Embed Size (px)
Citation preview

11
Chapter 2 Review of Literature

12
2.1 Plastic wastes: An aesthetic nuisance
Increasing amounts of plastic waste are being generated following the rapid
rate of urbanization in India. Today, there is a staggering demand for plastic
products with the rising affluence and public embracement of western
consumerism. However, this expansion of plastic production and consumption is
having a significant impact both visibly and invisibly on the environment and
society in India. The problems with plastic waste may seem surprising in a country
where traditional materials fulfilling the current role of plastics have existed. The
winning factor for plastics is its functional superiority (convenience) and cost
effectiveness. By sheer economies of scale, plastics have eroded the traditional
industries in India and have slowly perpetuated the throwaway culture in the
Indian society. The bottled water, fast food and Pepsi Coke culture in the country
contributes to the increasing plastic waste generation in India. The problem
becomes very visible when there is no effective end of life management to take
care of the litter, and this creates an environmental and social problem. The
widespread use of plastics as a packaging medium has resulted in the landscapes
of India being littered with non-biodegradable plastic bags and PET bottles, with
plastic bags dominating the litter. Much plastic waste has a value, and is
consequently taken care of by the informal recycling sector. Market forces guide
the informal sector, and they contribute to the waste system immensely by
collecting waste material that has a value, thereby taking over a part of the burden
on the municipalities. Despite the attempts from the formal and the informal
sector, significant quantities of the plastic waste remain uncollected. Waste
management is also constrained by the lack of public awareness and low municipal
budgets in the country. Most municipalities are starved of budgets and this impairs
the system of waste collection and disposal in many cities in India. Even when
budgets are adequate for collection, safe disposal remains a major problem. In
essence, inefficient waste management leads to a number of environmental
problems. The situation is more acute in countries such as India where economic
growth as well as urbanization is quite rapid. In view of the limited resources and
availability of land for disposal, especially in the metropolitan cities, there is a

13
need for a concerted effort to develop cost-effective and feasible policy options for
tackling the waste management problems.
Figure 2.1: Linkage between waste prevention and waste minimization
2.2 Plastic Industry in India
The growth of the Indian plastic industry has been phenomenal - the growth
rate (17%) is higher than for the plastic industry elsewhere in the world. India has a
population of over 1 billion and a plastic consumption of 4 million tonnes. One third
of the population is destitute and may not have the disposable income to consume
much in the way of plastics or other goods. The virgin industry does not target this
population to expand its markets. However, one third of the population is the middle
class whose aspirations could be molded to increase consumption. Plastic
manufacturers create needs for this segment of population. The rising needs of the
middle class, and abilities of plastics to satisfy them at a cheaper price as compared to
other materials like glass and metal, has contributed to an increase in the consumption
of plastics in the last few years. The consumption trends for key commodity plastics
are presented below in Table 2.1.
A total of 36.5 million tonnes/year (36.5 kg/individual) of municipal solid
waste is generated in the country. Considering the fact that the plastic consumption in
the country is 4 million tonnes and 52% of the plastics is used for packaging, and then
we could estimate that the plastic waste generated is at least 2 million tonnes and not
more than 4 million tonnes. Since plastics constitute only between 1-4% of the waste,
and then the total waste generated should be between 50 million tonnes – 400 million
tonnes /year that constitute 50-400 kg/individual, which is higher than the reported
figures.

14
Table 2.1: Total predicted increase in consumption of resins
Polymer 1995-1996 2001-2002 2006-2007
Polyethylene (PE) 823 1835 3267
Polypropylene (PP) 340 885 1790
Polyvinyl chloride (PVC) 489 867 1287
Polyethyleneterephthalate (PET) 34 140 289
Others 203 647 1415
Total 1889 4374 8054
Plastics in Packaging 976 2272 4037
% of Plastics in Packaging 52% 52% 50%
(Source: National Plastic Waste Management Task Force -1997). Figures in thousand tonnes
Table 2.2: Demand scenario for key commodity plastics in India
1995-1996 2001-2002 2006-2007
Total polymers 1889 4374 8054
Process Waste (2%) 38 87 161
Post consumer Waste17 870 (46%) 1966(45%) 3624(45%)
(Source: National Plastic Waste Management Task Force -1997) Figures in thousand tonnes
2.3 Plastic waste generation in India
Recycling can be done but is very tedious. The sorting of the wide variety
of discarded plastic material is also a very time-consuming process. Moreover, the
presence of a wide variety of additives such as pigments, coatings, fillers, limits
the use of the recycled material. Thus significant step towards, replacement of
non-degradable polymers by degradable polymers and make environment pollution
free is a major interest both to decision-makers and the plastic industry (Anderson

15
and Dawes 1990, Song et al., 1999). In India plastic recycling is a lucrative
business, and its sustainability is maintained by the price difference between
virgin and reprocessed granules. Recycled granules are 60% cheaper than the
virgin granules. Much of this price difference is owing to the poor wages paid in
this sector and pilferage of electricity in the informal sector. As long as there is an
ever-growing demand for cheap plastic items in the country by the poor people,
plastic recycling in this low grade manner (using poor quality plastic waste in low
grade melters and extruders most of which do not have temperature control
mechanisms) with toxic additives and colours will continue. The range of end
products produced from recycling is seemingly endless, and suffices the demand
of a population living below the poverty line, which accepts products of lower
quality. Therefore the structure of the plastic recycling industry indicates that there
are several issues of concern that need immediate attention. Some of the issues
pertain to the health and hygiene of the workers involved in the reprocessing trade,
upgrading of processing equipments used in the recycling of plastics, quality of
the effluent from the recycling plants, and finally, the quality of the products from
recycled plastics waste. Besides the regulatory relationship, the industry has also
come up with voluntary initiatives of awareness generation in the view to protect
the image of the plastic industry. The Indian Centre for Plastics and Environment
(ICPE) has been set up as a result of the recommendations made in the plastic
waste management task force. Responsibility to protect the environment and
enforcing the existing regulation lies within the Ministry of Environment and
Forests (MOEF) in India. The Central Pollution Control Board (CPCB) reporting
to MOEF is an autonomous body, with no bona fide powers to enforce laws, with
its major function to provide advice and technical assistance. In addition, every
state has a State Pollution Control Boards (SPCB), which is an autonomous body
under the State Government that enforces rules and regulations. An increasing
number of environmental legislation requires the SPCB and CPCB to work
together in implementing rules and regulations.

16
Table 2.3: State specific initiatives to address the problem of plastic waste
States Date Action Taken
Himachal Pradesh (Shimla) July 1996
A ban was placed on plastic littering. The act also provides provision for imposing deterrent penalties.
Jammu and Kashmir November 1998
Ban on the use of plastic bags. They have been replaced by bags of alternative material such as jute and paper.
Maharashtra March 1999 Ban on thin plastic bags but failure of the rule due to poor enforcement.
Mumbai 15 August 2000
State law was passed upholding national law prohibiting manufacture of polybags less than 20 microns thick and also mandated names and addresses of manufacturers to be printed on all bags. It also stated that licences would be revoked for all who disobey the rule The Mumbai Municipal Corporation has appointed a team of 97 detectors who monitor the compliance and fine the defaulters. Harsh measures like a fine of Rs 2000 (USD 42) for any shopkeepers using the bags and Rs 500 (USD 10) to the user have been imposed. In fact the anti plastic drive had led to the collection of Rs 100,000 (USD 2127) in fines.
Goa January 1998
15 August 2000 October 2000
Goa Non-Biodegradable Garbage Disposal Act Goa State Government announced a total ban on the use of recycled plastic bags less than 20 microns. Eighty day clean up drive to remove plastic bags from drains, beaches and roads of the city.
2.4 Emergence of substitutes: Bioplastics
Sustainable product development has attracted a lot of attention in the last few
years, and there has been extensive research looking at ways to provide material needs
using energy efficient, non-toxic and renewable sources rather than finite materials. It
is increasingly being realized that the use of long-lasting polymers for short-lived
applications is not entirely justified, especially when increased concern exists about
the preservation of finite resources. Conventional plastics are persistent in the
environment, if improperly disposed; they are a significant source of environmental
pollution and have a costly impact on waste management. For these reasons,

17
replacement of non-degradable polymers by degradable plastics, particularly for
single-use disposables and packaging applications, is of major interest to decision-
makers. Further, researchers have been very optimistic on the enormous potential of
replacing oil-based plastics with materials from naturally occurring polymers.
2.5 Composition of bioplastics
Bioplastics are made by the building blocks of complex carbohydrates like
cellulose or starch and reconstructing (polymerizing) them into plastics chemically,
biologically or thermally using microorganisms. The degradation of the polymer may
be caused by naturally occurring micro-organisms, the assistance of UV and heat
radiation, sunlight, hydrolysis by water and oxidation by air. There are four main
types of bioplastics available in the market today:
• Thermoplastic starch
• Cellulose acetates
• Polyhydroxy alkanoates (PHA)
• Polylactides
The three types of biodegradable plastics introduced are photodegradable,
semi-biodegradable, and completely biodegradable.
Photodegradable plastics have light sensitive groups incorporated directly into
the backbone of the polymer as additives. Extensive ultraviolet radiation (several
weeks to months) can disintegrate their polymeric structure rendering them open to
further bacterial degradation (Kalia et al., 2000a,b). However, landfills lack sunlight
and thus they remain non-degraded.
Semi-biodegradable plastics are the starch-linked plastics where starch is
incorporated to hold together short fragments of polyethylene. The idea behind starch-
linked plastics is that once discarded into landfills, bacteria in the soil will attack the
starch and release polymer fragments that can be degraded by other bacteria. Bacteria
indeed attack the starch but are turned off by the polyethylene fragments, which
thereby remain non-degradable (Johnstone, 1990).
The third type of biodegradable plastics rather new and promising because of
its actual utilization by bacteria to form a biopolymer. Included are

18
polyhydroxyalkanoates (PHA), polylactides (PLA), aliphatic polyesters,
polysaccharides, copolymers and/or blends of the above.
2.6 Status of bioplastics in India
There is a lot of awareness among the research institutions on the potential of
bioplastics in India. However; India appears to be investing only in first generation of
oil based plastics that have starch to impart it with biodegradability. Biodegradation
of polymers can be achieved in two ways. One way is to synthesize polymer like
PLA/PHB, which are biodegradable in nature. The other way is to modify the
properties of non-biodegradable plastics by the incorporation of additives that help in
breaking the molecular chain and permit direct metabolisation by microbes when
disposed in nature. In India the R&D efforts have focused more on the second
approach of biodegradation. The National Research Development Corporation of
India has developed a biodegradable plastic by mixing plastics/ LDPE and starch
made from tapioca and a soluble chemical agent that will soon bring biodegradable
bags in the market. This plastic is found to have adequate mechanical strength, and
has taken only 2 months under soil burial for complete disintegration. The starch
component, being organic, degrades in the soil and once the molecule of the
compound breaks, its vulnerability to bacterial attacks increases, thus resulting in its
disintegration (CDC, 2001). Currently there are two units in India who have opted for
the above-mentioned technology for production of biodegradable plastic. One of these
units located in North India is producing 30-40 tonnes/year of biodegradable plastics
in the granular form. Their products have been quality tested in India’s premiere
research institutions. Their focus has been mainly on the packaging sector (carry bags,
shopping bags, woven sack, and disposable containers. etc. The selling price of the
biodegradable plastic is Rs 80-100/kg and the price of a normal plastic product is Rs
60-70/kg. Further there is an excise duty of 16% on all products. Unfortunately cost
constraints have limited their markets, and thereby production (CDC, 2001). The
company claims its products are biodegradable when buried or under continued
exposure in atmosphere in 4 to 24 weeks thereby avoiding environmental pollution.
The market size of biodegradable plastics is estimated at present to be 46,000
tonnes in India and is likely to go up to 96,000 tonnes in 2006-07 based on a 15%
penetration level of potential segments (CDC, 2001). Product applications for bioplastics

19
will largely depend on its material properties like its strength, lifespan, resistance to heat
and water and the ability to package food items. Further the successful introduction of
these products also depends on their functional advantage, cost, waste management
attributes (ways of dealing with bio waste), ways of marketing and legislation.
2.7 Challenges ahead
There are number of factors that impede the adoption of bioplastics. A few of
them are mentioned below:
• All bioplastics are relatively more expensive than oil based plastics. There are
high research and development costs associated with them. The scale of
production is low, and hence the price bias exists. There are technical
uncertainties with the right choice of material for selected applications.
• The legal framework for the utilization of biodegradable materials is still very
unclear. Within waste management, local authorities have not treated
bioplastics as compostable material
• Definitions of biodegradability and compostability are unresolved. An
international standard for degradable materials is now being developed, which
is vital for the bioplastic stream to operate successfully.
• The achievement of total biodegradability.
The development of starch-based biodegradable plastics looks very promising
given the fact that starch is inexpensive, available throughout the year, and
biodegradable in various environments. The main drawbacks the industry is running
into are low water-barrier properties of bioplastics and the migration of hydrophilic
plasticizers with consequent ageing phenomena. The bioplastic industry is still in its
infancy and there are still several uncertainties that prevent the large-scale adoption of
bioplastics but continuous research in this direction may ensure that bioplastics of the
future will be produced from renewable sources and will display in-use properties
similar to those of conventional plastics. This in turn may change the scenario of
plastic waste management to a large extent. One has to remember that moving from
one technology to another based on non–toxic renewable input, will not by itself
change the development paradigm. Bioplastics and products made from renewable
sources may address the issues of toxicity and conserving finite oil resources, but

20
bioplastic is just another manufactured material. Introduction of bioplastics may well
solve the waste problems of packaging, and more so single use disposals, but it will
still encourage a more conscience free consume and throwaway culture.
Bio-plastics are a promising technology that can change the scenario of plastic
waste management. There are still several aspects that are unclear about bio-plastics,
but further research may bring to light several features that might aid the replacement of
oil-based plastics. Such a development may of course solve several waste problems
related to packaging, but would probably still propagate a throwaway culture. Plastic
waste is a pressing issue in the country today. A large number of Indians have turned
away from traditional modes of consumption, and are moving towards more wasteful
patterns of resource use. The increasing purchasing power and consumerism of the
burgeoning Indian middle class is moving India into the vicious use-and-discard cycle.
Halting these consumption patterns seems difficult in the light of globalization and
modernity. Ideally, looking at more sustainable means of satisfying need should be the
goal for the near future, but we also have to face the reality and challenges of the
existing situation. The consumption of plastic will double to 8 million tonnes in 2006,
and subsequently, the plastic waste will also escalate. Given this scenario, it is crucial
for India to check the use of plastic in the country. The existing policies have not been
able to provide any respite against littering and its associated problems. Therefore there
is an urgent need to identify policy options that can help in establishing an efficient
waste management process, and ensure efficient resource use in the country. However,
finding solutions to these problems calls for an active involvement from the
stakeholders, particularly the Government, to translate the goals into reality, by taking
the required initiatives. Such a proactive approach by the Government, along with a
clear policy agenda, and the cooperation of all the stakeholders to realize the policy
goals, will only help to ensure sustainable use of plastics in the country.
2.8 Polyhydroxyalkanoates
2.8.1 History of polyhydroxyalkanoates
The first of the discovered PHAs was polyhydroxybutyate. Now the number of
identified PHAs exceeds 100. PHAs are stored in the form of granules by bacteria. The
stained granules are distinctly visible under the microscope. The first determination of
P(3HB) had to wait until 1926 by Lemoigne. He extracted two components from Bacillus

21
megaterium, which he considered as a product of P(3HB) hydrolysis. During the following
30 years, interest in PHA was scant. It was concluded that PHB was an intracellular
reserve material. This paper marks the time when interest of microbiologists and
biochemists in PHB began to increase. The following 40 years saw intense research on the
subject and featured important developments in the knowledge about the polymer’s
widespread occurrence in various microorganisms. Until the end of 1973, interest in PHB
had been directed almost solely at its physiological significance in the functioning of
microbes and at the influence of environmental factors on its synthesis and reutilization.
The oil crisis of 1973 and the subsequent increase in the price of oil, and basic material,
cast doubts on the future of petroleum based polymer industry and set the stage for the
search for alternative types of plastic material. In 1976, Imperial Chemical Industries (ICI)
of England started investigating whether PHB could be profitably produced by bacterial
fermentation from photosynthesis-derived carbohydrate feedstocks (Senior and Dawes,
1973). Not only could PHB be synthesized from renewable sources, but some of its
properties resembled those of polypropylene. In the following years, research on PHB and
other forms of PHAs included investigations with other microorganisms and the potential
use of these biopolymers was realized (Braunegg et al., 1998; Volova et al., 2005). Since
mid-1980s, PHB and P(3HB-co-3HV) (marketed under the trademark of BIOPOL) in
agricultural and pharmaceutical industry, Zeneca Ltd (Great Britain, Bellingham) was
established. In 1996, BIOPOL business was acquired by Monsanto. Recently, PHA
production has become the goal of many large companies in the USA, Germany, Italy,
Japan, Scandinavian countries. The cost of the first lots of Biopol was US $16/Kg as
compared to the cost of polyolefins on the world market US $1/Kg. So BIOPOL as a
packaging material is not economically competitive with them. PHAs of such high cost
can be reasonably used for specific purposes, e.g. materials for medical applications. Thus,
to produce and apply PHAs on a large scale, it is necessary first to reduce their cost. Many
firms and industrial companies such as Monsanto, Metabolix Inc., Tepha and Procter and
Gamble, deal with the commercialization of PHAs.
The most widely produced microbial bioplastics are polyhydroxyalkanoates
(PHAs) and their derivatives (Madison and Huisman, 1999). Poly(D-3
hydroxybutyrate) is the most ubiquitous and most intensively studied PHA. PHAs are
the only 100% biodegradable polymers. They are polyesters of various HAs, which
are synthesized by numerous microorganisms under unbalanced growth conditions,

22
such as limitation of an essential nutrient such as nitrogen, oxygen magnesium and
phosphorus and presence of excess carbon source. They possess properties similar to
various synthetic thermoplastics like polypropylene and hence can be used in their
place. They are also completely degraded to water and carbon dioxide under aerobic
conditions and to methane under anaerobic conditions by microorganisms in soil, sea,
lake water and sewage. PHAs are of commercial interest because chemical or
biological hydrolysis of PHAs can yield optically pure (R)-form hydroxycarboxylic
acids (Lee et al., 2000; Park et al., 2002), which are used in the manufacture of
antibiotics, vitamins, perfumes and pheromones. To date, over 150 types of
hydroxycarboxylic acid have been identified as components of PHAs (Steinbuchel
and Valentin, 1995).
2.8.2 Location of polyhydroxyalkanoates in the cell
In the prokaryotes, PHA occurs either as inclusion bodies or as complexes of
Ca2+ and polyphosphates in the cytoplasmic membranes. It is accumulated as a
membrane enclosed inclusion in many bacteria up to 80% of the dry cell weight.
Discrete granules of PHA generally occur in the cytoplasm as inclusion bodies of
irregular morphology with diameter of about 0.2 to 0.5µm. These granules appear as
refractile inclusion under electron and phase contrast microscope. Light microscopic
investigation of the cells, stained with Sudan black B or more specific with
epiflourescence microscope using Nile Blue A provides easy means for detecting
PHA in cells.
2.8.3 Structure of polyhydroxyalkanoates
2.8.3.1 Chemical structure
Polyhydroxyalkanoates are polyester of various hydroxyalkanoates,
hydroxylated at positions 3, 4, 5 and 6, all of which are (R)-form chiral molecules that
are synthesized by many gram-positive and gram-negative bacteria from at least 75
different genera. General structural formula is shown in Figure 2.2 to 2.4 (Park et al.,
2005). PHB has a perfectly isotactic structure with only the (R) - configuration.
P(3HB) isolated from bacteria possesses 55 to 80% crystallinity. Polyhydroxybutyrate
(PHB) was the first PHA to be discovered in Bacillus species by Lemoigne in 1926
and is also the most widely studied and best characterized PHA. The molecular

23
weight of these compounds range from 2×105 to 3×106 Da. Depending on the number
of carbon atoms in the monomer unit, PHAs can be divided into three groups:
(1) Short chain length PHAs (scl- PHAs), which consist of 3-5 carbon atoms.
(2) Medium chain length PHAs (mcl- PHAs), which consist of 6-14 carbon atoms.
(3) Long chain length PHAs (lcl- PHAs), which consist of 17 and 18 carbon atoms.
This division of polymers into groups is based on the substrate specificity of
PHA synthases that can only accept certain hydroxyalkanoic acids in course of
polymerization (Anderson and Dawes, 1990). The PHA synthase of A. eutrophus can
polymerize 3HAs consisting of 3 to 5 carbon atoms whereas that present in
Pseudomonas oleovorans can only accept 3HAs of 6 to14 carbon atoms.
n varies from 600 to 35000 R= hydrogen Poly(3-hydroxypropionate) R=methyl Poly(3-Hydroxybutyrate) R=ethyl Poly(3-hydroxyvalerate) R=propyl Poly(3-hydroxyhexanoate) R=pentyl Poly(3-hydroxyoctanoate) R=nonyl Poly(3-hydroxydodecanoate)
Figure 2.2: Structure of polyhydroxyalkanoates
Figure 2.3: Structure of co polymer 3-HB-3HV
Figure 2.4: Structure of co polymer 3-HB-4HB

24
2.8.3.2 Physical properties
PHB is similar to polypropylene with three unique features: thermoplastic
processability, 100% resistance to water, and 100% biodegradability (Hrabak, 1992).
PHB is an aliphatic homopolymer with a melting point of 179°C and highly
crystalline (3HB) molecules within bacteria are amorphous (Barnard and Sanders,
1989; Amor, et al., 1991; Kawaguchi and Doi, 1992) and exist as water insoluble
inclusions. Water is a minor component of PHA inclusions and therefore it was
suggested that water could act as plasticizer (Barnard and Sanders, 1989). About 5 to
10% of water was estimated to be present in the nascent PHB inclusions, which upon
removal allows for the polymer chains to rearrange into lamellar crystals.
The densities of crystalline and amorphous PHB are 1.26 and 1.18 g/cm3,
respectively. The Mw of P(3HB) produced from wild-type bacteria is usually in the
range of 1 x 103 to 3 x 106 g/mol. The family of PHAs exhibits a wide variety of
mechanical properties from hard crystalline to elastic, depending on composition of
monomer units which broadens its application area, for example, MCL-PHAs are
semi-crystalline elastomers with low melting point, low tensile strength and units to
form PHA copolymers can also improve other properties such as crystallinity, melting
high elongation to break and can be used as biodegradable rubber after cross linking.
The glass transition temperature of PHB is around 4oC while the melting temperature
is near 180oC. Mechanical properties like Young’s modulus (3.5 Gpa) and the tensile
strength (43Mpa) of P(3HB) material are close to those of isotactic polypropylene.
Bacterially produced polyhydroxybutyrate and other PHAs have sufficiently high
molecular mass to have polymer characteristics that are similar to conventional
plastics such as polypropylene (Madison and Huisman, 1999).
2.9 Factors affecting polyhydroxyalkanoate synthesis and its
composition
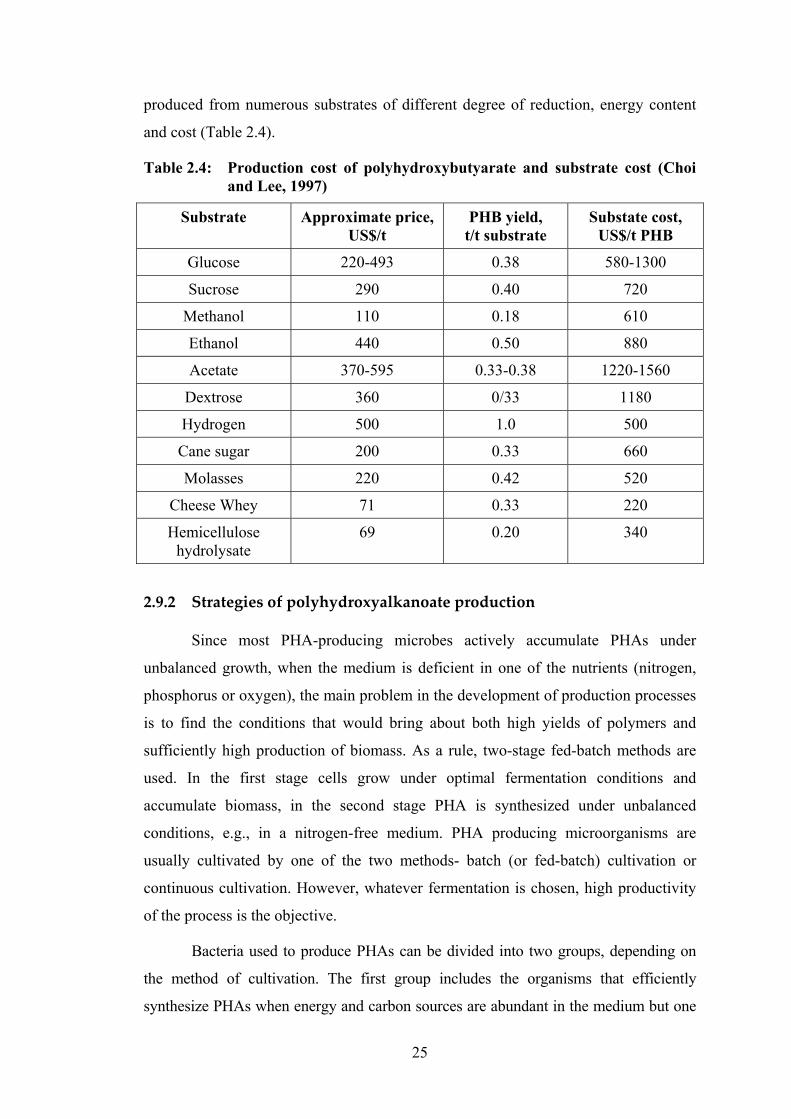
2.9.1 Feed substrate and growth conditions
The choice of the substrate used to produce PHAs is determined by
physiological-biochemical properties of PHA-producing microorganisms, economic
efficiency of the preferred strategy and the field of application of the ready produce.
The quality and cost of the substrates for different applications can vary. PHAs can be
25
produced from numerous substrates of different degree of reduction, energy content
and cost (Table 2.4).
Table 2.4: Production cost of polyhydroxybutyarate and substrate cost (Choi and Lee, 1997)
Substrate Approximate price, US$/t
PHB yield, t/t substrate
Substate cost, US$/t PHB
Glucose 220-493 0.38 580-1300
Sucrose 290 0.40 720
Methanol 110 0.18 610
Ethanol 440 0.50 880
Acetate 370-595 0.33-0.38 1220-1560
Dextrose 360 0/33 1180
Hydrogen 500 1.0 500
Cane sugar 200 0.33 660
Molasses 220 0.42 520
Cheese Whey 71 0.33 220
Hemicellulose hydrolysate
69 0.20 340
2.9.2 Strategies of polyhydroxyalkanoate production
Since most PHA-producing microbes actively accumulate PHAs under
unbalanced growth, when the medium is deficient in one of the nutrients (nitrogen,
phosphorus or oxygen), the main problem in the development of production processes
is to find the conditions that would bring about both high yields of polymers and
sufficiently high production of biomass. As a rule, two-stage fed-batch methods are
used. In the first stage cells grow under optimal fermentation conditions and
accumulate biomass, in the second stage PHA is synthesized under unbalanced
conditions, e.g., in a nitrogen-free medium. PHA producing microorganisms are
usually cultivated by one of the two methods- batch (or fed-batch) cultivation or
continuous cultivation. However, whatever fermentation is chosen, high productivity
of the process is the objective.
Bacteria used to produce PHAs can be divided into two groups, depending on
the method of cultivation. The first group includes the organisms that efficiently
synthesize PHAs when energy and carbon sources are abundant in the medium but one

26
of the biogenic elements (Nitrogen, Phosphorus, Sulfur, Potassium, Magnesium or
Oxygen) is deficient. These are A. eutrophus, Azotobacter vinelandii, P. oleovorans,
etc. The second group unites microorganisms that can efficiently synthesize PHAs at
high rates under optimal growth conditions. These are A. latus and recombinant
organisms harbouring the biosynthetic operon of A. eutrophus.
2.9.3 Bacterial strain
PHAs are produced by many different bacterial cultures. Cupriavidus necator
(formerly known as R. eutropha or A. eutrophus) is the one that has been most
extensively studied. Imperial Chemical Industries (ICI) were the first to use this
bacterial strain for the production of PHBV copolymer under the trade name Biopol.
Recently, Metabolix Inc. (USA) acquired the Biopol patents. At present, bacterial
fermentation of C. necator seems to be the most cost-effective process and even if
production switches to other bacteria or agricultural crops, these processes are likely
to use C. necator genes. A few important other strains that were recently studied
include: Bacillus sp., Alcaligenes sp., Pseudomonas sp., Aeromonas hydrophila,
Rhodopseudomonas palustris, Escherichia coli, Burkholderia sacchari and
Halomonas boliviensis.
PHA has been industrially produced by pure cultures including A. latus,
A. vinelandii, P. oleovorans, recombinant A. eutrophus and recombinant
E. coli (Grothe et al., 1999; Lee and Choi, 1999). With current advances in PHA
research, a PHA concentration of more than 80 g/l and productivity of more than 2 g/l/h
have been obtained in the laboratory using fed-batch cultivation (Lee, 1996b).
Similarly, recovery methods for PHAs of various purities from microorganisms have
received attention. Wider use of PHAs is prevented mainly by their high production
cost compared with the oil-derived plastics (Byrom, 1987; Lee and Yu, 1997). With the
aim of commercializing PHA, a substantial effort has been devoted to reducing the
production cost through the development of bacterial strains and more efficient
fermentation/recovery processes (Lee, 1996b; Grothe et al., 1999). From the literature,
the major cost in the PHA production is the cost of the substrate (Yamane, 1993).
The yields of PHA from the various substrates are similar, with one exception.
Consequently, the price of substrate has the largest influence on the cost of production
of PHA. The cheapest substrate costs $0.22/ kg of PHA compared with the cost of

27
polypropylene of $0.185/ kg. Productivity also has an effect on the production costs.
However, this is relative to the substrate, and downstream processing apparently has a
weak effect on the final cost. When the PHB productivity increased from 1.98 to 3.2
g/l/h, the PHB production cost decreased from $5.37/kg PHB to $4.91/ kg PHB (Lee
and Choi, 1999). In a laboratory fed-batch system using A. latus, the highest reported
productivity was 4.94 g/l/h which would lead to production costs of $2.6/ kg PHB. PHA
content of the produced biomass strongly affects the efficiency of the recovery process.
For example, a relatively low PHB content of 50% results in a high recovery cost of
$4.8 /kg PHB. On the other hand, the recovery cost for a process with 88% PHB
content was only $0.92/ kg PHB (Lee and Choi, 1999). A lower PHB content clearly
results in a high recovery cost. This is mainly due to the use of large amounts of
digesting agents for breaking the cell walls and to the increased cost of waste disposal.
Table 2.5: Overview of bacterial strains used to produce polyhydroxyalkanoates
Bacterial strain Carbon source (s) Polymer (s) produced Reference
Aeromonas hydrophila
Lauric acid, oleic acid mcl-PHAs Lee et al. 2000; Han et
al. 2004
Alcaligenes latus Malt, soy waste, milk waste, vinegar waste, sesame oil
PHB Wong et al. 2004, 2005
Bacillus cereus
Glucose, e-caprolactone, sugarbeet molasses
PHB, terpolymer
Labuzek and Radecka 2001; Yilmaz and Beyatli 2005; Valappil et al. 2007
Bacillus sp.
Nutrient broth, glucose, alkanoates, e-caprolactone, soy molasses
PHB, PHBV, copolymers
Katircioglu et al. 2003; Shamala et al. 2003; Tajima et al. 2003; Yilmaz et al. 2005; Full et al. 2006
Burkholderia cepacia
Palm olein, palm stearin, crude palm oil, palm kernel oil, oleic acid, xylose, levulinic acid, sugarbeet molasses
PHB, PHBV
Keenan et al. 2004; Nakas et al. 2004; Alias and Tan 2005; Elik et al. 2005
Caulobacter crescentus
Caulobacter medium, glucose PHB Qi and Rehm 2001

28
Bacterial strain Carbon source (s) Polymer (s) produced Reference
Escherichia coli mutants
Glucose, glycerol, palm oil, ethanol, sucrose, molasses
(UHMW)PHB
Mahishi et al. 2003; Kahar et al. 2005; Park et al. 2005; Nikel et al. 2006a; Sujatha and Shenbagarathai 2006
Halomonas boliviensis
Starch hydolysate, maltose, maltotetraose and maltohexaose
PHB Quillaguaman et al. 2005, 2006
Legionella pneumophila Nutrient broth PHB James et al. 1999
Methylocystis sp. Methane PHB Wendlandt et al. 2005
Microlunatus phosphovorus Glucose, acetate PHB Akar et al. 2006
Pseudomonas aeruginosa
Glucose, technical oleic acid, waste free fatty acids, waste free frying oil
mcl-PHAs Hoffmann and Rehm 2004
P. oleovorans Octanoic acid mcl-PHAs Durner et al. 2000; Foster et al. 2005
P. putida Glucose, octanoic acid, undecenoic acid
mcl-PHAs Tobin and O’Connor 2005; Hartmann et al. 2006
P. putida P. fluorescens, P. jessenii
Glucose, aromatic monomers
aromatic polymers
Tobin and O’Connor 2005; Ward and O’Connor 2005.
P. stutzeri Glucose, soybean oil, alcohols, alkanoates
mcl-PHAs Xu et al. 2005
Rhizobium meliloti, R. viciae, Bradyrhizobium japonicum
Glucose, sucrose, galactose, mannitol, trehalose, xylose, raffinose, maltose, dextrose, lactose, pyruvate, sugar beet molasses, whey
PHB Mercan and Beyatli 2005

29
Bacterial strain Carbon source (s) Polymer (s) produced Reference
Rhodopseudomonas palustris
Acetate, malate, fumarate, succinate, propionate, malonate, gluconate, butyrate, glycerol, citrate
PHB, PHBV Mukhopadhyay et al. 2005
Spirulina platensis (cyanobacterium) Carbon dioxide PHB Jau et al. 2005
Staphylococcus epidermidis
Malt, soy waste, milk waste, vinegar waste, sesame oil
PHB Wong et al. 2004, 2005
Cupriavidus necator
Glucose, sucrose, fructose, valerate, octanoate, lactic acid, soybean oil
PHB, copolymers
Kichise et al. 1999; Taguchi et al. 2003; Kahar et al. 2004; Khanna and Srivastava 2005a; Kim et al. 2005; Volova and Kalacheva 2005; Volova et al. 2005
C. necator H16 Hydrogen, carbon dioxide PHB Pohlmann et al. 2006
mcl-PHAs: medium-chain-length polyhydroxyalkanoates, PHB: poly(3-hydroxybutyrate), PHBV: poly(3-hydroxybutyrate-co-valerate), UHMW: ultra high molecular weight
2.9.4 Substrate
One of the problems preventing the commercial application of P(3HB) is its
high production cost. From an economical point of view, the cost of substrate (mainly
carbon source) contributes most significantly to the overall production cost of P(3HB)
(Choi and Lee 1997; Lee and Choi, 1999). To reduce the substrate cost, recombinant
strains utilizing a cheap carbon source and corresponding fermentation strategies have
been developed (Lee, 1996a). There have been several reports on the production of
P(3HB) from cheap carbon sources by wild-type P(3HB) producers (Kim and Chang,
1995). However, the P(3HB) concentration and P(3HB) content obtained were
considerably lower than those obtained using purified carbon substrates. Therefore,
more efficient fermentation strategies should be developed for efficient production of

30
P(3HB) from a cheap carbon source. Several bacterial strains can produce P(3HB)
from waste products (Steinbüchel and Valentin, 1995; Cho et al., 1997; Song et al.,
1999). If waste product:stream can be used as a substrate for the production of
P(3HB), combined advantages of reducing disposal cost and production of value-
added products can be realized. However, so far, only low P(3HB) content and
productivity were achieved from waste products. Since PHA production from glucose
or sucrose already had been optimized, development of fermentation technology to
use cheaper carbon sources would be a key factor in reducing the PHA production
cost.
The production of PHA by different microorganisms under different growth
conditions is summarized in Table 2.6.
Table 2.6: Polyhydroxybutyrate production from biowastes by diverse microorganisms PHB
Microorganism Substrate Culture mode
Time (h) Yield
(%) Conc. (g/l)
Reference
Archaea Extruded rice bran 55.6 77.8
Haloferax mediterranei Extruded
corn starch
Fed batch 120 38.7 24.2
Huang et al., 2006
Actinobacteria
Rhodococcus ruber
Low- rank coal liquefaction products
Batch 120 6 0.118
Füchtenbusch and Steinbüchel, 1991
Firmicutes
Bacillus megaterium
Date syrup/ Beet molasses
Batch 48 52 1.76 Omar et al., 2001
Staphylococcus epidermidis Malt wastes Batch 48 0.121 6.93 Wong et al.,
2000 α-Proteobacteria Methylobacterium sp. ZP24 Whey Batch 48 59.6 5.9 Yellore and
Desai, 1998
M. extorquens Methanol Fed batch 186 40 114 Bourque et al., 1995
M. rhodesianum Glycerol and Casein hydrolysates
Batch 45 50 11 Bormann and Roth, 1999
Sinorhizobium meliloti
Cheese Whey permeate
Batch 96 35 169 Povolo and Casella, 2003

31
PHB Microorganism Substrate Culture
mode Time (h) Yield
(%) Conc. (g/l)
Reference
β-Proteobacteria Alcaligenes eutrophaa
Starchy waste water Batch 48 34.1 1.2 Yu, 2001
A. eutropha DSM545
Potato processing wastes
Batch 120 77 5 Rusendi and Sheppard, 1995
A. eutropha strain H16 Plant oils Batch 72 79-82 2.9-3.3 Fukui and
Doi, 1998 A. eutropha NCIMP11599
Pulp fiber sludge Batch 30 78 2.5 Zhang et al.,
2004 Malt waste 32.4 18.4 A. latus
DSM1124 Soya waste Fed batch 69
22.7 6.0 Yu et al., 1999
Burkholderia cepacia
Palm oil Mill effluent Fed batch 672 57.4 2.46 Alias and
Tan, 2005 Comamonas testosterone
Vegetable oils Batch 48 53-58 41-50 Thakor et al.,
2005
Hydrogenophaga pseudoflava
Cheese Whey permeate
Batch 96 44 165 Povolo and Casella, 2003
Ralstonia eutrophaa
Topioca hydrolysate Fed batch 59 58 61 Kim and
Chang, 1995
R. eutropha DSM11348
Glycerol and Casein hydrolysates
Batch 67 47 15 Bormann and Roth, 1999
R. eutropha Food scraps Batch 80 72.6 11.3 Du and Yu, 2002
R. eutropha H16 Soybean oil Batch 96 76 0.76 Kahar et al., 2004
γ-Proteobacteria
Azotobacter vinelandiiUWD
Beet molasses fractions
Fed batch 24 70 7.8 Page, 1992
A. vinelandiiUWD Molasses Fed batch 36 66 22
Page and Cornish, 1993
Azotobacter vinelandiiUWD
Swine waste liquor Batch 18 58.3 9.4 Cho et al.,
1997 Fed batch 70 46 25 Kim, 2000 A. chroococcum
Starch Batch 58 73.9 0.864 Kim, 2000
Pseudomonas oleovorans
Low- rank coal liquefaction products
Batch 120 8 0.363 Steinbüchel and Hein, 2001
P. putida PGA1 Saponified Palm Kernel Oil
Batch 48 19-37 0.5-1.1 Tan et al., 1997

32
PHB
Microorganism Substrate Culture mode
Time (h) Yield
(%) Conc. (g/l)
Reference
49 80 69 Wong and Lee, 1998
36.5 87 168 Ahn et al., 2001
Fed batch
26 70 35.7 Park et al., 2002
Fed batch with O2 limitation
52 80 25
Recombinant Escherichia coli Whey
Fed batch without O2 limitation
35 57 32
Kim, 2000
Recombinant E. coli
Agro-industrial byproducts
Fed batch 24 72.9 51.1 Nikel et al., 2006b
Recombinant Klebsiella aerogenes
Molasses Fed batch 32 65 24 Zhang et al., 1994
Others Raw rice grain- based distillery spent wash
Batch 96 40 2.7 Khardenavis et al., 2007
Malt and Soya waste Batch 2 70 22.4 Wang et al.,
2007
Activated Sludge
Mixed liquor Batch 30 62 1.5 Satoh et al., 1998
(a: Renamed as Cupriavidus necator)
2.10 Polyhydroxyalkanoate biosynthesis pathways
2.10.1 R. eutropha polyhydroxyalkanoate biosynthetic pathway
Most of the organisms synthesize PHA using this pathway. The biosynthesis
pathways of R. eutropha, Zoogloea ramigera and A. beijerinckii are well established
(Doi et al., 1990). Firstly, a substrate is condensed to acetyl-coenzyme A (acetyl-
CoA). Two moles of acetyl-CoA are then used to synthesize one mole of PHB.
Acetyl-CoA is subjected to a sequence of three enzymatic reactions for PHB
synthesis.
When propionic acid is used as a sole substrate, PHB-PHV copolymer is
formed. Acetyl-CoA is formed by the elimination of carbonyl carbon from propionyl-

33
CoA. Two moles of acetyl-CoA are used to form a HB unit of the copolymer, while a
HV unit is formed by the reaction of acetyl CoA and propionyl-CoA. Figure 2.5 shows
the biosynthesis pathway of PHB-PHV copolymer by R. eutropha (Doi et al., 1990).
According to Doi et al., 1990, the degradation of PHA by R. eutropha can
occur simultaneously with its biosynthesis under nitrogen limitation. This observation
is called “a cyclic nature of PHA metabolism”. The author reported that the
composition of polymer was changed from PHB homopolymer to PHB-49%PHV
copolymer when the substrate was changed from butyric acid to pentanoic acid after
96 hours of nitrogen limitation accumulation period, i.e., there was a replacement of
PHB by PHB-PHV. Likewise, when R. eutropha with a PHV fraction of 56% of its
PHA content was fed with butyric acid as a sole substrate under nitrogen limitation,
the PHA composition changed markedly, i.e., the fraction of PHV decreased from
56% to 19% after 48 hours. These findings show the simultaneous synthesis and
degradation of PHA, i.e., the cyclic nature of PHA metabolism (Doi et al., 1990).
2.10.2 Rhodospirillum rubrum polyhydroxyalkanoate biosynthetic pathway
This pathway is similar to the R. eutropha pathway but two enoyl-CoA
hydratases are also involved in the second step of catalyzing the conversion of L-3-
hydroxybutyryl-CoA to D-3- hydroxybutyryl-CoA via crotonyl-CoA (Anderson and
Dawes, 1990; Doi, et al., 1990; Lee, 1996a). A simple schematic of this pathway is
shown as:
Acetate acetyl CoA aceto-acetyl-CoA L - 3- hydroxybutyryl-CoA
crotonyl CoA D-3-hydroxybutyryl-CoA PHB.
2.10.3 Pseudomonas oleovorans polyhydroxyalkanoate biosynthetic pathway
This biosynthesis pathway is found in P. oleovorans and most pseudomonads
from the rRNA homology group I (Lee, 1996a). These organisms produce medium-
chain-length (MCL) PHAs (from C6-C9) from MCL-alkanes, alcohols, or alkanoates.
According to Doi et al., 1990, productions of short-chain-length (SCL) PHAs, i.e.,
PHB homopolymer and PHB-PHV copolymer, cab be produced by these organisms
but the productions were less than 1.5%. This PHA biosynthesis involves the cyclic-
β-oxidation and thiolytic cleavage of fatty acids, i.e., 3- hydroxyacyl-CoA, and
intermediates of the β-oxidation pathways, are used for PHA biosynthesis.

34
2.10.4 Pseudomonas aeruginosa polyhydroxyalkanoate biosynthetic pathway
Most pseudomonads from the rRNA homology group I except P. oleororans
also produce MCL PHAs using this pathway. The pathway used in these organisms is
called the P. aeruginosa PHA biosynthetic pathway. Steinbuchel, 1991 stated that
MCL-PHAs produced by this pathway are from unrelated substrates, e.g., gluconate
or acetate. PHA is synthesized from acetyl-CoA via fatty acid synthetic pathways.
2.11 Genes and enzymes involved in polyhydroxyalkanoate synthesis
The organization of PHB metabolic genes in R. eutropha is shown in
Figure 2.5. The phaCBA cluster encodes three proteins: PhaA (β-ketothiolase), which
catalyses the synthesis of acetoacetyl–CoA from acetyl– CoA; PhaB (NADPH-
oxidoreductase), which stereospecifically reduces acetoacetyl–CoA to (R)-3-
hydroxybutyryl– CoA; and PhaC (PHB polymerase), which promotes the
incorporation of (R)-3-hydroxybutyryl– CoA enantiomers in the growing polymer
(Figure 2.5). PhaC is very active towards monomers containing less than five carbon
atoms, although it also synthesizes polymers containing small quantities of higher-
length monomers (C6–C8) (Sudesh et al., 2000a; Zinn et al., 2001; Salehizadeh et al.,
2004). The regulation of the PHB pathway seems to be complex. An excess of acetyl–
CoA reduces the synthesis of PHBs, whereas all the metabolic or environmental
conditions that cause a reduction in the pool of acetyl–CoA start, or restore, PHB
synthesis (Steinbu¨chel and Schegel, 1991; Zinn et al., 2001). Furthermore, B.
megaterium PhaC is synthesised as an inactive protein that requires a different
polypeptide (PhaR) to be converted into a functional enzyme (McCool and Cannon,
2001), suggesting that PHB regulation involves different environmental, metabolic
and genetic signals (Madison and Huisman, 1999, Luengo et al., 2003). Two
additional proteins, PhaZ and PhaP (phaZ and phaP gene products, Figure 2.5), also
participate either in the catabolism (PhaZ) or in the stabilization (PhaP) of the PHB
granule. PhaZ is a depolymerase (structurally related to esterases) that catalyses the
release of (R)-3- hydroxybutyrate from the polymer (or from oligomers longer than
dimers) (Figure 2.6) (Saegusa et al., 2001, Jendrossek and Handrick, 2002). In the
absence of PHB, PhaZ is produced as an inactive protein that requires PHB and an
activator (which could be replaced by trypsin) to be transformed into an active

35
enzyme. These observations suggest that either PhaZ is synthesized as a proenzyme,
or the attack of PhaZ to the granule surface requires the participation of a proteolytic
enzyme (Zinn et al., 2001). Recent studies have shown that the degradation of PHBs
is a complex mechanism that requires several depolymerases (PhaZ1, PhaZ2 and
PhaZ3) together with other as yet uncharacterized enzymes (Saegusa et al., 2001).
PhaP (phasin) is a low-molecular-weight protein (accumulated to high levels during
PHB synthesis) that enhances PHB production by binding to the granules (it regulates
the size, number and surface to volume ratio of PHB inclusions) (Madison and
Huisman, 1999; Sudesh et al., 2000b; Zinn et al., 2001). Recently, it has been reported
that the synthesis and accumulation of PhaP is a PHB-dependent mechanism
involving the participation of PhaR (an autoregulated repressor) (York et al., 2001).
However, regulation of the size and number of PHB inclusions is not only modulated
by PhaP but also by the quantity of PhaC present in the cells. PHAs are polyesters
containing monomers of medium chain length (mclPHAs, C5–C14) or long-chain
length (lclPHAs, >C14). Although PHAs are structurally related to PHBs (short-chain
length, sclPHAs) (Lageveen et al., 1998), the microbes that synthesize PHBs usually
fail to make PHAs. However, recombinant organisms containing mixed catabolic
pathways are able to synthesize either polymers (or co-polymers) containing scl, mcl
monomers, or both (Lee, 1996a; Matsusaki et al., 1998; Sudesh et al., 2000b; Zinn et
al., 2001). The organization of the mclPHA biosynthetic genes in P. oleovorans and
in P. putida is shown in Figure 2.6. The phaC1ZC2D operon encodes two
polymerases (PhaC1 and PhaC2), a depolymerase (PhaZ) and the PhaD protein
(Huisman et al., 1991; Steinbu¨chel and Hein, 2001). The two polymerases, which are
members of the a/b hydrolase subfamily, catalyze the condensation into PHAs of
several (R)-3- hydroxy-acyl–CoA derivatives (saturated, unsaturated, linear, cyclic,
branched or substituted with different functions such as halogen atoms, hydroxy,
cyano, carboxy, or phenyl groups) whose side chains range between C5 and C14
atoms (Lageveen et al., 1998; Sudesh et al., 2000b; Steinbu¨chel and Hein, 2001).
Both enzymes are quite similar in their amino acid sequence (about 50%) and
substrate specificity (3-OH-acyl–CoA derivatives and 4-, 5- or 6-OH-acyl-CoAs)
(Madison and Huisman, 1999), although when expressed in foreign hosts, they are
also able to polymerize other monomers (Lee, 1996b; Madison and Huisman, 1999;
Antonio et al., 2000; Sudesh et al., 2000a; Steinbu¨chel and Hein, 2001).
Identification of the amino acid residues required for catalysis (Rehm et al., 2001) has

36
allowed modification of the catalytic rates in several PHA syntheses (Taguchi et al.,
2002). Most mclPHA intermediates are obtained through fatty acid b-oxidation,
although other monomers (synthesized from different carbon sources) can also be
obtained via different pathways (Figure 2.6). The use of one or the other seems to be a
strain-specific trait. A different type of polymerase (type III) exists in Chromatium
vinosum, Thiocystis violacea, Thiocapsa pfennigii and Synechocystis sp. PCC 6803
(Sudesh et al., 2000b). This is formed by PhaC and PhaE, which mainly synthesise
sclPHAs but which also polymerise scl and mcl monomers (Sudesh et al., 2000b;
Steinbu¨chel and Hein, 2001). The existence of two polymerases in the same
microorganism (probably as a consequence of gene duplication) represents an
interesting evolutionary event that could have contributed to the biochemical
transition from PHBs (which only require a single polymerase) to PHAs (where two
enzymes are involved). Further studies are needed to confirm this hypothesis. phaC1
and phaC2 are separated by a third gene that encodes the PhaZ product (Figure 2.6).
This protein, containing a conventional lipase box (Madison and Huisman, 1999;
Sudesh et al., 2000a; Zinn et al., 2001; Williams and Martin, 2002), shows a certain
homology with depolymerases (enzymes involved in the mobilization of sclPHAs)
and with many hydrolytic enzymes, suggesting that it participates in the release of
hydroxyacyl–CoA derivatives from PHAs. The topological localization of PhaZ
(granule surface) (Sudesh et al., 2000b; Zinn et al., 2001) and the inability of certain
bacteria to mobilize mclPHAs when phaZ is mutated strongly supports its
physiological function (Madison and Huisman, 1999). Expression of the phaC1ZC2D
cluster in P. putida U must be under the control of promoter sequences located
upstream of phaC; otherwise, it would not be possible to account for the drastic
reduction in PHA synthesis that occurs when phaZ is disrupted, or when a plasmid is
introduced between the duplicated copies of phaC1 (Figure 2.6) (Zinn et al., 2001).
An additional cluster (phaFI), also involved in the biosynthesis of PHAs, is located
downstream from the phaC1ZC2D operon. These genes encode phasins PhaF and
PhaI. PhaF, a histone-H1-like protein, plays a dual function: it is involved in the
stabilization of the granule, and it acts as a regulator (Prieto et al., 1999). PhaF is a
granule-associated protein that represses phaC1ZC2D and phaI and contributes to the
stabilization of PHA granules; whereas PhaI, another granule-associated protein, only
participates in the formation and stabilization of PHA inclusions (Prieto et al., 1999).
In the absence of PHAs precursor, phaC1ZC2D and phaI are not expressed, whereas

37
when mcl monomers are synthesized, PhaF is removed from the DNA, initiating (or
restoring) PHAs production. Under these new conditions, PhaF and PhaI interact with
the hydrophobic nascent polymeric chains, contributing (in an isolated fashion or as a
complex) to granule formation (Prieto et al., 1999). The physiological role of PhaD
remains obscure. It is not a granule-associated protein, although it seems to be
required for PHA formation (Klinke et al., 2000). Very recently, we have observed
that the deletion of phaDFI in the overproducing mutant P. putida U DfadBA causes a
considerable reduction (>70%) in the synthesis of aliphatic PHAs, whereas poly(3-
hydroxyphenylalkanoates) are not produced. Furthermore, the restoration of PHA
synthesis in this double-deleted mutant (DfadBAD phaDFI) requires the expression of
phaF, whereas in its absence, even when phaD and phaI are expressed, this effect was
not reversed.
Figure 2.5: Organization of the genes and enzymes involved in the biosynthesis of bioplastics. (a) Biosynthesis of PHBs in Ralstonia eutropha (formerly Alcaligenes eutrophus) (b) PHAs in Pseudomonas oleovorans (c, d) PHAs in different mutants of Pseudomonas putida U designed to prove the existence of promoters downstream from phaC1 (c) Pseudomonas putida U mutant disrupted by the insertion of the integrative plasmid pK18::mob into the depolymerase gene (d) Pseudomonas putida U mutant in which the phaC1 gene has been duplicated and a new cluster phaC1ZC2DFI, without the promoter region (P1) located upstream from phaC1, has been generated. (Luengo et al., 2003)

38
Figure 2.6: Structural organization of a polyhydroxyalkanoates granule and metabolic interconnections between the different pathways involved in the biosynthesis and catabolism of polyhydroxybutyrates and polyhydroxyalkanoates. (a) Alkane oxidation pathway. (1) Alkane 1-monooxygenase, (2) alcohol dehydrogenase, (3) aldehyde dehydrogenase. (b) Fatty-acid b-oxidation. (4) acyl–CoA ligase, (5) acyl–CoA dehydrogenase, (6) enoyl–CoA hydratase, (7) 3-hydroxyacyl–CoA dehydrogenase, (8) 3-ketothiolase, (9) (R)-enoyl–CoA hydratase, (10) 3-ketoacyl–CoA reductase. (c) Biosynthesis from carbohydrates. (11) b-ketothiolase, (12) NADPH-dependent acetoacetyl–CoA reductase. (d) De novo fatty acid synthesis. (13) acetyl–CoA carboxylase, (14) ACP-malonyltransferase (15) 3-ketoacyl-ACP synthase, (16) 3- etoacyl-ACP reductase, (17) 3-hydroxyacyl-ACP reductase, (18) enoyl-ACP reductase, (19) 3-hydroxyacyl-ACP–CoA transacylase. (Luengo et al., 2003)
2.11.1 Polyhydroxyalkanoate production in recombinant Escherichia coli
In recent years, a combination of genetic engineering and molecular
microbiology techniques has been applied to enhance PHA production in
microorganisms. Several mutants with phenotypes in PHA synthesis were
characterized in order to develop optimal recombinant host strains. Over-expression

39
of pha genes in the natural PHA producer, however, resulted in little difference in
polymer accumulation. Natural producers, such as R. eutropha, are well adapted to
PHA accumulation in their cells. R. eutropha can store up to 90% of its dry weight
(dwt) in PHA granules. Most natural producers, however, take a long time to grow
during fermentation and extraction of polymers from their cells is difficult. Therefore,
these PHA producers are not suitable for industrial production of the biopolymer. On
the other hand, although E. coli does not naturally produce PHA, this bacterium is
considered to be appropriate host for generating higher yields of the biopolymer
because of its fast growth and the ease with which it can be lysed. Even after
extensive attempts at maximizing PHB production in non-PHB producing
microorganisms, the PHB accumulation level was not as high as what could be
obtained with the natural producers of the biopolymer. One of the major obstacles in
producing PHB in recombinant organisms is associated with the instability of the
introduced pha genes. Loss of the plasmid due to metabolic load often limits high
yields of the biopolymer (Steinbüchel and Pieper, 1992; Lee et al., 1996b; Madison
and Huisman, 1999). Other parameters have been adjusted to enhance PHB
production including increased carbon supply, changes in fermentation temperature,
changes in the number of plasmid copies and choice of bacterial strains (Kim et al.,
1994; Lee et al., 1996b, Nikel et al., 2006b). Growth of the recombinant cells was
impaired in many of these studies, especially in nutrient-rich medium (Lee, 1996a;
Wang and Lee, 1997). Recombinant E. coli cultured under optimal conditions has
been shown to accumulate PHB up to 85% of the cell dwt. PHB formed in these E.
coli, however, were of higher molecular weight than PHB produced by natural
producers (Zhang et al., 2004). The molecular mass of the PHB produced in E. coli
cells depended strongly on culture condition. In higher glucose concentration (20 g/l),
37 °C and pH 6.0, cells produced PHB with highest molecular mass value (20 MDa).
It has been suggested that a chain-transfer agent is generated in E. coli cells during the
accumulation of PHB. After the cloning of the R. eutropha PHA biosynthesis genes in
E. coli, recombinant E. coli has been investigated for the production of P(3HB)
because it has several advantages over other bacteria (Lee, 1996b). There have been a
series of papers that described the development of host–plasmid systems and the
strategies for producing a high concentration of P(3HB) with high productivity (Wang
and Yu, 2000). By the pH-stat fed-batch culture of recombinant E. coli harboring the
R. eutropha PHA biosynthesis genes, a cell dry weight of 206 g:l, P(3HB)

40
concentration of 149.7 g:l, and P(3HB) content of 73% were achieved in a chemically
defined medium, resulting in the P(3HB) productivity of 3.4 g:l per h (Wang and Yu,
2001). During the fed-batch culture of recombinant E. coli, a large amount of oxygen
was necessary to maintain the dissolved oxygen concentration above 10% of air
saturation. Since the use of a large amount the efficient production of P(3HB) by E.
coli. Recombinant E. coli strains harboring the initially cloned 6.4-kb DNA fragment
of A. latus produced P(3HB) to 50% of dry cell weight. A higher P(3HB)
concentration and P(3HB) content could be obtained by deleting the unnecessary
DNA fragment upstream of the PHA biosynthesis operon. By the pH-stat fed-batch
culture of recombinant E. coli harboring an optimally designed plasmid containing the
A. latus PHA biosynthesis genes in a chemically defined medium, final cell and
P(3HB) concentrations of 194.1 and 141.6 g:l, respectively, were obtained in 30.6 h,
resulting in a much higher productivity of 4.63 g P(3HB)/l/h. This should allow more
economical production of P(3HB) by recombinant E. coli.
2.11.2 Polyhydroxyalkanoate production in eukaryotic cells
The production of bioplastic in bacteria is limited by its high cost compared to
the costs associated with petroleum-derived plastics production. This aspect has been
one of the driving forces in exploring eukaryotic systems, especially crops, as
production hosts. Studies of PHA formation in yeast and insect cells can provide
valuable information about how these pathways can be incorporated into plants.
Synthesis of PHB has been demonstrated in Saccharomyces cerevisiae by expressing
the PHB synthase gene from R. eutropha. PHB accumulation, however, as very low
(0.5% of cell dwt), possibly because of insufficient endogenous β-ketoacyl- CoA-
thiolase and acetoacetyl- CoA reductase activities. To improve the yield and to
synthesize copolymers of PHAs, studies have focused on channeling the intermediates
of β-oxidation pathway into PHA assembly. Poirier et al., 1995, introduced a modified
phaC1 gene from P. aeruginosa into S. cerevisiae. Peroxisomal targeting (PTS1) of
the gene product was achieved by developing a construct which resulted in the
addition of a 34 amino acid stretch from the carboxylic end of Brassica napus
isocitrate lyase. When the recombinant yeast cells were grown in media containing
fatty acids, they accumulated mcl-PHAs demonstrating that peroxisomal PHA
synthase produces PHA in the peroxisomes using 3-hydroxyacyl- CoA intermediates
of fatty acid oxidation. In contrast to S. cerevisiae, Pichia pastoris grows vigorously

41
on fatty acids as a carbon source. Poirier et al., 1992, introduced the above PTS1-
modified P. aeruginosa phaC1 gene into P. pastoris and achieved mcl-PHA synthesis
in this yeast system with fatty acids in the growth medium. The yield of PHA in the
two described studies with yeast systems, however, was low, with accumulations
lower than 1% cell dwt. Possibilities have been explored of changing monomer
composition of PHA in recombinant yeast cells. The investigators demonstrated that it
was possible to alter the PHA monomer composition of mcl-PHAs produced in yeast
from the intermediates of the β-oxidation of fatty acids by using a modified form of
the peroxisomal multifunctional enzyme 2 (MFE-2, encoded by the fox2 gene). They
transformed yeast cells with genes coding for two mutant forms of the 3-hydroxyacyl-
CoA dehydrogenase domain of the MFE-2 of S. cerevisiae. The mutant MFE-2(aΔ)
retain a broad activity towards short-, medium- and long-chain (R)-3-hydroxyacyl-
CoAs, while the mutant MFE-2(bΔ), did not accept shortchain (R)-3-hydroxyacyl-
CoAs. Expression of MFE-2 (bΔ), along with PHA synthase, resulted in a substantial
increase in the proportion of the short-chain 3-hydroxyacid monomers at the expense
of longer monomers. These transformant yeast cells were inefficient at using short-
chain (R)-3-hydroxyacyl-CoAs generated by the β-oxidation cycle, leading to higher
levels of these intermediates available to the PHA synthase. Zhang et al., 2004,
engineered the synthesis of PHA polymers composed of monomers ranging from 4 to
14 carbon atoms in either the cytosol or the peroxisome of S. cerevisiae by harnessing
intermediates of fatty acid metabolism and achieved accumulation of PHA up to
approximately 7% of its cell dry weight. Insect cells have also been studied as a
model for PHA production in eukaryotes. The phaC gene from R. eutropha was
successfully expressed in cabbage looper cells and a soluble form of PHB synthase
that could be rapidly purified was obtained (Williams and Martin, 2002). In a separate
attempt, Williams and Martin, 2002 transfected fall armyworm cells with a modified
eukaryotic fatty acid synthase, which did not extend fatty acids beyond HB, along
with the phaC gene from R. eutropha. PHB production was achieved in the
transfected cells, although the yield was very low (% of cell dry weight).
2.11.3 Polyhydroxyalkanoate synthesis in transgenic plants
Polyhydroxyalkanoate production in bacteria and yeast requires growth under
sterile condition in a costly fermentation process with an external energy source such
as electricity. In contrast, PHA production in plant systems is considerably less

42
expensive because the system only relies on water, soil nutrients, atmospheric CO2
and sunlight. In addition, a plant production system is much more environmentally
friendly. Plants use photosynthetically fixed CO2 and water to generate the bioplastic,
which after disposal is degraded back to CO2 and water. Synthesis of PHAs in crops is
also an excellent way of increasing the value of the crops (Poirier et al., 1995). Since
starch and sugar are produced in plants at costs below the cost of commodity plastics,
it might be possible to produce PHA at a similar low cost. Unlike the bacterial cell,
the plant cell has different subcellular compartments in which PHA synthesis can be
metabolically localized. As mentioned earlier, PHB is synthesized in bacteria from
acetyl-CoA. This thioester is present in plant cells in the cytosol, plastids,
mitochondria and peroxisomes. Therefore, it should be possible to produce PHB in
any of these subcellular compartments.
2.12 Polyhydroxyalkanoate recovery processes
In addition to the costs of maintaining pure cultures and the high costs of
organic substrates, polymer recovery process is another factor that contributes to the
high overall cost of PHA production. In the past 2 decades, several recovery processes
have been investigated and studied in order find an economic way to isolate and
purify PHA. According to Doi et al., 1990; Lee, 1996a and Braunegg, 1998, several
methods have been used as a recovery process for PHA. These methods include
solvent extraction; the most common analytical technique used for PHA estimation
was a gravimetric method (Lemoigne, 1926), which consisted of PHB extraction from
lyophilized biomass with chloroform followed by precipitation with diethyl ether or
acetone. In 1958, Williamson and Wilkinson showed that under controlled conditions
of time and temperature all cell material, except PHB granules, dissolved in alkaline
sodium hypochlorite solution (Williamson and Wilkinson, 1958). Further
developments on this method were introduced (Law and Slepecky, 1969) wherein the
extracted PHB was converted (with concentrated sulphuric acid) to crotonic acid, and
estimated spectrophotometerically at 235 nm. Detection of PHA extracted with
chloroform can also be done by IR spectroscopy (at 5.75Å) (Jüttner et al., 1975), and
enzymatic digestion. Details of each method as well as their advantages and
disadvantages will be discussed and summarized here. In most cases, bacterial
biomass is separated from substrate medium by centrifugation, filtration or

43
flocculation. Then, the biomass is freeze dried (lyophilized). Basically, mild polar
compounds, e.g., acetone and alcohols, solubilize non-PHA cellular materials whereas
PHA granules remain intact. Non-PHA cellular materials are nucleic acids, lipids,
phospholipids, peptidoglycan and proteinaceous materials. On the other hands,
chloroform and other chlorinated hydrocarbons solubilize all PHAs. Therefore, both
types of solvents are usually applied during recovery process. Finally, evaporation or
precipitation with acetone or alcohol can be used to separate the dissolved polymer
from the solvent.
2.12.1 Solvent extraction
This method is used on a small scale for laboratory experiments as well as on
a large scale for commercial production. This method is a widely used method
because it is applicable to many PHA producing microorganisms. However, a large
amount of solvent is employed because PHA solution is highly viscous. According to
Lee, 1996b, approximately 20 parts of solvent is employed to extract 1 part of
polymer. This requirement makes solvent extraction a costly method. PHAs are
soluble in solvents, such as chloroform, methylene chloride or 1,2- dicholoroethane.
These 3 solvents can be used to extract PHA from bacterial biomass. In addition,
other solvents were also reported to be used to extract PHA, e.g., ethylene carbonate,
1,2-propylene carbonate, mixtures of 1,2-trichloroethane with water, and mixtures of
chloroform with methanol, ethanol, acetone or hexane. Doi et al., 1990, described a
chloroform extraction method. PHA is extracted with hot chloroform in a soxhlet
apparatus for over 1 hour. Then, PHA extracted is separated from lipids by
precipitating with diethyl ether, hexane, methanol, or ethanol. Finally, PHA is
redissolved in chloroform and further purified by precipitation with hexane. Ramsay
et al., 1995, examined the recovery of PHA from three different chlorinated solvents
(chloroform, methylene chloride, and 1,2-dichloroethane). They obtained the best
recovery and purity when biomass was pretreated with acetone. The optimum
digestion time for all three solvents was 15 minutes. Further digestion resulted in
degradation in the weight molecular weight (MW) of PHA. The degree of recovery
when the biomass were pretreated with acetone were 70, 24, and 66% when reflux for
15 minutes with chloroform, methylene chloride, and 1,2-dichloroethane,
respectively. Whereas the level of purity of these 3 solvents under these optimum
conditions were 96, 95, and 93%, respectively. Temperatures used of these three

44
solvents were 61, 40, and 83°C, respectively. The authors emphasized that extraction
conditions have a great impact on the degradation of PHA during the recovery
process.
2.12.2 Sodium hypochlorite digestion
Sodium hypochlorite solubilizes non-PHA cellular materials and leaves PHA
intact. Then, PHA can be separated from the solution by centrifugation. A severe
degradation of polymers during sodium hypochlorite digestion is frequently reported.
Because sodium hypochlorite is a strong oxidant, care has to be taken to select for
suitable digestion conditions in order to maintain a high molecular weight of the
polymers. Ramsay et al., 1995, examined the PHA recovery process from R. eutropha
using hypochlorite digestion with surfactant pretreatment. Two different surfactants
were investigated: Triton 100 X and sodium dodecyl sulfate (SDS). Improvements in
purity and molecular weight can be obtained by pretreating with surfactant prior to the
extraction with sodium hypochlorite. They reported that surfactant removed
approximately 85% of the total protein and additional protein (10%) was further
removed by sodium hypochlorite digestion. They also stated that this method resulted
in a high MW of extracted PHA and the recovery time was reduced when compared
to surfactant-enzymatic treatment or solvent extraction. In addition, a native PHA
granule could be maintained during this treatment, which allows PHA to be used for
more diverse applications in comparison to solvent extraction method.
The above-mentioned methods are time-consuming and not accurate
particularly for low PHB concentrations (Lafferty et al., 1988). Large sample volumes
and a high degree of replicates are required to minimize errors. Furthermore, cell
material can interfere with PHB extraction (Braunegg et al., 1978). These methods
allow for the quantification of total PHA but not the individual PHA components.
2.12.3 Gas chromatography method
The use of gas chromatography (GC) for the identification of PHA
components was then proposed. This method involves simultaneous extraction and
methanolysis of PHA, in mild acid or alkaline conditions, to form hydroxyalkanoates
methyl esters, which are then analyzed by GC (Braunegg et al., 1978). This method is
rapid (4 h), sensitive, reproducible and requires only small samples. It features
reaction and extraction in the same screw-capped tube. Another GC method for

45
increased PHA recovery was proposed by carrying out propanolysis in HCl (Riis and
Mai, 1988) rather than acidic methanolysis in sulphuric acid.
Other recent methods for PHA quantification include HPLC (Karr et al.,
1983), ionic chromatography, and enzymatic determination (Hesselmann et al., 1999).
HPLC measures only PHB and is based on conversion of PHB to crotonic acid
followed by UV detection at 210 nm. This method features 84% recovery of PHB.
PHA detection by ionic chromatography is based on the conversion of monomers to
alkanoic acids. The determination involves acid propanolysis followed by an alkaline
hydrolysis with Ca(OH)2 or acidic hydrolysis with concentrated H2SO4. After
centrifugation, the sample is injected into an anionic column with a conductivity
detector. 3HB is enzymically oxidized and the NADH produced from NAD+ was
reoxidised in the presence of iodonitro-tetrazoliumchloride to produce formazan,
which was spectrophotometerically measured at 492 nm. The determination of PHA
inside intact cells by two-dimensional fluorescence spectroscopy and flow cytometry
has also been proposed recently. Cells stained with Nile Blue, show a clear
fluorescence maximum between 570 and 605 nm when excited between 540 and 560
nm. A good correlation between fluorescence intensity and PHB concentration was
obtained. However, differentiation of PHA composition was not possible with this
method. Ease of recovery of PHA is a very important parameter in their economical
production. Thus, there is a need for development of methods for extraction of PHB
so that the overall process could be made much simpler and cheaper.
2.13 Applications of polyhydroxyalkanoates
2.13.1 Medical and pharmaceutical
The degradation product of P(3HB), D-3-hydroxybutyric acid, is a common
intermediate metabolic compound in all higher organisms (Lee, 1996a). Therefore, it
is plausible that it is biocompatible to animal tissues and P(3HB) can be implanted in
animal tissues without any toxic. Some possible applications of bacterial PHAs in the
medical and pharmaceutical applications include: biodegradable carrier for long term
dosage of drugs inside the body, surgical pins, sutures, and swabs, wound dressing,
bone replacements and plates, blood vessel replacements, and stimulation of bone
growth and healing by piezoelectric properties. The advantage of using biodegradable

46
plastics during implantation is that it will be biodegraded, i.e., the need for surgical
removal is not necessary.
2.13.2 Agricultural
PHAs are biodegraded in soil. Therefore, the use of PHAs in agriculture is
very promising. They can be used as biodegradable carrier for long-term dosage of
insecticides, herbicides, or fertilizers, seedling containers and plastic sheaths
protecting saplings, biodegradable matrix for drug release in veterinary medicine, and
tubing for crop irrigation. Here again, it is not necessary to remove biodegradable
items at the end of the harvesting season.
2.13.3 Biodegradable commodity packaging
PHA has a wide range of applications owing to their novel features. Initially,
PHA was used in packaging films mainly in bags, containers and paper coatings.
Similar applications as conventional commodity plastics include the disposable items,
such as razors, utensils, diapers, feminine hygiene products, cosmetic containers––
shampoo bottles and cups. In addition to potential as a plasticmaterial , PHA are also
useful as stereoregular compounds which can serve as chiral precursors for the
chemical synthesis of optically active compounds (Oeding and Schlegel, 1973; Senior
and Dawes, 1973). Such compounds are particularly used as biodegradable carriers
for long term dosage of drugs, medicines, hormones, insecticides and herbicides.
They are also used as osteosyntheticmaterial s in the stimulation of bone growth
owing to their piezoelectric properties, in bone plates, surgical sutures and blood
vessel replacements. However, the medical and pharmaceutical applications are
limited due to the slow biodegradation and high hydraulic stability in sterile tissues
(Wang and Bakken, 1998). PHA is considered as a source for the synthesis of chiral
compounds (enantiometrically pure chemicals) and is raw material for the production
of paints. PHA can be easily depolymerised to a rich source of optically active, pure,
bi-functional hydroxy acids. PHB for instance is readily hydrolyzed to R-3-
hydroxybutyric acid and used in the synthesis of Merck anti-glaucoma drug
“Truspot”. In tandem with R-1,3- butanediol, it is also used in the synthesis of β
lactams. Plant derived PHA can be depolymerized and used, directly or following
esterification, in the manufacture of bulk chemical. Besides helping to replace the
existing solvents, β-hydroxy acid esters and derivatives are likely to find growing use

47
as green solvents similar to lactic acid esters. The conversion of hydroxy acids into
crotonic acids such as 1,3-butanediol, lactones, etc. would help improve the market
value as they have an existing market demand in thousands of tonnes.
2.14 Economics of polyhydroxyalkanoate production
It is a prerequisite to standardize all the fermentation conditions for the
successful implementation of commercial PHA production systems. The price of the
product ultimately depends on the substrate cost, PHA yield on the substrate, and the
efficiency of product formulation in the downstream processing (Lee, 1996b). This
implies high levels of PHA as a percentage of cell dry weight and high productivity in
terms of gram of product per unit volume and time (de Koning and Witholt, 1997; de
Koning et al., 1997). Commercial applications and wide use of PHA is hampered due
to its price. The cost of PHA using the natural producer A. eutrophus is US$16 per Kg
which is 18 times more expensive than polypropylene. With recombinant E. coli as
producer of PHA, price can be reduced to US$4 per Kg, which is close to other
biodegradable plastic materials such as PLA and aliphatic polyesters. The
commercially viable price should come to US$3–5 per Kg (Lee, 1996b). The effect of
various substrate costs and the yield on the P(3HB) production cost are described in
Table 2.6. The development of PHB was begun by Imperial Chemical Industries (ICI)
in 1975–76 as a response to the increase in oil prices. ICI started making “Biopol” as
early as in 1982 from A. eutrophus (H16). Cargill Dow Polymers is marketing its
“EcoPla” resins in Japan, Novamont_s starch-based material, “Mater-Bi”, is marketed
in Japan by Nippon Gohsei. Mater-Bi has been used in transport packaging for
electrical goods, agricultural mulch film and in composting trials. Mitsubishi and
Nippon Shokubai under the trade names “LUNARE ZT” and “Lunare SE” market
“EnviroPlastic”. “Bionolle”, thermoplastic aliphatic polyester, is manufactured by
Showa Highpolymer and Denko of Japan. It is produced by the polycondensation of
glycol with dicarboxylic acids. “Lacea” is another type of bioplastic manufactured by
Mitsui Chemicals, Japan, from fermented starch, derived from a variety of renewable
resources, such as corn, beet, cane, and tapioca. Lacea is comparable to polyethylene
in terms of transparency and similar to polystyrene or polyethylene in terms of
processability. It also claims good mould resistance, low heat of combustion which is
similar to that of paper, superior stability in processing use and biodegradability

48
superior to that of earlier polylactic acid-based materials. Daicel Chemical Industries,
Japan has developed biodegradable blends of two different kinds of material,
polycaprolactone and acetyl cellulose resin with the brand name “Celgreen”.
Shimadzu developed a fermentation process for lactic acid and collaborated with
Mitsubishi Plastics Industries to develop poly-L-lactic acid. The resins are marketed
under the trade name “Lacty”. The other bioplastics manufactured by Japan based
firms are “Eco-Ware” and “Eco-Foam” (which are starch-based) and Cardoran and
Pulluran (which are based on polysaccharides).

![The Normal patched Allele Is Expressed in Medulloblastomas ...cancerres.aacrjournals.org/content/60/8/2239.full.pdf · [CANCER RESEARCH 60, 2239–2246, April 15, 2000] The Normal](https://img.pdfslide.us/doc/110x75/5a74e6ed7f8b9a93088bf69f/the-normal-patched-allele-is-expressed-in-medulloblastomas-cancer-research.jpg)



























