Embed Size (px)
Citation preview

# CHAPTER II # 8
CHAPTER II
Review of LiteRatuRe

# CHAPTER II # 9
2.1. Enzymes
Enzymes are proteins and are nature's own biocatalysts. Enzymes have many
advantages over their chemical counterparts in that they are more specific, and generally
possess high catalytic properties. Almost all processes in a biological cell need enzymes
to occur at significant rates. More than 4000 enzymes catalyzing a wide array of reactions
are known to exist. Since the 1950s, Enzyme technology has really taken off. It is the
basis of the new industry called biotechnology. There are great benefits in using enzymes
as catalysts to make products. They can be some 10,000 times more efficient than
ordinary inorganic catalysts used in industry. One enzyme molecule can catalyze 10
million reactions in a single second!
The enzymes are classified into six major categories based on the nature of the
chemical reaction they catalyze:
1. Oxidoreductases catalyses oxidation or reduction of their substrates
2. Transferases catalyses group transfer
3. Hydrolases catalyses bond breakage with the addition of water
4. Lyases remove groups from their substrates
5. Isomerases catalyses intramolecular rearrangements
6. Ligases catalyses the joining of two molecules at the expense of chemical
energy
More than 75% of industrial enzymes are hydrolases. And amongst them, amylase
account for about 30% of the world’s enzyme production (Maarel et al., 2002). Amylases

# CHAPTER II # 10
have even found new applications but their use in detergents is the major market. Figure
2.1 illustrates the diverse use of amylase in various sectors.
Figure 2.1. Use of amylase enzyme in diverse Business areas
2.2. Amylase
Amylases are starch degrading enzymes. They are widely distributed in microbial,
plant and animal kingdoms. However, enzymes from fungal and bacterial sources have
dominated applications in industrial sectors (Pandey et al., 2000). They degrade starch

# CHAPTER II # 11
and related polymers to yield products characteristic of individual amylolytic enzymes.
Initially the term amylase was used originally to designate enzymes capable of
hydrolyzing α-1,4-glucosidic bonds of amylose, amylopectin, glycogen and their
degradation products (Bernfeld, 1955; Fisher and Stein, 1960; Myrback and Neumuller,
1950). They act by hydrolyzing bonds between adjacent glucose units, yielding products
characteristic of the particular enzyme involved. Amylases have emerged as one of the
leading biocatalysts with proven potential to find usage in a wide array of industrial
applications, such as additives in processed food industries, additives in detergents,
waste-water treatment, biopulping, bioremediations and in molecular biology. The
spectrum of amylase application has widened in many other fields, such as clinical,
medical, analytical chemistry, as well as their wide spread application in starch
sacccharification, in the textile, food, fermentation, paper, brewing and distilling
industries (Pandey et al., 2000). Figure 2.2 illustrates the disparity of sale of amylase in
industries in year 2000 and 2010.
2.2.1 History of Amylase
In 1831, Erhard Friedrich Leuchs (1800-1837) described the diastatic action of
salivary ptyalin (amylase) on starch. The modern history of enzymes began in 1833 when
French chemists described the isolation of an amylase complex from germinating barley
and named it diastase. In 1862, Danielewski separated pancreatic amylase from trypsin.

# CHAPTER II # 12
2.2.2 Classes of amylases
1. α-Amylase
Most of the α-amylases (endo-1,4-α-D-glucan glucohydrolase, EC 3.2.1.1) are
metalloenzymes, which require calcium ions (Ca2+) for their activity, structural integrity
and stability. They belong to family 13 (GH-13) of the glycoside hydrolase group of
enzymes (Bordbar et al., 2005). The specificity of the bond attacked by α-amylases
depends on the sources of the enzymes. Currently, two major classes of α-amylases are
(a)

# CHAPTER II # 13
(b)
Figure 2.2. Distribution of sales of amylase by various industries (a) in 2000 and (b) in
2010
commercially produced through microbial fermentation. Based on the points of attack in
the glucose polymer chain, they can be classified into two categories, liquefying and
saccharifying. Bacterial α-amylase randomly attacks only the α-1,4 bonds yielding
maltotriose and maltose from amylose, or maltose, glucose and "limit dextrin" from
amylopectin, it belongs to the liquefying category. The hydrolysis reaction catalyzed by
this class of enzymes is usually carried out only to the extent that, for example, the starch
is rendered soluble enough to allow easy removal from starch-sized fabrics in the textile
industry. The paper industry also uses liquefying amylases on the starch used in paper
coating where breakage into the smallest glucose subunits is actually undesirable. On the
other hand, the fungal α-amylase belongs to the saccharifying category and attacks the
second linkage from the non reducing terminals (i.e. C4 end) of the straight segment,

# CHAPTER II # 14
resulting in a disaccharide called maltose. The bond breakage is thus more extensive in
saccharifying enzymes than in liquefying enzymes (Marrel et al., 2002).
Microbes are the major source of alpha-amylase. Microbial α-amylase has been shown to
produced by several fungi and moulds like Rhizopus, Aspergillus, etc. Bacterial amylase
might be produced by Bacillus, Pseudomonas, Saccharophilia, Clostridium and several
other species. But on industrial scale the strains of Bacillus species seem to be preferred
(Bordbar et al., 2005).
2. β-Amylase
Another form of amylase is β-amylase (1,4-α-D-glucan maltohydrolase, EC
3.2.1.2) is also synthesized by bacteria, fungi, and plants. Working from the non-reducing
end, β-amylase catalyzes the hydrolysis of the second α-1,4 glycosidic bond, cleaving off
two glucose units (maltose) at a time. During the ripening of fruit, β-amylase breaks
starch into maltose, resulting in the sweet flavor of ripe fruit.
Both α-amylase and β-amylase are present in seeds; β-amylase is present in an inactive
form prior to germination, whereas α-amylase and proteases appear once germination has
begun. Cereal grain amylase is key to the production of malt. Many microbes also
produce amylase to degrade extracellular starches. Animal tissues do not contain β-
amylase, although it may be present in microorganisms contained within the digestive
tract (Hill and Needham, 1970).

# CHAPTER II # 15
3. γ-Amylase
In addition to cleaving the last α-1,4 glycosidic linkages at the non-reducing end
of amylose and amylopectin, yielding glucose, γ-amylase also cleave α-1,6 glycosidic
linkage. Unlike the other forms of amylase, γ-amylase (Glucan 1,4-α-glucosidase, EC
3.2.1.3) is most efficient in acidic environments and has an optimum pH of 3.
2.2.3. Alpha Amylase Family
The α-amylase family, i.e. the clan GH-H of glycoside hydrolyses, is the largest
family of glycoside hydrolases, transferases and isomerases comprising nearly 30
different enzyme specificities (Henrissat, 1991). A large variety of enzymes are able to
act on starch. These enzymes can be divided basically into four groups: endoamylases,
exoamylases, debranching enzymes and transferases (Marrel et al., 2002);
1. endoamylases: cleave internal α-1,4 bonds resulting in α-anomeric products.
2. exoamylases: cleave α-1,4 or α-1,6 bonds of the external glucose residues resulting in
α- or β-anomeric products.
3. debranching enzymes: hydrolyze α-1,6 bonds exclusively leaving long linear
polysaccharides, and
4. transferases: cleave α-1,4 glycosidic bond of the donor molecule and transfer part of
the donor to a glycosidic acceptor forming a new glycosidic bond.
Glycoside hydrolases are able to metabolize a large variety of saccharides. They
have been divided into classes based on their mode of reaction and families based on
their well-defined amino acid sequence similarities. Most of the starch converting
enzymes belongs to GH-13 family. GH-13 family can be further classified based on a

# CHAPTER II # 16
larger unit called clan, which is the three dimensional structure of catalytic module. A
clan may consist of two or more families with the same three-dimensional structure of
catalytic domain but with limited sequence similarities, indicating that protein structure is
better preserved by evolution than amino acid sequence. Among the fourteen clans (A–N)
defined for glycosidases and transglycosidases, α-amylase family (GH-13) belongs to the
eighth clan, clan GH-H (MacGregor, 2005).
The concept of this group of enzymes as the α-amylase was proposed in 1992
(Takata, 1992). According to the definition, the members of this family must satisfy the
following requirements:
(i) They must act on α-glucosidic linkages and hydrolyze them to produce α-
anomeric monosaccharides and oligosaccharides or form α-glucosidic
linkages by transglycosylations,
(ii) Have four highly conserved regions in their primary structures consisting of
catalytic and important substrate-binding sites,
(iii) Have Asp, Glu, and Asp residues as catalytic sites corresponding to the Asp 206,
Glu 230 and Asp 297 of Taka amylase A, and
(iv) Possess a (β/α)8 or TIM barrel catalytic domain.
2.2.4 Characteristics of Catalytic Domain of α-Amylases
Four conserved sequence regions covering the strands β3, β4, β5 and β7 of the
catalytic (β/α)8-barreled domain were identified and used for defining the α-amylase
family. The two dimensional structure of α-amylase prototype consists of three domains,
namely A, B and C. Domain A is the N-terminal TIM barrel structure, domain B consists

# CHAPTER II # 17
of a long loop that protrudes between β-strand 3 and α-helix 3 and C domain with a β
sheet structure linked to A domain. The (β/α)8 barrel consists of eight alternate β-strands
and α-helices. The β strands are placed parallel to one another as if on a cylinder and α-
helices lie outside the cylinder. This TIM barrel contains four highly conserved regions
closely related to the active site present in all α-amylases (Kuriki et al., 2005). They are:
(i) First region: C-terminal end of β-strand 3 and histidine residue which interacts with
the glucose residue of the substrate,
(ii) Second region: β-strand 4 with Asp residue, which acts as the nucleophile during
catalysis,
(iii) Third region: β-strand 5 with glutamic acid residue acting as proton donor/acceptor,
and
(iv) Fourth region: β-strand 7 with a histidine residue and an Asp residue that may form
hydrogen bonds with glucose residue of the substrate.
While the four original conserved sequence regions contain the catalytic and substrate
binding residues of the individual members of the family, three additional conserved
sequences were later identified and they are shown to contain amino acid residues
connected to a given enzyme specificity. Two of these three regions roughly cover the
strands of α2 and α8 of the catalytic (β/α)8 barrel and one is located near the C-terminus of
domain B (Janecek, 2002).

# CHAPTER II # 18
2.3. COLD ACTIVE ENZYMES
2.3.1. Definition of psychrophile/psychrotolerant
The definition by Ingraham and Stokes (1959), that psychrophilic bacteria should
show visible growth at 0°C after 14 days of incubation, has been renewed by the seminal
paper of Morita (1975). Further publications and reviews (Baross and Morita, 1978;
Deming, 2002; Friedmann, 1994; Herbert, 1986; Inniss and Ingraham, 1978; Russell,
1990) have set the frame for defining psychrophilic organisms and their environments.
The classical definition of Morita (1975) is frequently used in the literature. This
definition proposes that psychrophilic microorganisms have optimum growth
temperatures of <15°C and upper cardinal temperatures of ~20°C. He further
distinguished psychrophiles from psychrotrophs (psychrotolerants) on the basis of their
cardinal temperatures, in that the latter have a minimum growth temperature, which is at
or just above zero, and optimum and maximum growth temperatures above 20°C.
Bacteria Minimum temp.
(Tmin)
Optimum temp.
(Topt)
Maximum temp.
(Tmax)
Psychrophilic <0°C <15°C <20°C
Psychrotolerant <0°C >15°C >20°C
The definition of psychrophilic microorganisms is ambiguous for three main reasons.
First, the temperature limits have been arbitrarily selected and do not correspond to any
clear separation of biological processes or environmental conditions. Second, Morita’s

# CHAPTER II # 19
definition does not apply to most eukaryotes. Finally, and most important,
microorganisms behave as thermodynamic units: increasing the culture temperature
increases reaction rates and the growth rate. However, they also behave as biological
units: at a given temperature, key metabolic steps are heat labile and impair the
functioning of some pathways. It has been shown, for instance, that although the growth
rate of some cold-adapted bacteria increases with a temperature shift from 5°C to 25°C,
the physiological state is strongly altered, as judged by the decrease in viable counts,
exoenzyme production, protein synthesis and membrane permeability (Orange, 1994;
Feller et al., 1994b). This emphasizes that the use of growth rates to define the optimum
growth temperature is inappropriate.
2.3.2. Habitats of psychrophilic/psychrotolerant bacteria
Around 85% of earth is occupied by cold ecosystems including the ocean depths,
polar and alpine regions. Out of which ~70% is covered by oceans that have a constant
temperature of 4-5°C below a depth of 1000 m, irrespective of the latitude. Remaining
15% included Polar Regions to which the glacier and alpine regions must also be added.
Extremophiles successfully colonized on these eternally cold environments which we can
call as psychrophiles (which literally means cold-loving). Psychrophilic microorganisms,
including bacteria, yeasts, fungi and microalgae, can be found in soils, in water (fresh and
saline, immobile and flowing) and associated with plants and cold-blooded animals.
Numbers of pathogenic psychrotolerant pseudomonads are also reported in plant
kingdom causing variety of diseases, including soft rots and wilts. These pseudomonads

# CHAPTER II # 20
produce ice-nucleation proteins, as part of their pathogenic mechanism which has
biotechnological potential.
2.3.3. Cold active amylases
Alpha-amylases are ubiquitous enzymes produced by plants, animals and
microbes, where they play a dominant role in carbohydrate metabolism. Cold-adapted
amylolytic microorganisms produce amylases, which function effectively at low
temperatures with high rates of catalysis in comparison to the amylases from mesophiles
or thermopiles, which shows little or no activity at low temperature. These amylases have
evolved a range of structural features that confer a high level of flexibility, particularly
around the active site are translated into low activation enthalpy, low-substrate affinity,
and high specific activity at low temperatures. Moreover, the maximum level of activity
of these amylases is shifted towards lower temperatures with a concomitant decrease in
thermal stability. The knowledge of cold-active amylolytic enzymes is increasing at a
rapid and exciting rate.
2.3.4. Sources of cold-active amylases
Enzyme adapted to cold can be produced by prokaryotic as well as by eukaryotic
organisms. Up to now, most of them originate from bacteria and fish living in polar
regions and especially in Antarctic sea water which represent a permanently cold (0±2°C)
and constant temperature habitat (Feller et al., 1996).
Psychrophilic (cold-loving) or psychrotolerant (cold-adapted) microorganisms
producing cold-active amylases are found inhabiting the low temperature environments
of the Earth, including polar regions, high mountains, glaciers, ocean deeps, shallow

# CHAPTER II # 21
subterranean systems (i.e. caves), the upper atmosphere, refrigerated appliances and the
surfaces of plants and animals living in cold environments, where temperature never
exceed 5°C. The potentials of psychrophiles and psychrophilic enzymes including
amylase have been reviewed by Cavicchioli et al. (2002); Deming (2002); Margesin et al.
(2002); Feller and Gerday (2003) and Georlette et al. (2004). Many psychrophiles live in
biotopes having more than one stress factors, such as low temperature and high pressure
in deep seas (piezo-psychrophiles), or low temperature and high salt concentration in
salty seas (halo-psychrophiles) also produces amylase (Cavicchioli et al., 2002; Deming,
2002; Margesin et al., 2002; Feller and Gerday, 2003; Georlette et al., 2004).
A diverse range of psychrophilic microorganisms, belonging to bacteria, archaea,
yeast and fungi have been isolated from these cold environments (Cavicchioli et al.,
2002; Deming, 2002; Margesin et al., 2002; Feller and Gerday, 2003; Georlette et al.,
2004). In spite of the wide distribution of amylases, microbial sources, namely fungal and
bacterial amylases, are used for the industrial production due to advantages such as cost
effectiveness, consistency, less time and space required for production and ease of
process modification and optimization (Burhan et al., 2003). Among bacteria, Bacillus sp.
is widely used for α-amylase production to meet industrial needs. B. subtilis, B.
stearothermophilus, B. licheniformis and B. amyloliquefaciens are known to be good
producers of α-amylase and these have been widely used for commercial production of
the enzyme for various applications. Similarly, fungi belonging to the genus Aspergillus
have been widely used for the production of amylases for centuries (Pandey et al., 2000).

# CHAPTER II # 22
In addition, some recent examples of cold-active amylase producing bacteria are shown
in the Table 2.1.
Table 2.1: Cold-active amylase producing microbes
Microorganisms Source Reference
Bacillus cereus GA6
Microbacterium foliorum GA2
Wangia sp. C52
P. arctica GS230
β-proteobacteria
Gangotri glacier, India
Gangotri glacier, India
Southern Okinawa
Seawater from Gaogong island,
China
Roopkund Glacier, Himalayan
range, India.
Kuddus et al. (2012)
Roohi et al. (2011)
Liu et al. (2011b)
Lu et al. (2010)
Suman et al. (2010)
Nocardiopsis aegyptia
Marine sediment of Abu Qir Bay,
Alexandria, Egypt
Abou-Elela et al.
(2009)
Streptomyces 4 Alga Soil and vegetation (East Antarctica) Cotarlet et al. (2009)
Micrococcus antarcticus Antarctica Fan et al. (2009)
Bacterial strains Sediment samples from Midtre
Lovenbreen Arctic glacier
Reddy et al. (2009)
Culturable bacteria Sediment and a soil from
Kongsfjorden and Ny-Alesund,
Svalbard, Arctic.
Srinivas et al.
(2009)
Nocardiopsis sp. 7326 Deep sea sediment of Prydz Bay,
Antarctic.
Zhang and Zeng
(2008)

# CHAPTER II # 23
Lactobacillus plantarum
MTCC 1407
Central Tuber Crop Research
Institute, Bhubaneswar, India
Smita et al. (2008)
Amy I and Amy II Eisenia foetida
Earthworm, Osaka, Japan.
Ueda et al. (2008)
Arthrobacter
psychrolactophilus
ATCC 700733
Pennsylvania soil Michael et al.
(2005)
Aspergillus oryzae Culture from University of
Agriculture, Abeokuta, Nigeria
Akpan and Adelaja,
(2004)
Gamma-Proteobacteria Permanently cold fjords of
Spitzbergen, Arctic Ocean.
Tatiana et al. (2004)
Aspergillus ochraceus Soil Nahas and Mirela
(2002)
Alteromonas sp. Antarctic sea water Chessa et al. (1999)
2.4. STRUCRURAL MODIFICATIONS FOR COLD ADAPTATION
Temperature is one of the most important environmental factors for life, as it
influences most biochemical reactions. A reduction in temperature slows down most
physiological processes, changes protein-protein interactions, reduces membrane fluidity
and provokes an increased viscosity of water. Moreover, Enzymes are also subject to cold
denaturation, leading to the loss of enzyme activity at low temperatures (Yancey and
Somero, 1978; Somero, 1981). Psychrophilic microorganisms producing cold-active
amylases are structurally modified by an increasing flexibility of the polypeptide chain

# CHAPTER II # 24
enabling an easier accommodation of substrates at low temperature. They must modify
their lipid composition to maintain membrane fluidity at environmental temperatures.
This can be done in many ways;
a) Unlike cold-adapted proteins, which improve their structural mobility, the thermal
adaptation of membrane lipids does not involve the synthesis of fatty acyls that have
increased degrees of freedom, but rather the introduction of steric constraints that reduce
the packing of acyl chains in the membrane. These steric constraints destabilize the
membrane and reduce the lipid viscosity (Margesin et al., 2002; Russell and Hamamoto,
1998; Russel, 1997).
b) Presence of a cis-unsaturated double bond in the chain that induces a 30° bend. Such
bending creates a cavity in the lipid layer and perturbs the packing density. Trans-
unsaturated double bonds are also observed, but are less efficient as they only produce a
modest kink of the acyl chain.
c) The occurrence of branched lipids mainly methyl-branched fatty-acyl chains also
perturbs the compactness of neighbouring chains owing to the steric hindrance that is
caused by the side-chain group. The position of this branching along the chain also
modulates the gel-phase transition temperature.
d) Finally, shorter fatty-acyl chains reduce the contacts between adjacent chains and
increase fluidity (Jagannadham, 2000; Fong et al., 2001).
Besides the variations in membrane structure and its lipid composition, another
important feature in these cold-adapted microorganisms is the presence of cold-
acclimation proteins (CAPs). These are a set of ~20 proteins which is permanently
synthesized during steady-state growth at low temperatures, but not at milder

# CHAPTER II # 25
temperatures (Hebraud and Potier, 2000; Hebraud et al., 1994; Berger et al., 1996).
Interestingly, some of the few CAPs that have been identified in cold-adapted bacteria
actually acting as cold-shock proteins in mesophiles, such as the RNA chaperone CspA
(Berger et al., 1997). It has been proposed that these CAPs are essential for the
maintenance of both growth and the cell cycle at low temperatures (Hebraud and Potier,
2000) but their function is still poorly understood.
Cold-shock proteins (Csp) are an additional type of adaptation for psychrophilic
organisms in cold regions. These proteins act mainly on the regulation of cellular protein
synthesis, particularly at the level of transcription and the initiation of translation; and
they also act as chaperone by preventing the formation of mRNA secondary structures.
Advantage of these Csp’s is that the synthesis of housekeeping gene products is not
inhibited by cold-shock which is normally occurring in their mesophilic and thermophilic
homologues (Cavicchioli et al., 2000; Berger et al., 1996; Mayr et al., 1996).
Antifreeze proteins, AFPs (are peptides and glycopeptides of various sizes) are
more frequently occurring in fishes, insects, plants, fungi and some microorganisms
which decrease the freezing point of cellular water by binding to ice crystals and prevent
the destruction of cell membranes and the disruption of osmotic balance. Besides
contributing to freeze resistance and freeze tolerance, AFPs also helped to increase
species diversity in some of the harshest and most inhospitable environments (Barrett,
2001; Jia and Davies, 2002). Although antifreeze proteins have been reported in several
eukaryotes, there is no supporting evidence for the occurrence of such glycopeptides in
psychrophilic prokaryotes.

# CHAPTER II # 26
To define the properties of a cold-active enzyme, the effect of temperature on the
activity of psychrophilic and mesophilic enzymes is illustrated in Figure 2.3.
Figure 2.3. Temperature dependence of enzyme activity.
[Psychrophilic enzymes (left curve; blue) are up to tenfold more active at low and
moderate temperatures (up to 20–30°C) than their mesophilic homologues (right curve;
red). Such high activity compensates the cold-induced inhibition of reaction rates.
However, the activity of cold-adapted enzymes is also heat labile, as judged by the
downshift of the apparent optimal temperature of activity (Feller et al., 1992)].

# CHAPTER II # 27
2.4.1. Structural adaptations at the active site
To date, five crystal structures of bacterial psychrophilic enzymes have been
solved by X-ray crystallography. These include the bacterial α-amylase (Aghajari et al.,
1998), malate dehydrogenase (Kim et al., 1999), citrate synthase (Russell, 1998), triose-
phosphate isomerase (Alveraz et al., 1998) and the fish trypsin (Smalas et al., 1994).
Unfortunately, only these limited data of X-ray structures have been compared with
mesophilic or thermophilic homologues and analyzed in detail with regards to the
structural parameters related to cold activity. Nevertheless, this limited set of data has
already provided valuable insights into the molecular basis of cold adaptation (Russell,
2000; Schroder et al., 2000; Leiros et al., 1999; Gianese et al., 2002).
As far as the active site of these psychrophilic enzymes is concerned, all reactive
side chains as well as most side chains pointing towards the catalytic cavity are strictly
conserved. This means that the overall catalytic mechanism and reaction pathway are not
modified in cold-active enzymes. This was demonstrated by the X-ray structure solved at
high resolution of both the cold-active α-amylase and of its closest structural homologue
from pig (Qian et al., 1994; Aghajari et al., 2002).
Two other types of molecular adaptation have been shown by X-ray structures.
Frequently, the catalytic cavity seems to be larger and more accessible to ligands in
psychrophilic enzymes than in mesophilic enzymes (Russell, 1998; Aghajari, 2003). This
is achieved by the deletion of residues in loops bordering the active site, by the distinct
conformation of these loops or by the replacement of bulky side chains with smaller
groups at the entrance to the active site. This improved accessibility is thought not only to

# CHAPTER II # 28
be responsible for the accommodation of the substrate at low energy cost, but also to
facilitate the release and exit of the reaction products. In some enzymes, the electrostatic
potential around the active-site region is also improved, so as to attract the oppositely
charged ligand and channel the substrate towards the catalytic cavity (Russell, 1998;
Aghajari, 2003; Kim et al., 1999; Smalas et al., 1994; Brandsdal et al., 2001). As a result
of the low temperature of unfolding, which prevents aggregation, psychrophilic enzymes
frequently show a high degree of unfolding reversibility that allows analysis of their
stability in the context of equilibrium thermodynamics (Feller et al., 1999). Such an
analysis is illustrated in Figure 2.4 by the stability curves of psychrophilic, mesophilic
and thermophilic proteins.
Figure 2.4. Gibbs free energy of denaturation of a psychrophilic α-amylase from
Antarctic Pseudoalteromonas haloplanktis (blue line), of mesophilic proteins (yellow
lines) and of one thermophilic protein (red line). ΔG is given in specific units (cal mol–1
of residues). By increasing order of melting temperatures (Tm) from left to right: the

# CHAPTER II # 29
psychrophilic Pseudoalteromonas haloplanktis α-amylase, the mesophilic proteins T4
lysozyme, barnase, Rnase T1, Rnase A, spectrin SH3, barstar, phosphocarrier HPr,
chymotrypsin inhibitor ACI2, Protein G IgG binding domain, ovomucoid third domain,
thioredoxin, ubiquitin and the thermophilic protein Sso7d (Feller et al., 1999).
2.4.2. Structural alterations create low stability
All the structural factors that stabilize a protein molecule can be attenuated in
both strength and number in psychrophilic enzymes (Smalas et al., 2000; Russell, 2000).
The number of proline and arginine residues (which restrict backbone rotations and can
form multiple hydrogen bonds and salt bridges, respectively) is reduced, whereas clusters
of glycine residues (which essentially have no side chain) provide localized chain
mobility. All weak interactions (ion pairs, aromatic interactions, hydrogen bonds and
helix dipoles) are less abundant, and non-polar core clusters have a weaker
hydrophobicity, making the protein interior less compact. Frequently, stabilizing
cofactors bind weakly, and loose or relaxed protein extremities seem to favor unzipping.
The protein surface is generally characterized by the disappearance of several solvent
exposed ion pairs, the exposure of a higher proportion of non-polar groups to the
surrounding medium (an entropy-driven destabilizing factor) and an excess of negative
charges that favor interactions with the solvent. These factors are thought to improve the
resilience or the breathing of the external shell. In multimeric enzymes, the cohesion
between monomers is also reduced by decreasing the number and strength of interactions
that are involved in association (Bell, 2002). However, each protein family adopts its own
strategy to decrease stability by using one or a combination of these structural alterations

# CHAPTER II # 30
(Leiros et al., 1999; Schroder-Leiros et al., 2000; Gianese et al., 2001; Gianese et al.,
2002).
2.4.3. Kinetic optimization of cold-active enzymes
Improving the turnover number kcat (catalytic constant; kcat is the maximal
enzyme reaction rate at a given temperature, which is expressed as the number of
substrate molecules that are transformed by one molecule of enzyme per unit of time) is
the main physiological adaptation, because it offsets the inhibitory effect of low
temperatures on reaction rates and therefore provides adequate raw metabolic activity to
the growing organism. However, both kcat and Km is the substrate concentration that is
required to produce 50% of the maximal activity. The specificity constant kcat/Km is
generally a better indication of catalytic efficiency than kcat alone. In principle, cold-
adapted enzymes can optimize the kcat/Km ratio by increasing kcat, decreasing Km or by
changes in both kcat and Km (Hoyoux et al., 2004).
2.4.4. Structure of Antarctic psychrophile Alteromonas haloplanctis α-amylase
The cold-active α-amylase from the Antarctic psychrophile, Alteromonas
haloplanctis has been studied extensively (Feller et al., 1992). The enzyme has a
molecular mass of 49 kDa with few salt bridges, aromatic interactions, small hydrophobic
cluster, few arginine residues and weak stabilization of helix dipoles. It is the first cold-
active α-amylase, which has been successfully crystallized and the 3-D structure resolved
at 1.85 Å (Aghajari et al., 1996). Also, this α-amylase was successfully expressed in
mesophilic host E. coli preserving genuine properties of a psychrophilic enzyme (Feller
et al., 1998).

# CHAPTER II # 31
The overall fold of A. haloplanctis α-amylase is very similar to those reported for
mesophilic α-amylases (Aghajari et al., 1998). Three characteristic domains as well as
ion-binding sites are found: domain A (residues 1-86 and 147-356); the central N-
terminal domain with a (β/α)8-barrel fold; a minor domain B (residues 87-146, an
insertion between α3 and β3) that protrudes from domain A and comprises a loop
structure, short β-strands and a short α-helix; and the C-terminal domain C consisting of
eight β-strands that form a Greek-key motif (the number of β-strands in other α-amylases
varies from five in barley to ten in human salivary). The largest variations in primary
structures between these enzymes from different species have been found in domain C
(Jespersen et al., 1993; MacGregor, 1988) and domain B (Janecek et al., 1997), but it
should also be mentioned that, throughout the α-amylase family, only eight residues are
invariant in the (β/α)8 barrel (Svendsen, 1994). These include seven residues at the active
site and a structurally important glycine. As in the mammalian α-amylases, binding sites
for calcium and chloride ions have been located in the structure of A. haloplanctis α-
amylase (Figure 2.5 a,b) (Aghajari et al., 1998).

# CHAPTER II # 32
Figure 2.5 (a) Figure 2.5 (b)
Figure 2.5(a). The three dimensional structure of amylase from Aspergillus sp.
(Matsuura et al., 1984)
Figure 2.5(b). Overall structure of psychrophilic AHA with residues (Gln 58 and Ala 99)
that replace the cysteines involved in the disulfide bridge between domains A and B in
MAA highlighted (in the upper right part of the figure). Domain A is colored in cyan,
domain B in pink and domain C in blue. The active site with the three catalytic amino
acids is shown, as are the calcium ion (yellow sphere) and chloride ion (green sphere)
(Aghajari et al., 1998). Several laboratories are now applying the techniques of gene
cloning and over expression (Table 2.2), followed by enzyme purification and
crystallization. Moreover draft genome sequences have been produced from two cold-
adapted Archaea: Methanogenium frigidum and Methanococcoides burtonii (Saunders et
al., 2003).

# CHAPTER II # 33
Table 2.2. Enzymes from psychrophilic bacteria, which have been cloned and
sequenced
Enzyme Bacterium Reference
Uracil-DNA glycosylase Psychrophilic strain
BMTU3346
Jaeger et al. (2000)
β-galactosidase Carnobacterium piscicola BA
Coombs and
Brenchley (1999)
Subtilisin (proteinase) Bacillus sp. TA39 Davail et al. (1994)
Lactate dehydrogenase Bacillus psychrosacchardyticus Vckovski et al.
(1990)
Esterase Pseudomonas sp. LS107d2 Mckay et al. (1992)
Isocitrate dehydrogenase Vibrio sp. ABE-1 Ishii et al. (1993)
β-Galactosidase Arthrobacter sp. B7 Gutshall et al.
(1995)
3-Isopropylmalate
dehydrogenase
Vibrio sp. 15 Wallon et al. (1997)
Alcohol dehydrogenase Moraxella sp. TAE 123 Tsigos et al. (1998)
Citrate synthase Arthrobacter sp. DS2-3R Gerike et al. (1997)
Malate dehydrogenase Aquaspirillium articum Kim et al. (1999)
Elongation factor 2 Methanococcus burtonii Thomas and
Cavicchioli (1998)
α-Amylase Alteromonas haloplanctis Feller et al. (1992)

# CHAPTER II # 34
Lipase* Moraxella sp. TA144
Psychrobacler immobilis
Pseudomonas sp. B11-1
Feller et al. (1991)
Arpigny et al. (1993)
Choo et al. (1998)
Triosephosphate
isomerase
Moraxella sp. TA137 Rentier et al. (1993)
β -Lactamase Psychrobacter immobilis Feller et al. (1996)
* Three isozymes were cloned
2.5. OPTIMIZATION PRODUCTION OF COLD-ACTIVE ALPHA-
AMYLASE ENZYME
Cold-active amylases are mostly extracellular and are highly influenced by
nutritional and physicochemical factors such as temperature, agitation, pH, nitrogen
source, carbon source, inducers, inorganic sources and dissolved oxygen. To meet the
demand of industries, low-cost medium is required for the production of α-amylase. Both
solid state fermentation (SSF) and submerged fermentation (SmF) could be used for the
production of α-amylases, although traditionally these have been obtained from
submerged cultures because of ease of handling and greater control of environmental
factors such as temperature and pH. SSF has been used for long to convert moist
agricultural polymeric substrates such as wheat, rice, soy, cassava, etc. (Table 2.3) into
fermented food products including industrial enzymes (Pandey et al., 1995). Solid state
fermentation is generally defined as the growth of microorganisms on moist solid
substrates with negligible free water (Selvakumar et al., 1998). The solid substrate may

# CHAPTER II # 35
provide only support or both support and nutrition. SSF constitutes an interesting
alternative since the metabolites so produced are concentrated and purification
procedures are less costly (Pandey et al., 1995; Benjamin and Pandey, 1997; Pandey,
1991). SSF is preferred to SmF because of simple technique, low capital investment,
lower levels of catabolite repression and end-product inhibition, low waste water output,
better product recovery, and high quality production (Zadrazil and Puniya, 1995). Among
the different substrates used for SSF, wheat bran has been reported to produce promising
results (Duenas et al., 1995; Muniswaran et al., 1994).
Table 2.3. Agro substrates used for cold-active α-amylase production
Substrate Organism Activity Reference
Wheat bran Paenibacillus
amylolyticus
275.95
(U/g/min)
Haq et al. (2012)
Wheat bran Aspergillus flavus, F2Mbb 119.1 (U/mg/ml) Sidkey et al. (2010)
Brown Lentils Bacillus sp. B4M bl 89 (U/mg/ml) Sidkey et al. (2010)
Maize meal Penicillium sp. 137 (U/g) Gouda and Elbahloul (2008)
Wheat bran:
Groundnut oil
cake (1:1)
Bacillus
amyloliquefaciens ATCC
23842
62470 (U/g) Gangadharan et al. (2006)
Wheat bran Bacillus sp. PS-7 464000 (U/g) Sodhi et al. (2005)
Rice bran Bacillus sp. PS-7 145000 (U/g) Sodhi et al. (2005)

# CHAPTER II # 36
2.5.1. Temperature
The influence of temperature on amylase production is related to the growth of
the organism. Hence, the optimum temperature depends on whether the culture is
mesophilic, thermophilic or psychrophilic. Among the fungi, most amylase production
studies have been done with mesophilic fungi within the temperature range of 25–37°C
(Francis et al., 2003; Ramachandran et al., 2004). Bacterial α-amylases are produced at a
Corn bran Bacillus sp. PS-7 97600 (U/g) Sodhi et al. (2005)
Coconut oil
cake
A. oryzae 3388 (U/g) Ramachandran et al. (2004)
Spent brewing
grain
A. oryzae NRRL 6270 6583 (U/g) Francis et al. (2003)
Rice husk B. subtilis 21 760 (U/g) Baysal et al. (2003)
Amaranthus
grains
Aspergillus flavus 1920 (U/g) Vishwanathan & Surlikar
(2001)
Maize bran B. coagulans 22956 (U/g) Babu and Satyanarayana
(1995)
Mustard oil
cake
B. coagulans 5953 (U/g) Babu and Satyanarayana
(1995)
Gram bran B. coagulans 8984 (U/g) Babu and Satyanarayana
(1995)

# CHAPTER II # 37
much wider range of temperature. B. amyloliquefaciens, B. subtilis, B. licheniformis and
B. stearothermophilus are among the most commonly used Bacillus sp. reported to
produce α-amylase at temperatures 37-60°C (Syu and Chen, 1997; Mishra et al., 2005;
Mendu et al., 2005; Mielenz, 1983). A cold-active α-amylase from Antarctic
psychrophile Alteromonas haloplanktis was reported to exhibit maximum α-amylase
production at 4°C (Feller et al., 1998).
2.5.2. pH
pH is also one of the important factors that determine the growth and morphology
of microorganisms as they are sensitive to the concentration of hydrogen ions present in
the medium. Earlier studies have revealed that fungi required slightly acidic pH and
bacteria required neutral pH for optimum growth. pH is known to affect the synthesis and
secretion of α-amylase just like its stability (Fogarty, 1983). Bacterial cultures such as B.
subtilis, B. licheniformis and B. amyloliquefaciens required an initial pH of 7.0 (Syu and
Chen, 1997; Tanyildizi et al., 2005; Haq et al., 2005). Rhodothermus marinus was
reported to yield good enzyme levels at initial pH range 7.5 to 8.0 (Gomes et al., 2003).
Hyperthermophilic archae such as Pyrococcus furiosus, P. woesei and Thermococcus
profundus yielded optimum α-amylase at pH 5.0 (Vieille and Zeikus, 2001).
Thermophilic anaerobic bacteria Clostridium thermosulfurogenes gave maximum titres of
α-amylase at pH 7.0 (Swamy and Seenayya, 1996). A list of various cold active α-
amylase producing psychrophillic and psychrotrophic bacteria and their production
parameters are presented in Table 2.4.

# CHAPTER II # 38
Table 2.4. Production parameters for cold-active α-amylase
Microbes Incubation
period (hr)
Optimum
temp.(°C)
Optimum
pH
Reference
Bacillus cereus GA6 96 20 10.0 Kuddus et al. (2012)
Microbacterium
foliorum GA2
120 20 9.0 Roohi et al. (2011)
Wangia sp. C52 NM 20 7.18 Liu et al. (2011a)
Pseudoalteromonas
arctica GS230
24 20 8.0 Lu et al. (2010)
Nocardiopsis aegyptia 48 25 5.0 Abou-Elela et al.
(2009)
Streptomyces sp. 1 20 NM Cotarlet et al. (2009)
Micrococcus
antarcticus
64 12 8.0 Fan et al. (2009)
Lactobacillus
plantarum
36 35 7.0 Smita et al. (2008)
Penicillium sp. 48 30 4.0 Gouda and Elbahloul
(2008)
Nocardiopsis sp. 7326 NM 20 8.0 Zhang and Zeng (2008)
P. amylolyticus 0.5 35 6.0 Xiaohong et al. (2007)
Arthrobacter
psychrolactophilus
NM 22 NM Michael et al. (2005)

# CHAPTER II # 39
NM: not mentioned
2.5.3. Carbon and Nitrogen sources
These are necessary for the growth and metabolism of organisms. Various carbon
sources are tried to optimize the maximum production of cold-active α-amylase for
different bacterial species. For enhanced growth and metabolism of organisms, nitrogen
also play very important role just like carbon. A large variety of carbon and nitrogen
sources for bacterial species are available which give rise to maximum production of
cold-active α- amylase (Table 2.5 and 2.6).
Aspergillus oryzae NM 4 NM Akpan & Adelaja
(2004)
Penicillum sp. FS
010441
NM 15 6.0 Zhang et al. (2002)
Aspergillus ochraceus 48 30 5 Nahas and Mirela
(2002)
Flavobacterium
balustinum A201
NM 30 NM Morita et al. (1997)
Bacillus sp. A-001 NM 35 7.5 Lealem and Gashe
(1994)

# CHAPTER II # 40
Table 2.5. Carbon sources used for maximum cold-adapted α-amylase production
Bacterial species Best C-source Reference
Bacillus cereus GA6 Glycerol Kuddus et al. (2012)
Microbacterium foliorum GA2 Lactose Roohi et al. (2011)
Pseudoalteromonas arctica GS230 Soluble starch Lu et al. (2010)
Bacillus sp. strain TSCVKK Dextrin Kiran and Chandra (2008)
Bacillus sp.1 Starch Varalakshmi et al. (2008)
Nocardiopsis aegyptia Starch Abou-Elela et al. (2009)
B. subtilis IMG22 Starch and Glycerol Sodhi et al. (2005)
Bacillus sp. PS-7 Starch and Glycerol Tanyildizi et al. (2005)
Bacillus sp. I-3 Starch and Glycerol Goyal et al. (2005)
Bacillus sp. Lactose Hamilton et al. (1999)
B. stearothermophilus Soluble starch Srivastava & Baruah (1986)
B. licheniformis Galactose, glycogen and inulin Chandra et al. (1980)

# CHAPTER II # 41
Table 2.6. Nitrogen sources used for maximum cold-active α-amylase production
Bacterial species Best N-source Reference
Bacillus cereus GA6 Ammonium acetate Kuddus et al. (2012)
Microbacterium foliorum GA2 Yeast extract Roohi et al. (2011)
Wangia sp. C52 Yeast extract Liu et al. (2011a)
Pseudoalteromonas arctica
GS230
Beef extract Lu et al. (2010)
Pseudoalteromonas
haloplanktis
Glutamic acid Wilmes et al. (2010)
Nocardiopsis aegyptia Potassium nitrate Abou-Elela et al. (2009)
Bacillus sp. strain TSCVKK Yeast extract Kiran and Chandra (2008)
Bacillus sp. I-3 Soya bean meal Sodhi et al. (2005) and
Francis et al. (2003)
B. licheniformis SPT 278 Peptone Aiyer (2004)
A. oryzae A1560 Casein hydrolysate Pederson and Nielson
(2000)
B. amylolyticus Peptone and yeast extract Dettori et al. (1992)

# CHAPTER II # 42
2.5.4. Other fermentative conditions
Surfactants in the fermentation medium are known to increase the production of
extracellular amylase enzymes by increasing cell membrane permeability. Some common
surfactants are Tween 80, polyethylene glycols, Cholic acid which is used in different
concentrations. Supplementation of salts of certain metal ions provided good growth of
microorganisms and thereby better enzyme production (as most α-amylases are known to
be metalloenzymes). Some frequently used metal ions are CaCl2, NaCl, LiSO4, MgSO4,
FeCl3, Mn2+, Zn2+, etc. In SSF system some additional fermentative conditions play very
vital role viz selection of a suitable substrate and microorganism; pre-treatment of the
substrate; particle size (inter-particle space and surface area) of the substrate; water
content; relative humidity; type and size of the inoculum; removal of metabolic heat;
period of cultivation; maintenance of uniformity in the environment of SSF system, and
the gaseous atmosphere, i.e. oxygen consumption rate and carbon dioxide evolution rate.
2.6. COLD-ACTIVE AMYLASES: PURIFICATION AND
CHARACTERIRIZATION
Industrial enzymes produced in bulk generally require little downstream
processing and hence are relatively crude preparations. The commercial use of α-amylase
generally does not require purification of the enzyme, but enzyme applications in
pharmaceutical and clinical sectors require high purity amylases. The enzyme in purified
form is also a prerequisite in studies of structure-function relationships and biochemical
# CHAPTER II # 43
properties. The purification of α-amylases from microbial sources in most cases has
involved classical purification methods. These methods involve separation of the culture
from the fermentation broth, selective concentration by precipitation using ammonium
sulfate or organic solvents such as chilled acetone. The crude enzyme is then subjected to
chromatography, usually affinity, ion exchange and/or gel filtration. The need for large-
scale cost effective purification of proteins has resulted in evolution of techniques that
provide fast, efficient and economical protocols in fewer processing steps (Amritkar et
al., 2004). Purification techniques that produce homogeneous preparation of α-amylases
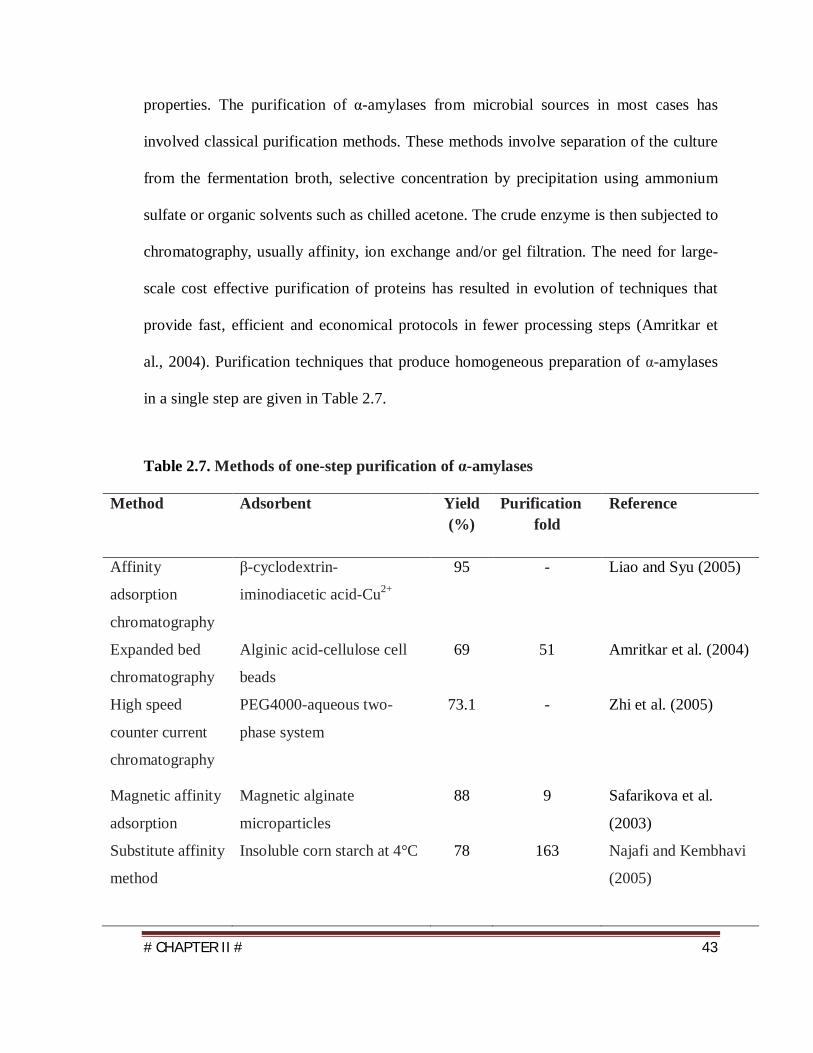
in a single step are given in Table 2.7.
Table 2.7. Methods of one-step purification of α-amylases
Method
Adsorbent Yield (%)
Purification fold
Reference
Affinity
adsorption
chromatography
β-cyclodextrin-
iminodiacetic acid-Cu2+
95 - Liao and Syu (2005)
Expanded bed
chromatography
Alginic acid-cellulose cell
beads
69 51 Amritkar et al. (2004)
High speed
counter current
chromatography
PEG4000-aqueous two-
phase system
73.1 - Zhi et al. (2005)
Magnetic affinity
adsorption
Magnetic alginate
microparticles
88 9 Safarikova et al.
(2003)
Substitute affinity
method
Insoluble corn starch at 4°C 78 163 Najafi and Kembhavi
(2005)

# CHAPTER II # 44
Ultrafiltration α-Cyclodextrin coupled with
Sepharose 6B (pH 7.0)
78 140 Iefuji et al. (1996)
Gel filtration
chromatography
Sephacryl-S200 HR 17 9 Marco et al. (1996)
Ion exchange
chromatography
DEAE-Cellulose DE52 (pH
5.3)
66 33 Morgan and Priest
(1981)
Ultrafiltration Cellulose 35 20 Giraud et al. (1993)
Mono Q
adsorption
chromatography
Sephadex G-25 (pH 7.5 50 6.9 Freer (1993)
Drum humidifier
chromatography
25% (NH4)2SO4, 70%
acetone
- 1.036 Robyt and Ackerman
(1971)
The pH optima of α-amylases vary from 2.0 to 12.0 (Vihinen and Mantsala, 1989).
Alpha-amylases from most bacteria and fungi have pH optima in the acidic to neutral
range (Pandey et al., 2000). Alpha-amylases from Alicyclobacillus acidocaldarius
showed an acidic pH optima of 3.0 (Schwermann et al., 1994), in contrast to the alkaline
amylase with optima of pH 9.0-10.5 as reported from an alkalophilic Bacillus sp. (Saito,
1973; Krishnan and Chandra, 1983; Lee et al., 1994; Shinke et al., 1996). Extremely
alkaliphilic α-amylase with pH optima of 11.0-12.0 has been reported from Bacillus sp.
GM8901 (Kim et al., 1995). In some cases, the pH optimum was observed to be
dependent upon temperature as in the case of Bacillus stearothermophilus DONK BS-1
(Ogasahara et al., 1970) and on calcium as in the case of B. stearothermophilus (Pfueller
and Elliott, 1969). Alpha-amylases are generally stable over a wide range of pH from 4.0
to 11.0 (Fogarty and Kelly, 1979; Vihinen and Mantsala, 1989; Hamilton et al., 1999a;

# CHAPTER II # 45
Hamilton et al., 1999b; Saito, 1973; Khoo et al., 1994), however, α-amylases with
stability in a narrow range have also been reported (Coronado et al., 2000; Krishnan and
Chandra, 1983; Robyt and Ackerman, 1971).
The optimal activity temperature of Nocardiopsis sp. 7326 amylase was 35°C,
and the enzyme was stable between pH 5.0 and 10.0 with maximal activity at pH 8.0
(Zhang and Zeng, 2008). The activity of α-amylase from Streptococcus bovis JB1 was
optimal at pH 5.0 to 6.0. The enzyme was relatively stable at temperatures below 50°C
(Freer 1993). Ueda and his coworkers (2008) purified and characterized the novel cold-
adapted α-amylase (Amy I and Amy II) which are most active at pH 5.5 and stable at pH
7.0-9.0. Both Amy I and Amy II exhibited activities at 10°C. A cold-active α-amylase
from Antarctic psychrophile Alteromonas haloplanktis was reported to exhibit maximum
α-amylase production at 4°C (Feller et al. 1998).
The optimal temperature and pH for the purified amylase from Micrococcus
antarcticus were 30°C and 6.0, respectively (Fan et al. 2009). It still showed high activity
at low temperature 10-15°C. It was sensitive to high temperature but was stable at pH
6.0-10.0 with at least 70% activity remained. Liu et al. (2011b) reported a 58 kDa cold-
active amylase from marine Wangia sp. C52 having an optimum pH and temperature of
6.0 and 30°C, also in the presence of Ca2+ and Co2+, the enzyme activity was stimulated
while Cu2+, Hg2+, Mn2+, Zn2+, Fe3+, Al3+, EDTA, EGTA and SDS reduced the activity.
Km and Vmax values of the purified enzyme for soluble starch were 2.08±0.3 mg/ml and
1.26±0.02 mg/ml/min, respectively. Nouadri et al. (2010) purified 60.5 kDa α-amylase
from Penicillium camemberti PL21 with specific activity of 154.2 units/ml/mg protein
with 38.5 folds purification. The optimum substrate concentration for soluble starch was

# CHAPTER II # 46
1% (w/v) at the optimum temperature of 30ºC. The purified α-amylase enzyme had a
maximum activity at pH 6.0 and the Km value for soluble starch was found to be 0.92
mg/ml.
Alpha-amylase from Alteromonas sp. TAC 240B collected form Antarctic sea water
showed a molecular mass of about 50 kDa and a pI of 5.2. The enzyme is stable from pH
7.5 to 9.0 and has a maximal activity at pH 7.5. This psychrophilic alpha-amylase
requires both Cl- and Ca2+ for its amylolytic activity. Br- is also quite efficient as an
allosteric effector. The comparison of the amino acid composition with those of other
alpha-amylases from various organisms shows that the cold-active alpha-amylase has the
lowest content of Arg and Pro residues (Chessa et al., 1999). Psychrophilic amylase
produced by Penicillum sp. FS010441 isolated from sea mud of the Yellow Sea and the
East China Sea. The optimum growth temperature and pH for this α-amylase enzyme is
15°C and 6.0, respectively and even at 0°C, the enzyme shows some activities (Zhang et
al., 2002). A cold-active amylase producing P. amylolyticus strain showed an optimum
temperature of 35°C and pH 6.0 when incubated for 30 hours. This amylase was not
stable when the temperature was above 40°C (Xiaohong et al., 2007). Alpha-amylases
from B. subtilis, B. amyloliquefaciens I and B. amyloliquefaciens II were strongly
inhibited by Zn2+, Ag+, Cu2+ and Fe2+ (Elif and Velittin, 2000). However, the activity of
cold-adapted Nocardiopsis sp. 7326 amylase was not affected by Zn2+, Ni2+ and Fe2+, but
was even activated by Cd2+ and Cu2+ (Zhang and Zeng, 2008).

# CHAPTER II # 47
2.7. BIOTECHNOLOGICAL ASPECTS OF COLD ACTIVE ALPHA-
AMYLASE
An emerging area of research in the field of enzymology is to develop radically
different and novel biocatalysts through various molecular approaches including
recombinant DNA technology, protein engineering, directed evolution and the
metagenomic approach. As a whole, amylase biotechnology has just reached the end of
lag phase and the beginning of the exponential phase: it demands extension in terms of
both quality and quantity. Qualitative improvements in restructuring amylase gene and its
protein can be achieved by employing already established recombinant DNA technology
and protein engineering. Quantitative enhancement needs strain improvement, especially
through site-directed mutagenesis and standardizing the nutrient medium for the
overproduction of cold-active α-amylases.
2.7.1. Gene cloning
To date, a very few number of cold-active α-amylase genes were isolated and the
related studies have been carried out. Early successes in the production of heterologous
proteins were achieved using Escherichia coli as host and various kinds of proteins were
expressed in E. coli. However, expression of eukaryotic proteins in E. coli became very
difficult due to formation of inclusion bodies, protein misfolding and safety issues. Other
expression systems were developed among yeasts, fungi, plants and animals. Cloning and
expression of the cold-active amylase gene from Alteromonas haloplanctis has been
reported (Feller et al., 1992, 1998).

# CHAPTER II # 48
2.7.2. Protein engineering
Psychrophilic organisms and their enzymes have, in recent years, increasingly
attracted the attention of the scientific community due to their peculiar properties that
render them particularly useful in investigating the possible relationship existing between
stability, flexibility and specific activity and as valuable tools for biotechnological
purposes. Although α-amylase carry significant commercial value, biotechnologically
produced or engineered cold-active α-amylases may represent the focus of industrial
interest in future. Cold-active α-amylases could generate avenues for industrial
applications, once their specific properties are improved through enzyme engineering.
Determination of three dimensional structures of more cold-active amylases would allow
the detailed analysis of protein adaptation to temperatures at molecular level. This may
include increased thermolabile nature and/or catalytic activity at low temperatures, or the
modification of pH profiles. Cold-active α-amylases from microorganisms retaining high
catalytic activity at low temperatures are successfully produced using site directed
mutagenesis and directed evolution. Alpha-amylase from the Antarctic psychrophile
Alteromonas haloplanctis is synthesized at 0±2°C by the wild strain. This heat-labile α-
amylase folds correctly when over expressed in Escherichia coli, providing the culture
temperature is sufficiently low to avoid irreversible denaturation (Feller et al., 1998). The
thermal stability of the cold-active α-amylase (AHA) secreted by the Antarctic bacterium
Alteromonas haloplanctis has been investigated by intrinsic fluorescence, circular
dichroism and differential scanning calorimetry. It was found that this heat-labile enzyme
is the largest known multidomain protein exhibiting a reversible two-state unfolding
(Feller et al., 1998).

# CHAPTER II # 49
2.8. BIOTECHNOLOGICAL POTENTIAL OF PSYCHRO-
TOLERANT ALPHA-AMYLASES
Amylases are among the most important hydrolytic enzymes for all starch based
industries and the commercialization of amylases is oldest with first use in 1984, as a
pharmaceutical aid for the treatment of digestive disorders. In the present day scenario,
amylases find application in all the industrial processes such as in food, detergents,
textiles and in pulp-paper industry, for the hydrolysis of starch, etc. In this light,
microbial amylases have completely replaced chemical hydrolysis in the starch
processing industry. They can also be of potential use in the pharmaceutical and fine
chemical industries. Today, amylases have the major world market share of enzymes
(Aehle and Misset, 1999). Several different amylase preparations are available with
various enzyme manufacturers for specific use in varied industries.
2.8.1. Detergent applications
Dirt and stains on clothes comes in many forms and includes proteins, starches
and lipids. In addition, clothes that have been starched must be freed of the starch. Using
detergents in water at high temperatures and with vigorous mixing, it is possible to
remove most types of dirt and stains but the cost of heating the water is high and lengthy
mixing or beating will shorten the life of clothing and other materials. The use of
enzymes allows lower temperatures to be employed and shorter periods of agitation are
needed, often after a preliminary period of soaking. Psychrophilic enzymes can be very
useful for domestic processes as amylases; especially cold-active alkaline α-amylases can
be used in detergents since washing clothes at low temperatures protect the colors of

# CHAPTER II # 50
fabrics and reduce energy consumption. In general, enzymatic detergents remove stains
from clothes far more effectively than non-enzymatic detergents.
2.8.2. Bread and baking industry and as an antistaling agent
The baking industry has made use of these enzymes for hundreds of years to
manufacture a wide variety of high quality products. For decades, enzymes such as malt
and microbial α-amylases have been widely used in the baking industry (Hamer et al.,
1995; Si, 1999). These enzymes were used in bread and rolls to give these products a
higher volume, better color and a softer crumb. It is the malt preparation that has led the
way and opened the opportunities for many enzymes to be used commercially in baking.
Today, many enzyme preparations such as proteases, lipases, xylanases, pullulanases,
cellulases, glucose oxidases, lipoxygenases etc. are being used in the bread industry for
varied purposes (Kulp, 1993; Prieto et al., 1995; Si, 1999; Monfort et al., 1996), but none
had been able to replace α-amylases.
Till date, α-amylases used in baking have been cereal enzymes from barley malt
and microbial enzymes from fungi and bacteria (Hebeda et al., 1990, 1991). Fungal α-
amylases have been permitted as bread additives since 1955 in the US and in 1963 in UK
after confirmation of their GRAS status (Pritchard, 1992). Presently they are used all over
the world to different extents. Supplementation of flour with exogenous fungal α-amylase
having higher activities is common in the present day modern and continuous baking
process (Pritchard, 1992). Cold-active α-amylase supplementation in flour not only
enhances the rate of fermentation but also consume less amount of energy at industrial
scale and moreover reduces the viscosity of dough, resulting in improvements in the

# CHAPTER II # 51
volume and texture of the product, also generates additional sugar in the dough, which
improves the taste, crust color and toasting qualities of the bread (Van and Hille, 1992).
One of the new applications of α-amylase in the industry has been in retarding the
staling of baked products, which reduces the shelf life of these products. Upon storage the
crumb becomes dry and firm, the crust loses its crispness and the flavor of the bread
deteriorates. All these undesirable changes in the bread are together known as staling.
The importance of retrogradation of starch fraction in bread staling has been emphasized
(Kulp and Ponte, 1991). A loss of more than US $1 billion is incurred in USA alone
every year due to the staling of bread. Conventionally various additives are used to
prevent staling and improve the texture and flavor of baked products. Additives include
chemicals, small sugars, enzymes/their combinations, milk powder, emulsifiers,
monoglycerides/diglycerides, sugar esters, lecithin, etc; granulated fat, anti-oxidant
(ascorbic acid or potassium borate), sugars/salts (Spendler and Jorgensen, 1997).
Recently emphasis has been given to the use of enzymes in dough improvement as anti-
staling agents, e.g. α-amylase (Cole, 1982), branching enzymes (Okada et al., 1984) and
debranching enzymes (Carroll et al., 1987), maltogenic amylases (Oleson, 1991), β-
amylases (Wursch and Gumy, 1994) amyloglucosidases (Vidal and Gerrity, 1979).
Pullulanases and α-amylase combination are used for efficient antistaling property
(Carroll et al., 1987). However, a slight excess of α-amylases was also used which is
undesirable as it causes stickiness in bread (Oleson, 1991). Therefore, a recent trend is to
use intermediate temperature stable (ITS) α-amylases or better cold-active α-amylase
(Kulp, 1993; Hebeda et al., 1990 and 1991; Ahuja et al., 1998). They are active after
starch gelatinization and become inactive much before the completion of the baking

# CHAPTER II # 52
process. Further, the dextrin with 4-9 degree of polymerization produced by these shows
the anti-staling properties. Although a wide variety of microbial α-amylases is known, α-
amylase with ‘ITS’ property has been reported from only a few microorganisms (Prieto et
al., 1995; Monfort et al., 1996; Kraus and Hebeda, 1993; Gigras et al., 2002).
2.8.3. Manufacturing of maltose
Maltose is a disaccharide made up of glucose units. It is the main component of
malt-sugar syrup (Matsumoto et al., 1982). Maltose is commonly used as sweetener and
also as intravenous sugar supplement. It has a great value in food industries since it is
non-hygroscopic and does not easily crystallize. For the manufacturing of variety of
grade of maltose, starch (potato, sweet potato, corn and cassava starches are frequently
used) is hydrolyzed by the use of alpha-amylase. To reduce the time and energy
consumption and to increase the efficiency, cold-active α-amylase is better option. For
production of medical grade, the concentration of starch slurry is adjusted to be 10-20%
maltose and for food grade, 20-40% maltose is used.
2.8.4. Manufacturing of high fructose containing syrups
High fructose containing syrups 42 F (Fructose content, 42%) is prepared by
enzymatic isomerization of glucose with glucose isomerase. The starch is first converted
to glucose by α-amylase enzymatic liquefaction and saccharification.

# CHAPTER II # 53
2.8.5. Manufacturing of maltotetraose syrup
Maltotetraose syrup (G4 syrup) is produced by breaking of starch into
maltotetraose by the action of α-amylase enzyme. The sweetness of the syrup is as low as
20% of sucrose. Therefore in foods, G4 syrup can be successfully used in place of
sucrose which reduces the sweetness without altering their inherent taste and flavor. It
has high moisture retention power which maintains integrity of starch particles and
retains suitable moisture in foods. G4 syrup improves the food texture because of its high
viscosity than sucrose. It further lowers down the freezing point of water than sucrose or
high fructose syrup, so can be used to control the freezing points of frozen foods. G4
syrup imparts gloss and can be used in industry such as a paper sizer.
2.8.6. Manufacturing of high molecular weight branched dextrins
High molecular weight branched dextrins are used as extender for production of
powdery foods and a glazing agent for rice cakes. These are produced by the action of α-
amylase on corn starch. Degree of hydrolysis depends on the type of starch and the
physical properties desired. Branched dextrins can be collected as powder after
chromatography and spray drying.
2.8.7. Removal of starch sizer from textile (desizing)
Modern production processes for textiles introduce a considerable strain on the
warp during weaving. The yarn must, therefore, be prevented from breaking. For this
purpose a removable protective layer is applied to the threads. The materials that are used
for this size layer are quite different. Starch is a very attractive size, because it is cheap,
easily available in most regions of the world, and it can be removed quite easily. Good

# CHAPTER II # 54
desizing of starch sized textiles is achieved by the application of α-amylases, which
selectively remove the size and do not attack the fibers. It also randomly cleaves the
starch into dextrins that are water soluble and can be removed by washing. It also
prevents the loss of string by friction, cutting and generation of static electricity on the
string by giving softness to the surface of string due to laid down warp. The use of α-
amylases in warp sizing of textile fibres for manufacturing fibers with great strength has
been reported (Hendriksen et al., 1999).
2.8.8. Direct fermentation of starch to ethanol
For large-scale processing, the bioconversion of biomass resources, especially
starchy materials, to ethanol, is very useful because it can be used as a bio-fuel and as the
starting material for various chemicals. However, in the present scenario its cost of
production is very high because of three main reasons; ethanol production from starchy
materials via fermentation consists of two or three steps, large quantity of α-amylase is
needed and starchy materials need to be cooked at a high temperature (140° to 180°C).
Now methods are developed in which cost of production can be minimized; by
fermenting starch to ethanol in one step using co-cultures of two different strains and by
using low temperature-cooking fermentation systems (that succeeded in reducing energy
consumption by approximately 50%), but it is still necessary to add large amounts of
amylolytic enzymes to hydrolyze the starchy materials to glucose (Matsumoto et al.,
1982, 1985).

# CHAPTER II # 55
2.8.9. Treatment of starch containing waste water
Food processing wastewater offers a unique challenge to any treatment system.
Often containing multiple types of contaminants that pose serious threats to the ability of
a standard sewage treatment facility, causes pollution problem also. Biotechnological
treatment of food processing starch waste water by the action of α-amylase can produce
valuable products such as microbial biomass protein and also purifies the effluent at low
temperature (Aiyer, 2005).
2.8.10. Starch liquefaction and saccharification
The major market for α-amylases lies in the production of starch hydrolysates
such as glucose and fructose. Starch is converted into high fructose corn syrups (HFCS).
Because of their high sweetening property, these are used in huge quantities in the
beverage industry as sweeteners for soft drinks. The process requires the use of α-
amylase for starch liquefaction. The use of α-amyalse in starch liquefaction is well
established and has been extensively reviewed (Marrel et al., 2002; Pandey et al., 2000).
2.8.11. Paper and pulp industry
The use of α-amylase for the production of low viscosity, high molecular weight
starch for coating of paper is reported (Bruinenberg et al., 1996). The use of amylases in
the pulp and paper industry is in the modification of starches for coated paper. As for
textiles, sizing of paper is performed to protect the paper against mechanical damage
during processing. It also improves the quality of the finished paper. The size enhances
the stiffness and strength in paper. It also improves the erasibility and is a good coating

# CHAPTER II # 56
for the paper. Starch is also a good sizing agent for the finishing of paper. Starch is added
to the paper in the size press and paper picks up the starch by passing through two rollers
that transfer the starch slurry. A constant viscosity of the starch is required for
reproducible results at this stage. The mill also has the flexibility of varying the starch
viscosity for different paper grades. The viscosity of the natural starch is too high for
paper sizing and is adjusted by partially degrading the polymer with α-amylases in a
batch or continuous processes. The conditions depend upon the source of starch and the
type of α-amylase at different temperatures (Tolan, 1996).
2.8.12. Analysis in medicinal and clinical chemistry
With the advent of new frontiers in biotechnology, the spectrum of amylase
applications has expanded into many other fields, such as clinical, medicinal and
analytical chemistry. There are several processes in the medicinal and clinical areas that
involve the application of amylases. The application of a liquid stable reagent, based on
α-amylase for the Ciba Corning Express clinical chemistry system has been described
(Becks et al., 1995). A process for the detection of higher oligosaccharides, which
involved the application of amylase, was also developed (Giri et al., 1990). This method
was claimed to be more efficient than the silver nitrate test. Biosensors with an
electrolyte isolator semiconductor capacitor (EIS-CAP) transducer for process monitoring
were also developed (Menzal et al., 1998).

# CHAPTER II # 57
2.8.13. Other applications
In food industry cold-active α-amylase can be used for the reduction of haze
formation in juices. Psychrophilic microorganisms have also been proposed for the
bioremediation of polluted soils and waste waters during the winter in temperate
countries, when the degradative capacity of the endogenous microflora is impaired by
low temperatures. Glycosidases are often used in the baking industry, but can retain
residual activity after cooking that alters the structure of the final product during storage;
this can be avoided by the use of psychrophilic glycosidases. An important achievement
in the field has been the construction of a host-vector system that allows the over
expression of genes in psychrophilic bacteria (Tutino, 2001): expression at low
temperatures prevents the formation of inclusion bodies and protects heat-sensitive gene
products. Using enzymes having highest activity below 20°C in food processing, limits
the growth of other contaminating microorganisms, shorten the process times, and avoid
designing expensive heating steps. Cold-active α-amylases could be used in the brewing
industry to speed the mashing phase at low temperatures. The expansion of experimental
models to include plants, nematodes, some cold blooded animals such as fish and frogs,
and other microorganisms may create the need for enzymes with higher activities at
lower temperatures. In addition, reporter genes making cold-active enzymes would be
valuable additions to the arsenal of molecular tools.

# CHAPTER II # 58
2.9. Future prospects
The emerging picture of cold adaptation indicates that to catalyze biochemical
reactions at low temperatures, psychrophilic enzymes improve the flexibility of the
structural elements that are involved in the catalytic cycle, thereby resulting in an activity
that is markedly heat labile. The high flexibility of cold-active enzymes is strongly
supported by experiments using dynamic quenching of fluorescence that show increased
permeability to a small quencher molecule (D’Amico et al., 2003; Collins et al., 2003;
Georlette et al., 2003). Obviously, the next challenge will be to define this flexibility in
terms of the type, amplitude and timescale of molecular motions. In fact, the intricate
relationships between these factors and activity or stability are still poorly understood,
not only in psychrophilic enzymes but also in proteins and enzymes in general (Jaenicke,
2000; Hernandez et al., 2000; Tehei et al., 2001). As a result of their biophysical
peculiarities, psychrophilic enzymes are interesting and useful models in protein research
for protein evolution, folding and dynamic studies.





























