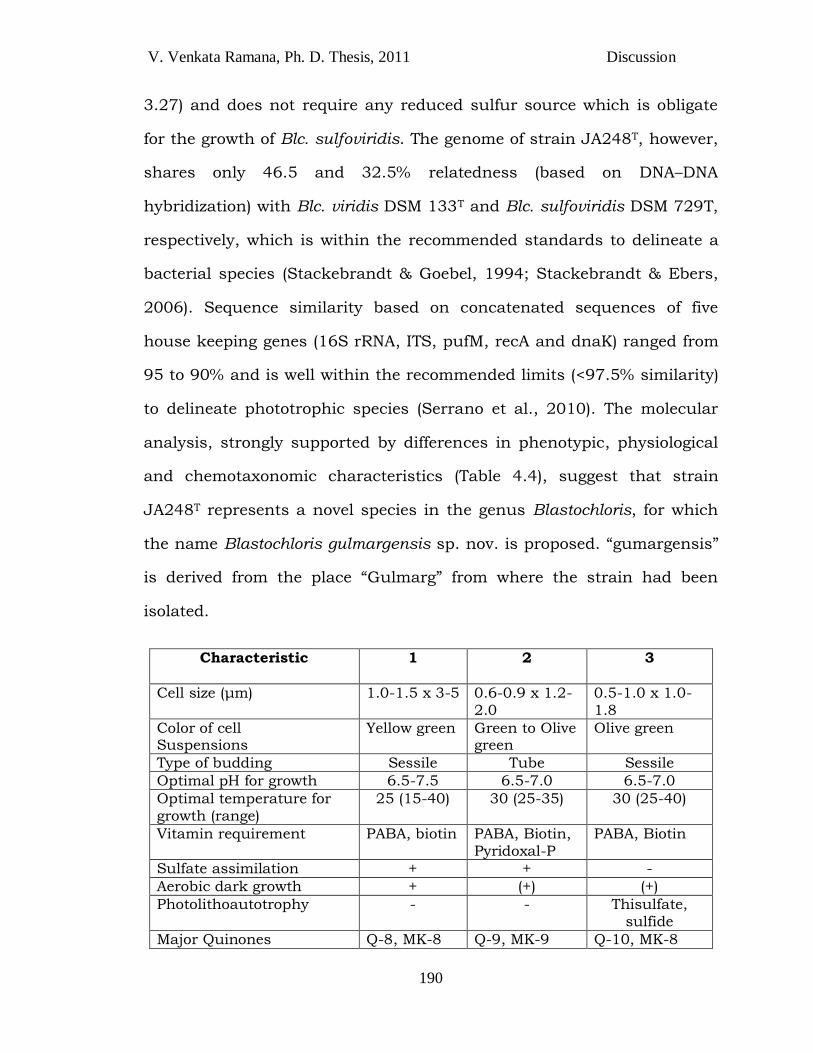
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
190
3.27) and does not require any reduced sulfur source which is obligate
for the growth of Blc. sulfoviridis. The genome of strain JA248T, however,
shares only 46.5 and 32.5% relatedness (based on DNA–DNA
hybridization) with Blc. viridis DSM 133T and Blc. sulfoviridis DSM 729T,
respectively, which is within the recommended standards to delineate a
bacterial species (Stackebrandt & Goebel, 1994; Stackebrandt & Ebers,
2006). Sequence similarity based on concatenated sequences of five
house keeping genes (16S rRNA, ITS, pufM, recA and dnaK) ranged from
95 to 90% and is well within the recommended limits (<97.5% similarity)
to delineate phototrophic species (Serrano et al., 2010). The molecular
analysis, strongly supported by differences in phenotypic, physiological
and chemotaxonomic characteristics (Table 4.4), suggest that strain
JA248T represents a novel species in the genus Blastochloris, for which
the name Blastochloris gulmargensis sp. nov. is proposed. ―gumargensis‖
is derived from the place ―Gulmarg‖ from where the strain had been
isolated.
Characteristic 1 2 3
Cell size (μm) 1.0-1.5 x 3-5 0.6-0.9 x 1.2-2.0
0.5-1.0 x 1.0-1.8
Color of cell Suspensions
Yellow green Green to Olive green
Olive green
Type of budding Sessile Tube Sessile
Optimal pH for growth 6.5-7.5 6.5-7.0 6.5-7.0
Optimal temperature for growth (range)
25 (15-40) 30 (25-35) 30 (25-40)
Vitamin requirement PABA, biotin PABA, Biotin, Pyridoxal-P
PABA, Biotin
Sulfate assimilation + + -
Aerobic dark growth + (+) (+)
Photolithoautotrophy - - Thisulfate, sulfide
Major Quinones Q-8, MK-8 Q-9, MK-9 Q-10, MK-8
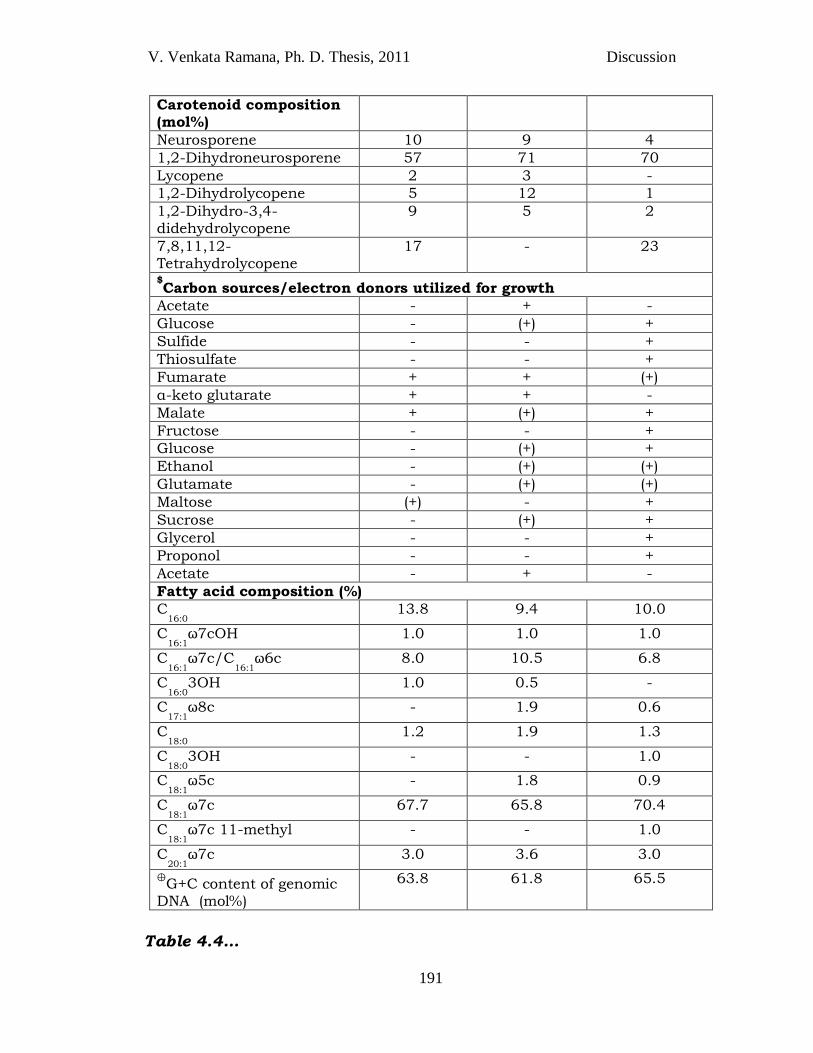
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
191
Carotenoid composition (mol%)
Neurosporene 10 9 4
1,2-Dihydroneurosporene 57 71 70
Lycopene 2 3 -
1,2-Dihydrolycopene 5 12 1
1,2-Dihydro-3,4-didehydrolycopene
9 5 2
7,8,11,12-Tetrahydrolycopene
17 - 23
$Carbon sources/electron donors utilized for growth
Acetate - + -
Glucose - (+) +
Sulfide - - +
Thiosulfate - - +
Fumarate + + (+)
α-keto glutarate + + -
Malate + (+) +
Fructose - - +
Glucose - (+) +
Ethanol - (+) (+)
Glutamate - (+) (+)
Maltose (+) - +
Sucrose - (+) +
Glycerol - - +
Proponol - - +
Acetate - + -
Fatty acid composition (%)
C16:0
13.8 9.4 10.0
C16:1
ω7cOH 1.0 1.0 1.0
C16:1
ω7c/C16:1
ω6c 8.0 10.5 6.8
C16:0
3OH 1.0 0.5 -
C17:1
ω8c - 1.9 0.6
C18:0
1.2 1.9 1.3
C18:0
3OH - - 1.0
C18:1
ω5c - 1.8 0.9
C18:1
ω7c 67.7 65.8 70.4
C18:1
ω7c 11-methyl - - 1.0
C20:1
ω7c 3.0 3.6 3.0
⊕G+C content of genomic
DNA (mol%)
63.8 61.8 65.5
Table 4.4…

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
192
Table 4.4 *Differentiating characteristics of species of the genus Blastochloris. *Sources: Data pertains to comparative analysis done at authors‘ laboratory. 1,
strain JA248T; 2, Blc. viridis DSM133T; 3, Blc. sulfoviridis DSM729T. $All the
results are analysis after 3 subcultures with the respective substrates and compared against control (without the test substrate). Substrates that were utilized by all the strains include- pyruvate and succinate; those that could not be utilized include- tartrate, citrate, formate and benzoate. Other similarities of the three strains include rod shape, cell division by budding, presence of a single polar flagellum, rosette formation, lamellar internal membrane and presence of BChl b. Type strains do not have NaCl requirement for growth. &The reassociation values are the mean + 1 SD of two independent
determinations. Symbols: +, substrate utilized; -, substrate not utilized; (+), poorly utilized; PABA = para-aminobenzoic acid; NT, not tested, ND, not
determined. ⊕HPLC analysis.
Description of Blastocholris gulmargensis sp. nov.
Blastochloris gulmargensis (gul.mar.gen'sis. N.L. fem. adj. gulmargensis
pertaining to Gulmarg, a hilly resort in Kashmir, India, from where the
type strain was isolated).
Cells are rod shaped, 1.0-1.5 μm wide and 3.0-5.0 μm long,
budding division and motile. Growth occurs well under anaerobic
conditions in the light. Lamellar intracytoplasmic membrane structures.
Phototrophic cultures are yellow green. The in vivo absorption spectra of
intact cells exhibit maxima at 404, 455, 485, 605, 812, 1022 nm.
Bacteriochlorophyll b and carotenoids of spirilloxanthin series are
present. The type strain is mesophilic (25oC) and grows well at a pH
optimum of 6.5-7.5. The preferred mode of growth is
photoorganoheterotrophy with a few organic compounds. Good growth
occurs on fumarate, pyruvate, succinate, α-ketoglutarate and malate.
Photoautotrophic and fermentative growth are absent. PABA and biotin

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
193
are required as growth factors. C18:1
ω7c, C16:0
are the dominant fatty
acids; sufficient amounts of C16:1 ω7c/C16:1 ω6c and C20:1 ω7c are also
found. Major quinones are Q-8 and MK-8. The DNA G + C content of the
type strain is 63.8 mol% (by HPLC). Natural habitats are cold sulfur
springs. The type strain, JA248T (=JCM 14795
=DSM 19786), was
isolated from an epilithic biofilm sample from a cold sulfur spring at
Gulmarg, Jammu and Kashmir, India.
GENUS Ectothiorhodospira (Ect)
Ectothiorhodospira is a phototrophic genus of purple sulfur
bacteria classified in the family Ectothiorhodispiriaceae, order
Chromatiales, class Gammaproteobacteria. Members of the family
Ectothiorhodospiraceae form a distinct lineage from Chromatiaceae on the
basis of 16S rRNA gene sequence analysis (Imhoff and Süling, 1996).
Among the 13 genera recognized in the family Ectothiorhodospiraceae,
only 4 genera (Ectothiorhodosinus, Ectothiorhodospira, Halorhodospira
and Thiorhodospira) are represented by members of true phototrophic
species(http://www.bacterio.cict.fr/classifgenerafamilies.html#Ectothiorh
odospiraceae)
All the phototrophic members of the family Ectothiorhodospiraceae
accumulate sulfur granules outside the cell and thus phenotypically
distinct from the members of the Chromatiaceae (Imhoff, 2006b). After
reclassification due to revision of species delineation, the genus
Ectothiorhodospira was left with only 4 species (Ect. mobilis, Ect.
shaposhnikovii, Ect. marina and Ect. Haloalkaliphila. E. vacuolata and E.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
194
marismortui are considered as heterotypic synonyms of Ect.
shaposhnikovii and Ect. mobilis respectively based on DNA-DNA
reassociation and ribotyping (Ventura et al., 2000). With the
subsequently described Ect. variabilis (Gorlenko et al. 2009), the number
of species became five.
The species of the genus Ectothiorhodospira represent by few
characteristic features such as rod-shaped or vibrioid or true spiral
shaped cells, gram-negative, motile by polar tuft of flagella, multiply by
binary fission and many contain gas vesicles. Internal photosynthetic
membranes are lamellar type and photosynthetic pigments are
bacteriochlorophyll-a and carotenoides. Sulfide oxidized to sulfate, with
S0 as an intermediate, which is deposited outside the cells. Most species
live in marine and saline environments that contain sulfide and have
slight to extreme alkaline pH and occasionally found in soil.
Strain JA430T
Strain JA430T was isolated from a solar saltern, a novel habitat,
though Ectothiorhodospira spp have been reported from other marine and
halophilic environments (Imhoff, 2006). Strain JA430T was enriched in
the medium supplemented with pyruvate / bicarbonate as carbon source
and sulfide as electron donor. Since, strain JA430T did not have
photolithoautotrophy, it must have used the pyruvate as carbon source
and reduced sulfide as electron donor, in contrast to all other species of
Ectothiorhodospira which are having photolithoautotrophic growth. By
not supplementing the medium with an organic carbon source, the strain
could have been missed. This may explain why all the species described

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
195
till now are photoautotrophic. Thus, it becomes clear that restricting to
one medium composition will restrict the discovery of novel species.
A 16S rRNA gene sequence similarity of < 97% with the species of
the genus Ectothiorhodospira (Table 3.2) has clearly indicated the strain
JA430T representing a novel species, which necessitated detailed
polyphasic characterization. Though phylogenetic tree constructed by
16S rRNA gene sequence of strain JA430T (Fig. 3.21) show closest
similarity with the type strain Ect. variablis WN22T (96.1% sequence
similarity), it clustered with type strain Ect. mobilis DSM 237T.
Polyphasic characterization of strain JA430T has revealed its
differentiating characteristics with existing type strains of the genus
Ectothiorhodospira such as size of cells, colour of the cell suspension and
type of carbon source utilization. In addition, requirement of vitamins p-
aminobenzoic acid, pantothente and pyridoxal phosphate was observed
for the growth of strain JA430T, whereas none of the other strains of the
same genus require vitamin source. Sulfate assimilation was not
observed in any of the strains except JA430T and Ect. variablis. In
contrast to the property of having diazotrophy in species of the genus
Ectothiorhodospira, strain JA430T did not have the same.
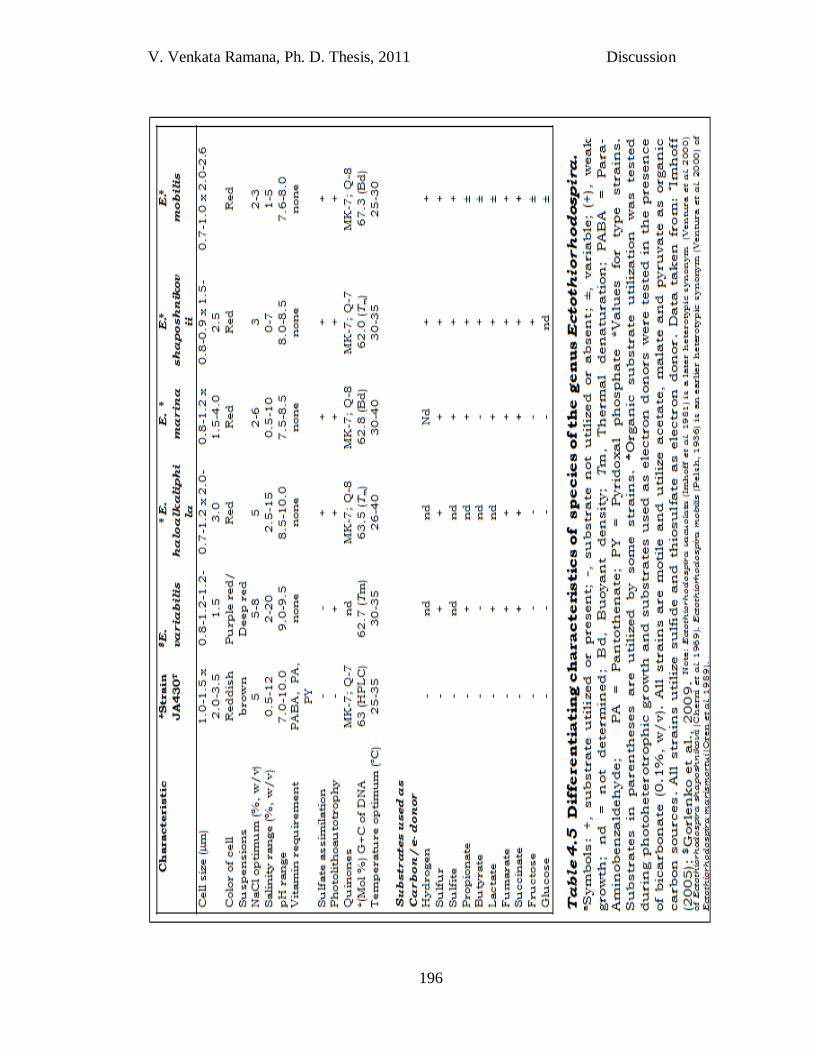
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
196

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
197
Apart from phenotypic and physiological differences, 16S rRNA
gene sequence similarity (96.1%) of strain JA430T is also within the
recommended standards to delineate a bacterial species (Stackebrandt
and Goebel, 1994; Stackebrandt and Ebers, 2006). The above differences
of strain JA430T from other Ectothiorhodospira spp. (Table 4.5) enable
description of a novel species of the genus Ectothiorhodospira as
Ectothiorhodospira salini.
Description of Ectothiorhodospira salini sp. nov.
Ectothiorhodospira salini (sa.li'ni. L. gen. n. salini, of a salt-cellar).
Cells are vibrioid to spiral shaped, 1.0–1.5 m wide and 2.0–3.5 m long.
Cells are motile by means of polar flagella and divide by binary fission.
Growth occurs under anaerobic conditions in the light. Internal
photosynthetic membranes are of lamellar type arranged parallel to the
cytoplasmic membrane. Phototrophic cultures are reddish brown. The in
vivo absorption spectra of intact cells exhibit maxima at 311, 377, 512-
518, 590-593, 797 and 860 nm. Bacteriochlorophyll a and carotenoids;
spirilloxanthin, rhodopin, anhydrorhodovibrin, tetrahydrolycopene and
rhodovibrin are present. The type strain is mesophilic (300C) growing at a
pH optimum at 7.5 (range, pH 7.0-10.0) and requires 5% NaCl for
optimal growth (NaCl range, 0.5-12.0%). The preferred mode of growth is
photoorganoheterotrophy with a few organic compounds. Good growth
occurs on acetate, malate and pyruvate. Photoautotrophic and
fermentative growth is absent. PABA, pantothenate and pyridoxal
phosphate are required as growth factors. C18:1ω7c is the dominant fatty
acid; sufficient amounts of C16:0, C19:0cyclo8c and C16:1 ω7c/C16:16c are

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
198
also found. Major quinones are MK-7 and Q-7. The DNA G+C content of
the type strain is 63 mol% (by HPLC). Natural habitats are solar salterns.
The type strain JA430T (=NBRC105915 =KCTC 5805) was isolated from
sediment sample from a solar saltern at kanyakumari, Tamil Nadu,
India.
STRAINS OF THE GENUS Marichromatium (Mcr)
Marichromatium (Mcr) is a phototrophic genus of purple sulfur
bacteria, classified in the family Chromaticaea, order Chromatiales, class
Gammaproteobactreia. Marichromatium species are distinct from other
members of the family Chromatiaceae by their specific salt requirement
and the very high mol% G+C content of their DNA. Species of this genus
have previously been classified as belonging to the genus Chromatium
(Pfennig and Truper, 1974; Pfennig, 1989b). However because of
significant phenotypic and genetic differences to true Chromatium spp.
and other members of the family Chromatiaceae, they have been
transferred to the new genus Marichromatium (Imhoff et al., 1998b).
Till date, 5 Marichromatium species have been validly published
(http://www.bacterio.cict.fr/m/marichromatium.html). Mcr. gracile
biotype thermosulfidiphilum (Serrano et al., 2009) is a newly described
biotype, the only one among anoxygenic phototrophic bacteria. The
common property among all the members of the genus Marichromatium
is that they represent the true marine species with wide growth
capabilities. Marichromatium species had been distinguished based on
the differences in colour of cell suspension, substrate utilization, cell

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
199
size, salt responses, carotenoides group, growth modes and whole
genome DNA-DNA hybridization.
In the present study, the highest number of strains of purple
sulfur bacteria (JA550, JA548, JA349, JA546, JA350, JA387, JA547,
JA536, JA637, JA552, JA537, JA551, JA540, JA638, JA553 and JA260)
isolated belong to the genus Marichromatium (16 strains out of 24 = 67%
of PSB) and all were from the marine habitats. Most of the strains (8 out
of 16) among the above showed closest 16S rRNA gene sequence
similarity (Table 3.2) to Marichromatium gracile revealing their
predominance.
Though the species of the genus Marichromatium (except Mcr.
bheemlicum and Mcr. fluminis) share more than 99% 16S rRNA gene
sequence similarity, they exhibit differences in ribotyping (Acinas et al.,
2004). The DNA-DNA hybridization of the Marichromatium species
(Serrano et al., 2009) revealed that Mcr. gracile and Mcr. purpuratum are
71% similar which are to be considered as heterotypic synonyms.
However, this conflict is resolved by the use of MLSA approach where
they have used concatenated sequences of 6 protein-coding genes (gyrB,
recA, fusA, dnaK, pufM and soxB) together with 16S rRNA gene and the
internal transcriber spacer ITS region for comparison (a minimum use of
7 genes is recommended for MLSA [Gevers et al., 2005; Maiden et al.,
1998]) and have concluded that the established taxonomy of the genus
Marichromatium with four validly described species (the recently
described species, Marichromatium fluminis (Sucharita et al., 2010a) is
not included in their analysis) is acceptable.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
200
A noteworthy observation in the present study is that use of
glutamate as carbon source in the media enhanced the enrichment of
most strains of Mcr. purpuratum, whereas pyruvate as carbon source
enriched strains of other species of the genus Marichromatium. Based on
16S rRNA gene sequence similarity (Table 3.2), strain JA387 is close to
Mcr. indicum (Arunasri et al., 2005), while JA536 and JA637 are close to
Mcr. gracile DSM 203T (Table 3.2). But all these three strains closely
clustered with Mcr. purpuratum in the phylogenetic tree (Fig. 4.4).
Percentage of 16S rRNA gene sequences similarity of remaining 9 strains
of Marichromatium coincides with the percentage of distance in
phylogenetic tree (Fig. 4.4). This kind of unconcurrent result
(phylogenetic distances caliculated by dendrogram and by 16S rRNA
Blast) is observed with genomospecies, because they are genotypically
very close to each other.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
201
Fig. 4.4 Dendrogram showing phylogenetic relationship of strains ((JA550, JA548, JA349, JA546, JA350, JA387, JA547, JA536, JA637, JA552, JA537, JA551, JA540, JA638, JA553 and JA260) with phototrophic members of Gammaproteobacteria. (Tree was constructed using NJ method of MEGA4 software. The gene bank accession numbers for 16S rRNA gene sequences are shown in parentheses. Bar, one nucleotide substitution for 100 nucleotides. Bootstrap values below 50 have been removed from
the dendrogram)

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
202
STRAINS OF THE GENUS Rhodovulum (Rdv)
Till date, the genus Rhodovulum (Rdv) comprises twelve validly
published species: Rdv. sulfidophilum, Rdv. adriaticum, Rdv.
euryhalinum, Rdv. strictum, Rdv. iodosum, Rdv. robiginosum, Rdv.
steppense, Rdv. lacipunicei, Rdv. kholense, Rdv. imhoffii, Rdv.
visakhapatnamense and Rdv. marinum
(http://www.bacterio.cict.fr/qr/rhodovulum.html). The major
differentiating characterstic of the genus Rhodovulum with other genera
is 16S rRNA gene sequence dissimilarity. Species of this genus (Rdv.
sulfidophilum, Rdv. adriaticum) have formerly been included in the genus
Rhodopseudomonas. The recognition of morphologically and
chemotaxonomically distinct characteristics and phylogenetic analysis
led to their separation from Rhodopseudomonas and classification in the
genus Rhodobacter (Imhoff et al., 1984b). Later, the data of 16S rRNA
gene sequences phylogenetically separated fresh water and marine forms
of the genus Rhodobacter. Hence, Rhodobacter sulfidophilum and
Rhodobacter adriaticum were transferred into the new genus Rhodovulum
(Hiraishi and Ueda, 1994).
In the present study, four strains of genus Rhodovulum, JA255,
JA390, JA545 and A595 were isolated (Fig. 3.2). Based on rapid typing
(Table 3.2), strain JA595 was sorted and subjected for detailed
polyphasic characterization. Strian JA595 showed distinct physiological
and 16S rRNA gene sequence similarities with the species of the genus
Rhodovulum, but exhibited all similar characteristics and close 16S rRNA
gene sequence similarity (99.5%) with strain JA580T (which is recently

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
203
proposed as a novel species named Rhodovulum phaeolacus [Lakshmi et
al., 2011]) (Fig. 4.5). Therefore, strain JA595 is propsed as an additional
strain of Rhodovulum phaeolacus.
Fig. 4.5 Phylogenetic tree based on almost-complete 16S rRNA gene
sequences showing the relationship of strain JA595T within the class
Alphaproteobacteria. The tree was constructed by the neighbor joining method using the MEGA4 software and rooted by using Rhodospirillum rubrum as out group as the out-group. Numbers at nodes represent bootstrap values (based on 100 resamplings). The GenBank accession numbers for 16S rRNA gene sequences are shown in parentheses. Bar, 1 nucleotide substitutions per 100 nucleotides. Other three strains JA255, JA390 and JA545 are not included in the dendrogram for having insufficient length and quality of 16S rRNA gene sequence.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
204
STRAINS OF THE LESS PREDOMINANT GENERA: Rhodothalassium,
Rubrivivax, Roseospira, Rhodomicrobium, Rhodoplanes AND
Rhodospirillum.
The obligate requirement for salt, cell morphology and internal
membrane structure clearly distinguish Rhodothalassium from other
spiral shaped phototrophic Alphaproteobacteria. Strain JA389 and JA473
of genus Incertae Sedis Rhodothalassium, were isolated from halophilic
(saltpan) habitats, like that of existing type strain Rhodothalassium
salexigens DSM 2132T, which prooved their wide occurence in
hypersaline environments. Based on rapid typing (Table 3.2), strains
JA389 and JA473 are similar to Rhodothalassium salexigens (Table 4.6).
Two strains of the genus Rubrivivax, 2 of Roseospira and one each
of Rhodomicrobium, Rhodoplanes and Rhodospirillum were found to be
very similar to their respective closest type strains (Table 3.2), based on
colour of the culture, pigments and carotenoides, microscopic
observation and 16S rRNA gene sequence similarity. Hence, they were
considered as strains of their respective closest type strains. Now the
genus Rhodomicrobium consists of only one validly published species
Rhodomicrobium vanielli. Isolation of strain of Rhodomicrobium vannielli
(present study) is the first report from India.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
205
STRAINS OF THE GENERA Thiophaeococcus, Allochromatium
Thiohalocapsa.
Genera Thiophaecoccus, Allochromatium and Thiohalocapsa are belong to
the purple sulfur bacteria classified under the family Chromatiales, order
Rhodocyclales of the class Betaproteobacteria distributed under the class
Fig. 4.6 Phylogenetic tree based 16S rRNA gene sequences showing the relationship of strains JA389 and JA473 within the class Alphaproteobacteria. The tree was constructed by the neighbor joining method within the MEGA4 software and rooted by using Escherichia coli as the out-group. Numbers at nodes represent bootstrap values (based on 100 resamplings). The GenBank accession numbers for 16S rRNA gene sequences are shown in parentheses. Bar, 2
nucleotide substitutions per 100 nucleotides.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
206
Betaproteobacteria. Thiophaeococcus is a recently described (Anil Kumar
et al., 2008b) genus isolated from brackish water of mangrove forest
(Bhitarkanika) from India on the east coast, whereas the strains JA535,
JA541 and JA592 of Thiophaeococcus (of present study), were isolated
from microbial mat and mud of saltpan. All strains of Thiophaeococcus
have obligate requirement for NaCl. 16S rRNA gene sequence similarity of
strains JA541, JA592 and JA535 (from the present study) with
Thiophaeococcus mangrovi is 95.5%, 95.6% and 95.7% respectively (
Table 3.2 and Fig. 4.7). All these three strains were isolated from Tamil
Nadu coast. The similarity between each of the three strains is 99%.
These three strains are yet to be characterized completely to reveal the
differentiating characteristics at species level.
Strains JA543 and JA549 have similar morphological and
microscopic characteristics with Allochromatium phaeobacterium JA144T,
Allochromatium vinosum DSM 180T respectively. 99.4% similarity of 16S
rRNA gene sequence of strain JA549 with Allochromatium vinosum (Table
3.2 and Fig. 4.7) also supports close relatedness with type strain.
One strain JA534 of the genus Thiohalocapsa was isolated which is
from seashore, Akkarpettai,Tamil Nadu and its 16S rRNA gene sequence
similarity is 98.9% with Thiohalocapsa marina JA142T (Table 3.2).
Thiohalocapsa spp. abound in purple ponds imparting bright purple
colour to the water body (Fig. 4.1).
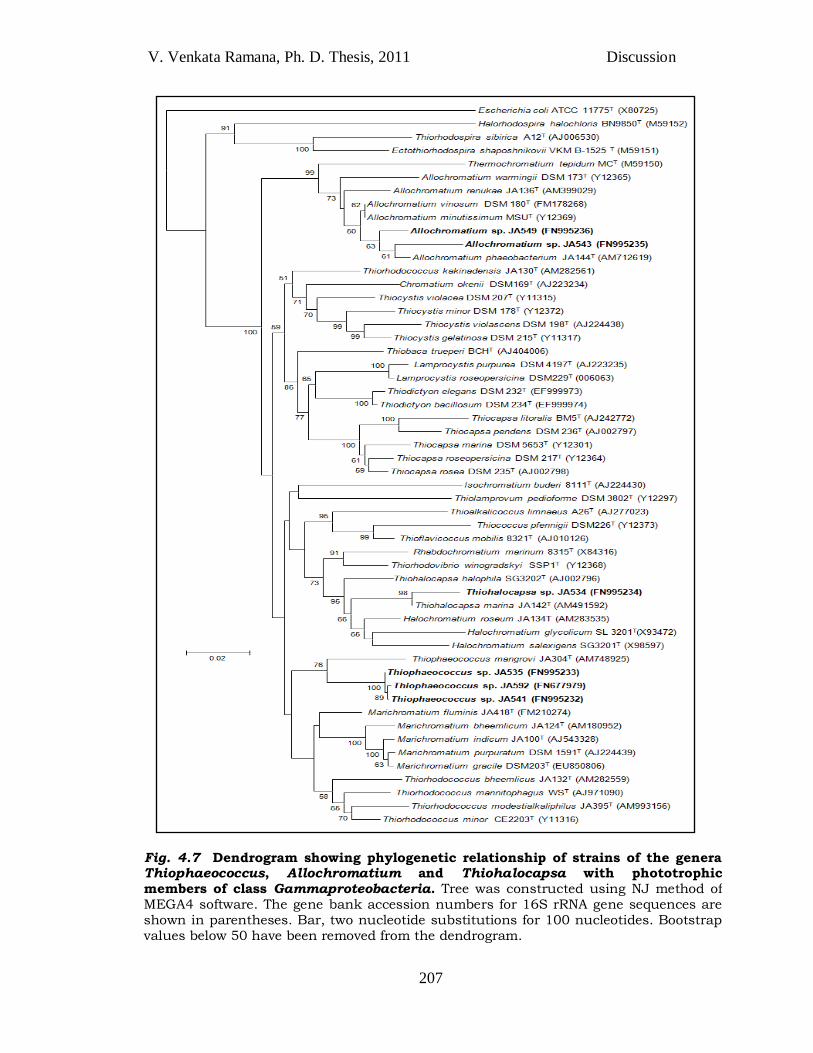
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
207
Fig. 4.7 Dendrogram showing phylogenetic relationship of strains of the genera Thiophaeococcus, Allochromatium and Thiohalocapsa with phototrophic members of class Gammaproteobacteria. Tree was constructed using NJ method of MEGA4 software. The gene bank accession numbers for 16S rRNA gene sequences are shown in parentheses. Bar, two nucleotide substitutions for 100 nucleotides. Bootstrap values below 50 have been removed from the dendrogram.

V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
208
From the results it was found that all purple sulfur bacteria (with
the exception of Allochromatium) could be isolated from marine habitats,
whereas most of the purple nonsulfur strains were from fresh water
habitats (Table 3.1 and 3.2). Among purple sulfur bacteria isolated, most
of strains belong to the genus Marichromatium. Among purple non sulfur
bacteria, most of the strains/species belong to the genera Rhodobacter
and Rhodopseudomonas.
Most of the strains (47%) isolated in the present study are from
coastal habitats of south India, followed by strains isolated from
Himalayan region of north India (24%). Remaining percentage of samples
include all other habitats (section, 2.9). From the present study it was
observed that Purple sulfur bacteria (mostly Genus Marichromatium) are
predominant in marine and coastal habitats and purple nonsulfur
bacteria (mostly Rhodobacter and Rhodopseudomonas) are predominant
in fresh water habitats.
As a whole, among 59 strains, it possible to describe 7 novel
species and 3 additional strains of type strains, which had been isolated
from different regions of India. Six among 7 novel species belongs to
purple nonsulfur bacteria and one is of purple sulfur bacteria.
Until now, a total number of 137 (75 PNSB + 62 PSB) purple
bacterial species were described and validly published from 13 different
countries. Among 137 novel purple bacterial species, 38 species are
from India, which contributes 27.7% to total novel purple bacterial
species of the world (Fig. 4.8).
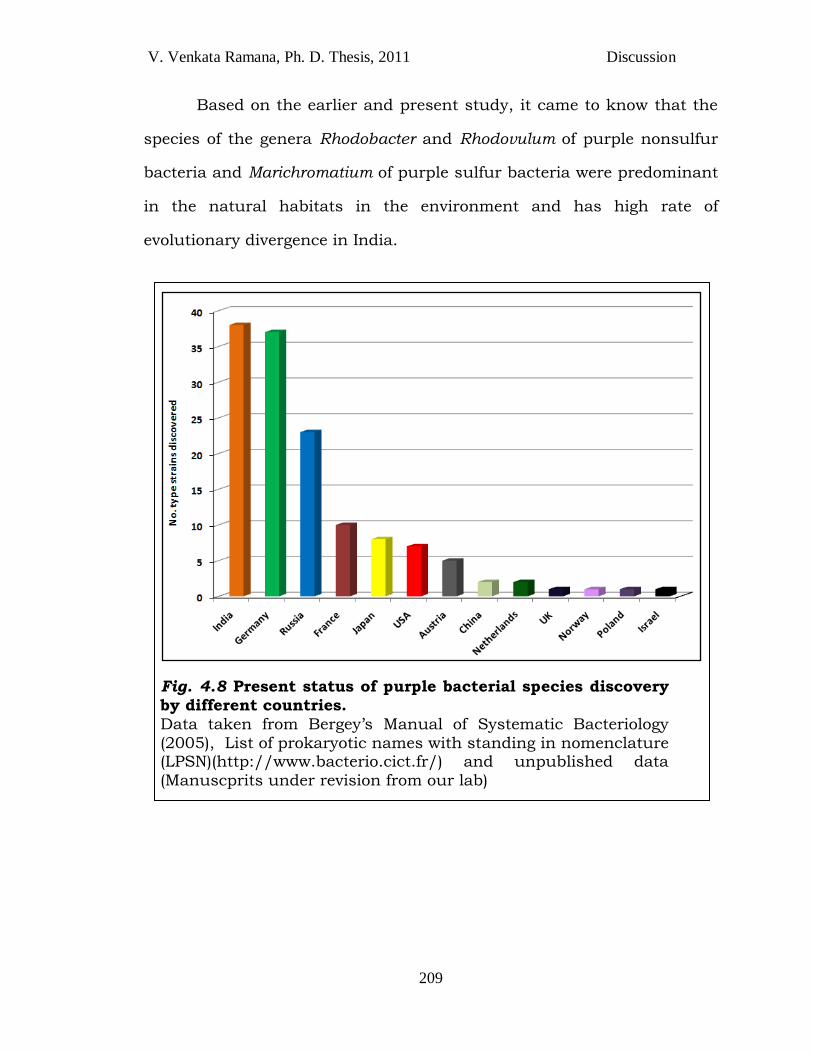
V. Venkata Ramana, Ph. D. Thesis, 2011 Discussion
209
Based on the earlier and present study, it came to know that the
species of the genera Rhodobacter and Rhodovulum of purple nonsulfur
bacteria and Marichromatium of purple sulfur bacteria were predominant
in the natural habitats in the environment and has high rate of
evolutionary divergence in India.
Fig. 4.8 Present status of purple bacterial species discovery
by different countries.
Data taken from Bergey‘s Manual of Systematic Bacteriology
(2005), List of prokaryotic names with standing in nomenclature (LPSN)(http://www.bacterio.cict.fr/) and unpublished data
(Manuscprits under revision from our lab)





























