Embed Size (px)
Citation preview

76
Chapter-2
Micropropagation and Large-
scale Culture of C. borivilianum in Bioreactor

77
2.1 INTRODUCTION
Multiplication of genetically identical copies of plant species by method of asexual
reproduction is called “clonal propagation” and plant population derived from a single
individual by asexual reproduction constitutes a “clone”. In nature, clonal propagation
occurs by apomixis (seed development without meiosis and fertilization) and/or vegetative
reproduction (regeneration of new plants from vegetative parts). Apomixis being restricted
to only few species, the method of vegetative propagation of plants has been practiced for
centuries through improved conventional methods. Plant tissue culture has become a
popular method for vegetative propagation of plants. Large scale clonal propagation of
plants popularly known and described as “micropropagation” is the first major practical
application of plant biotechnology. The beginning of this technique based on certain
important discoveries dates back to about five to six decades. Morel and Martin (1952) for
the first time recovered virus free Dahlia plants from infected individuals by excising and
culturing their shoot tips in vitro. Morel (1960) also realized the potential of this method
for rapid propagation of these plants. This contribution of Morel not only revolutionized
the orchid industry, but also gave impetus to the utilization of shoot-bud culture for rapid
cloning of other plant species.
Skoog and Miller (1957) put forth the concept of hormonal control of organ
formation. They regarded organ formation to be determined by quantitative interaction, i.e.
ratios rather than absolute concentration of phytohormones participating in growth and
development. This concept of hormonal regulation of organogenesis is now applicable to a
large number of plant species. They discovered the role of plant hormones, especially of
cytokinins in shoot morphogenesis. The elucidation of the role of cytokinins in apical
dominance inhibition, which subsequently results in the release of axillary meristems from
dormancy, was another major break-through in this field (Sachs and Thimann, 1964). The
practical and applied use of plant cell, tissue and organ culture techniques has developed
tremendously since Vasil and Hildebrandt (1965) demonstrated that whole plant can be
regenerated from single cell of tobacco cultured in an in vitro environment. Murashige
(1974, 1977, 1980) was instrumental in giving the technique of in vitro culture a status of
a viable approach to the propagation of many plant species. He worked extensively for the
popularization of technique developing standard methods for in vitro propagation of
several species ranging from ferns to foliage, flower and fruit plants. Indeed the
Murashige’s name became intimately associated with the technique.

78
The successful application of these developments to the in vitro multiplication of
plants by micropropagation has been a key factor in the refinement and development of
this technology into an important worldwide industry. Practical documentation of
morphogenetic and biochemical totipotency in varied plant species e.g. ornamental,
horticultural and medicinal crops has pushed micropropagation to the realm of commercial
application that itself has got further strengthened with the advancements made in the
frontline areas of molecular biology and genetic engineering. Plant tissue culture
techniques now occupy a core sector position in overall plant biotechnology domain.
Globally the annual throughput of commercial micropropagation efforts is about 500
million propagules including ornamentals, fruits and forest plants. The requirement for
ornamental plant species alone is estimated to be around $2 billion with a net turnover of $
50 billion by the year 2010.
The micropropagation technology is simple involving the culture of isolated
shoots/shoot bases/shoot meristems on suitable nutrient media to stimulate the
development of multiple axillary buds by overcoming apical dominance. Generally three
to six-fold increase in the shoot numbers at regular 4-6 weeks interval results in the
production of a large number of plants from each cultured shoot meristem in one year
(Hussey, 1978; Walkey, 1978; Pierik, 1979). The simplicity of the methodology has
allowed development of hundreds of small and large industries in many countries around
the world.
There are two other methods of in vitro propagation also. These are based on the
de novo formation of shoot meristems in callus cultures or induction of somatic embryos
from somatic cells, these phenomenon generally referred to as indirect regeneration (Crisp
and Walkey, 1974; Murashige, 1974; de Fossard, 1977; Conger, 1981). Differentiation of
plant cells from callus cultures has been suggested as a potential method for rapid
propagation of selected plant species because hundreds and thousands of plants can be
raised from a small amount of tissue and in a continuous process. Wherever applicable,
this is often the fastest method of multiplication. However, there are few drawbacks in this
procedure and as far as possible it should be avoided in clonal propagation of any specific
cultivar. One of the most serious drawback for shoot multiplication is the genetic
instability in long term cultures resulting in high incidences of genetically variant plants
and even chimeras. Therefore, the plants propagated through this method are not the true
copies of the original resource plant due to epigenetic effects and genetic differences
arising due to genes/chromosomal changes. These variations are called “somaclonal

79
variations”. The other method of in vitro plant propagation is based on the induction of
somatic embryogenesis (Vasil and Vasil, 1991). Somatic embryogenesis i.e., the formation
of embryo like structures from somatic cells and their germination into complete plants
(Bhaskaran and Smith, 1990) has now been proved as potentially most efficient and
economic method for the large scale clonal propagation of plants. It is a versatile
technique for rapid multiplication of plants and offer a superior possibility for developing
scale-up technology as compared to organogenesis.
The distinction between these routes of plant multiplication is important because
the genetic uniformity of the plants produced is generally affected. The success of
micropropagation technology largely depends upon the production of genetically uniform
plant offsprings. Sometimes plants propagated through tissue culture may not consistently
resemble the original resource plant due to epigenetic effects and genetic differences
arising due to genes/chromosomal changes. The shoot meristems are generally highly
stable and therefore multiplication by precocious axillary shoots in case of dicotyledonous
plants and shoot multiplication through shoot base meristems in case of most
monocotyledonous plants is generally preferred because mutation is least likely to occur.
Axillary bud proliferation method is currently the most popular approach to in vitro clonal
propagation of plants because the cells of the shoot apex are uniformly diploid and are
least susceptible to genotypic changes under culture conditions ensuring maximum genetic
uniformity of the resulting plants and avoiding breakdown of chimeras which is common
during adventitious bud culture (Cassells et al., 1980). However, the abnormalities
whatsoever observed in systems multiplied by axillary bud proliferation result due to the
development of some adventitious shoots, lasting effect of in vitro environment in some
cases (particularly growth regulators) and/or in perennial species by the appearance of
juvenile traits. On the contrary, the adventitious meristems are more susceptible to
mutations as they are usually derived from single cell or a small group of cells. Mutation is
a single celled event and a mutant cell (either spontaneous or induced) can divide to form
only a limited tissue within one layer resulting in chimera that generally prove unstable
and short lived. In some species, abnormal plants are readily formed adventitiously. In
Brassica oleracea, the plants regenerated directly from the leaf and stem explants include
over 70% tetraploids and 15% octaploids (Hoark et al., 1975; Hussey and Falavigna,
1980; Hussey, 1982). Despite this disadvantage, this phenomenon can enable the recovery
of new plant types. Multiplication of adventitious shoots is appropriate only where axillary
shoots are infrequent or unresponsive. The plants developed from somatic embryos are

80
normally true to type or less variable as compared to the plants developed via shoot and
root morphogenesis (Lorz et al., 1988). The distinction between these organogenic and
embryogenic routes of differentiation are based on morphological, anatomical,
biochemical and more recently the molecular evidence (Dudits et al., 1995; Merkle et al.,
1995; Yeung, 1995; Mayer and Jurgens, 1998; Arnold et al., 2002). Micropropagation
generally involves four steps: (1) Initiation of aseptic cultures: the selection, sterilization
and establishment of suitable aseptic cultures on defined nutrient culture medium and
under in vitro culture conditions; (2) Multiplication: growth and proliferation of multiple
shoots on an optimized shoot multiplication medium; (3) Rooting: induction and growth of
roots in in vitro regenerated shoots and (4) Hardening: acclimatization of the in vitro
raised plantlets under glasshouse conditions and subsequently transplantation of hardened
plants in fields. These steps have been formally designated as stage I, stage II, stage III
and stage IV (Murashige, 1974) and this terminology has been widely adopted (Street,
1977; Hussey, 1980; Conger, 1981 Debergh and Maene, 1981). Debergh and Maene
(1981) introduced the stage 0, making micropropagation a five step process. This stage
involves the preparation of mother plants to provide quality explants for better
establishment of cultures in stage 1. It includes exposing the stock plants to suitable light,
temperature and growth regulator treatments to improve the quality of explants. To reduce
the contamination problem in stage 1, the mother plants should be grown under controlled
conditions in a glasshouse and regularly watered. It would not only help minimize the
incident of infection in the stage 1 but may also reduce the neeed for a harsh sterilization
treatment. Each stage has its special requirements.
The success of a micropropagation protocol depends on a number of factors. The
age, nature, origin and physiological state of the inoculum/explant and seasonal variation
play a crucial role in the establishment of cultures and subsequent plant regeneration.
Sterilization method, media and culture conditions also affect the micropropagation. Pierik
(1987) has discussed the problem related to the rejuvenation and maturation in
micropropagation. On the other hand, a few problems often encountered during the
process are: (a) the browning/blackening of the culture medium and/or the explant due to
the injury caused during the excision of the explant inducing the cells to leach out phenolic
compounds which are readily oxidized. The oxidation products of phenols can be
phytotoxic and cause necrosis and eventually death of the explant. This problem is
especially common in woody species; (b) hyperhydricity or vitrification (morphological,
physiological and metabolic derangements frequently affecting herbaceous and woody

81
plants during their in vitro culture). Hyperhydrated shoots generally show poor growth,
become necrotic and finally die (Gaspar, 1991). The plants showing hyperhydration have
low survival rate during acclimatization (Ziv, 1991a,b). The problem of hyperhydricity
can be avoided by increasing the concentration of agar, using an agar with higher gelling
strength, lowering cytokinin levels, replacing one type of cytokinin by another or by
lowering NH4+ concentration.; (c) contamination: it is one of the most serious problems in
commercial micropropagation as it can cause a disastrous situation if it occurs at an
advanced stage of the production schedule. It is therefore extremely important to maintain
high standards of hygiene in and around the sterile area.
Various factors affecting adventitious rhizogenesis have also been discussed
(Moncousin, 1988). To increase the survival rate of plants humidity tents, antitranspirants,
supplementary light and CO2 enrichment have been employed (Kozai et al., 1988). A
computerized acclimatization unit has been developed to provide better environmental
control, high survival and rapid growth of the plants.
While there are numerous advantages associated with micropropagation, two major
limitations (a) customer acceptance; and (b) high production costs to its extended
application exist since the commercialization of this technology. One of the most
important aspect of commercial micropropagation is the economics involved and the cost
of plantlet production. According to Kozai et al. (1992a, b), this high production cost can
be traced to several technical reasons described as follows:
1. Micropropagation is labour intensive involving manual handling at 3 or 4 stages.
High labour costs alone accounts for nearly 60-70% of the total production cost
especially in developed countries. Therefore there is considerable interest in partial
or full automation of the process (Vasil, 1991a,b; Aitken-Christie et al., 1995).
2. Losses due to contamination during in vitro culture and hardening phases.
3. Poor throughput due to high mortality during acclimatization phase on account of
physiological/ developmental abnormalities in regenerated progeny.
4. High energy cost related to power consumption
5. Heterotrophic mode of in vitro growth that requires costly medium compounds like
sugar, agar, hormones etc.
6. Insufficient production against the market demand and lack of buy-back
arrangements.

82
As a result, the micropropagated plants are generally 10-50 times more expensive
than the conventional propagules such as cuttings or seeds (Pierik, 1991). Such an
escalation is bound to limit the scope of the technology unless an element of exclusivity is
attached to its demand. Therefore, micropropagation can be justified only when the
conventional methods of clonal propagation do not work or the product is of very high
commercial value. For some crops micropropagation can be economical only to produce
pathogen free foundation stock which can be further used to produce pathogen free elite
germplasm stock. Nevertheless, the micropropagation industries are adopting different
strategies for cost reduction (Debergh and Zimmerman, 1991; Takayama and Akita, 1994;
Kozai et al., 1997; Etienne and Berthouly, 2002). Various strategies have been attempted
and adopted to reduce the cost of plant production by manipulating methods, culture
conditions, nutrient media, adopting mechanical gadgets and resorting to automation.
Some of the promising strategies are:
1. Automation of various steps to reduce labour in-puts. To some extent automation
has been introduced at the media preparation stage by using dispensing machines.
Bioreactors are being used for the propagule multiplication stage in some cases.
2. Cost reduction through energy saving, resource recycling and resource
optimization.
3. Modulation of in vitro growth environment to promote autotrophic growth.
4. Use of cheaper substratum, culture vessels and energy sources.
5. Improvement in through-put index by refinement of acclimatization protocols.
Alternatively, the approach should be to use the the technique as an interphase
wherein only the base populations are generated in vitro and their subsequent enlargement
is achieved by conventional means. Another approach leading to reduction in labour cost
is to sub-contract the task of in vitro multiplication of their elite material in developing
countries where labour is comaparatively cheap. This strategy is being currently followed
by some of the developed countries. This has resulted in establishment of several
commercial tissue culture laboratories in the developing countries. Initially about 125
tissue culture units with a total production capacity of 240 million plants was set up in
India but at present there are only about 46 established commercial tissue culture units.
These establishments have so far been largely concentrating on exploiting the international
markets. Their production capacity ranges between 1 million to 5 million plants per annum

83
with an aggregate production capacity of around 180 million plants per year. Other small
units may account for an additional 20 million plants. Two tissue culture pilot plant
facilities were also set up for large scale production of elite planting material with the
main objective of demonstrating the field performance of tissue cultured plants. Each pilot
plant has production capacity of 1 million plantlet per annum. These pilot plants have been
converted to Microrpogation Technology Parks (MTPs) to provide an effective platform
for transfer of proven technologies to entrepreneurs. The plants prioritized by different
consumer segments include ornamentals, spices, forest trees, medicinal and aromatic
plants. The overall market for tissue cultured plants is expected to grow by 20-25%. The
micropropagation industry is capital and labour intensive. Rapid multiplication of plants
under disease free in vitro condition requires the production of quality plants at
competitive price and according to the demand of the market which will increase customer
acceptance. Any attempt to reduce the cost of production by accelerating the rate of
multiplication or automation should not be at the expense of the quality of the product.
Pushing a system beyond a limit with regards to the rate of multiplication or the life span
of a culture may introduce genetic aberrations which if not identified and discarded at the
right time, would get rapidly multiplied with normal type.
Bioreactors provide a rapid and efficient plant propagation system for many
horticultural, ornamental and medicinal plant species utilizing liquid media to avoid
intensive manual handling. The use of bioreactors for micropropagation was first reported
in 1981 for Begonia propagation (Takayama and Misawa, 1981). Since then it has proved
applicable to many species and plant organs including shoots, bulbs, microtubers, corms
and somatic embryos (Paek et al., 2001). Liquid media have been used for plant culture in
both agitated vessels and in bioreactors for somatic embryogenesis (Styer, 1985; Preil,
1991; Scarg, 1992) and as a system for plant propagation through the organogenic
pathway (Takayama, 1991; Ziv, 1992a). Automation of micropropagation in bioreactors
has been advanced by several authors as a possible way of reducing cost of
micropropagated plants (Preil, 1991; Sharma, 1992; Aitken-Christie et al., 1995; Leathers
et al., 1995; Son et al., 1999; Ibaraki and Kurata, 2001; Paek et al., 2001). Most reviews
and research reports on bioreactors for plant cells are concerned with the production of
plant secondary metabolites as the key product. There are only few reports on the use of
bioreactors for large-scale propagation of plants (Kessel and Carr, 1972; Ammirato and
Styer, 1985; Takayma, 1986; Wheat et al., 1986; Chen et al., 1987; Asenjo and O’ Carroll,
1988; Ziv, 1995, 2000). Different reactor configurations for plant cell suspensions, plant

84
tissue and organ cultures, relative advantages and selection criteria for various reactor
configurations for specific process applications have been studied in detail (Takayama and
Akita, 1994; Heyerdahl et al., 1995; Walker, 1995; Lee, 1997; Sajc et al., 2000; Paek et al.
2001; Paek and Chakrabarty, 2003). Some of the major differences between the different
reactors designed for the production of plant tissues have been reviewed by Cazzulino et
al., 1991. The mixing requirements of the reactor can be expected to be quite different in
these type of conditions. Various types of bioreactors have been used in plant tissue
cultures, though the hardware configuration of the bioreactors for plant tissue cultures are
basically same as for microbial and animal cell cultures (Cazzulino et al., 1991;
Takayama, 1991). Some of the problems associated with the operation of bioreactors were
reviewed by Ziv (1995, 2000), Takayama and Akita (1998). Bioreactor configuration must
be determined according to the mixing and aeration requirements of specific plants or
tissues, as well as for minimization of shear stress. Micropropagation in bioreactors
depends on a better understanding of plant responses to signals from the
microenvironment and on specific culture manipulations to control the morphogenesis of
the plants in liquid cultures (Ziv, 2005).
To further overcome higher production cost automation/mechanization is being
introduced in commercial micropropagation laboratories. Various types of robots and
microcomputers have been developed for the surface sterilization of tissue, media
distribution, inoculation and transfer of plants to soil. Computerized green houses to
control the growth environment are likely to further help in achieving the high survival
rates. The use of artificial intelligence, image analysis, video cameras, mistifiers,
automatic production lines and computerization of information management willl further
facilitate micropropagation.
The roots of safed musli (Chlorophytum borivilianum) are of immense commercial
importance because they are source of steroidal saponins and glycosides which are of great
pharmaceutical value. Tuberous roots of plants are widely used as general health purpose
tonic and aphrodisiac due to the prescence of steroidal saponins, viz. neotigogenin,
neohecogenin, stigmasterol and tokorogenin (Tandon and Shukla, 1995b). Safed musli
roots are used as a major ingredient of many formulations used for the treatment of
diseases like piles, diabetes, rheumatoid arthritis, various gynaecological problems and as
anti-pyretic, sialogogue, galactogue. It also possess haemostatic properties. It is a major
ingredient in ‘Chyavanaprasha’ which is used in the treatment of general debility and as a
vitalizer. Overall tuberous roots are used as an important ingredient in more than hundred

85
Herbal, Unani, Homeopathic and Allopathic formulations. The steroidal saponins are also
used in the industrial production of sex-hormones, corticosteroids and steroid derivatives
in general. Therefore, there is increased demand for dried roots of the safed musl
especially C. borivilianum in Indian and International drug market and conservation and
mass-multiplication of the plant for its domestication to meet the present demand is
urgently needed. C. borivilianum is having restricted distribution in India and
overharvesting without replenishment has further reduced its distribution. The cultivation
of safed musli is also limited in the country. Though plant propagates through seeds but
seed-setting and germination are low. Safed musli being mainly a vegetative crop,
vegetative propagation through conventional methods is difficult and has slow propagation
rate and time consuming. Therefore, poor seed seeting and germination and slow growth
rate are major constraints in the large-scale cultivation of safed musli.

86
2.2 REVIEW OF LITERATURE
2.2.1 Micropropagation: The most commercially exploited tool of plant
biotechnology
Plant tissue culture defined as the culture of plant cells, tissues and organs under
controlled environment on artificial medium, has an important role to play in the
production of agricultural, horticultural and medicinal plants. Among the many faceted
applications of plant tissue culture, micropropagation is one of the most commercially
exploited area of plant tissue culture. Conceptually, the primary goal of micropropagation
effort is to exploit the micropropagation competence of a living plant cell or tissue to
provide a large number of genetically identical, physiologically uniform and
developmentally normal population of an elite parent selection that must possess high
photoautotrophic capability to survive in field. Currently the most popular application of
micropropagation is the mass clonal multiplication of desired genotypes of plants.
Through micropropagation methods phenomenal rates of clonal multiplication running
into millions have been achieved. Industrial application of this technique started between
late 1960s and early 1970s. The development of micropropagation technique is based on
certain fundamental discoveries. The initial milestone in large scale production of
important plant species through this technique of micropropagation was laid down in the
mid of 20th centuary, when for the first time, cultured shoot meristems were used to get
virus-free plants of Dahlia and potato (Morel and Martin, 1952, 1955). The observation
that apical meristem culture in orchid could be a better alternative to vegetative
multiplication by Morel (1960) initiated the practical application of micropropagation in
horticulture. The other milestones included elucidation of the role of cytokinins in shoot
morphogenesis (Skoog and Miller, 1957) and in the inhibition of apical dominance, which
in turn releases the axillary meristems from dormancy (Sachs and Thimann, 1964).
The successful application of such fundamental discoveries to the multiplication of
plants by micropropagation has been a key factor in the development of this technology.
Presently the micropropagation technique is especially being used not only for those plants
which are difficult to propagate through conventional practices, but also for mass
multiplication of existing stocks of germplasm for biomass energy production and
conservation of economically important, elite and rare plant species that are threatened or
on the verge of extinction (Normah et al., 1997; Pania et al., 2000; Tewari et al., 2000;
Wawrosch et al., 2001). At present, being the most commercially exploited tool of plant

87
biotechnology, the micropropagation technique has been applied to many plant species
including crop plants, ornamental plants and trees (Murashige, 1989) and some important
medicinal and aromatic plants which are enumerated in Table-3.
The conventional propagation practices for clonal propagation of such plants are
time consuming and labour intensive. Ideally the micropropagation cycle must operate
over a shorter time frame and at a lower cost. Micropropagation is especially relevant
where starting plant material is limited, where clonal progenies are required, where quality
propagule are needed in bulk, or where speciallity plants (sterile somatic hybrids,
transgenics) are required for breeding and other research and development activities
(Bhojwani, 1990; George, 1993; Vasil and Thorpe, 1994; Thorpe, 1995).
Table-3: Some of the important medicinal and aromatic plants micropropagated in
vitro
Family Plant species Active constituent(s) Explant(s) used for micropropagation
Nutrient medium used
Reference(s)
Apocynaceae Catharanthus roseus
Alkaloids (Vinblastine and vincristine)
Seeds, meristem tip MS + 1.0 mgl-1
BAP Hirata et al., 1987
Rauvolfia serpentina
Alkaloids (reserpine, ajmalicine)
Nodal explants MS + 2.65 µM NAA + 0.33 µM Kn
Mathur et al., 1987
Apiaceae Centella asiatica Glycosides (asiaticoside, indocentelloside, brahmoside theankuniside)
Nodal segments MS + 22.2 µM BAP + 2.68 µM NAA
Tewari et al., 2000
Quercus spp. Tannins (Tannic acid) Axillary buds, nodal explants
MS + 0.1-1.0 mgl-1 BAP
Favre et al., 1993
Araliaceae Panax spp. Saponins (Ginsenoside)
Cotyledons MS + 30.0 µM GA3 Choi et al., 1998
Asclepiadaceae Gymnema sylvestre
Gymnemic acid Nodal explants from seedlings
MS + 1.0 mgl-1 BAP + 0.5 mgl-1 Kn + 0.1 mgl-1 NAA + 100.0 mgl-1 malt extract +100.0 mgl-1 citric acid
Komalavalli and Rao, 2000
Tylophora indica Alkaloids (Tylophorine, Tylophorenine and Tylophorinidine)
Nodal explants MS + 5.0 mgl-1 BAP + 0.5 mgl-1
NAA + 100.0 mgl-1
ascorbic acid
Sharma and Chandel, 1992
Asteraceae
Artemisia annua Sesquiterpene lactone (artimisinin, artimisinic acid)
Nodal explants, shoot tips Green parts from seedlings
MS + 0.2 mgl-1 BAP + 0.2 mgl-1
NAA Half strength MS + 0.2 mgl-1 BAP + 0.05 mg l-1 NAA
Jha et al., 1988 Woerdenbag et al., 1993
Chrysanthemum cinerariaefolium
Ester (Pyrethrins) Shoot tips MS + 3.0 mgl-1 BAP MS + 0.2 mgl-1 NAA
Zieg et al., 1983 Wambugu and Rangan, 1981

88
Saussurea lappa Alkaloid (Saussurine) Leaf segments from seedlings
MS + 5.0 µM BAP + 0.1 µM IAA
Arora and Bhojwani, 1989
Stevia rebaudiana
Diterpenoid glycoside (Stevioside, rebsudiosides)
Nodal segments MS + 10.0 mgl-1 Kn/2iP
Yang et al., 1981
Vanilla planifolia
Vanillin Shoot tips MS + 0.5 mgl-1 BAP
Philip and Nainar, 1986
Berberidaceae Podophyllum peltatum
Lignan (Podophyllotoxin)
Rhizome tips MS + 4.4 µM BAP + 0.025% (w/v) activated charcoal
Cerdeira et al., 1998
Burseraceae Commiphora wightii
Oleoresins (guggulsterols and guggulsterones)
Axillary buds MS + 17.8 µM BAP + 18.6 µM Kn + 100.0 mgl-1 glutamine + 100.0 mgl-1 thiamine HCL + 0.3% charcoal
Barve and Mehta, 1993
Colchiceaceae Gloriosa superba
Akaloids (Colchicine, glorisine)
Corms MS + 2.5 mgl-1 BAP or Kn + 0.25 mgl-1 NAA
Finnie and Van Staden, 1989
Dioscoreaceae Dioscorea spp. Steroids (Diosgenin) Shoot meristem MS + 15.0 mgl-1 AS + 0.1 mgl-1 NAA
Chaturvedi, 1975
Drosaraceae Drosera spp. Plumbagin (Naphthoquinone)
Shoot tips MS + 0.5 mgl-1 zeatin + 0.5 mgl-1 Kn
Jayaram and Prasad, 2007
Euphorbiaceae Phyllanthus amarus
Phyllanthin and Hypophyllanthin
Shoot tips MS + 0.1 mgl-1 Kn + 0.1 mgl-1 IAA
Bhattacharyya and Bhattacharya, 2001a
Fabaceae Glycyrrhiza glabra
Glycrrhizin Nodal explants NN + 0.05 mgl-1 Kn + 1.0 mgl-1 IAA
Kukreja, 1998
Gentianaceae Eustoma grandiflorum
Xanthones, flavonoides
Shoot tips, nodal segments
MS + 2.0 mgl-1 IAA + 2.0 mgl-1 Kn
Zenkteler and Zenkteler, 1987
Hypericaceae Hypericum perforatum
Bisanthron derivative (Hypericin), Phenolics (Hyperin, uercitrin)
Hypocotyl segments from seedlings
MS salts + B5 vitamins + 5.0 µM TDZ
Murch et al., 2000
Lamiaceae Coleus forskohlii Labdane diterpenoid (Forskolin)
Nodal segments Stem tip
MS +2.0 mg l-1 Kn + 1.0 mgl-1 IAA MS + 0.57 µM IAA + 0.46 µM Kn
Sharma et al., 1991 Bhattacharyya and Bhattacharya, 2001b
Mentha spp. Monoterpenes menthol, menthone, methyl esters, monoterpene derivatives (pulegone, piperitone, menthofurone)
Leaf disks Leaf and node explants
MS + 44.4 µM BAP + 250 mll-1
coconut water MS + 3.0 mgl-1 Kn + 1.0 mgl-1 IAA
Eck and Kitto, 1992 Kukreja, 1996
Lathyraceae Heimia salicifolia
Alkaloids (Cryogenin and nesodine)
Shoot tips, nodal explants
MS + 1.0 µM BAP + 2.3 µM Kn + 21.0 nM NAA
Rother, 1989
Lauraceae Cinnamomum camphora
Polyphenolics (Tannins) phenyl propanoid (Cinnamaldehyde)
Shoot tips and nodal explants
WPM + 13.32 µM BAP + 4.65 µM Kn
Babu et al., 2003

89
Liliaceae
Allium sativum Aloe barbadensis Lilium spp.
Allicin Anthraquinone glycoside (barbaloin) aloesin, aloesone glucomannans
Stem disc Meristem tips (Apical meristem) Shoot meristem bulbscales
Phytohormone free LS Medium MS + 80.0 mgl1AS + 0.1 mgl-1 IAA + 0.1 mgl-1 Kn MS + 0.25 mgl-1
2,4-D + 1.0 mgl-1 Kn MS + 0.1 mgl-1 NAA
Ayabe and Sumi, 1998. Conci and Nomes, 1991 Sanchez et al., 1988 Takayama and Misawa, 1979
Nyssaceae Camptotheca accuminata
Camptothecin Axillary buds B5 + 4.4 µM BAP + 0.54 µM NAA
Liu and Li, 2001
Orchidaceae Dendrobium chrysotoxum
Bisbenzyl (erianin) Shoot tips Modified Knudson’s C medium + 8.0 µM BAP or 1.0 µM TDZ
Roy et al., 2007
Poaceae Vetiveria zizanioides
Alcohol (vetiverol) Rhizomes, floral primordia, nodal segments
MS + 1.0 mgl-1 BAP + 1.0 mgl-1 IAA MS + 1.0 mgl-1 BAP + 1.0 mgl-1 NAA
Jagadishchandra and Sreenath, 1987 Sreenath and Jagadish chandra, 1990
Polygonaceae Rheum emodi Anthraquinone Shoot tip Leaf explant
MS + 2.0 mgl-1 BAP + 1.0 mgl-1
IBA MS + 2.0 mgl-1 BAP + 1.0 mgl-1 IAA or 0.25 mgl-1 IBA
Lal and Ahuja, 1989 -do-
Plumbaginaceae Plantago zeylanica
Alkaloid (Plumbagin) Nodal explant MS + 27.2 µM AS + 2.46 µM IBA
Selvakumar et al., 2001
Rosaceae Coluria geoides Essential oil (Eugenol) Hypocotyl segments with cotyledons and shoot tips
NN+ 0.3 mgl-1 BAP + 0.05 mgl-1 IBA + 10.0 mgl-1 AS
Olszowska and Furmanowa, 1993
Rubiaceae Gardenia grandiflora
Irridoid glycosides Shoot tips MS + 5.0 mgl-1 IAA + 10.0 mgl-1 BAP
Scaramizzi and D’Elia, 1984
Cephalis ipecacuanha
Isoquinolene alkaloid (emetine, emetamine etc.)
Nodal segments B5 + 0.01 mgl-1 NAA +1.0-3.0mgl-1 BAP
Ikeda et al., 1988
Saxifragaceae Bergenia crassifolia
Hydroquinone, bergenin, arbutin
Rhizome cuttings ML + 3.0 mgl-1 BAP + 0.1 mgl-1 NAA + 80.0 mgl-1 AS
Furmanowa and Rapczewska, 1993
Scrophulariaceae Picrorhiza kurroa
Glycosides (Picroside and kutkoside)
Shoot tips MS + 5.0 mgl-1 Kn + 1.0 mgl-1 IAA
Lal et al., 1988
Verbascum thapsus
Iridoid glycosides (harpagoside, aucubin), Flavonoids (3'- methyl guercitin, hesperedin, verbascoside) and saponins
Leaf discs MS + 13.32 µM BAP + 5.37 µM NAA
Turker et al., 2001
Solanaceae Datura spp. Tropane alkaloids (scopolamine etc.)
Nodal explants MS + 1.0 mgl-1 BAP
Santos et al., 1990

90
Duboisia myoporoides
Tropane alkaloids (hyoscine and hyoscyamine)
Nodal explants MS + 3.0 mgl-1 Kn + .01 mgl-1 IAA
Kukreja and Mathur, 1985
Withania somnifera
Alkaloids, steroidal lactones
Leaf explants, seeds, shoot tips from seedlings
MS + 4.4 µM BAP + 7.99 µM IAA
Sen and Sharma, 1991; Kulkarni et al., 1996
Taxaceae Taxus spp. Taxol Bud explants Half strength MS + 2.5 mgl-1 BAP + 1.0 gl-1 activated charcoal + 100.0 mgl-1 silver nitrate
Chang et al., 2001
Valerianaceae Valeriana wallichii
Valepotriates Shoot tip and axillary bud
MS + 5.0 mgl-1 Kn + 1.0 mgl-1 IAA
Mathur et al., 1988
Zingiberaceae Zingiber officinale
Sesquiterpenes Shoot bud MS + 1.0 mgl-1 BAP
Hosoki and Sagawa, 1977
Nothapodytes foetida
Quinoline alkaloids (camptothecin and 9-methoxycamptothecin) and Nothapodytines
Hypocotyl segments from seedlings
MS + 2.2 µM TDZ Rai, 2002
2.2.1.1 Priority medicinal plants for micropropagation
Medicinal plants with established market demand coupled with some inherent problems
associated with their field cultivation such as prolonged juvenile phase, poor flowering,
low seed set or viability and strong recalcitrant nature, are therefore fast attracting the
attention of micropropagation industry to meet their demand. Medicinal herbs in general
are characterized by slow growth rate, long gestation period between planting to economic
harvesting, predominance of vegetative mode of propagation or cross pollination
behaviour and tedious extraction and purification methods for the isolation of their active
principles. Lack of quality propagules of superior genotypes is perhaps the most deterring
factor in attracting farmers to take up the commercial cultivation of these herbs at a scale
that can commensurate with their supply to drug industries. Based on several successfull
stories and established industrial scenario in ornamental, forestry and vegetable crop
business, plant tissue culture based micropropagation technologies are being viewed now
to provide solutions to these problems of medicinal plant sector. Medicinal herbs that are
occupying the core attention of the industry (Mathur and Mathur, 2003) include the one
that: (i) are endangered, facing high level of threat of extinction or becoming rare (Ginkgo,
Picrorrhiza, Podophyllum, Rheum etc.); (ii) have established commercial demand under
low-volume-high value category of products (Echinacea, Catharanthus, Hypericum,
Ocimum, Papaver, Plantago, Taxus etc.); (iii) find excessive use in traditional systems of
medicine (Acorus, Adathoda, Asparagus, Centella, Chlorophytum, Emblica, Ephedra,
Holarrhena, Phyllanthus, Swertia, Terminalia, Tinospora etc.); (iv) are identified as a
potential source of lead molecules for modern drugs (Coleus, Coptis, Erythroxylum,

91
Gymnema, Hypericum, Morinda, Taxus, Vinca etc.); (v) have prominent vegetative mode
of propagation with narrow genetic base (Allium, Curcuma, Dioscorea, Gloriosa, Zingiber
etc.); (vi) Crops that are in prime focus of genetic manipulation efforts (Artemisia,
Catharanthus, Hyoscyamus, Papver, Taxus etc.); (vii) have been consciously protected by
tribals as courtyard cultigens in sacred mangroves (Argyreia, Clerodendron, Crataeva,
Elaeocarpus, Ficus, Gymnema, Morinda, Zanthoxylum etc.); (viii) yield costly
pharmaceutical, nutraceutical or cosmetic compounds for which no synthetic substitute is
available (Artemisia, Catharanthus, Chlorophytum, Silybum, Taxus etc.).
2.2.1.2 Shoot meristem culture: A means of large-scale clonal propagation
The concept of using shoot meristem culture for in vitro propagation originated with the
application of this technique to produce virus free Cymbidiums from infected donor plants
through meristem culture (Morel, 1960) as it was reported that root and shoot apices of
virus infected plants are frequently devoid of pathogens (White, 1934; Kassanis, 1957).
Later on, the technique was established as a process of large-scale clonal propagation for
virus-free orchids (Morel, 1963).
The technique of shoot meristem culture allows the stem tip to grow on nutrient
media which supports the plant growth. This stem tip is characterized by the prescence of
apical meristem comprising of meristemetic cells capable of division (Esau, 1960) and few
leaf primordia present at sub-apical region. If a portion beyond the meristematic sub-apical
region is included, the procedure is referred to as shoot tip culture rather than meristem
culture (Cutter, 1971; Wang and Hu, 1980, 1985). The cells of apical meristem possess a
high degree of morphogenetic potential for generating plants having the similar genotypic
and phenotypic composition as that of mother source, as these cells tend to be genetically
stable. Endogenous contaminants do not easily invade the meristem, often resulting in the
formation of disease free plants. The nutrient medium and phytohormones stimulate and
support the growth and development of multiple axillary buds. Three to six-fold increase
in shoot number can be generally achieved at every 4-6 weeks, which results in the
production of millions of plants from each cultured shoot meristem in one year. Besides
this, there are two other methods through which the plants can be propagated in vitro.
These include: (a) de novo formation of shoot meristems in callus tissues and (b) somatic
embryogenesis.

92
2.2.2 Routes of propagation: Distiction between various routes of regenaration in
micropropagation is important regarding uniformity of clonal progenies
2.2.2.1 Regeneration through callus cultures
The production of plants through callus tissue is generally avoided because of frequent
occurrence of genetically variant plants and even chimeras. These variations are called
‘somaclonal variations’ and can be of two types, either epigenetic variations or heritable
variations (Lorz et al., 1988; Skirvin et al., 1994). The epigenetic or developmental
variations are the temporary variations which include phenotypic changes due to the
expression of specific genes under a particular set of environmental conditions (Hartman
and Kester, 1983). Stable or heritable variations involve phenotypic and biochemical traits
due to karyological variations (Lorz et al., 1988) involving either single or multiple gene
changes.
2.2.2.2 Somatic embryogenesis
On the other hand somatic embryogenesis i.e. the formation of embryo like structures from
somatic cells and their germination into complete plants (Bhaskaran and Smith, 1990) has
now been proved as potentially most efficient and economic method for the large scale
propagation of plants. The plants developed from somatic embryos are normally true to
type or less variable as compared to the plants developed via shoot and root
morphogenesis (Lorz et al., 1988).
Therefore, the differentiation of whole plants in tissue cultures with minimum
variation may occur via shoot and root differentiation through axillary branching or
alternatively, the cells may undergo embryogenic development to give rise to bipolar
embryos, reffered to as ‘somatic embryos’ to distinguish them from zygotic embryos. A
shoot bud and an embryo are distinguishable on the basis of recognizable morphological
differences between the two. The former is a monopolar structure. It develops procambial
strands which establish a connection with the pre-existing vascular tissue dispersed within
the cultured explant. On the other hand, an embryo is a bipolar structure with a closed
reticular end. It has no vascular connection with the maternal callus tissue or the cultured
explant.
The adventitious shoot buds on the other hand do not form cotyledons and resultant
leaves are borne in phyllotaxy similar to the one present in parent plant. These shoots are
open ended with vasculature typically joined with mother tissue. Shoots are formed

93
independent of roots and vasculature of roots (if present) may not be in continuity with
that of shoots.
2.2.3 Factors affecting micropropagation
There are several other factors, which directly or indirectly play a major role in clonal
propagation through meristem culture or somatic embryogenesis in any plant system
(Cassells and Minas, 1983).
2.2.3.1 Explant: Type and nature of the explant
Since the objective of micropropagation is large-scale production of genetically uniform
planting material, the role of explant becomes very important in determining the success
of any micropropagation system. An explant is an important independent living unit
capable of presenting all genetic information that the excised cell(s) or tissue(s) perform at
their original location. Murashige (1974) recognized several factors that should be
considered in explant selection viz., the organ that is to serve as tissue source (nature and
origin of the explant), the physiological and ontogenic age of organ, the season in which
the explant is obtained, the size of explant, the overall quality of the mother plant, the
position of the explant on the mother plant or stock plant, orientation of the explant on the
medium and the inoculation density. Therefore, it is imperative to consider these points
before selecting any explant for micropropagation system because the ability of
regenereation of any explant is influenced by above mentioned factors. The influence of
the type and nature of the explant on establishment and subsequent plant regeneration
through micropropagation has been very well studied in a number of plant species
including Chrysanthemum (Lazar and Cachita, 1985), Drosera species, Gerbera and
Saintpaulia (Cachita, 1984, 1987). In recent years also, work has been done in this
direction in many plant species including Morus alba (Bhau and Wakhlu, 2001), neem
(Salvi et al., 2001), grapes (Thomas, 2001) etc. The physiological status of an explant is
affected by the age of the donor plant which has a direct bearing on the regenereability of
the explant. The use of young and meristematic tissues has, in many cases enabled raising
of regenerative cultures when mature and differentiated explants failed to show such a
response. The source of explant has been considered as an important factor in the indirect
regeneration of plants in Chrysanthemum (Bush et al., 1976; De Jong and Custers, 1986).
Earlier Kassanis and Verma (1967) reported that the prescence of leaf primordia
determines the capability of a shoot meristem explant to develop either into callus or into a
plant. In some plants, such as tobacco, almost all parts are ameneable to in vitro plant

94
regeneration, in others this potential is restricted to only certain tissues. In plants where
different explants respond, some may be more regenerative than the others. In Crotalaria
juncea (Ramawat et al., 1977) and Glycine (Kameya and Widholm, 1981) the hypocotyls
exhibits higher potential for shoot formation than the root segments. In Lactuca sativa
(Doerschug and Miller, 1967) and B. juncea (Sharma, 1987) cotyledon was the best
explant for plant regeneration.
2.2.3.2 Preparation and orientation of explant
The preparation of explants is also important. In cotyledon cultures of B. juncea, shoot
buds or roots are formed only at the cut end of the petiole depending on the culture
medium. The lamina lacks this potential. However, the prescence of laminar tissue is
essential for the petiolar tissue to exhibit totipotency. Therfore, the ideal explant to achieve
regeneration is the lamina together with petiole. This is also true for B. oleracea (Lazzeri
and Dunwell, 1986; Horeau et al., 1988). Orientation of the explant on the medium also
proved to be a critical factor for organogenic differentiation in cotyledon cultures of B.
juncea (Sharma et al., 1990) and guava (Chen and Chang, 2002).
2.2.3.3 Size of the explant
The size of the explant to be cultured is of great significance. Smaller explants are always
better for regenerative response with low chances of contamination. The influence of the
size of the explant on in vitro clonal propagation has also been studied in detial in various
plant species including Cassava (Kartha and Gamborg, 1975). The explant of varying
sizes have been tried for micropropagation. However, best results have been achieved with
explant size ranging between 1- 4 cms. The size of the explant plays a vital role in
meristem culture. The appropriate size of the meristem tip ranges between 0.2-0.5 mm.
Size smaller than the lower critical limit leads the explant to mortality. Silmilarly larger
explants (>0.5mm) lead to establishment of in vitro cultures which will not be free from
viruses.
2.2.3.4 Preconditioning of explant
The preconditioning of the stock plant itself is very important in determining the inductive
process that results in the morphogenetic response in vitro. The environmental factors such
as light, temperature and application of plant growth regulators can yield better, hygienic
and responsive explant for micropropagation.

95
2.2.3.5 Genotype
Genotype specificity to regeneration has been reported in a number of plants. Intraspecefic
variation for regeneration in tissue cultures of B. oleracea was observed (Lazzeri and
Dunwell, 1984; Murata and Orton, 1987; Horeau et al., 1988). Genetic variation for
regeneration occurs between varieties and in outbreeding species, even within varieties
such as in tomato (Padmanabhan et al., 1974; Kurtz and Lineberger, 1983). Different
cultivars of alfalfa exhibited variations in regeneration capacity when subjected to the
same culture regime (Saunders and Bingham, 1972).
2.2.3.6 Media composition and plant growth regulators
Basic nutritional compositions are used on the basis of physical state of the culture system
as well as plant system. Different plant systems have shown their varied in vitro nutritional
requirements (Gamborg and Wetter, 1975). Even tissues from different parts of a plant
may have different requirements for optimal growth (Murashige and Skoog, 1962).
Similarly, different culture systems, including callus cultures, plant regeneration through
micropropagation, meristem and embryo culture, etc. have different requirements in the
form of basal medium compositions. NN (Nitsch and Nitsch, 1969), B5 (Gamborg et al.,
1968), WPM (Lloyd and Mc Cown, 1981) media have been employed in various studies
and plant systems, but the most commonly used medium is Murashige and Skoog’s (MS;
Murashige and Skoog, 1962) basal medium. Some times, it is also used with some
modifications because of the growth additives and salt concentration requirements within
the media, which vary from plant species to species and even from one stage of culture
development to another (Hughes et al., 1978; Evans et al., 1981).
In addition to nutrients, it is generally necessary to add one or more growth
substances, the plant growth regulators (plant growth hormones) to support good growth
of plant tissues and organs. However, the requirement for these substances varies
considerably and it is believed that it depends on their endogenous levels in the plant
system which is variable with the type of plant tissue, plant type, and the phase of plant
growth and mode of differentiation i.e. organogenesis, embryogenesis or rhizogenesis
(Reinert et al., 1977). The growth regulators are required in very minute quantities (µ mol
l-1 values). The main plant growth regulators used in tissue culture studies are auxins and
cytokinins. In tissue culture auxins have been used for cell division and root
differentiation. The auxins commonly used in tissue culture are: indole-3-acetic acid
(IAA), indole-3-butyric acid (IBA), naphthalene acetic acid (NAA), naphthoxyacetic acid

96
(NOA), para-chlorophenoxyacetic acid (p-CPA), dichlorophenoxyacetic acid (2,4-D) and
trichlorophenoxyacetic acid (2,4,5-T). Of the various auxins, IAA is the least stable in the
medium. Therefore, synthetic auxins such as NAA and IBA have been preferred for use in
tissue culture media. They are widely used for rooting and in interaction with a cytokinin
for shoot proliferation. Because of its strong tendency to induce callusing, 2,4-D is
avoided when shoot multiplication is attempted through axillary branching or adventitious
shoot development. For somatic embryogenesis, however 2,4-D is the most effective
auxin.
In tissue culture media, cytokinins are incorporated mainly for shoot proliferation
by the release of axillary buds from apical dominance (Einset, 1985). These compounds
are also used for cell division and differentiation of adventitious shoots from callus and
organs. Most commonly used cytokinins are: 6-benzylaminopurine (BAP), isopentenyl-
adenine (2-iP), furfuryl amino purine (Kinetin), thidiazuron (TDZ) and zeatin. BAP, a
synthetic cytokinin, is most frequently used (Bhojwani, 1980a). 2-iP has been also
effective in the cases where other cytokinins have not given optimal response (Bhojwani,
1980b; Cohen, 1980). Zeatin is not preferred because of its high cost. However, in
Japanese Persimmon both BAP and 2-iP were ineffective and best shoot growth and
multiplication occured in half strength MS medium containing zeatin (Fukui et al., 1989).
Since 1982, thidiazuron, a urea derivative, has been used as cytokinin in several studies on
shoot multiplication (Lu, 1993). It is especially more effective than the other cytokinins
(with purine ring) particularly in recalcitrant woody species. Thidiazuron is effective at
much lower concentration than other cytokinins. For shoot multiplication, the prescence of
an auxin in the medium is not obligatory. In a number of cases, a cytokinin alone is
enough for optimal shoot multiplication (Lane, 1979; Stoltz, 1979; Bhojwani, 1980 b;
Garland and Stoltz, 1981).
Compared to auxins and cytokinins, gibberellins are used less frequently
(Schnabdrauch and Sink, 1979; Wochok and Sluis, 1980; Brand and Lineberger, 1992).
Abscissic acid is most often required for the normal growth and development of somatic
embryos and in many cases its prescence is essential (Ammirato, 1988). More recently,
there has been considerable interest in the application of growth retardants, such as
paclobutrazol to reduce hyperhydricity and regulate leaf growth and function in relation to
control of water stress during the acclimatization stage of micropropagation (Ziv, 1992b).
Ancymidol has been used to inhibit leaf formation and promote shoot formation in
gladiolus (Ziv, 1989; Ziv and Ariel, 1991). Growth retardants like ancymidol or

97
paclobutrazol (Ziv, 1992b, 2000) have been used in many cases where they reduced water
uptake during cell proliferation, decreased vacuolation and intercellular spaces, shortened
the stems, inhibited leaf expansion and minimized shoot malformation (Ziv, 2005). Use of
growth retardants controlled shoot growth and provided culture conditions that reduced
abnormal leaves and enhanced formation of bud clusters in potato, gladiolus and
Ornithogalum dubium resulting in a high proliferation rate and limited hyperhydricity
(Takayama, 1991; Ilan et al., 1995; Levin et al., 1997; Takayama and Akita, 1998)
2.2.3.7 Light, temperature and pH requirements
The other environmental conditions i.e., intensity and quality of light, photoperiod,
temperature, relative humidity, culture vessel type etc. play a key role in the in vitro plant
growth and its overall development. In general, shoot cultures require the prescence of
light (Wang and Hu, 1980). Several reports have been published revealing the optimal
need of light and darkness for different plant species (Pillai and Hildebrandt, 1968; Huth
and Bode, 1970; Hasegawa et al., 1973; Skoog, 1944). The quality of light also influences
differentiation (Weis and Jaffe, 1969; Bagga et al., 1985). Involvement of phytochrome in
shoot induction is also reported (Bagga et al., 1985). The most commonly used
photoperiod regime in the culture rooms is 16/8 hrs day/night period (Murashige, 1977). In
micropropagation studies, white light emitted from cool white tube lights is mostly used. It
is generally accepted that a light intensity of around 1000 to 5000 lux in culture rooms
may be required (Pierk, 1987). The optimum incubation period required for the normal
growth and development lies at 25±2°C (Hasegawa et al., 1973; Lane, 1979; Minocha,
1980; Pierk, 1987).
Generally the pH of the medium is 5.8±0.1 before sterilization. The pH of the
medium changes at various stages of preparation and culture. Such pH changes later on
influence the availability of various mineral ions in the medium and their uptake by the
plant tissue. Howard and Marks (1989) studied the effect of change in pH on shoot
production in Magnolia and Dianthus.
2.2.3.8 Oxidative browning
The phenolic products released from cultured explants readily oxidize to produce quinines
and cause browning/blackening of the culture medium and/or the explant and cause
necrosis and, eventually death of the explant. Some of the methods that have been used to
overcome this problem are: (1) Quick transfer of the explant within the same vessel or to a
fresh medium 2 or 3 times at short intervals (Morel, 1972; Preece and Compton, 1991); (2)

98
the initial culture in liquid medium before planting on semi-solid medium; (3) the addition
of antioxidants, such as cysteine-HCL, ascorbic acid or citric acid to the culure medium
(Sondahl and Sharp, 1977; Skirvin and Chu, 1979); (4) addition of adsorbants such as
activated charcoal (Ziv and Halevy, 1983) or polyvinypyrolidone (polycar AT or PVP);
(Walkey, 1972; Gupta et al., 1980).
2.2.4 Rhizogenesis: Root induction in in vitro regenerated shoots
To obtain complete plantlets, the in vitro regenerated shoots must be transferred to a
rooting medium which is normally different from the shoot multiplication medium,
particularly in its hormonal and salt composition. For in vitro rooting nutrient salts in the
medium have a dramatic effect both on rooting efficiency and number of roots induced per
microcutting. Often, where shoot multiplication was induced on full strength MS medium,
the salt concentration was reduced to half (Seabrook et al., 1976; Zimmerman and
Broome, 1981; Anderson, 1984) or a quarter (Skirvin and Chu, 1979) for rooting. The
effect of dark treatment and temperature variations (Zimmerman, 1984; Zimmerman and
Fordham, 1985) and number of shoot multiplication cycles on rooting (Gupta et al., 1981)
has been also studied. For rooting, individual shoots measuring generally about 2-3 cm in
length are excised and transferred to the rooting medium. However, it would not only be
convenient but also economical if shoot culture biomass formed after a shoot
multiplication cycle, can be handled as units during the rooting and transplantation stage
and individual parts separated at a later stage as in garlic (Bhojwani, 1980b; Bhojwani et
al., 1982). It has been experienced that handling of plants during transplantation is made
convenient if the roots are not of too much length. Longer roots may be damaged during
transplantation and thus reduce the chances of survival of plants.
It is a common practice to induce roots in in vitro regenerated shoots but many
commercial laboratories prefer to treat the in vitro multiplied shoots as microcuttings and
root them in vivo (ex vitro), if possible because of advantages associated with it (Debergh
and Read, 1991; Preece and Sutter, 1991).
2.2.5 Transplantation: The biological hardening
Poor adaptability of most of the in vitro raised plants to soil or acclimatization is the major
factor limiting the widespread use of micropropagation technique at the industrial level.
The acclimatization is the climatic adaption of the plant, that has been transferred to a new
environment (Conover and Poole, 1984). Low survival index of the micropropagated
progenies at the time of their transfer in natural conditions is generally due to the change

99
of the environment (sudden switch from heterotrophic mode of life in in vitro cultures to
autotrophic mode of life as they are transferred to soil under glass or greenhouse
conditions).
The normal plant development during acclimatization stage is mandatory to ensure
a high percentage of survival after transplanting ex vitro (Preece and Sutter, 1991; Ziv,
1991a). The low light, high humidity, lack of adequate CO2 and sufficient energy through
carbohydrate source i.e. sucrose in the medium create heterotrophic mode of nutrition for
in vitro raised plantlets. Consequently, these plants are poor in chlorophyll content and the
enzymes responsible for photosynthesis are inactive or absent altogether (Grout and
Aston, 1977; Wetzstein and Sommer, 1982; Donnelly and Vidaver, 1984). Therefore,
gradual acclimatization is necessary for the survival of these plants during transition phase
from culture room to the greenhouse/field. In the first phase of acclimatization the main
environmental stress to the plants is the change from a substrate rich in organic nutrients to
one providing only inorganic nutrients. This probably restores the photosynthetic activity
of the plants enabling them to withstand the subsequent reduction in the ambient relative
humidity and survival under field conditions. Acclimatization of micropropagated plants
on a large scale is generally carried out in a polyhouse or glasshouse where a gradation of
high-to-low humidity and low-to-high levels of irradiance are maintained. High humidity
is maintained by a misting or fogging system.
The problems related to the survival of plants during hardening or acclimatization
has been a matter of research since beginning. Brainerd and Fuchigami (1981) examined
the response of micropropagted apple plants to relative humidity at the acclimatization
stage (Sutter and Hutzell, 1984). Physiological state of micropropagated plants during
acclimatization also plays an important role in the adaption of plants to new environment
(Huylenbroeck and Debergh, 1996; Huylenbroeck et al., 1998). Many other aspects of the
acclimatization of plants, changed environmental conditions and consequent response of
the plant systems have been reported for many plant species (Diaz-Perez et al., 1995;
Pospisilova et al., 1999). It was observed that CO2 enrichment and providing
supplementary illumination i.e. exposure to high light intensity during acclimatization
resulted in autotrophy and better growth, and improved hardening in a number of plant
species (Lee et al., 1985; Lakso et al., 1986; Desjardins et al., 1987; Kozai et al., 1988;
Tal et al., 1992).

100
Some other methods recently recommended to improve the transplantation success,
presumably at lower costs are: (i) in vitro acclimatization, and (ii) in vitro formation of
storage organs. During in vitro hardening, the roots of micropropagated plants, particularly
of tree species are treated with Arbuscular Mycorrhizal Fungi (AMF), symbiotic bacteria,
growth promoting rhizobacteria such as Rhizobium species, pathogen antagonists or
hypervirulent strains of plant pathogens (Lovato et al., 1996; Gange and Ayres, 1999)
which make symbiotic association with plants to ensure better survival and growth of the
plants after transfer to field (Perinet and Lalonde, 1983; Bertrand and Lalonde, 1985;
Dhawan and Bhojwani, 1987). These microorganisms show functional host-specificity for
their efficacy (Nowak, 1998). They must be therefore assessed for their host range and
capacity to provide acclimatization advantage to tissue progeny.
The high relative humidity (RH) in the head space of the culture vessel and water
potential of the medium influence plant growth and development. The culture of the plants
at relatively low humidity reduces their wilting after transfer to soil by improving stomatal
movement (Wardle et al, 1983; Short et al., 1987) and increasing the deposition of
epicuticular wax on the leaves (Wardle et al, 1983; Maene and Debergh, 1986; Short et
al., 1987).
Certain growth retardants have been reported to promote in vitro root and shoot
growth (Chin, 1982; Khunachak et al., 1987), reduce shoot elongation and leaf area,
increase chlorophyll content and improve stress resistance of in vitro grown plants by
promoting epicuticular wax deposition and stomatal response and development of thick
roots (Ziv, 1991a, 1992a; Novello et al., 1992; Roberts et al., 1992). Addition of
paclobutrazol to the rooting medium enhanced dessication tolerance of micropropagated
chrysanthemum, rose and grapwine plants (Smith et al., 1990; Roberts et al., 1992). In
Philodendron, application of paclobutrazol or ancymidol during shoot proliferation
enhanced ex vitro survival of micropropagated plants (Ziv and Ariel, 1991).
In several plant species increase in plant growth and photoautotrophic behaviour
could be achieved by reducing or deleting sucrose from the medium, increasing CO2 level
around the plants (plants deriving their carbon requirement from CO2), and increasing
irradiance (Kozai, 1991a, c). Some of the advantages of photoautotrophic
micropropagation are: development of structurally and physiologically normal plants;
simplified rooting; reduced use of growth regulators; reduced problem of microbial
contamination due to lack of sucrose in the medium and facilitation of automation and

101
computerization of micropropagation (Kozai, 1991b, c). Hayashi et al. (1988) developed a
microcomputer controlled acclimatization chamber in which levels of CO2, relative
humidity, irradiance, temperature and air flow could be monitored. Acquired autotrophy
for growth in the abscence of sugars, vitamins and hormones would not only make shoot
and root growth more normal but will also contribute towards cost economization of the
protocol. However, photoautotrophic micropropagation is still very much at the laboratory
stage and many aspects, including its economic feasibility and practical utility have to be
worked out before it can be taken up at commercial level.
2.2.6 Advantages of micropropagation
There are many advantages of in vitro technique of plant propagation compared to the
conventional procedures (Murashige, 1974, 1978; George and Sherrington, 1984; Pennell,
1984) which made this technique to become a routine procedure for large-scale production
of many plant species including crop plants, vegetables, spices, medicinal and aromatic
plants etc. This technique has some significant advantages:
1. The advantage of this aseptic method of clonal propagation, popularly called
‘micropropagation’ over the conventional methods is that in a relatively short time
and space a large number of plants can be produced starting from a single
individual independent of seasonal constraints. Once established, actively dividing
culture is a continuous source of microplants for further multiplication and
production under field/glass house conditions.
2. An enhanced rate of plant multiplication in vitro would reduce considerably the
period between the selection and release of a new cultivar. The amount of plant
material released from quarantine being usually limited, micropropagation should
also hasten the introduction of new crops following their passage through
quarantine.
3. Since micropropagated plants are genetically homogenous, therefore plants which
are more or less true-to-type to the parent plant can be produced by this method.
4. Since shoot tip culture allows the plants to become virus free, therefore disease
free plants can be produced from diseased ones.
2.2.7 Cost-effectiveness in micropropagation
Inspite of these merits, the micropropagation technique has certain demerits also which
have limited the use and exploitation of this technique at industrial level. The major
limitation is the higher cost of plant production. Hence, the most challenging aspect at

102
present is to reduce the production cost, thereby improving the production efficiency so
that it can compete with seed production or traditional vegetative propagation methods e.g,
cuttings, tubers and bulbs, grafting (Anderson and Meagher, 1977; Sluis and Walker,
1985; Donnan, 1986; Levin and Vasil, 1989; Aitken-Christie, 1991; Vasil, 1991a). To
overcome this limitation, a number of cost reduction strategies have now been developed.
Employing liquid medium in shake flask cultures, reduction in plant production cost can
be achieved. (Takayama and Misawa, 1981; Weathers and Giles, 1988; Takayama, 1991;
Paque et al., 1992; Bhagyalakshmi and Singh, 1995).
2.2.8 Physical states of the culture medium
The rate at which cultures grow and produce shoots during micropropagation can largely
be influenced by physical states of the medium. In certain cases, even the morphogenetic
pattern of differentiation of tissues or organs may be completely modified accordingly on
their culture in different states i.e. semi-solid or in the liquid nutrient medium of the same
composition (Skoog, 1944; Dougall and Shimbayashi, 1960). Basically used for plant
cells, somatic embryos and organ cultures, both in agitated flasks and bioreactors (Smart
and Fowler, 1984; Attree et al., 1994; Tautorus and Dunstan, 1995), the liquid culture
medium has been relatively less used for the purpose of micropropagation. However,
certain merits of this technique are helpful in proving it now as one of the important
methods for cost reduction during micropropagation. Culture of somatic embryos and
organs grown in liquid medium often results in faster rate of growth than on semi-solid
medium because greater surface area of the explants is in contact with the medium,
diffusion gradient between the medium and explants is reduced which results in more
effective uptake of nutrients and phytohormones, leading to the better shoot and root
growth (Maene and Debergh, 1985; Ziv, 1989; Smith and Spoomer, 1994; Sandal et al.,
2001). Besides, the toxic metabolites accumulated in the vicinity of the tissue are
effectively dispersed in the liquid medium. The disappearance or the lesser expression of
activity of apical dominance due to continuous shaking of the tissues in the medium is
another important feature of liquid cultures, which generally leads to the induction and
proliferation of numerous axillary buds, thus favouring the production of large number of
plants which are more or less true-to-type (Takayama and Misawa, 1981; Harris and
Mason, 1983; Douglas, 1984; Pierik, 1987; Chu et al., 1993). Furthermore, within the
shake culture conditions, the growth and multiplication rate of shoots is enhanced by
forced aeration, since continuous shaking of the medium provides sufficient oxygen
supply to the tissue, which ultimately leads to their faster growth. Agitation speed of liquid

103
culture media is generally slow for organ culture i.e. around 60-70 rpm and higher for cell
culture i.e. around 100-150 rpm (Saxena and Sharma, 2003). The response of cultured
tissue to media manipulation and selection pressure is also more rapid. In addition to these
advantages, the preparation of liquid medium and handling of shake cultures is easier as
compared to the semi-solid one. The advantages of liquid culture medium for enhancing
shoot proliferation and growth have been reported in several plant species (Chu et al.,
1993; Sandal et al., 2001; Kim et al., 2003).
For several experimental plant systems liquid medium has proved either critical for
their survival in the culture or beneficial for multiplication (Simmonds and Cumming,
1976; Snir and Erez, 1980; Molnar, 1987; Saxena and Bhojwani, 1993). In Cattleya and
most of the bromeliads cultures could be initiated only in liquid medium (Murashige,
1974; Hosoki and Asahira, 1980). On the other hand in solid medium, the rate of growth
may be slow as the toxic exudates may not diffuse from the explants quickly. Some brands
of most commonly used gelling agent-agar contain inhibitory substances, which may
prevent morphogenesis in certain cases (Powell and Uhrig, 1987). The agar besides adding
(upto 65%) to the cost of the culture medium also results in local accumulation of heat and
hinders the access of dissolved oxygen to cultured cells (Kohlenbach et al., 1978).
2.2.9 Hyperhydricity or vitrification
Liquid medium generally increases nutrient uptake and faster growth of in vitro cultures
due to lack of polarity and orientation. However, this continuous contact of plant tissues
with the liquid medium results into the problem of hyperhydricity or vitrification in some
of the cultured plant species (Debergh and Zimmermann, 1991). The resultant propagules
are normally fragile, have a glossy appearance with succulent leaves and a poor root
system (Ziv, 1995). The leaves normally develop an unorganized mesophyll tissue with
large intercellular spaces, deformed vasculature and an abnormal epidermis that lacks
cuticle and possesses malfunctioning guard cells that do not respond to closure signals
(Ziv, 2000). When such malformed plants are transplanted into soil they exhibit high
mortality rate due to abnormal photosynthetic and transpiration mechanism. To overcome
this problem, tissue culturists have initiated experiments with a two laye culture system
(double-phase culture system) for culture of plants by adding a static liquid phase medium
over semi-solid medium (Maene and Debergh, 1985; Viseur, 1987). In this system,
nutrients can be replenished by adding a layer of liquid medium over a solid medium,
which has already been exhausted, thus reducing the labour cost and stress during
subculturing (Aitken-Christie and Jones, 1987).

104
Hyperhydration can be reduced and the quality of in vitro raised plants improved
by lowering the humidity in the headspace of the culture vessel by: (1) increasing the
concentration of agar (Debergh et al., 1981); (2) improving the aeration of the container
(Dillen and Buysens, 1989; Rossetto et al., 1992); and (3) bottom cooling of the culture
vial (Maene and Debergh, 1987). Other treatments which may help to reduce
hyperhydration are diluting MS medium minerals (Riffaud and Cornu, 1981; Letouze and
Daguin, 1983) and addition of phlorizin, phloroglucinol (Hegedus and Phan, 1983,1987)
or CoCl2 (Gaspar and Kevers, 1985) to the medium. The vitrification problem in liquid
medium could also be obviated by suppressing leaf growth with the addition of
paclobutrazol to the medium (Ziv, 1989; Ziv and Ariel, 1991). A number of studies have
been undertaken suggesting that not all the plant species show vitrification when cultured
on liquid medium. In some culture systems this problem can be avoided by taking
advantage of surface tension and floating explants, so that the developing tissue do not get
submerged in the liquid medium (Debergh et al., 1981; Skidmore et al., 1988; Dillen and
Buysens, 1989). In bioreactors also employing liquid medium, cultures impose stress
signals that are expressed in developmental abnormalities. Submerged tissue exhibit
oxidative stress, with elevated concentrations of reactive oxygen species, associated with a
changes in antioxidant enzyme activity. These changes may affect the anatomy and
physiology of the plants and their survival. Attempts to control hyperhydric deformations
have focussed on better aeration and intermittent plant submergence in the medium using
temporary immersion bioreactors (Teisson et al., 1996; Escalona et al., 1999; Etienne and
Berthouly, 2002).
Techniques that have the potential to further increase the efficiency of
micropropagation, but still awaits further improvements, include simplified large-scale
bioreactors, cheaper automation facilities, efficient somatic embryogenesis and synthetic
seed production, greater utilization of the autotrophic growth potential of cultures, and
good repeatability and quality assurance of the micropropagated plants. Cost reduction at
various stages of growth, development, multiplication and transfer of plantlets to the
greenhouse has also been dealt in detail (Levin et al., 1988).
2.2.10 Mass propagation of plants through bioreactors
Micropropagation is currently applied to a large number of agricultural and forestry,
medicinal and aromatic plant species, but is still costly due to intensive manual
manipulation throughout the various culture phases. Automation of micropropagation in

105
bioreactors has been advanced as a possible way to overcome the limitations imposed by
labour intensive methods and high production costs (Aitken-Christie, 1991; Takayama and
Akita, 1994; Leathers et al, 1995; Paek et al., 2001).
Progress in tissue culture automation will depend upon the use of liquid cultures in
bioreactors (Alper et al., 1993; Sakamoto et al., 1995). Liquid medium can provide much
more uniform growth environment, easy renewal without changing the containers,
membrane filteration for sterilization and ease for scaling up in large containers.
Employing bioreactors with liquid medium for large-scale production of horticultural,
forestry and medicinal plants either through cell, organogenic propagules e.g. meristem
tips, bulblets, corms, nodules, microtubers, shoot clusters (Takayama, 1986; Ziv, 1990;
Takahashi et al., 1992a, b; Akita et al., 1994; Ziv and Shemesh, 1996; Ziv and Lilien-
Kipins, 1997; Lim et al., 1998; Lee, 1999; Seon et al., 2000) or somatic embryos (Stuart et
al., 1987; Greidziak et al., 1990; Gupta et al., 1991; Terashima and Nishimura, 1991;
Denchev et al., 1992) is advantageous due to the ease of scaling-up the cultures in these
systems (Preil, 1991) and year round production of the propagules of useful plants
resulting in comparatively less labour cost and time (Levin et al., 1988; Preil et al., 1988).
Bioreactors are usually described in a biochemical context as self-contained, sterile
environment which capitalize on liquid nutrient or liquid/air inflow and outflow systems,
designed for intensive culture and affording maximum opportunity for monitoring and
control over microenvironmental conditions (agitation, aeration, temperature, dissolved
oxygen, pH etc.). The basic function of a bioreactor is to provide optimum physical and
chemical conditions, which influence the optimum growth of tissues in a high yielding
culture system. In general bioreactor-culture offers many advantages compared with agar-
based cultures including better control of the culture conditions; optimal supply of
nutrients and growth regulators, aeration and medium circulation, renewal of the culture
atmosphere as well as changing the medium during the culture period according to the
developmental stage, filtration of the medium for exudates, contamination control and
production of clusters of buds or somatic embryos for the automatic handling of the
propagules (Ziv, 2005). The use of bioreactors as a system for plant propagation through organogenic or
embryogenic pathway is presently being applied to several ornamental, vegetable, fruit
crop, medicinal and aromatic plants (Table-4). The various propagation aspects of several
plant species in bioreactors and some major problems associated with the operation of
bioreactors were recently reviewed (Takayama and Akita, 1998).

106
Table-4: Some important plants propagated in different types of bioreactors Plant species Propagable units produced Reference(s)
Allium sativum Multiple shoots, microbulbs, bulblets
Kim, 2002; Kim et al., 2004
Amaryllis hippeastrum Buds, bulblets Takayama and Akita, 1998
Ananas comosus Shoot clusters Escalona et al., 1999 Araceae spp. Plants Takayama and Akita, 1998
Artemisia annua Plants Park et al., 1989 Begonia spp. Multiple shoots Takayama, 1991 Callistephus hortensis Shoots Tisserat and Vandercook, 1985 Chrysanthemum spp. Shoots Hahn and Paek, 2005; Kim et al., 2005
Colocasia spp. - Takayama, 1991
Coffea arabica Shoot clusters, plants Teisson and Alvard, 1995
Crescentia cujete Shoot, plantlets Murch et al., 2004
Dianthus caryophyllus Shoots, plants Chatterjee et al., 1997
Dioscorea spp. microtubers Jova et al., 2005 Dieffenbachia spp. - Takayama, 1991 Fragaria ananasa Shoots, plants Takayama and Akita, 1998 Gentiana spp. Plants Hosokawa et al., 1998 Gladiolus grandiflorum Bud clusters, corms, plants Ziv, 1990; Ziv et al., 1998 Gloximia spp. Shoots Takayama, 1991 Hevea brasiliensis Buds, plants Teisson and Alvard, 1995
Hyacinthus orientalis Bulblets, plants Takayama and Akita, 1998 Lilium spp. Bulblets, plants Takayama and Akita, 1998; Kim et al., 2001;
Lian et al., 2003 Mallus spp. Shoots, plantlets Chakrabarty et al., 2003
Mitragyna inermis Shoots Tisserat and Vandercook, 1985
Musa spp. Buds, plants Alvard et al., 1993; Teisson and Alvard, 1995
Nephrolepis exaltata Buds, plants Levin et al., 1988; Ziv et al., 1998 Nerine samiensis Bulblets Ziv et al., 1994 Ornithogalum dubium Shoots, bulblets, plants Ziv and Lilien-Kipnis, 1997 Pinus radiata Shoots Aitken-Christie and Jones, 1987
Populus tremula Bud clusters, shoots, plants McCown et al., 1988
Pinellia ternata Shoots, corms Takayama, 1991 Pelargonium graveolense Shoots Katagi et al., 1986 Phalaenopsis Protocorm like bodies Park et al., 2000 Saccharum spp. Shoots Lorenzo et al., 1998
Solanum tuberosum Clusters, shoots, microtubers, Takayama and Akita, 1998 ; Ziv et al., 1998 ; Piao et al., 2003
Stevia rebaudiana Shoots Akita et al., 1994 Vitis spp. Shoots, plantlets Shim, 2002
Majority of the studies involving the use of bioreactor systems have followed the
concept of batch cultures. A schematic outline of mass propagation of certain
economically important plants using shake flask and bioreactor technique has been
proposed earlier (Bajaj, 1991; Kukreja and Ahuja, 1994) which involves the following
steps (a) establishment of aseptic cultures; (b) rapid growth and multiplication of apical
and axillary buds in shake flasks; (c) rapid growth and multiplication of shoot buds in

107
bioreactor culture; (d) root formation and hardening of the in vitro raised plants; (e)
acclimatization of the in vitro raised plants under field conditions.
2.2.10.1 Factors affecting culture of plant tissues in bioreactor
Various physical and chemical factors affecting the growth and proliferation of tissues in a
bioreactor culture vessel containing liquid medium have been studied in detail (Lee,
1997). To obtain normal and healthy plants, accurate monitoring of such factors is very
necessary during the growth phase of culture. These physical and chemical factors include
dissolved oxygen (DO) of the liquid medium, pH of the medium, inoculum density,
culture period, light and temperature conditions and the configuration and type of culture
vessel.
(a) pH of the culture medium
Lee (1997) studied the precise monitoring and controlling of pH changes during the
growth phase of culture and showed that it helped to control the changes of internal
physical and chemical factors which subsequently helped in improving plant growth.
Changes in pH during culture have also been reported by other workers (Dussert et al.,
1995; Yu et al., 2000; Lian et al., 2002). These changes appeared to be related to the
balance between ammonium in the medium as shown by several authors (Escalona et al.,
1999; Lian et al., 2002). Precise recording of fluctuations in parameters like pH in
computer controlled bioreactor cultures will improve the repeatability of complex
biological process.
(b) Dissolved oxygen
Oxygen concentrations in liquid cultures depend on the prescence of dissolved O2 in the
medium, the air bubbles inside the medium as well as O2 in the gaseous phase. The
dissolved oxygen content (DO) indicates the amount of oxygen available to the tissue in
the liquid medium. This available oxygen for plant cell is determined by oxygen transfer
coefficient (KLa values). As the tissue grows, the need for oxygen increases which can be
regulated by modifying operational parameters such as aeration rate, agitation speed, gas
flow and air bubble size, which helps in enhancing the availability of dissolved oxygen to
the tissue (Takahashi et al., 1992a, b; Drew, 1997; Lim et al., 1998; Seon et al., 2000). In
such cases oxygen enriched environment in bioreactor culture vessel leads to the better
plantlet growth.

108
Current interest towards photoautotrophich micropropagation revealed that carbon
dioxide and light enriched environment during the in vitro culture facilitates and supports
the shoot growth (Kozai et al, 1987; Infante et al., 1989). It has been observed that CO2
enrichment in a bioreactor culture vessel during the growth phase of shoots of sweet
potato, potato, Chrysanthemum and Chinese fox glove enhanced the growth and
production of healthy plantlets (Paek et al., 2001).
(c) Mixing or distribution of air
The other key parameter is mixing, which is necessary to distribute air equally to cells or
tissues, and nutrients throughout the liquid phase (Leathers et al., 1995; Sajc et al., 2000;
Honda et al., 2001). Mixing is normally carried out by sparging, mechanical agitation or a
combination of these two, but the magnitude of hydrodynamic forces associated with
mixing should be small enough not to cause cell or tissue damage, but sufficient to
stimulate selected function. The cultures growing in a bioreactor vessel containing liquid
medium exhibit the following conditions (Takayama, 1991): (a) floating just beneath the
medium surface (Strawberry plants) (Takayama et al., 1987); (b) freely moving in the
medium as in the case of Begonia and Gloxinia (Takayama and Misawa, 1981); (c)
sinking or submerged to the bottom of the vessel (Lilies, gladiolus and potatoes).
2.2.10.2 Bioreactor: basic configuration of culture vessel
As the bioreactors are the most suitable vessel for large-scale culture resulting in the
production of a large number of plantlets in one single batch, once the culture conditions
have been established at the shake flask level or in a small scale bioreactor of 250 ml to 1l
capacity, cultures can be easily manipulated in a large-scale bioreactor. The size and
various designs or configurations according to the test culture systems have been reported
by different workers (Katagi et al., 1986; Takayama, 1991; Teng et al., 1993; Akita and
Takayama, 1994). The basic configuration of bioreactors used for plant propagation is
similar to that of used for microbial, animal or plant cell cultures (Akita et al., 1994).
Standard configuration of a bioreactor includes bioreactor culture vessel, which provides
optimum space for the tissue to interact with the medium and to grow under aseptic
conditions. Impeller or agitator is attached to a rotating shaft, which is mechanically
driven. The main function of impeller is agitation and proper mixing of the medium in
order to supply ample oxygen and mineral substrates to the cultured tissue for growth.
Shoot and embryo cultures are more sensitive to damages caused by shear stress generated
by mechanical agitation. According to their growth habits, the multiple shoots generally

109
do not form dense clumps during growth where mass transfer is severly limited and
therefore the mechanical agitation is not always necessary for mass propagation of shoots
in bioreactors (Akita et al., 1994). Many other modifications in impellar design and their
use during shoot growth in the bioreactor culture vessel, such as non-mechanical agitation
(Chen et al., 1987; Stuart et al., 1987) or mechanical agitation (Ammirato and Styer, 1985;
Styer, 1985) have been studied. Aerator or sparger is a stainless steel tube with pin holes
provided at the base of the bioreactor vessel, through which compressed and filtered sterile
air is sparged in the medium. The type of sparger is important because the efficiency of
shoot and embryo growth and development depends upon the level of dissolved oxygen
and carbon dioxide. Even the size of bubbles produced by sparger holes are important as
their larger size along with the velocity of movement can cause damage to the growing
cultured tissue due to shear stress (Terashima and Nishimura, 1991; Jay et al., 1992). Air
line filter is an autoclavable disposable filter which allows to sparge sterile air into the
vessel. In addition to this an bioreactor unit has on line measurement of different culture
conditions i.e. pH, dissolved oxygen, temperature, gas flow rate, sugar level, foaming etc.
2.3.10.3 Types of bioreactors used for micropropagation
The design and operation of a bioreactor is determined by biological needs and engineering
requirements according to the concerned plant species and type of tissues, which often
includes a number of factors: aeration and mixing, low shear and hydrodynamic forces,
effective control of the physico-chemical environment and ease of scale-up (Paek et al.,
2005). The primary considerations of these bioreactors is to minimize tissue damage and
foaming, better conrols for maintainence of medium pH, dissolved O2 or CO2 concentration
in head space and medium rheology. Various types of bioreactors have been developed and
used in different laboratories for plant shoot culture. Generally they are modified in terms of
devices providing agitation and vessel configuration (Kessell and Carr, 1972; Wheat et al.,
1986; Chen et al., 1987; Asenjo and O’Carroll, 1988). Some of the major differences
between the bioreactors designed and used for secondary metabolite production and for
plant propagation deal with the media manipulations, the relatively large size of tissue
structures in the vessel, and in later case in contrast the relatively small anticipated scale of
production system (Vasil, 1991b). The different types of (Fig-3) bioreactors used for large-
scale plant propagation are presented in Table-5.

110
Fig.-3: Different types of bioreactors used for micropropagation

111
Table-5: Types of bioreactors generally used for plant propagation Types of bioreactors Configuration Reference(s) Mechanically agitated bioreactors
(a) Aeration agitation type bioreactors
(b) Rotating drum bioreactors
(c) Spin filter bioreactor (d) Stirred tank bioreactors
Kessel and Carr, 1972; Stuart et al., 1987; Preil et al., 1988 Tanaka et al., 1983 Styer, 1985; Wheat et al., 1986 Hooker and Lee, 1990; Lee, 1997
Pneumatically agitated bioreactor
(a) Simple aeration bioreactor
(b) Bubble column bioreactor
(c) Airlift bioreactor (d) Ebb and flow bioreactor (e) Balloon type bubble
bioreactor
Takayama, 1986, 1991 Lee, 1999 Paek et al., 2005
Non-agitated bioreactors (a) Gaseous phase bioreactor
(b) Overlay aeration bioreactor
Ushiyama et al., 1984a,b Ishibashi et al., 1987; Ushiyama, 1988
Some other bioreactor types have also been reported for plant propagation in which
light conditions have been developed in culture vessels (Ikeda, 1985). Inoue (1984)
described the use of transparent pipes fitted into the bioreactor through which light was
emitted. Ikeda (1985) reported an airlift bioreactor equipped with a photo inducing draft
tube, which consists of optical fibers introducing light into the reactor vessel.
2.3.10.4 Advantages and disadvantages of the use of bioreactors for
micropropagation
The major advantages of using bioreactor culture system for micropropagation of
economically important plants are the potential for scaling-up in lesser time, reduction in
the production cost as well as automated control of physical and chemical environment
during growth phase of the plant cultures. However, a lack of systematic and factorial
experimental knowledge about the interaction of plant physiology and physical parameters
of bioreactor design affects the frequent use of this technology (Paek et al., 2001).
Different designs and modifications in the agitation systems have made the technique
more advantageous.
Takayama and Akita (1994), Heyerdahl et al. (1995), Walker (1995), Lee (1997),
Sajc et al. (2000), Honda et al. (2001), Paek et al. (2001), Paek and Chakrabarty (2003)
reviewed different reactor configurations for plant cell suspensions, plant tissue and organ
cultures. The relative advantages and selection criteria for various reactor configurations

112
were discussed for specific process applications. Various types of simple bioreactors with
mechanically sparged or gas-sparged mixing were used in plant and cell culture to provide
stirring, circulation and aeration (Takayama, 1991; Scragg, 1992; Takayama and Akita,
1998; Ziv, 2000). Stirred tank bioreactor (STR) was designed to ensure the proper mixing
of medium and generating large sized bubbles. Mechanically stirred bioreactors depend on
impellers including a helical ribbon impeller (Archambault et al.,1994), magnetic stirrers
or vibrating perforated plates. The use of mechanically agitated stirrers resulted in an even
flow of the medium in different directions of the vessel which enhanced the proper
oxygenation of the cultured tissue (Schiigerl and Sitting, 1987; Hooker and Lee, 1990)
Numerous modifications of the conventional STRs with bubble aeration have been
developed that have a variety of impellar designs (Honda et al., 2001). The stirred tank
reacrtors (STR) present several limitations such as high power consumptions, high shear
forces, complicated configuration, problems with sealing and stabiliy of rotating shafts in
tall bioreactors, high contamination rates, difficulty in optimizing culture conditions etc.
Rotary drum reactors have significantly higher surface to volume ratio than other reactor
types. As a result mass transfer is achieved with comparably less power consumption. It
has been shown that mixing by gas sparging in bubble column or airlift bioreactors lacking
impellers or blades was far less damaging for clusters than mechanical stirring, since
shearing stress was minimized (Ziv and Hadar, 1991; Ilan et al., 1995; Ziv and Shemesh,
1996). In a bubble column bioreactor the bubbles create less shear forces so that it is
useful for plant organ cultures especially for propagation of various species through tissue
culture of shoots, bulbs, corms and tubers. Aeration, mixing and circulation in bubble
column or airlift bioreactors is provided by air entering the vessel from the side or from
the basal opening through a sparger. As the air bubbles rise they lift the plant biomass and
provide the require oxygen (Styer, 1985; Merchuk, 1990, Cazzulino et al., 1991; Preil,
1991). In case of airlift and column type bioreactors the positive points are simplicity of
design and construction, low shear stress, less energy requirements, while the
disadvantages are foaming induced by large volumes of air leading to impaired gas
transfer within the medium (Leathers et al., 1995; Lee, 1997) and growth of cells in the
head space. The phenomenon of foaming and cell growth on the wall of the vessel is due
to the diameter and top of the vessel being the same (Paek et al., 2001). Another problem
faced in airlift type bioreactors is the evaporation of culture medium. To overcome this
problem, addition of a sterile water column or condenser could be helpful. This can also
extend the cultivation period (Lee, 1997). To overcome the problems encountered during

113
the use of airlift and bubble column bioreactors, the bottom-type bubble bioreactor
(BTBB) was designed, in which by using a concentric tube for cell lifting at the vessel
base, foaming was drastically reduced. This bioreactor was found to be much reliable for
cell, tissue and organ culture of various species (Paek et al., 2001). In addition there was
provision of gas recycling system which allowed the examination of different gases in the
medium (Son et al., 1999). Disposable pre-sterilized plastic bioreactor (2-5 l) were used
for the proliferation of bud or meristematic clusters of several ornamental, vegetable and
woody plant species. The plastic bioreactor inuced minimal shearing and foaming
resulting in an increased biomass/proliferation as compared to the glass bubble-column
bioreactor (Ziv et al., 1998).
Attempts to control hyperhydric deformations have focussed on better aeration and
intermittent plant submergence in the medium by using temporary immersion bioreactors
(Teisson et al., 1996; Escalona et al., 1999; Etienne and Berthouly, 2002). The use of ebb
and flow type of bioreactor for the mass propagation of several plant species has overcome
a number of problems faced during the culture in other types of bioreactors. The principle
equipment in an ebb and flood bioreactor is the same as that in the BTBB. However in this
type, a supporting net was used to hold the plant material. This is a periodic immersion
system in which to avoid the complete submersion of explants in the liquid medium, a
fixed or floating support system has been provided inside the vessel, which initially helps
to hold the explant apart from the agitator (If provided) and later supports the growing
shoots. In this system medium is pumped from a storage tank into the culture vessel. A
series of channels help to supply nutrients evenly to the plant material resulting in an faster
and uniform growth. The medium remains in the vessel for a few minutes, after which it is
drained back to the storage tank for reuse. The drainage process is controlled by a solenoid
volve at different intervals depending upon the plant species and explant type (Paek et al.,
2005). This has eliminated the problems of those culture systems where different growth
phases need different compositions of nutrients in case of somatic embryo development.
Under these circumstances the ebb and flow type systems would be helpful because each
step can be separated from the other and can be easily monitored and controlled
independently (Merkle et al., 1995). In such a system each step of plantlet production can
be carried out in the same culture vessel simply by changing the cultute medium. Besides,
the system has the advantage of short term exposure of growing cultures to plant growth
hormones for the induction of different and desirable growth responses i.e. embryogenesis,
organogenesis or multiple shoot induction etc. This system completely eliminates regular

114
subculturing which is the most labour intensive and adds to the production cost of
micropropagated plants. Temporary immersion was used for banana, rubber and coffee
(Teisson and Alvard, 1995; Teisson et al., 1996; Etienne et al., 1997b), for tea (Akula et
al., 2000) and on a much larger scale for pineapple and sugarcane (Lorenzo et al., 1998;
Escalona et al., 1999). Temporary immersion was found to enhance potato microtuber
growth in a rotating bioreactor (Yu et al., 2000).
However, inconsistencies in optimizing bioreactor types and culture parameters
have been reported. Although the main source of these inconsistencies may be species
specific variations, attention is needed to interpret these results. So, once the culture
conditions have been established in a small-scale bioreactoer, cultures can be easily scaled
up in a large-scale (500-1000 l) bioreactor (Paek et al., 2005).
2.2.10.5 Automation in micropropagation
Current micropropagation techniques require a large number of culture vessels, media
preparation containers, manual media dispensing and labour intensive steps of explant
preparation, microshoot cutting and inoculation. The integrated management of whole
process as well as transplantation of plantlets for hardening, packaging, labelling and
transport further adds to the labour cost of the individual batches (Chu, 1995). While most
of the media preparation can be easily subjected to computerized automation (Ibaraki and
Kurata, 2001), the efforts towards automation of in vitro operation steps are still far from
adequate (Aitken-Christie et al., 1990). In this direction advancement has been made in
the application of robots (with or without vision system) for cutting and implantation of
explants in plant species with upright shoots, bulbous propagules and distinct nodes. The
robots employed in micropropagation industry today can recognize the node position
through a laser scanner attached to a image processing unit. The excising unit of the robot
then grips the shoots and cuts the nodal section through a single or multiknife cutter whose
position can be oriented in any desired direction and angle (Kozai et al., 1992a, b).
Two other areas of serious concern in automation of micropropagation through
bioreactor technology are contamination losses and sorting tools to identify quality
propagules. Researchers will have to develop protocols of shorter duration so that entire
multiplication cycle in bioreactors is completed in a short span of time. Alternatively,
bioengineers must take provision for continuous periodic filteration of the medium to
minimize microbial growth within the culture vessel (Levin et al., 1997). Offline analysis

115
of changes in nutrient and hormone concentrations during bioreactor culture will present
new possibilities for the better manipulation of embryogenesis and organogenesis.
Systems using image sorting principles are urgently required to isolate quality
micropropagtion units on the basis of their size, shape, texture, colour and density which if
done manually involves high labour inputs. A comparision of the external morphology
with the anatomical and biochemical maturity of the propagules can also provide
important tools to determine their developmental or establishment competence. For
evaluating all these parameters through image analysis of contour, skeleton and geometric
features of the micropropagules, several algorithm based computer programmes are being
developed (Sharaki, 1991).
Mechanization of transplanting steps of the micropropagation protocol is another
area of focus to reduce the production cost. Though prototypes of transplanting robots
based on fuzzy theory have been prepared to transplant plants of right developmental stage
but success rate during hardening is still low.
2.2.11 PCR-based Random Amplified Polymorphic DNA technique for molecular
characterization of the in vitro raised plants
Random Amplified Polymorphic DNA technique is a modified PCR based technique
which overcomes the drawbacks of the selective gene amplification as a method for
genetic analysis. The modification involves the use of a single short primer (8-12
nucleotides long unlike regular PCR where primer length ranges between 20-40
nucleotides) of arbitrary nucleotide sequence or selected at random, in the amplification
reaction (Welsh and McClelland, 1990; Williams et al., 1990). In this technique, a single
primer of arbitrary sequence anneals to the genomic DNA at two different sites on
opposite strands and if these primer annealing sites are within an amplifiable distance of
each other, discrete products on genomic size and homology between primers and
tempelate DNA sequence are produced. Unlike traditional PCR anlysis, RAPD does not
require any specific knowledge of the nucleotide and amino acid sequence data of the
target organism for the design of amplification primers because the primer sequences are
arbitrarily defined. The primers will or will not amplify a segment of DNA, depending on
positions that are complementary to the primer’s sequence. In RAPD technique, the
segments of DNA that are amplified, are random. The differences between two genotypes
due to mutations that inhibit primer annealing or creates newer stable primer annealing
sites and/or due to the insertion/ deletion of sequences, are also reflected as differences in

116
the length of amplification products. This is the basis of determining the genetic variability
or similarity by this PCR based RAPD technique (Samee, 1993; Tingey and Tufo del,
1993).
Investigation of the genetic relationship among plant species, varieties, cultivars
and hybrids require recent DNA technologies. Random Amplified Polymorphic DNA
(RAPD) technique of DNA fingerprinting is capable of providing various markers
(Williams et al., 1990) which can be used for the estimation of genetic relationship as well
as the cultivar identification or differentiation among various plant species. The technique
has been used in cultivar identification of many plants (Koller et al., 1993; Stiles et al,
1993; Yang and Quiros, 1993). There are also reports on the use of RAPD in
chracterization of soybeans (Lark et al., 1992, Paiva et al., 1994; Prabhu and Gresshoff,
1994). This technique can be successfully used for the assessment of genetic similarity as
well as for the detection of somaclonal variations among the in vitro regenerated plants
(Gavida et al., 1996; Khanuja et al., 1998; Raina et al., 2001; Modgil et al., 2005; Lu et
al., 2007; Palombi et al., 2007; Sreedhar et al., 2007). The technique of RAPD described
by Williams et al. (1990) in comparision to other DNA based techniques (AFLP, RFLP) is
faster, less expensive and known to be more efficient alternative as it is able to reveal the
polymorphism at greater extent as well as the RAPD profiles are independent of age,
physiological status, tissue source or environmental impressions. The success of any
micropropagation protocol dependes upon the production of large number of true-to-type
plants. Various physical and chemical factors during in vitro culture of plants can involve
mutation inducing processes during explant establishment, maintenance and callus
induction (Lorz et al., 1988). These can induce variations in cultured cells and tissues and
they should be detected at the molecular level. The genetic uniformity of many
micrpropagated plants have been established on the basis of RAPD technique (Rani et al.,
1995; Rani and Raina, 2002; Carvalho et al., 2004; Ray et al., 2006). In general RAPD
can be applied to differentiate subspecies, varieties, ecotypes and cell lines. RAPD
markers are well suited for genetic mapping for breeding applications and for DNA
fingerprinting with particular utility for studies of population genetics. Previously Lattoo
et al. (2006) reported true-to-type character of micropropagated C. arundinaceum plants
raised through multiple shoot bud induction and growth employing RAPD analysis.
In the present study, an attempt has been made to establish a method for the in
vitro propagation of C. borivilianum through multiple shoot regeneration by shoot base
culture using different culture conditions and thereafter the molecular characterization of

117
the randomly selected in vitro raised plants has been performed by RAPD analysis to
establish their genetic uniformity.
2.2.12 Chemistry of the genus Chlorophytum
Traditionally roots of different Chlorophytum species are reported to possess various
pharmacological utilities having saponins as one of the important phytochemical
constituents. Saponins consist of sugar moiety, usually containing glucose, galactose,
glucuronic acid, xylose, rhamnose or methylpentose, glycosidically linked to hydrophobic
aglycone (sapogenin) which may be triterpenoid or steroid in nature. The oligosaccharide
chain is normally attached at C3 position (monodesmosidic) but many saponins have an
additional sugar moiety at the C26 or C28 positions (bidesmosidic). Kaushik (2005) in his
review summarized the work done on chemical investigation of various Chlorophytum
species with specific reference to saponins.
(a) C. malayense
C. malayense is indigenous to South-East Asia and South-West of Yunnan province of
China. Li et al. (1990) isolated four steroidal saponins from C. malayense rhizome by
methanolic extraction followed by n-butanol partitioning to get crude saponin and
subsequent separation on silicagel column chromatography. These four saponins, termed
as chloromaloside-A, B, C and D have neo-hecogenin and neotigogenin as the aglycone
moiety with various substitutions of sugar moiety. Chloromaloside-A, C and D belong to
25-S spirostane series, while chloromaloside-B is found to be furostane type.
Chloromaloside-A isolated as colourless needle, is the major saponin of C. malayense. In a
bioassay guided fractionation, chloromaloside-A showed broad cytotoxicity against
various human cancer cell lines (Qiu et al., 2000). A new steroidal saponin named as
chloromaloside-E has been isolated (Yang and Yang, 2000) having neohecogenin as
aglycone.
(b) C. comosum
C. Comosum is native to South Africa and is used for the treatment of bronchitis fracture
and burns as part of traditional medicine in China. It is commonly known as ‘Spider
Plant’. Seven anti-tumour promoter steroidal saponins have been isolated from crude
saponin obtained by partioning of methanol extract with n-butanol from this species
(Mimaki et al., 1996). The saponins of C. comosum are different from saponins of C.
malayense having aglycone based on 25-R-spirostan series as tigogenin, gitogenin and

118
hecogenin, while saponins of C. malayenese are based on 25-S spirostan series as
neotigogenin and neo-hecogenin. The isolated saponins have been evaluated for in vitro
anti tumour promoter activity by measurement of the inhibitory activity on TPA-
stimulated 32P incorporation into phospholipids of HeLa cells as a primary screening test
to identify new anti tumour promoter compounds (Mimaki et al., 1996).
(c) C. arundinaceum
C. arundinaceum is found in India and is reported to possess adaptogenic activity (Tandon
and Shukla, 1995b). Gupta et al. (1979) isolated galactoglucan from the fruits of C.
arundinaceum. Tuberous roots also contain sugars (arabinose, glucose and xylose). Four
sapogenins- stigmasterol, tigogenin, neotigogenin and tokorogenin-identified on the basis
of mass spectral fragmentation, were isolated from this species (Tandon and Shukla,
1992). The same group further identified several compounds viz., nonacosane,
tetracosanoic and triacontanoic acid, 4-hydroxyl-8, 11-oxidoheneicosanol (a disubstituted
tetrahydrofuran) and pentacosyl docosanoate (Tandon et al., 1992). 2, 2', 4, 4'-tetrahydro
bibenzyl xyloside has also been reported from the same species for the first time in the
family Liliaceae (Tandon and Shukla, 1993). Later they also reported two new saponins,
arundinoside A and arundinoside B in the roots of this species. (Tandon and Shukla,
1995a, 1997).
(d) C. borivilianum
C. borivilianum is commonly known as ‘safed musli’ in India. Its roots are widely used for
various therapeutic applications in various systems of medicine (Oudhia and Tripathi,
1999). It has spermatogenic property and is found useful in curing impotency. It is also
reported to cure diabetes, arthritis and increasing general body immunity. However, in
recent years its effectiveness in increasing male potency has become very popular and is
now considered as an alternative to ‘Viagra’. This species has not been investigated
properly for its phytochemical constituents. Recently, Narasimhan et al. (2006) reported
(2/<1) fructo/oligopolysaccharide fraction of C. borivilianum having antidiabetic and
antioxidant activity. Govindarajan et al. (2005) also reported in vitro antioxidant activity
in ethanolic extract of C. borivilianum. The roots are reported to contain 42% of
carbohydrates, 8-9% of proteins, 3-4% of fibers, and 2-17% of saponins (Bordia et al.,
1995).

119
(e) C. tuberosum
Narasimhan et al. (2006) found that this species of Chlorophytum possess antioxidant and
free radical scavenging activity.
(f) C. laxum
Gao et al. (2005) reported a new labdane diterpene glycoside (chlorophytoside-A) from C.
laxum.
(g) C. inomatum
O’ Donnell et al. (2006) isolated a new homoisoflavanone [3/(4'/methoxybenzyl)/7,8/
methylene dioxy/chroman/4/one] and another metabolite 7/(1'/hydroxyethyl)/2/
(2''/hydroxyethyl)/3,4/dihydrobenzopyran and reported that antibacterial activity in this
plant is mainly due to the prescence of homoisoflavonoid.
(h) C. nimonii
Isolation of tigogenin pentaglycoside from aerial parts of C. nimonii as well as its use in
antihyperglycemic and hypolipidemic activities have been reported (Lakshmi et al., 2007).
2.2.13 Tissue culture studies in Chlorophytum species
2.2.13.1 In vitro shoot cultures
A few studies on the in vitro tissue culture aspects of C. borivilianum have been
conducted. Amongst various species of Chlorophytum, three species viz. C. comosum, C.
borivilianum and C. arundinaceum have been investigated in detail. In vitro multiplication
of C. comosum was achieved by apical meristem culture (Apollonio et al., 1990). The
stem-disc posed a serious problem of contamination whenever used as explant to raise
cultures (Apollonio et al., 1990; Purohit et al., 1994a; Suri et al., 1998).
In vitro clonal multiplication of C. borivilianum was reported by Purohit et al.
(1994a). In vitro cultures were established from stem discs (possessing shoot buds and
young shoot bases) obtained from field-grown plants. In further experiments shoot bases
were used as a source of initial explants because of less contamination rate as compared to
stem discs. MS medium supplemented with 22.2 µM BAP was found best for shoot
proliferation. Shoots multiplied at a rate of four-fold every three weeks on this medium.
Individual shoots separated from clusters were trimmed from the top leaving only 5 mm at
the base. Such shoots were subcultured on fresh medium as secondary explants.
Rhizogenesis was observed in shoots when they were transferred to MS medium with ¾

120
strength inorganic and organic constituents and 9.8 µM IBA. Four weeks old plantlets
were transferred to pots containing a soil-soilrite mixture (1:1, v/v). These pots were
initially maintained under high humidity conditions in mist chambers and were finally
exposed to external environment. Sixty seven percent of the micropropagated plants were
successfully acclimatized in pots. Such established plants produced normal fasciculated
storage roots similar to their wild parents. The entire procedure could be completed
without intermediate callus formation. None of the plants showed any phenotypic
variation. The basic MS salt formulation was found to be best among different nutrient
media (MS, B5 media) tested during shoot multiplication.
Later Suri et al. (1998) also reported in vitro shoot multiplication in C.
borivilianum using stem discs as explants. In their studies, B5 medium (Gamborg et al.,
1968) supplemented with BAP (2.5-5.0 mgl-1) supported shoot proliferation from the stem
disc explants. Cultures were maintained on B5 medium with 5.0 mgl-1 BAP. Higher
concentrations (>7.5 mgl-1) were inhibitory for shoot growth and multiplication. B5
medium supplemented with IBA (0.25 mgl-1) was optimal for root induction in isolated
shoots.
Dave et al. (2003a) reported strategies for scaling-up production and evaluation of
field performance of micropropagated safed musli plants. Young shoot buds proliferating
from tuberous roots attached to stem discs of the identified nursery-grown plants served as
the source of explant. Surface-sterilized explants were inoculated on MS medium
containing 22.2µM BAP. Proliferated shoots were subsequently transferred to the same or
modified fresh medium on every three weeks of interval for multiplication. In their studies
they also observed best response in terms of shoot multiplication on MS medium
supplemented with 22.2 µM BAP and 3% sucrose as previously reported by Purohit et al.
(1994a). Amongst different gelling agents used, phytagel at 0.2% showed slightly better
response than BDH agar in terms of shoot multiplication, but the use of BDH agar was
preffered due to its low cost. Based on experiments they concluded that the optimal (>3.5
fold) shoot multiplication rate could be achievd on MS medium supplemented with 22.2
µM BAP, 0.8% standard BDH agar and 3% sucrose. For induction of rooting the shoots
were inoculated on modified rooting medium i.e. MS medium (MS salts and vitamins
reduced to three-quarters) supplemented with 0.8% agar, 3.0% sucrose and 9.8 µM IBA
(Purohit et al., 1994a). Root induction could be uniformly obtained after 7 days of
inoculation of shoots while roots suitable for soil transfer developed in 21 days. Plantlets
subjected to hardening under shade conditions during the monsoon months of high

121
humidity showed better survival rate and growth compared to plantlets hardened in vitro
and subsequently transferred to the greenhouse for acclimatization. Plantlet survival rate
was 87% and 90% under openfield and shade conditions, respectively. Plantlets grown ex
vitro under agro-shadenet and field conditions produced tuberous roots. The tubers of
previous years were used as propagules for field plantations next season. Multi-location
trials of in vitro raised plants showed similar performance in different field conditions.
Pudake and Dhumale (2003) also reported in vitro shoot multiplication of C.
borivilianum through shoot base and stem disc culture. Shoot multiplication was achieved
on MS medium supplemented with 2.0 mgl-1 BAP. Among the different combinations of
IBA or NAA tested for rooting, IBA (3.0 mgl-1) was found to be relatively better. Up to
90% plantlets were established in pots following in vivo hardening.
Dave et al. (2003b) achieved in vitro propagation of safed musli using
encapsulated shoot buds to save time and labour required for regular subculture. Four mm
long shoot buds encapsulated in 3.0% sodium alginate matrix polymerized by 100 mM
solution of hydrated calcium chloride gave best results. Subsequently plants were
produced from these buds. Storage conditions, gel matrix medium and duration of storage
influenced the in vitro potential of these encapsulated shoot buds when transferred to the
standard multiplication medium containing MS salts and 5.0 mgl-1 BAP. Encapsulated
shoot buds stored on wet agar-gel and kept under culture room conditions of light and at
28°C temperature showed more than 80% sprouting within 3 weeks. Encapsulated shoot
buds stored in dark at 4ºC temperature on agar-gelled wet medium showed more than 90%
sprouting after 7 days of storage. Encapsulated buds sprouted readily while decline in
sprouting was observed with increase in storage period irrespective of the light or dark
conditions used. Supplementing alginate matrix with sucrose and MS salts produced better
results as compared to that with or without MS salts. All the sprouted shoot buds
irrespective of their storage conditions, produced normal shoots on shoot multiplication
medium and recording 2.5-fold increase/subculture after 21 days. More than 70%
microshoots could be rooted on the medium containing 3/4 strength MS salts and 2.0 mgl-1
IBA (Purohit et al., 1994a).
Lattoo et al. (2006) reported an in vitro multiplication system via multiple shoot
bud induction and regeneration in C. arundinaceum using shoot crown explants possessing
juvenile adventitious shoot buds. Optimum regeneration frequency (87%) and
organogenetic response in the form of de novo organized multiple shoot buds without an

122
intervening callus phase was obtained on MS medium supplemented with 3% sucrose
(w/v), 4x10-6 M Kn and 2x10-6 M IBA. Shoot number per explant averaged 2.77 after eight
weeks of culture passage. Axenic secondary explants with multiple shoot buds on
subculturing exhibited best response on MS medium containing 1x10-5 M Kn and 5x10-6M
IBA, giving rise to an average of 18.74 shoots per culture. It was observed that the shoot
bud differentiation and regeneration was influenced by the ratios of cytokinins/auxin
rather than their relative concentrations. Four week old in vitro regenerated shoots
developed roots in half strength MS basal medium containing 3% sucrose (w/v)
supplemented with 5 x 10-6 M IBA. Four week old in vitro rooted plants after hardening in
controlled environment chamber for three weeks were directly transpalnted in earthen pots
in sand-soil mixture (1:1; v:v) under field conditions with more than 90% success. Plants
maintained under field conditions developed normal fascicualted roots. Regenerated plants
were also free of any noticeable phenotypic variability.
Sharma and Mohan (2006) described in vitro shoot regeneration from immature
floral buds along with inflorescence axis in C. borivilianum. MS medium supplemented
with 2.0 mgl-1 Kn and 0.1 mgl-1 2, 4-D proved best for multiple shoot induction. Maximum
number of shoots were obtained on MS medium containing 2.0 mgl-1 BAP. Rooting of
shoots (86.7%) with maximum fasciculated roots occurred on Knop’s medium containing
iron and vitamins of MS medium with 2.0 mgl-1 IBA and 0.1% activated charcoal. Plants
hardened in four weeks and 80% survival rate was reported.
Bhardwaj and Murali (2006) patented the protocol of in-vitro mass culture of C.
borivilianum through pieces of root top with crown meristem. These explants were
inoculated on culture initiation medium comprising of half strength MS medium for
induction of multiple shoots. The shoot cultures were transferred to proliferation and
elongation medium comprising of half strength MS medium supplemented with 0.05% to
0.5% of BAP. The multiple shoots in the range of 3-9 shoots per explant were obtained.
The elongated shoots were transferred to phytohormone free half strength MS medium for
rooting. They reported upto 100% survival rate of in vitro regenerated plants.
Rizvi et al. (2007a) reported efficient and cost-effective micropropagation of C.
borivilianum employing shoot base explants and liquid culture medium supplemented with
BAP (5.0 mgl-1) and reported about 7.0 to 7.5-fold increase in total number of shoots per
flask against only 4.5-fold increase on agar-solidified medium. In vitro regenerated plants

123
exhibited 80% survival rate. Use of liquid culture medium resulted upto 92.31% reduction
in shoot production cost compared to semi-solid medium.
2.2.13.2 In vitro tuberous root formation
Suri et al. (1998) studied in vitro tuberous root formation in C. borivilianum. Plantlets
produced 3-4 tuberous roots when kept in vitro for 2-3 more months after induction of
fibrous roots on in vitro regenerated shoots. These plantlets were maintained on a filter
paper bridge in liquid B5 medium containing IBA (0.1 mgl-1) and sucrose (60 gl-1). Fresh
liquid medium was added at a regular interval of 15 days. Such plantlets, on transfer to the
soil showed 100% survival in their studies, and after overwintering, sprouted in the
following season. According to Suri et al. (1998) tuberous roots are produced de novo
from the stem on attaining maturity and not formed by tuberization of fibrous roots.
Organogenic cultures have been maintained in the laboratory for the past six years without
loss of any regenerative potential.
2.2.13.3 Root culture in liquid medium
Arora (1999) made attempts to grow isolated roots of C. borivilianum in culture as an
alternate to natural roots. Roots were initiated from callus culture and germinating
embryos. Increase in root biomass was obtained by inoculating these roots to 1 l flasks and
2l stirred tank bioreactor containing MS medium supplemented with 500 mg N/l [100 mg
N/l (NH4)2SO4 and 400 mg N/l KNO3] and 0.1 mgl-1 each of Kn and NAA, but the roots
grew as thin fibrous roots and no thickening of the roots was observed on any of the
treatments used.

124
2.3 MATERIALS AND METHODS
2.3.1 Plant material: Maintenance of mother stock plants
Plants of Chlorophytum borivilianum (Family-Liliaceae) were collected from natural
habitat (Rajsamand district near Udaipur) in Rajasthan and maintained in earthen pots
containing mixture of sand, soil and farmyard manure (1:1:1, v/v). These plants were used
as the mother source for obtaining explants for tissue culture experiments. These stock
plants were maintained under glass house conditions at 60-70% relative humidity and
25±5°C temperature.
2.3.2 Initiation of aseptic cultures
2.3.2.1 Chemicals
All chemicals and phytohormones used during the entire study were procured from M/s
British Drug House (BDH), India; M/s Fine Chemicals, India; M/s Hi-Media Laboratories
Pvt. Ltd., India; M/s Merck Pvt. Ltd., India; M/s Qualigens India; M/s Sigma Aldrich
Chemicals, USA and M/s Sisco Research Laboratories (SRL) Pvt. Ltd., India.
2.3.2.2 Glassware and plasticware
‘Borosil’, ‘Corning’ or ‘Schott Duran’ make glasswares were used for all experiments.
Tarsons (India) Pvt. Ltd. make plasticwares were also used. Erlenmeyer flasks (100, 150,
250 and 500 ml), both wide mouth and narrow mouth and culture tubes (25x150 mm)
were mainly used as culture vessels. Besides this screw cap glass bottles (100-500 ml),
glass petri dishes (90 mm), beakers (50-1000 ml), measuring cylinders (25-1000 ml),
pipettes of various capacity (0.01-10 ml), micropipettes (0.5-1000µl), funnels and glass
rods were also used. Disposable plastic petri-dishes were also used in the present study.
All the plasticwares and glasswares used in media or solution(s) preparation were initially
washed with a liquid detergent (Teepol) and then thoroughly washed with tap water to
remove traces of detergent. The washed glassware and plasticwares were then rinsed with
double distilled water. Washed and rinsed glasswares were kept in oven at 90-100°C for 3-
4 hrs for drying.
2.3.2.3 Sealing devices
Parafilm ‘M’®(American Can. Co.) obtained from Sigma Aldrich Corporation, USA was
used for sealing purpose wherever required.

125
2.3.2.4 pH meter and weighing balance
pH meter (Control Dynamics Systems Pvt. Ltd., India) and weighing balance (Metller-
Toledo International Inc., USA) were used during media preparation and for preparing
other solutions as per requirement. Besides this microwave oven (IFB Industries Ltd.,
India) water purification unit (Millipore Corporation, France) and horizontal laminar air
flow (Micro Flit, India) were also used at different stages of media preparation and/or
inoculation.
2.3.2.5 Nutrient media
MS medium (Murashige and Skoogs, 1962) and its modified formulations were used in
the present investigation. Details of the basal media composition are provided in Table-25.
Stock solutions of different medium constituents were prepared and stored in a refrigerator
at 4°C. For preparing semi-solid medium, 0.8% (w/v) agar was used as gelling agent. In
liquid medium agar was excluded. Medium was supplemented with 3% (w/v) sucrose and
0.1% (w/v) myoinositol. The phytohormones were added to the medium according to their
requirement throughout the experiments. The heat labile phytohormones were filter
sterilized and added to the medium after filter sterilization. The pH of both semi-solid and
liquid media was adjusted to 5.8±0.1 using 0.1 N HCl and/or 0.1 N NaOH prior to steam
sterilizing at 121°C temperature and 15 lbs pressure for 20 minutes using
horizontal/vertical sterilizer (M/s Nat Steels Pvt. Ltd., Mumbai).
2.3.2.6 Preparation of stock solutions of phytohormones
The stock solutions of different phytohormones used in the present study were prepared at
the concentration of 1.0 mg/ml (w/v) by dissolving 100 mg of plant hormone in 0.5 to 1.0
ml of 0.1 N NaOH or 0.1 N HCL or EtOH (depending upon the nature of solubility of
plant hormone) and final volume was made to 100 ml by adding sterile distilled water.
These stock hormone solutions were stored in a refrigerator at 4°C and were used for
different experiments according to their requirement.
2.3.3 Establishment and maintenance of mother stock cultures
2.3.3.1 Sterilization and preparation of explants
In vitro cultures were initiated from young shoot buds proliferating from tuberous roots
attached to the stem disc of these glass house grown plants. Stem disc portion containing
shoot buds was separated from tuberous roots and cut into pieces of stem disc each
containing 1-2 shoot buds. These explants were washed thoroughly in a gentle continuous

126
flow of tap water for 2-3 hrs and later surface sterilized with 0.1% (w/v) mercuric chloride
(HgCl2) solution for 4-6 min. Following surface sterilization explants were rinsed 4-5
times with sterile distilled water. These explants were transferred aseptically under a
clean air laminar hood on semi-solid medium fortified with different phytohormone
concentrations as mentioned in results.
2.3.3.2 Culture conditions
All aseptic cultures were maintained in a culture room having controlled environmental
conditions i.e.16 hrs photoperiod (16:8 hrs light: dark alternating regime), 3000-4000 lux
intensity provided by cool white fluorescent tubes, 25±2°C incubation temperature and
about 60% relative humidity.
2.3.3.3 Multiplication of propagable units
After 3-4 weeks of incubation on the culture medium, shoots proliferated from shoot buds.
Individual shoot or a bunch of 2-3 shoots were excised from the cluster of in vitro
regenerated shoots and trimmed from the top leaving about 1.0-1.5 cms portion along with
shoot base part. Such prepared explants were used throughout the study. Shoots which
proliferated from these explants were subcultured regularly on the same fresh medium
every 3-4 weeks for further multiplication. A bunch of 2 shoots (1.0-1.5 cms portion of
shoots) along with a part of shoot base was used as initial inoculum in all experiments
except where number of shoots was mentioned.
2.3.4 Establishment of aseptic cultures in shake flasks
Explants having 1.0-1.5 cms shoot length along with a part of shoot base obtained from
regular sub-culturing of mother stock cultures maintained on MS semi-solid medium
supplemented with 5.0 mgl-1 BAP were transferred to liquid MS medium (without agar) of
the same composition. Thirty ml of liquid medium was dispensed per 250 ml wide mouth
Erlenmeyer flask and cultures were kept for 40 days on a New Brunswick Scientific
Rotary Shaker (USA) at 70 rpm speed as well as in static condition. All the cultures were
incubated in a culture room under 16 hr photoperiod (16:8 hrs light:dark alternating
regime), 25±2°C temperature and about 60% relative humidity.
2.3.5 Rhizogenesis in the in vitro raised plantlets
Root induction in in vitro regenerated shoots is an essential step involved in
micropropagation. For rooting, shoots regenerated in vitro on semi-solid as well as liquid
medium were seperated from the cluster of in vitro regenerated shoots and transferred to

127
the rooting medium. Various concentrations of different auxins as mentioned in results
were supplemnted to the MS medium for rooting of in vitro raised plants. Different
strengths of the MS medium as mentioned in the results were also tested for their effect on
root induction and growth.
2.3.6 Hardening of the in vitro raised plants under glass house conditions
In vitro rooted plantlets growing under different in vitro culture conditions i. e. plants
growing in agar-solidified medium and plants growing in liquid medium shake flaks were
gently washed under tap water to remove agar based semi-solid medium or liquid medium
without damaging the delicate root system. Plants were hardened under culture room
conditions by transferring to test tubes containing distilled water for 15-20 days and then
carefully transferred to earthen pots containing a mixture of sand:soil:farmyard manure
(1:1:1, v/v) for further hardening.
2.3.7 Analysis of data
In all experiments standard deviation was calculated by using following formula:
√ n ∑ χ2 – ( ∑ χ )
n (n-1)
Where,
n = number of replicates observed; ∑ = summation; χ = observation of the replicate
2.3.8 Mass propagation of C. borivilianum plants in bioreactor
2.3.8.1 Bioreactor configuration
In the present study experiments for large-scale culture of C. borivilianum plants were
done by using a bench top/stirred bioreactor (model Bio Flow-110, M/s New Brunswick
Scientific, USA) having working volume of 5.0 litres. The impeller in this air sparged top
driven system provided mechanical agitation. The bioreactor consisted of a thick glass
jacketed vessel assembly having inner vessel with removable baffle assembly consisting of
different probes including dissolved oxygen (DO), pH and temperature probes to control
and optimize various culture conditions. The inner vessel assembly was surrounded by an
outer thick glass jacket. The external jacket was used as a water temperature controller.
The whole glass culture vessel rests on a double walled stainless steel base through which
cold water circulates and maintains the temperature of the culture medium. An exhaust gas
condenser was installed to the unit for minimizing the medium evaporation during culture
period. The refrigerated water (temperature 4°C) was circulated through double walled

128
stainless steel condenser with the help of a refrigerated water bath circulator. All parts of
the bioreactor assembly were thouroughly washed and then surface sterilized with ethyl
alcohol prior to assembling the unit and lubricated with silicone grease to make the unit air
tight. About 2.5 litre liquid MS medium supplemented with 5.0 mgl-1 BAP, 3% (w/v)
sucrose and 0.1% (w/v) myo-inositol (pH 5.8±0.1) was poured in the culture vessel.
During the experiments, an autoclavable nylon mesh (pore size 200 µM) tightened just
beneath the medium surface on the lower stainless steel semi-circular ring of the baffle
assembly prevented the explants from sinking to the bottom or getting submerged in the
medium. The explants remain floating near the medium surface with the support of mesh
during the culture period. The incoming air was passed within the medium through air
sparger after sterilization by a hydrophobic membrane filter (Whatman, USA, 0.22 µ).
This complete unit was properly assembled and autoclaved at 120°C and 15 lbs pressure
for 25 minutes in a vertical cylindrical sterilizer (M/s Yarco, India).
2.3.8.2 Inoculation of the bioreactor vessel
Inoculum, consisting of one week old in vitro regenerated shoots along with shoot base
part (2-3 cms in length) were transferred from Erlenmeyer liquid medium shake flasks to
the sterilized reactor culture vessel under a clean air laminar air flow. The head plate of the
reactor was cleaned and surface sterilized using alcohol and inoculation port was
thoroughly heated/flamed before and after opening. Approximately 120 explants were
transferred through inoculation port with the help of a sterile forecep. Culture conditions
included 16:8 hrs alternating light and dark regime, 25±2°C temperature and 60% relative
humidity. After inoculation, culture vessel unit was installed on the main control module.
Water lines were connected to the water jacket inlet and to the exhaust condenser. All air
and/or gas lines were also reconnected. The pH and Do2 cables were connected to their
respective probes and to the pH and DO2 control module. The temperature probe was
reinstalled into the thermowell. Air sparger was connected to air compressor through a
silicone tubing fitted with 0.22 µ Whatman filter.
2.3.9 Molecular analysis of plants obtained from different in vitro culture
conditions through PCR based Random Amplified Polymorphic DNA technique
For determining genetic similarity between in vitro raised plants and wild type (control) at
molecular level, their RAPD analysis was performed. In this technique high molecular
weight DNA was isolated and subsequently this DNA was amplified through PCR using

129
primers of known sequences. The protocol for DNA isolation (Khanuja et al., 1999),
amplification and analysis is described below:
2.3.9.1 DNA isolation of selected plants
Approximately 2.0 to 2.5 g of tissue from young leaves was properly washed with sterile
distilled water and grinded into fine powder in liquid nitrogen. The fine powder thus
obtained from each sample was transferred to sterile oakridge tube and 5.0 ml of freshly
prepared and pre-warmed (60°C for half an hour) extraction buffer was added to it and
gently mixed by inversion into slurry (details of buffer are presented in Table-6). The
oakridge tubes having samples were then incubated at 65°C for one hour in a water bath.
After one hour incubation, they were taken out from water bath and 5.0 ml of chloroform:
isoamyl alcohol (24:1 respectively) was added and gently mixed by inversion for about
10-15 min to form an emulsion. Tubes were then centrifuged at 10,000 rpm for 10 min at
24° C. After centrifugation three layers were formed: upper aqueous layer containing high
molecular weight nucleic acid and dissolved polysaccharides, middle layer contains cell
rests and dissolved green cell walls while lower layer contains chloroform with dissolved
proteins, lipids and chlorophylls. The upper aqueous layer from each tube was transferred
to a fresh sterile oakridge tube followed by addition of 2.0 ml of 5 M NaCl solution to
each tube and mixed properly. After this 3.0 ml isopropanol (0.6 volume of the total
solution) was added to it. The mixture was kept for one hour at room temperature. After
one hour slow and careful mixing will produce fibrous nucleic acid. The mixture was then
centrifuged at 10,000 rpm for 10 min at 24°C. The supernatant was discarded while the
pellet (containing DNA) was washed with 800 µl of 80% ethyl alcohol. This mixture was
then centrifuged again at 10,000 rpm for 5 min at 24° C. Supernatant was discarded while
the pellet was vacuum dried for 10-15 min and dissolved in 750 µl high salt TE (Table-6)
by gentle mixing. Then 10µl of RNAse was added to each tube and tubes were incubated
at 37° C for half an hour. Then entire solution was transferred into sterile eppendorf tubes
and 750 µl of chloroform: isoamyl alcohol (24:1) was added to it and mixed gently. The
tubes containing mixture were then centrifuged at 10,000 rpm for 10 min at 24°C and the
upper aqueous layer was transferred to a fresh sterile eppendorf tube while lower layer
was discarded. This process was repeated and to the upper aqueous layer thus obtained
double volume of 100% cold ethanol was added. After gentle mixing the mixture was kept
at -20° C for 2 hrs. After 2 hrs, the tubes containing mixture were centrifuged at 10,000
rpm for 10 min at 4°C. Supernatant was discarded while the pellet was vacuum dried for

130
10-15 min. Depending upon the pellet size, the vacuum dried pellet was then dissolved
into 50-100µl of sterile triple distilled (Milli- Q) water and stored at 4°C. The DNA
concentration was quantified by running aliquotes on 0.8% agarose gel (M/s Hi-Media
Laboratories Pvt. Ltd., India) as well as in spectrophotometer (Nanodrop ND-1000,
U.S.A.) at 260 nm absorbance.
Table-6: Details of the buffers used for isolation of DNA
Extraction buffer (2xCTAB) 100ml High salt TE buffer (100ml) NaCl (Sodium chloride) 1.5M NaCl 1.0M
CTAB (Cetyl trimethylammonium bromide)
2.5% Tris 100Mm (pH 8.0)
Tris 100mM (pH 8.0)
EDTA 10mM (pH 8.0)
EDTA
(Ethylenediamine tetraacetic acid)
20mM (pH 8.0)
ß-mercaptoethanol
0.2% (v/v)
PVP (Polyvinylpyrrolidone) 1% (w/v)
2.3.9.2 PCR amplification reactions
Polymerase chain reactions were performed in a total volume of 25 µl with each reaction
tube containing 25 ng of DNA, 0.6 units Taq DNA polymerase, 10mM of each dNTPs
(ATP, GTP, CTP, TTP), 1.5 mM MgCl2 buffer and 5 pmol of decanucleotide primers.
Twenty different primers MAP 01 to MAP 20 procured from M/s Bangalore Genei (India)
were used. The PCR amplification reactions were carried out using DNA engine thermal
cycler (MJ Research, USA) as per following protocol :
Step 1: Denaturation at 94°C for 5min.
Step 2: Denaturation at 94°C for1min.
Annealing at 35°C for1min.
Extension at 72°C for 2 min.
Step 3: Final extension at 72°C for 5 min.
Step 2 is repeated 45 times
After completion of reaction, the PCR product was stored at -20°C. The amplified DNA
was loaded on 1.2% agarose gel containing 0.5 µg/ml ethidium bromide and after
completion of gel electrophoresis, photographed on a polaroid system. The polymorphic

131
bands were analyzed and the graphic dendrogram of the genetic relationship between the
in vitro raised plants and their control type was produced.
A set of 20 primers (MAP01 to MAP20) with the sequences as described below
were used to study DNA polymorphism for determining the similarity index (degree of
similarity) among the 14 randomly selected in vitro raised plants of C. borivilianum along
with parent plant. These randomly selected plants were obtained from the various
experimental conditions i.e. plants obtained from cultures maintained on semi-solid and
liquid media. Plants were grown and maintained in glass house conditions along with
control plant (parent plant).
The sequences of twenty different primers used in the present study are as follows (Table-7):
Table-7: The nucleotide sequences of primers used for RAPD analysis
Code Sequence (5' to 3') Code Sequence (5' to 3')
MAP 01 AAATCGGAGC MAP 11 CCCTGCAGGC
MAP 02 GTCCTACTCG MAP 12 CCAAGCTTGC
MAP 03 GTCCTTAGCG MAP 13 GTGCAATGAG MAP 04 TGCGCGATCG MAP 14 AGGATACGTG MAP 05 AACGTACGCG MAP 15 AAGATAGCGG
MAP 06 GCACGCCGGA MAP 16 GGATCTGAAC
MAP 07 CACCCTGCGC MAP 17 TTGTCTCAGG MAP 08 CTATCGCCGC MAP 18 CATCCCGAAC
MAP 09 CGGGATCCGC MAP 19 GGACTCCACG
MAP 10 GCGAATTCCG MAP 20 AGCCTGACGC

132
2.4 RESULTS
2.4.1 Establishment of aseptic cultures from shoot base
explants
Shoot base explants of Chlorophytum borivilianum were cultured on semi-solid MS basal
medium fortified with various levels (1.0-10.0 mgl-1) of different cytokinins i.e. Kn, BAP,
combinations of Kn and BAP, 2iP or Z. All cytokinins (Kn, BAP, 2iP or Z) independently
or in combinations viz. Kn+BAP, were supplemented to the MS basal medium at different
levels i.e. 1.0, 3.0, 5.0, 7.5 or 10 mgl-1. AS was supplemented at 1.0, 3.0, 5.0, 7.5, 10, 25,
50 or 100 mgl-1 levels. Kn and BAP each were supplemented to the MS medium in equal
concentration. TDZ was added at 0.1, 0.2, 0.5, 1.0, 2.0 or 3.0 mgl-1. Similarly, different
auxins i.e. IAA, IBA, NAA, 2,4-D or P were also added individually in the range of 0.05-
2.0 mgl-1 to the MS basal medium for selecting optimal medium for direct shoot
regeneration and growth from shoot base explants. MS basal medium without any plant
hormones served as control during these experiments.
Establishment of aseptic cultures was difficult because the explants were used from
underground tuberous roots (Plate-1a) of field grown plants. Around 50-60% aseptic
cultures were observed in initial experiments. On MS medium containing different
cytokinins the explants swelled during the initial culture period and inoculated shoot bases
initiated shoot primordia at all the concentrations within 18 days of inoculation (Plate-1b).
Shoot regeneration and growth was observed on all media combinations as well as on
control medium. Though the morphogenetic response in inoculated shoot bases was
observed on all media combinations, the frequency of morphogenetic response and
number of shoots regenerated varied depending upon the type and concentrations of
different hormones added to the medium. The data for various growth parameters such as
number of shoots/explant, shoot length (cms) and number of roots/explant in the
regenerated shoots were recorded after 30 days of incubation of shoot bases on MS basal
medium fortified with different concentrations of various cytokinins (Table-8, 9, 10).
Best response in terms of multiple shoot bud induction and shoot regeneration was
observed on MS basal medium supplemented with 0.1 mgl-1 TDZ (Table-10) followed by
medium containing 5.0 mgl-1 BAP (Table-8). On MS medium containing TDZ (0.1 mgl-1)
an average of 18.4 shoots per explant were regenerated after 30 days of culture. An

133
average number of 10.4 shoots could be obtained on medium supplemented with 3.0 mgl-1
level of TDZ.
MS medium supplemented with BAP (5.0 mgl-1) during the same culture period
could support regeneration of an average number of 11.4 shoots per explant (Plate-1d).
Amongst all cytokinins tested viz. Kn, BAP, Kn+BAP, 2-iP, Z or AS it was observed that
BAP at levels ranging between 1.0-7.5 mgl-1 exhibited best response. Though TDZ (0.1
mgl-1) showed better response than BAP ( 5.0 mgl-1) in terms of shoot regeneration (Plate-
1c) but shoot growth and length were comparatively more in BAP supported medium as
compared to TDZ containing media combinations. At optimal level of TDZ (0.1 mgl-1) for
multiple shoot regeneration, an average shoot length of 0.74 cms was recorded compared
to optimal level of BAP (5.0 mgl-1) where an average length of 2.0 cms was recorded
(Fig.-4). BAP levels above 5.0 mgl-1 were not effective for multiple shoot regeneration
and shoot growth. In case of medium supplemented with kinetin at 5.0 mgl-1, an average
number of 7 shoots/explant were regenerated, while maximum 2.48 cms length of shoots
was recorded. Less number of shoots regenerated on the above mentioned level of kinetin
than on medium fortified with same level of BAP, on the other hand shoot growth was
better in kinetin supplemented medium but callus formation was observed at the base. 2-iP
also induced multiple shoot formation and the response was somewhat similar as that on
kinetin supplemented medium. At 5.0 mgl-1 level of 2-iP, the number of shoots
regenerated was same as that on medium supplemented with same level of kinetin, but
shoot length was lesser (1.46 cms) as compared to kinetin supplemented medium (2.48
cms). In order to improve upon the shoot growth and length, Kn was added to the culture
medium with BAP in equal proportions at different concentrations. The medium although
supported shoot growth but shoot multiplication response was low compared to the
medium supplemented with BAP as the sole phytohormone. Besides, on the BAP and
kinetin supplemented medium callusing response was observed from the base of
regenerated shoots. The multiple shoots produced on 2-iP fortified medium were
comparatively less in number than shoots produced on BAP fortified medium and shoot
length was also less than recorded on Kn+BAP combinations. Higher level of cytokinins
(>7.5 mgl-1) could not evoke optimal morphogenetic response and shoot regeneration was
less in case of all cytokinins tested. Amongst all cytokinins tested Z or AS were least
effective at all levels. Zeatin (Z) was effective at lower levels and at 3.0 mgl-1
concentration could induce regeneration of an average number of 3.8 shoots while AS was

134
effective at higher levels and an average number of 4 shoots were obtained at 10 mg l-1
level of AS (Table-8, 9).
Table-8: Effect of cytokinin supplementation on the growth response of cultured
shoot bases of C. borivilianum after 30 days of culture in MS medium
S. No.
Growth parameter(s)
Cytokinin (mgl-1)
Different cytokinin(s) tested
Control* Kn BAP Kn+BAP† 2iP Z
1. No. of shoots/ explant
1.00 6.00± 0.70** 5.00 ± 1.22 8.80 ± 1.30 7.00 ± 1.00 6.00 ± 1.00 3.20 ± 0.45
Shoot length (cms)
2.26 ± 0.38 2.00 ± 0.49 1.19 ± 0.20 3.30 ± 0.43 2.54 ± 0.76 1.60 ± 0.23
No. of roots/explant
1.40 ± 0.54 1.00 ± 0.00 - 1.00 ± 0.00 2.60 ± 0.54 1.20 ± 0.83
2. No. of shoots/ explant
3.00 5.60 ± 1.34 10.60 ± 1.14 9.40 ± 1.94 5.80 ± 1.09 3.80 ± 0.84
Shoot length (cms)
1.51 ± 0.37 0.90 ± 0.11 3.78 ± 0.87 1.73 ± 0.37 2.10 ± 0.32
No. of roots/ explant
1.20 ± 0.44 - - 1.80 ± 0.44 1.40 ± 0.54
3. No. of shoots/ explant
5.00 7.00 ± 1.58 11.40 ± 1.14 9.00 ± 1.58 7.00 ± 1.00 2.20 ± 0.44
Shoot length (cms)
2.48 ± 0.37 2.00 ± 0.12 2.98 ± 0.57 1.46 ± 0.18 1.40 ± 0.18
No. of roots/ explant
1.00 ± 0.00 1.40 ± 0.54 1.00 ± 0.00 1.40 ± 0.54 -
4. No. of shoots/ explant
7.50 5.00 ± 1.00 9.00 ± 1.22 5.00 ± 0.70 7.00 ± 1.14 2.00 ± 0.00
Shoot length (cms)
1.90 ± 0.20 1.36 ± 0.08 2.50 ± 0.36 1.48 ± 0.26 0.90 ± 0.20
No. of roots/ explant
1.00 ± 0.00 - - - -
5. No. of shoots/ explant
10.00 2.60 ± 0.54 1.40 ± 0.54 2.00 ± 0.00 3.40 ± 0.54 1.60 ± 0.55
Shoot length (cms)
1.50 ± 0.15 1.10 ± 0.20 2.00 ± 0.27 2.20 ± 0.16 0.70 ± 0.23
No. of roots/ explant
- - - - -
*MS medium without phytohormones; ** Average value ± SE (standard error); † Kn and BAP in equal amounts; - No
response
Table-9: Effect of different levels of AS in MS medium on the morphogenetic response
of cultured shoot bases of C. boriviianum after 30 days of culture
S. No.
Growth parameter(s)
AS concentration (mgl-1) 1.00 3.00 5.00 7.50 10.00 25.00 50.00 100.00
1. No. of shoots/explant
2.60± 0.55* 2.80± 0.45 2.40± 0.54 3.00± 0.00 4.00± 0.71 2.20± 0.84 1.80± 0.44 1.60± 0.55
2. Shoot length (cms)
1.66 ± 0.21 1.86± 0.34 1.48± 0.24 2.20± 0.40 2.42± 0.45 1.60± 0.31 1.34± 0.23 1.38± 0.18
3. No. of roots/explant
1.20 ± 0.83 1.00± 0.00 - 1.00± 0.00 1.40± 0.55 - - -
* Average value ± SE (standard error); - No response
Table-10: Effect of TDZ supplementation in the MS medium on the growth response
of cultured shoot bases of C. borivilianum after 30 days of culture
S. No.
Growth parameter(s)
TDZ concentration (mgl-1) 0.10 0.20 0.50 1.00 2.00 3.00
1. No. of shoots/ explant 18.40 ±1.51* 2.00 ± 0.00 4.00 ± 1.14 2.00 ± 0.00 3.60 ± 0.54 10.40 ± 1.51

135
2. Shoot length (cms) 0.74 ± 0.02 1.00 ± 0.00 1.29 ± 0.16 1.00 ± 0.00 1.45 ± 0.13 0.75 ± 0.12 3. No. of roots/explant - - - - 2.00 ± 0.00 -
* Average value ± SE (standard error); - No response
MS basal medium without any hormones served as control medium in the present
study, supported an average number of 6 shoots per explant and also supported shoot
growth but morphogenetic response in terms of shoot regeneration and growth was not
comparable to that observed on MS medium fortified with 5.0 mgl-1 BAP (Table-8).
Thus, MS basal medium fortified with 5.0 mgl-1 BAP supported optimal shoot
regeneration although regenerated shoots were relatively smaller in size than those
obtained on medium either supplemented with Kn alone or combinations of Kn+BAP
but no intervening callus phase was observed. On the other hand callusing response was
observed on media supplemented with Kn alone or combinations of Kn and BAP. Media
containing different levels of Kn, Kn+BAP, 2iP, Z or AS although responding in terms
of shoot regeneration and growth could not be used for further experiments due to low
shoot regeneration rate in general as compared to media supplemented with 5.0 mgl-1
BAP (Fig.-4).
0
2
4
6
8
10
12
14
16
18
20
Control† Kn (3) BAP (5) Kn+BAP(5)
2-iP (5) Z (3) AS (10) TDZ (0.1)
No. of shoots / explant Length of shoots (cms)
Fig.-4: Effect of different cytokinins (optimal level of each cytokinin in mgl-1) on shoot multiplication and
growth from shoot base explants of C. borivilianum after 30 days of culture; † MS basal medium without
phytohormones.
Thus, for the growth parameters tested for shoot regeneration and growth in the
present study viz. number of shoots and shoot length MS basal medium supplemented with
5.0 mgl-1 BAP was found to be most suitable and without inducing the callusing response it
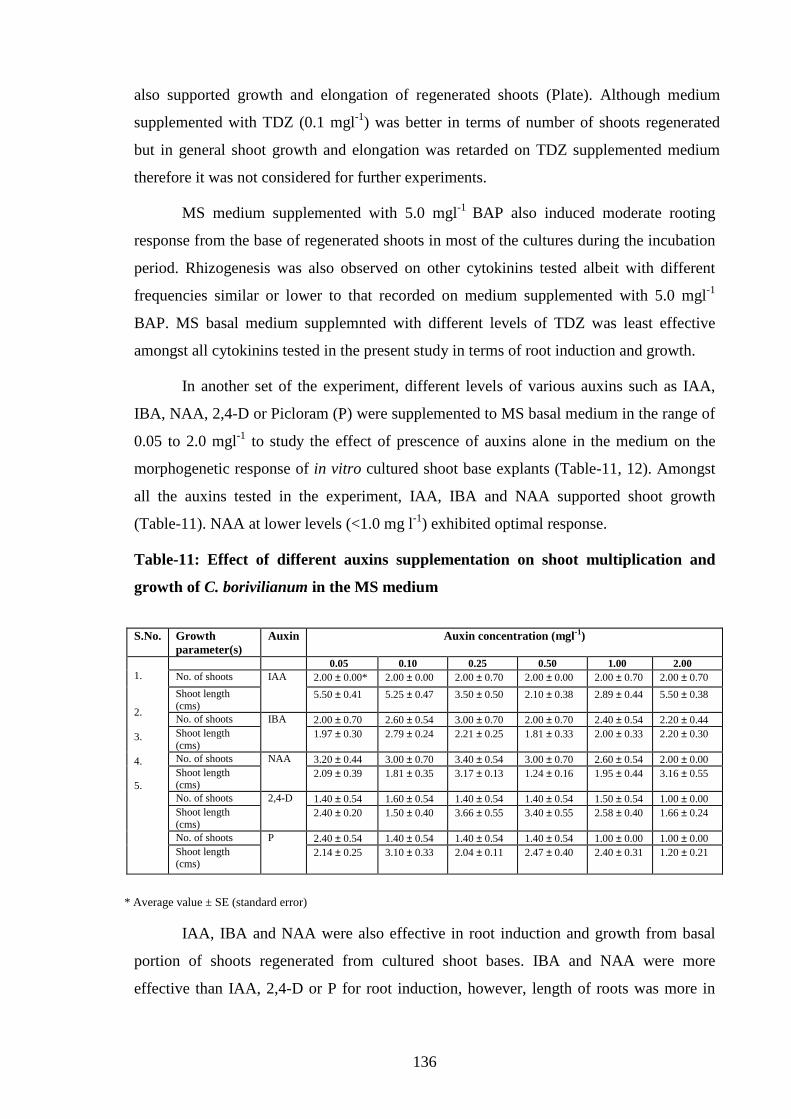
136
also supported growth and elongation of regenerated shoots (Plate). Although medium
supplemented with TDZ (0.1 mgl-1) was better in terms of number of shoots regenerated
but in general shoot growth and elongation was retarded on TDZ supplemented medium
therefore it was not considered for further experiments.
MS medium supplemented with 5.0 mgl-1 BAP also induced moderate rooting
response from the base of regenerated shoots in most of the cultures during the incubation
period. Rhizogenesis was also observed on other cytokinins tested albeit with different
frequencies similar or lower to that recorded on medium supplemented with 5.0 mgl-1
BAP. MS basal medium supplemnted with different levels of TDZ was least effective
amongst all cytokinins tested in the present study in terms of root induction and growth.
In another set of the experiment, different levels of various auxins such as IAA,
IBA, NAA, 2,4-D or Picloram (P) were supplemented to MS basal medium in the range of
0.05 to 2.0 mgl-1 to study the effect of prescence of auxins alone in the medium on the
morphogenetic response of in vitro cultured shoot base explants (Table-11, 12). Amongst
all the auxins tested in the experiment, IAA, IBA and NAA supported shoot growth
(Table-11). NAA at lower levels (<1.0 mg l-1) exhibited optimal response.
Table-11: Effect of different auxins supplementation on shoot multiplication and
growth of C. borivilianum in the MS medium
S.No. Growth parameter(s)
Auxin Auxin concentration (mgl-1)
1. 2. 3. 4. 5.
0.05 0.10 0.25 0.50 1.00 2.00 No. of shoots IAA 2.00 ± 0.00* 2.00 ± 0.00 2.00 ± 0.70 2.00 ± 0.00 2.00 ± 0.70 2.00 ± 0.70
Shoot length (cms)
5.50 ± 0.41 5.25 ± 0.47 3.50 ± 0.50 2.10 ± 0.38 2.89 ± 0.44 5.50 ± 0.38
No. of shoots IBA 2.00 ± 0.70 2.60 ± 0.54 3.00 ± 0.70 2.00 ± 0.70 2.40 ± 0.54 2.20 ± 0.44 Shoot length (cms)
1.97 ± 0.30 2.79 ± 0.24 2.21 ± 0.25 1.81 ± 0.33 2.00 ± 0.33 2.20 ± 0.30
No. of shoots NAA 3.20 ± 0.44 3.00 ± 0.70 3.40 ± 0.54 3.00 ± 0.70 2.60 ± 0.54 2.00 ± 0.00 Shoot length (cms)
2.09 ± 0.39 1.81 ± 0.35 3.17 ± 0.13 1.24 ± 0.16 1.95 ± 0.44 3.16 ± 0.55
No. of shoots 2,4-D 1.40 ± 0.54 1.60 ± 0.54 1.40 ± 0.54 1.40 ± 0.54 1.50 ± 0.54 1.00 ± 0.00 Shoot length (cms)
2.40 ± 0.20 1.50 ± 0.40 3.66 ± 0.55 3.40 ± 0.55 2.58 ± 0.40 1.66 ± 0.24
No. of shoots P 2.40 ± 0.54 1.40 ± 0.54 1.40 ± 0.54 1.40 ± 0.54 1.00 ± 0.00 1.00 ± 0.00 Shoot length (cms)
2.14 ± 0.25 3.10 ± 0.33 2.04 ± 0.11 2.47 ± 0.40 2.40 ± 0.31 1.20 ± 0.21
* Average value ± SE (standard error)
IAA, IBA and NAA were also effective in root induction and growth from basal
portion of shoots regenerated from cultured shoot bases. IBA and NAA were more
effective than IAA, 2,4-D or P for root induction, however, length of roots was more in

137
IAA fortified media combinations. Although IAA, IBA or NAA were effective in root
induction but IBA and NAA supported thickening of roots also (Table-12).
Picloram (P) or 2,4-D were least effective amongst all auxins tested. In case of
media supplemented with different levels of 2,4-D or P, except 0.05 mgl-1 level of both the
auxins other levels of either auxin did not support rhizogenesis from in vitro cultured
shoots. Callusing was observed at the basal portion of shoots cultured on 2,4-D
supplemented media. Amongst all auxins tested NAA at 0.25 mgl-1 level induced optimal
root regeneration (an average number of 19.6 roots per explant) but with callusing at basal
region. While IBA (2.0 mgl-1) supplemented MS basal medium could induced an average
number of 16.6 roots per explant without any intervening callus phase (Fig.-5). Shoot
regeneration on 2,4-D or P supplemented media was repressed and was not comparable to
that observed on IAA, IBA or NAA supplemented media.
In a separate experiment NAA was supplemented to optimal shoot regeneration
medium (MS basal medium containing 5.0 mgl-1 BAP) in the range of 0.05-2.0 mgl-1to
have both shoot multiplication and growth and root induction responses in the same
culture medium. Both shoot regeneration and growth were comparatively less to that
observed on MS medium fortified only with BAP (5.0 mgl-1). Besides, callusing response
was also observed from the basal portion of shoot bases cultured in vitro on the medium
containing combinations of BAP and NAA.
Thus, based on number of roots/explant, length of roots, number of roots showing
thickening/tuberization and number of roots having secondary rooting, MS medium
fortified with IBA at 2.0 mgl-1 was found optimal and roots regenerated directly without
any callus formation. Although more roots were produced on NAA fortified media, the
roots were shorter and callusing was observed at the base of roots. IAA, IBA or NAA
tested at various levels were effective for shoot growth, root induction and growth but they
could not induce the shoot multiplication as observed previously on media supplemented
with different cytokinins.

138
Table-12: Effect of auxin(s) on root induction and growth in C. borivilianum after 30 days of culture in MS medium
S. No.
Growth parameter(s) Auxin Auxin concentration (mgl-1)
0.05 0.10 0.25 0.50 1.00 2.00
1. Root induction (days) IAA 10.00 9.00-12.00 10.00 11.00 11.00 11.00
No. of roots/explant 4.33 ± 0.89* 3.40 ± 1.14 2.60 ± 0.54 5.00 ± 1.22 9.00 ± 1.00 6.00 ± 1.00
Length of roots (cms) 4.66 ± 0.80 5.86 ± 1.00 4.76 ± 0.71 2.44 ± 0.33 2.81 ± 0.46 3.26 ± 0.35
No. of thick roots - - - - - -
No. of roots showing secondary roots - 1.60 ± 0.54 - - - -
2. Root induction (days) IBA 11.00 10.00 12.00 12.00 12.00 12.00
No. of roots/ explant 13.00 ± 2.07 14.20 ± 1.30 15.60 ± 1.81 9.60 ± 1.51 8.00 ± 1.58 16.60 ± 1.94
Length of roots (cms) 1.42 ± 0.04 1.33 ± 0.07 1.49 ± 0.08 0.91 ± 0.14 1.07 ± 0.14 1.59 ± 0.24
No. of thick roots 3.80 ± 0.83 3.80 ± 0.83 4.40 ± 0.54 2.60 ± 0.54 1.20 ± 0.44 3.80 ± 0.83
No. of roots showing secondary roots 1.80 ± 0.44 1.60 ± 0.44 1.60 ± 0.54 1.60 ± 0.54 1.00 ± 0.00 2.00 ± 0.70
3. Root induction (days) NAA 9.00 9.00 11.00 12.00 12.00 12.00
No. of roots/explant 13.00 ± 1.58 14.60 ± 2.30 19.60 ± 1.81 15.40 ± 1.81 10.00 ± 1.87 1.40 ± 0.54
Length of roots (cms) 1.42 ± 0.08 0.56 ± 0.09 0.97 ± 0.15 0.87 ± 0.09 0.50 ± 0.00 0.50 ± 0.00
No. of thick roots 4.60 ± 0.89 3.00 ± 0.70 3.40 ± 0.54 2.50 ± 0.89 1.00 ± 0.00 -
No. of roots showing secondary roots 1.60 ± 0.54 - - - - -
4. Root induction (days) 2,4-D
13.00 - - - - - No. of roots/explant 1.40 ± 0.54 - - - - -
Length of roots (cms) 1.20 ± 0.19 - - - - -
No. of thick roots - - - - - -
No. of roots showing secondary roots - - - - - - 5. Root induction (days) P 13.00 - - - - -
No. of roots/explant 1.00 ± 0.00 - - - - - Length of roots (cms) 2.80 ± 0.73 - - - - - No. of thick roots - - - - - - No. of roots showing secondary roots - - - - - -
* Average value ± SE (standard error); - No response

99
Different strengths of MS basal medium supplemented with different levels of IBA
(0.05-2.0 mgl-1) were also tested to standardize best nutritional requirements for root
induction and growth (Table-13). In this experiment, strength of basal MS medium was
reduced to half and three-fourth of the normal. It was observed that three-fourth strength
of MS salts gave better response than half-strength MS salts and an average number of 15
roots/shoot with an average length of 1.5 cms were obtained on three-fourth strength MS
medium supplemented with 2.0 mgl-1 IBA (previously standardized for rhizogenesis)
against an average number of 12 roots per shoot with an average length of 1.27 cms on
half-strength MS medium supplemented with same level of IBA.
Thus, for all parameters tested viz. number of roots/explant, length of roots,
number of roots showing thickening/tuberization, number of roots showing secondary
rooting, three-fourth strength MS medium supplemented with 2.0 mgl-1 IBA gave best
response. Based on these experiments it can be concluded that MS basal medium fortified
with 5.0 mgl-1 BAP supported optimal shoot regeneration and growth while three-fourth
strength MS medium supplemented with IBA (2.0 mgl-1) is optimal for root induction in in
vitro regenerated shoots.
Table-13: Effect of IBA supplementation in MS medium (half and three-fourth
strength) on root induction in cultured shoots of C. borivilianum after 30 days of culture
S. No. Strength of medium
IBA conc. (mgl-1)
Growth parameter(s)
No. of roots/ explant
Length of roots (cms)
No. of thick roots
No. of roots showing
secondary roots
1. Half -strength 0.05 6.00 ± 1.00* 1.01 ± 0.06 1.00 ± 0.00 1.00 ± 0.00 0.10 8.00 ± 1.58 0.95 ± 0.09 1.00 ± 0.00 1.00 ± 0.00 0.25 11.00 ± 1.58 1.21 ± 0.03 1.60 ± 0.54 1.00 ± 0.00 2.00 12.00 ± 1.87 1.27 ± 0.07 1.20 ± 0.44 1.00 ± 0.00
2. Three-fourth 0.05 10.60 ± 1.51 1.36 ± 0.08 2.60 ± 0.54 1.20 ± 0.44 0.10 11.80 ± 2.16 1.28 ± 0.14 2.20 ± 0.44 1.20 ± 0.44 0.25 14.00 ± 1.87 1.40 ± 0.08 3.40 ± 0.54 1.60 ± 0.54 2.00 15.00 ± 1.58 1.50 ± 0.08 3.00 ± 0.70 1.80 ± 0.44
* Average value ± SE (standard error)

100
0
5
10
15
20
25
IAA (1) IBA (2) NAA (0.25) 2,4-D (0.05) P (0.05)
No. of roots / explant Root length (cms)
Fig.-5: Effect of different auxins (optimal level of each auxin in mgl-1) on root induction and growth in C.
borivilianum after 30 days of culture period
The healthy shoots regenerated on standardized shoot multiplication medium after
30 days of culture were transferred to rooting medium (three-fourth MS salts fortified with
2.0 mgl-1 IBA). About 2.5-3.0 cms long shoots were found to be suitable for the purpose
of rooting. The fibrous adventitious roots originated from basal portion of in vitro
regenerated shoots. Root induction could be uniformly obtained after 10-12 days of
inoculation of shoots on standard rooting medium while roots suitable for soil transfer
developed in 25-30 days.
The plantlets obtained after four weeks of culturing on root induction medium
were washed gently with tap water to remove sticking agar carefully so that roots are not
damaged. Thereafter, they were transferred to test tubes containing distilled water for two
weeks for culture room hardening and subsequently transplanted to the earthen pots
containing a mixture of sand, soil and farm yard manure in 1:1:1 (v/v) ratio under glass
house conditions. The plantlets removed from culture vessels and transferred directly to
pots under natural environmental conditions could not withstand the initial transplantation
shock and died within 8-10 days. Potted plants were covered with transparent polythene
bags with small holes for air ventilation and to ensure high humidity in initial stages.
Inside the green house, relative humidity (RH) was maintained at 80-90% and temperature
was around 28±2°C. During first week the plants were kept close to the cooling source (air
conditioners) and then they were gradually shifted away. The polythene bags were
removed after two weeks. In vitro regenerated plants exhibited 78-80% survival rate. In
this study the entire procedure could be completed without callus formation. It is an

101
advantage since callus formation may induce genetic variability. Regenerated plants were
free of any noticeable phenotypic variability.
2.4.2 Effect of liquid culture medium on shoot multiplication and growth in C.
borivilianum
In C. borivilianum shoot base is the basic culturing unit which is used for regeneration of
multiple shoots. Multiple shoots grew from shoot primordia induced on shoot base
explants used in the present investigation. Therefore, these shoot bases can be used as
propagable units in C. borivilianum. The objective of the present investigation was to
maximize the production of such propagable units. Use of liquid culture medium for
multiplication of C. borivilianum will lower down the production cost of in vitro
regenerated plants. To develop a protocol for large-scale multiplication of propagable
units i.e. shoot bases their culture in bench top bioreactor is desirable. It was therefore
necessary to use and test the efficiency of liquid culture medium as only liquid culture
medium can be employed in bioreactor culture vessel. To achieve this experiments were
performed to study the effect of liquid culture medium on different growth parameters of
C. borivilianum. In our earlier experiments amongst different cytokinins (Kn, BAP, 2-iP
or Z) tested for their efficacy on in vitro shoot multiplication and growth at a range of 1.0
to 10.0 mgl-1, BAP at 5.0 mgl-1 exhibited optimal response. Therefore, MS basal medium
containing BAP (5.0 mgl-1) with (treated as control) or without agar was used. Two shoot
base explants (inoculum density) were inoculated per culture flask. A comparative study
for the growth of C. borivilianum in semi-solid and liquid culture media was carried out
by inoculating shoot bases from in vitro grown plants maintained on shoot multiplication
medium (Table-8). A clump of 2-3 shoot base explants, each measuring about 1.0-1.5 cms
were inoculated in semi-solid and liquid medium of the same composition. All the cultures
were incubated under uniform culture conditions. The liquid cultures were incubated in
static or agitated (75 rpm) conditions on a rotary shaker (Plate-2). The morphogenetic
response in terms of number of shoots/flask, mean length of shoots, number of
roots/explant, length of roots, total biomass/culture flask (f. wt. and dry wt.) were recorded
in both liquid and semi-solid media after 40 days of culture period (Table-14). The
observations recorded clearly revealed that liquid culture (agitated) supported better shoot
multiplication and growth of regenerated shoots than semi-solid medium (Plate-3a). An
average of 15 shoots/explant (7.5-fold increase over two shoots/flask inoculum density)
could be obtained in agitated liquid medium against 9 shoots/explant (4.5-fold increase) in
semi-solid medium during the same culture duration. A significant difference in the length
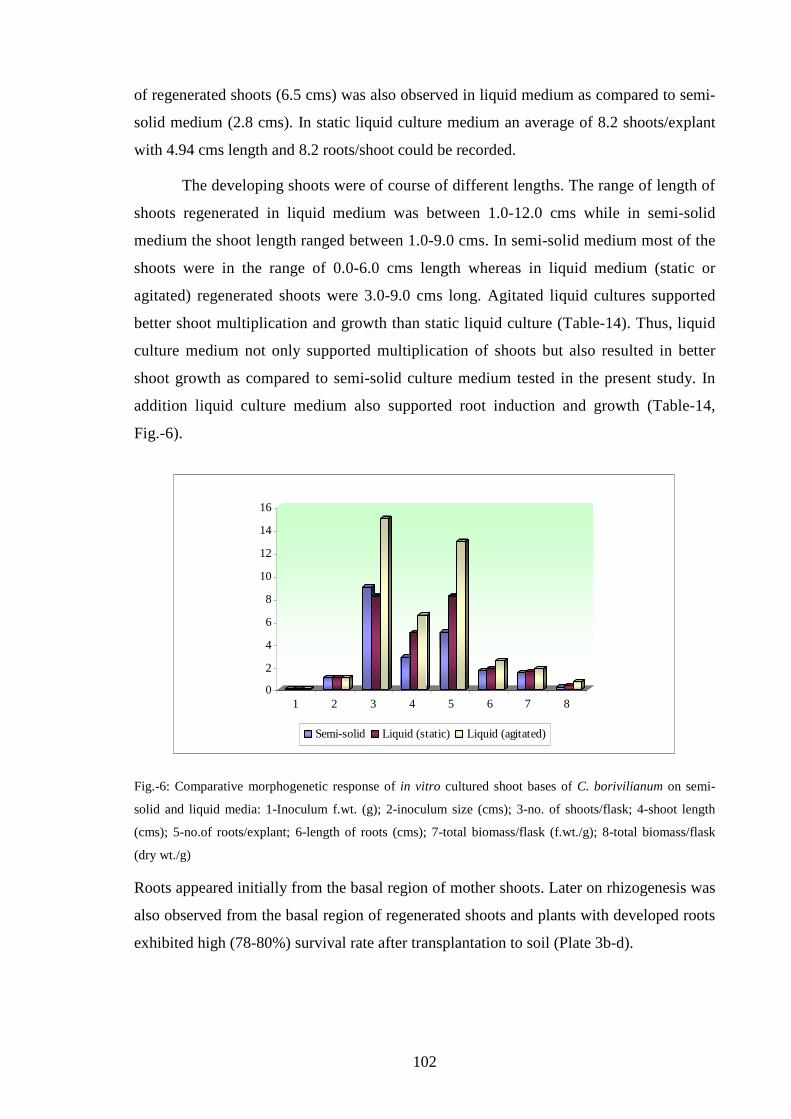
102
of regenerated shoots (6.5 cms) was also observed in liquid medium as compared to semi-
solid medium (2.8 cms). In static liquid culture medium an average of 8.2 shoots/explant
with 4.94 cms length and 8.2 roots/shoot could be recorded.
The developing shoots were of course of different lengths. The range of length of
shoots regenerated in liquid medium was between 1.0-12.0 cms while in semi-solid
medium the shoot length ranged between 1.0-9.0 cms. In semi-solid medium most of the
shoots were in the range of 0.0-6.0 cms length whereas in liquid medium (static or
agitated) regenerated shoots were 3.0-9.0 cms long. Agitated liquid cultures supported
better shoot multiplication and growth than static liquid culture (Table-14). Thus, liquid
culture medium not only supported multiplication of shoots but also resulted in better
shoot growth as compared to semi-solid culture medium tested in the present study. In
addition liquid culture medium also supported root induction and growth (Table-14,
Fig.-6).
0
2
4
6
8
10
12
14
16
1 2 3 4 5 6 7 8
Semi-solid Liquid (static) Liquid (agitated)
Fig.-6: Comparative morphogenetic response of in vitro cultured shoot bases of C. borivilianum on semi-
solid and liquid media: 1-Inoculum f.wt. (g); 2-inoculum size (cms); 3-no. of shoots/flask; 4-shoot length
(cms); 5-no.of roots/explant; 6-length of roots (cms); 7-total biomass/flask (f.wt./g); 8-total biomass/flask
(dry wt./g)
Roots appeared initially from the basal region of mother shoots. Later on rhizogenesis was
also observed from the basal region of regenerated shoots and plants with developed roots
exhibited high (78-80%) survival rate after transplantation to soil (Plate 3b-d).

103
Table-14: Effect of different physical states (semi-solid or liquid) of culture medium
on morphogenetic response of in vitro cultured shoot bases of C. borivilianum
S. No.
Growth parameter(s) Physical state of the culture medium
Semi-solid Static liquid Agitated liquid
1. 2. 3. 4. 5. 6. 7. 8. 9.
Inoculum , f.wt. (g) Inoculum size (cms) No. of shoots/flask No. of shoots having different lengths (cms) 0.0-3.0 3.1-6.0 6.1-9.0 9.1-12.0 Length of shoots (cms) No. of roots/flask Length of roots (cms) Total biomass/ flask (f.wt./g)** Total biomass/flask (dry wt./g)
0.09 1.00-1.50 9.00 ± 1.00* 5.00 3.00 1.00 0.00 2.80 ± 0.42 5.00 ± 1.00 1.60 ± 0.30 1.47 ± 0.05 0.20 ± 0.02
0.09 1.00-1.50 8.20 ± 1.30 1.40 4.20 2.60 0.00 4.94 ± 0.17 8.20 ± 1.78 1.80 ± 0.18 1.54 ± 0.06 0.30 ± 0.04
0.09 1.00-1.50 15.00 ± 1.22 1.60 4.00 7.00 2.40 6.50 ± 0.79 13.00 ± 1.58 2.50 ± 0.30 1.79 ± 0.04 0.69 ± 0.06
* Average value ± SE (standard error); ** Biomass was harvested after 40 days of culture
Because of overall supportive effect of agitated liquid medium on most of the
growth parameters tested in the present study, about 7.5-fold increase in propagable units
(shoots of which shoot base part is utilized) could be obtained after 40 days of culture
which makes liquid culture method useful for large scale multiplication of this
commercially important plant species.
2.4.3 Effect of different sucrose levels on morphogenetic response of in vitro
cultured shoot base explants
Plant organogenesis is regarded as a complex process triggered by hormones and sustained
by carbohydrates. Exogenous carbohydrates play an important role in plant tissue culture.
They have an essential function in plant metabolism since they serve as carbon and energy
source and also act as osmotic agents. The most commonly used carbohydrate for plant
tissue culture (especially for shoot regeneration and multiplication) is sucrose. Sucrose as
a carbon and energy source has significant effect on the overall growth of plants, therefore
it was intended to test the efficacy of different sucrose levels on shoot multiplication and
growth from in vitro cultured shoot bases of C. borivilianum. Carbohydrate requirements
have to be optimized for each micropropagation system (for a specific species/genotype).
Different sucrose levels ranging from 1-9% (w/v) were supplemented to MS liquid culture
medium fortified with 5.0 mgl-1 BAP (Table-14). Liquid MS basal medium without
sucrose served as control in this experiment. Amongst all the sucrose concentrations
tested, 4% sucrose level promoted highest (15.2) average number of shoots/explant and
average shoot length (5.5 cms) followed by 3% sucrose level resulting in an average
number of 14 shoots/explant and average length of 5.0 cms. All other sucrose

104
concentrations tested could not evoke growth response comparable to 3% and 4% sucrose
levels. At lower (<3%) and higher (>6%) concentrations, reduction in shoot multiplication
and growth was observed. In the absence of an exogenous carbon supply (control) shoot
multiplication and growth were significantly lower than other concentrations of sucrose
studied in the present study (Table-15, Fig.-7). The shoots turned brown and died after 2-3
weeks in culture. Amongst different sucrose concentrations tested, 3% sucrose level was
found optimal for shoot multiplication and development since no significant differences
were found when explants were cultured at a higher (4%) concentration of sucrose.
Therefore from cost reduction point of view 3% sucrose level was used for further
experiments in the present study.
Fig.-7: Effect of different sucrose levels (%) on morphogenetic growth response of cultured shoot base
explants of C. borivilianum
Table-15: Effect of different sucrose concentrations on growth response of cultured
shoot bases of C. borivilianum
Sucrose levels (%) Growth parameters(s)
No. of shoots/explant
Shoot length (cms)
No. of roots/explant
Length of roots (cms)
Total biomass/flask
( f. wt./g)
Total biomass/flask
(dry wt./g) Control (without sucrose) 1.20 ± 0.45* 0.58 ± 0.13 - - 0.25 ± 0.03 0.06 ± 0.02 1 6.00 ± 1.00 3.38 ± 0.31 5.00 ± 1.14 1.26 ± 0.26 1.09 ± 0.03 0.20 ± 0.02 2 9.00 ± 1.00 3.70 ± 0.24 10.00 ± 1.30 1.50 ± 0.15 1.34 ± 0.02 0.35 ± 0.02 3 14.00 ± 1.22 5.00 ± 0.44 15.20 ± 1.48 2.73 ± 0.15 1.74 ± 0.05 0.60 ± 0.04 4 15.20 ± 1.30 5.50 ± 0.38 14.00 ± 1.22 2.30 ± 0.40 1.76 ± 0.06 0.64 ± 0.05 5 12.00 ± 1.22 2.10 ± 0.22 11.00 ± 1.22 1.75 ± 0.10 1.66 ± 0.14 0.40 ± 0.02 6 9.00 ± 1.00 2.35 ± 0.26 10.00 ± 1.58 1.75 ± 0.11 1.37 ± 0.06 0.32 ± 0.05 7 8.75 ± 1.30 2.10 ± 0.34 7.60 ± 1.14 1.44 ± 0.12 1.15 ± 0.10 0.20 ± 0.02 8 8.00 ± 1.22 1.97 ± 0.14 6.00 ± 1.14 1.50 ± 0.19 0.95 ± 0.03 0.17 ± 0.01 9 3.00 ± 0.00 1.34 ± 0.11 2.00 ± 0.70 0.50 ± 0.00 0.35 ± 0.01 0.09 ± 0.01
* Average value ± SE (standard error); - No response
0
2
4
6
8
10
12
14
16
0 1 2 3 4 5 6 7 8 9
Sucrose conc. (%)
No. of shoots / explantLength of shoots (cms)Total biomass / flask (f. wt. / g)

105
2.4.4 Effect of different pH levels in culture medium on shoot growth and
development
The basal MS liquid medium supplemented with 5.0 mgl-1 BAP was employed in the
present experiment. As this plant species thrives best in various types of soils having
different pH ion concentrations under natural conditions, it was of considerable interest to
test the effect of pH variation on growth and development of cultured shoot bases of C.
borivilianum. The pH of the culture medium was adjusted at two levels below (3.86 and
4.86) and two levels above (6.86 and 7.86) the normal pH (5.86) which served as control
in the present experiment. However, none of the pH levels tested gave better response in
terms of growth and development of shoots and roots as compared to control under in vitro
culture conditions (Table-16, Fig.-8 ).
Table-16: Effect of pH variation in liquid culture medium on growth response of
cultured shoot bases of C. borivilianum
S. No. Growth parameter(s) pH of medium 3.86 4.86 5.86 (control) 6.86 7.86
1. No. of shoot/explant 7.66 ± 1.14* 7.20 ± 1.30 14.00 ± 1.22 7.60 ± 1.14 5.00 ± 1.00 2. Shoot length (cms) 4.23 ± 0.26 4.66 ± 0.16 5.00 ± 0.44 5.30 ± 0.59 4.00 ± 0.37
3. No. of roots/explant 15.00 ± 1.58 13.80 ± 1.30 15.20 ± 1.48 14.75 ± 1.81 9.60 ± 1.67
4. Length of roots (cms) 2.29 ± 0.30 2.39 ± 0.39 2.73 ± 0.15 2.46 ± 0.18 2.26 ± 0.43
5. Total biomass/flask (f. wt./g)
1.64 ± 0.05 0.99 ± 0.07 1.74 ± 0.05 1.16 ± 0.01 0.92 ± 0.05
6. Total biomass/flask (dry wt./g)
0.22 ± 0.01 0.16 ± 0.03 0.60 ± 0.04 0.15 ± 0.02 0.12 ± 0.02
*Average value ± SE (standard error)
Fig-8: Growth response of in vitro cultured shoot base explants of C. borivilianum at different pH levels
0
2
4
6
8
10
12
14
3.86 4.86 5.86 6.86 7.86
Culture medium pH
No. of shoots / explantLength of shoots (cms)Total biomass / flask (f. wt. / g)

106
An average number of 14 shoots/explant were obtained at pH level 5.86 (control),
while number of shoots/explant at all other pH levels was comparably lower than control.
Root number/explant and length was also highest on control medium compared to any
other pH levels tested in the present study. Highest biomass (1.74 g, f.wt.) was recorded
on control medium. This may be due to the production of highest number of shoots and
higher shoot length at this pH level.
2.4.5 Effect of inoculum density and volume of medium on shoot multiplication and
growth of in vitro cultured shoot bases
Inoculum density levels generally affect the total number of shoots produced which in
turn affect the biomass (f.wt./d.wt.) production. During the experiments where liquid
medium was used it was observed, especially at high inoculum densities that some
explants submerged in liquid medium while others remain floating and/or partially
submerged. This situation resulted in differential morphogenetic response when shoot
bases were cultured under controlled conditions. Submerged explants were less
responsive and showed hyperhydricity (vitrification) as compared to partially submerged
explants which contribute towards multiplication of shoots thus affecting biomass (f.wt.)
production. Dissolved oxygen (DO2) in the medium may play a critical role in such
situations. On the other hand limited nutritional supply due to high inoculum densities
may also result in low response. With a objective to produce maximum number of
propagable units in a defined volume of culture medium which may ultimately result in
higher efficiency of shoot multiplication and growth and low production cost, these
factors need special attention. Keeping this in mind an experiment was conducted to
study the effect of different inoculum densities (number of explants inoculated/culture
flask) versus different volume of medium per culture flask on the shoot multiplication
and growth. Shoot base explants of uniform size (1.0-1.5 cms) weighing about 0.09 g
were inoculated in 250 ml wide mouth Erlenmeyer flasks containing different volumes
(10, 20, 30, 40 or 50 ml) of shoot multiplication liquid culture medium (MS basal
medium+5.0 mgl-1 BAP) and increasing the inoculum density up to 5-fold (10 shoot base
explants per flask) in the culture flask. Two shoot bases inoculated in 30 ml culture
medium served as control. Observations revealed that two shoot base explants/culture
flask inoculum density resulted in about 7.0-7.5 fold increase in number of shoots/flask
in 30-50 ml medium after 40 days of culture period while 4 and 6 shoot base inoculum
densities exhibited only 2-5 fold increased response. Higher inoculum densities resulted
in noticeable decrease in shoot multiplication response. It was observed that when

107
inoculum density was increased beyond 6 shoot bases/culture flask, percentage increase
in terms of number of shoots produced/culture flask and biomass/flask (f.wt./g) was
lowered. The decrease in these parameters was mainly due to regeneration of lesser
number of shoots (Table-17: A-E). It was observed that as the volume of culture
medium/flask increased percentage response also increased. The best response was
obtained with 40 ml of media volume/culture flask at 2 shoot base explants/culture flask
inoculum density level followed by 30 ml of culture medium/flask at same inoculum
density level. The shoot multiplication response at 30 or 40 ml of media volume were
almost similar. An average number of 15.4 shoots/flask were obtained at 40 ml of
medium volume against 15.0 shoots/flask at 30 ml medium volume. 30 ml of medium
volume gave better response and an average shoot length of 6.5 cms was obtained
against 3.22 cms in 40 ml of medium volume. (Table-17: A-E, Fig.-9). In view of better
Fig.-9: Effect of volumes of medium on number of shoots/culture and biomass yield of C. borivilianum
plants after 40 days
shoot growth response and utilization of lower volume of culture medium which will
ultimately result in cost reduction, 30 ml of medium volume was preferred over 40 ml
medium volume. Low volume of the medium (10 and 20 ml) were least responsive with
all levels of inoculum densities tested in the present study. 30 to 50 ml medium/culture
flask supported rhizogenesis in cultured shoot bases.
0
2
4
68
10
12
14
16
10 20 30 40 50
Volume of medium / culture flask (ml)
No. of shoots / explant Length of shoots (cms)Total biomass / flask (f. wt. / g)

108
Table 17-A: Effect of inoculum density (2 shoot bases) and different volumes of
medium on shoot multiplication and growth in C. borivilianum
S. No.
Growth parameter(s) Volume of the medium in the culture flask (ml)
10 20 30 40 50
1. No. of shoots/flask 2.00 ± 0.00* 7.60 ± 0.89 15.00 ± 1.22 15.40 ± 1.67 14.40 ± 1.14 2. Shoot length (cms) 0.94 ± 0.08 3.59 ± 0.30 6.50 ± 0.79 3.22 ± 0.61 4.54 ± 0.60 3. No. of roots/flask - 4.00 ± 0.70 13.00 ± 1.58 14.60 ± 2.07 7.33 ± 1.14 4. Length of roots (cms) - 0.96 ± 0.16 2.50 ± 0.30 2.32 ± 0.33 1.24 ± 0.22
5. Total biomass / flask (f. wt./g)
0.01 ± 0.00 0.72 ± 0.03 1.79 ± 0.04 1.76 ± 0.10 2.22 ± 0.40
6. Total biomass / flask (dry wt./g)
0.004 ± 0.00 0.27 ± 0.01 0.69 ± 0.06 0.68 ± 0.07 0.89 ± 0.19
* Average value ± SE (standard error); - No response
Table 17-B: Effect of inoculum density (4 shoot bases) and different volumes of
medium on shoot multiplication and growth in C. borivilianum
S. No.
Growth parameter (s) Volume of the medium in the culture flask (ml) 10 20 30 40 50
1. No. of shoots/flask 2.40 ± 0.54* 8.00 ± 1.00 19.80 ± 2.16 14.00 ± 2.00 20.60 ± 2.30 2. Shoot length (cms) 0.82 ± 0.13 5.01 ± 0.52 3.69 ± 0.28 4.36 ± 0.64 3.70 ± 0.73
3. No. of roots/flask - 11.60 ±1.67 13.00 ± 1.87 22.00 ± 2.12 21.00 ± 2.54 4. Length of roots (cms) - 1.17 ± 0.20 1.45 ± 0.20 2.49 ± 0.25 1.64 ± 0.29 5. Total biomass/flask
(f. wt./g) 0.02 ± 0.03 1.21 ± 0.06 3.09 ± 0.14 2.83 ± 0.30 4.07 ± 0.24
6. Total biomass/flask (dry wt./ g)
0.01 ± 0.001 0.44 ± 0.05 1.31 ± 0.05 1.42 ± 0.22 1.78 ± 0.13
* Average value ± SE (standard error); - No response
Therefore 30 ml of culture medium/flask was regarded as optimal volume for
multiplication of shoots, resulting in 7.5-fold increase in shoot numbers/flask. Thus, only
30 ml liquid medium per 250 ml Erlenmeyer flask is required against 55 ml semi-solid
medium/250 ml Erlenmeyer flask, where only 9 shoots/flask could be obtained against 15
shoots/flask on liquid culture medium. In this way, 33 or 18 culture flasks can be prepared
using one litre of liquid or semi-solid culture medium respectively, which will ultimately
result in more number of shoots, thereby resulting in lower cost of plant production in
liquid culture medium.

109
Table 17-C: Effect of inoculum density (6 shoot bases) and different volumes of
medium on shoot multiplication and growth of C. borivilianum
S. No.
Growth parameter(s)
Volume of the medium in the culture flask (ml) 10 20 30 40 50
1. No. of shoot /flask 3.80 ± 0.83* 10.60 ± 1.51 14.40 ± 1.67 16.80 ± 1.92 23.00 ± 2.44 2. Shoot length (cms) 1.27 ± 0.25 3.40 ± 0.63 4.73 ± 0.77 5.78 ± 0.69 5.02 ± 0.64 3. No. of roots/flask 1.00 ± 0.00 10.00 ± 1.22 10.00 ± 1.87 9.60 ± 1.64 19.33 ± 2.58 4. Length of roots (cms) 0.50 ± 0.00 1.14 ± 0.13 1.38 ± 0.09 1.32 ± 0.25 1.02 ± 0.21 5. Total biomass/flask
(f. wt./ g) 0.30 ± 0.01 2.83 ± 0.34 1.89 ± 0.12 2.31 ± 0.22 2.87 ± 0.15
6. Total biomass/flask (dry wt./ g)
0.03 ± 0.01 1.35 ± 0.14 0.95 ± 0.09 1.20 ± 0.05 1.37 ± 0.10
* Average value ± SE (standard error); - No response
Table 17-D: Effect of inoculum density (8 shoot bases) and different volumes of
medium on shoot multiplication and growth of C. borivilianum
S. No.
Growth parameter(s) Volume of the medium in the culture flask (ml)
10 20 30 40 50
1. Total no. of shoots/flask 4.00 ± 0.70* 10.00 ± 1.58 14.40 ± 1.67 17.00 ± 1.58 21.00± 2.12 2. Shoot length (cms) 1.20 ± 0.15 3.50 ± 0.38 5.00 ± 0.52 5.20 ± 0.80 5.40 ± 0.56 3. No. of roots/explant - 9.00 ± 1.58 12.00 ± 1.87 15.00 ± 1.87 18.00± 2.34 4. Length of roots (cms) - 1.00 ± 0.18 1.90 ± 0.14 2.30 ± 0.20 2.10 ± 0.31 5. Total biomass/flask
(f. wt./ g) 0.38 ± 0.04 2.30 ± 0.18 2.44 ± 0.09 2.63 ± 0.13 2.85 ± 0.20
6. Total biomass/flask (dry wt./g)
0.06 ± 0.007 1.09 ± 0.06 1.18 ± 0.06 1.16 ± 0.07 1.33 ± 0.10
* Average value ± SE (standard error); - No response
Table 17-E: Effect of inoculum density (10 shoot bases) and different volumes of
medium on shoot multiplication and growth of C. borivilianum
S. No.
Growth parameter(s)
Volume of the medium in the culture flask (ml) 10 20 30 40 50
1. No. of shoots/flask 3.00 ± 0.70* 11.00 ± 1.58 14.00 ± 1.14 16.00 ± 1.67 18.00 ± 1.87 2. Shoot length (cms) 0.85 ± 0.07 3.50 ± 0.48 4.60 ± 0.40 5.00 ± 0.53 5.10 ± 0.54 3. No. of roots/flask - 9.00 ± 1.58 11.00 ± 1.58 17.00 ± 2.54 14.00 ± 2.82 4. Length of roots (cms) - 1.00 ± 0.21 2.00 ± 0.40 2.51 ± 0.09 2.75 ± 0.39 5. Total biomass/flask
( f. wt./g) 0.42 ± 0.04 1.92 ± 0.17 2.68 ± 0.24 2.82 ± 0.21 2.89 ± 0.20
6. Total biomass/flask (dry wt./g )
0.09 ± 0.02 0.88 ± 0.05 1.14 ± 0.05 1.26 ± 0.03 1.36 ± 0.10
* Average value ± SE (standard error); - No response
Thus, based on the cost of 30 ml of liquid MS culture medium/flask employed in
the experiments using Hi-Media make chemical reagents, the production cost of single
shoot on liquid medium is Rs. 0.02 compared to Rs. 0.26 on semi-solid medium, thereby
reducing the production cost of single shoot in liquid medium by 92.31% of the production
cost in semi-solid medium (Table-18, Fig.-10).

110
Table-18: Comparative cost of shoot production of C. borivilianum in semi-solid and
liquid medium
Steps in cost reduction
Cost analysis Physical state of medium Percent (%) cost reduction
Semi-solid Liquid
(agitated)
Step1 Cost of one liter medium* (Rs.) 41.88 7.56 81.95
Volume of medium used/flask (ml) 55.00 30.00 -
Step2 Cost of medium/flask (Rs.) 2.30 0.23 90.00
Step3 Approximate cost of single shoot (Rs.) 0.26 0.02 92.31
*Cost of chemical ingredients (Hi-Media make as per price list 2004-2005) used as per Murashige and
Skoog (1962) for preparation of one liter MS basal culture medium
Fig.-10: Percent cost reduction for in vitro shoot production of C. borivilianum in liquid and semi-solid
medium
2.4.6 Up-scaling of shoot cultures of C. borivilianum in bioreactor
Earlier studies carried out at shake flask level had revealed comparatively better shoot
multiplication and growth in the agitated liquid medium than semi-solid medium. In
earlier experiments, MS basal medium supplemented with 5.0 mgl-1 BAP was employed.
These observations prompted to conduct an experiment for large scale cultivation of C.
borivilianum plants using a bench top stirred bioreactor of 5.0 l working capacity. About
120 shoot base explants (length of each explant ranging between 2-3 cms) obtained from
shoots regenerated in vitro in liquid medium shake flasks (Plate-4a), were inoculated in
bioreactor. The initial biomass (f. wt./g) of all explants was 11.9 g. The culture vessel

111
contained 2.5 l of liquid basal MS medium supplemented with 5.0 mgl-1 BAP. An
autoclavable nylon mesh was provided as a septum in the bioreactor culture vessel just
below the medium surface dividing the vessel into two equal halves (Plate-4b, c). The
culture vessel contained two spargers, one above the autoclavable nylon mesh and
another just below the mesh through which filtered sterile air was blown in the culture
medium at 0.5 l/min rate throughout the culture duration. The vessel also contained an
agitator working at a speed of 75 rpm. The agitation results in proper mixing and
distribution of the medium evenly in all directions of the culture vessel. The nylon mesh
support prevented sinking of the explants to the bottom of the culture vessel. Growing
shoots grew normally (negatively geotropic) and were above the surface of the medium
in upper section of culture vessel. The support provided by nylon mesh also avoided
vitrification of the growing shoots. Without the support of nylon mesh few explants
always remained in suspended condition in the culture medium due to air current and
agitation in the medium, many explants submerged and settled down at the bottom of the
culture vessel. Agitation in the medium through impeller forced the submerged explants
to float and remain suspended in the medium which resulted in their poor growth due to
hyperhydricity. Similar phenomenon was observed in earlier experiments with liquid
shake flask cultures containing 50 ml culture medium. The nylon septum divided the
culture vessel into two halves and impeller placed in the lower half was separated
avoiding the direct contact of explants with agitator otherwise some of these explants
may come into contact of impeller blades and got injured. Such injured explants
exhibited poor response in terms of shoot regeneration and growth resulting in low
biomass yield. After three weeks of incubation period, the growing shoots were
harvested and the growth response of inoculated explants was recorded (Plate-4d).
About 75% response in terms of shoot regeneration and growth was observed and
biomass yield of 52.36 g (f. wt.) was recorded. Thus, approximately 4.4-fold increase in
biomass yield over the initial inoculum (f. wt.) was observed in the present experiment
(Table-19). The plantlets thus regenerated were hardenend in culture room conditions for
5 weeks as described earlier in the Materials and Methods section and planted into soil
containing a mixture of sand, soil and farmyard manure (1:1:1, v/v) for further hardening
in the glass house conditions (Plate-4e).

112
Table-19: Shoot regeneration and growth response of shoot base explants of C.
borivilianum cultured in bioreactor
Volume of medium (l)
Inoculum density Bioreactor culture vessel
configuration
Culture period (days)
Shoot growth
response (%)
Final f. wt. of
harvested biomass
Percentage increase
over inoculum
Number of explants
inocualted
F. wt. (g) of the
inoculum
2.50 120 11.90 Modifiaction of basic configuration by addition of an
autoclavable nylon mesh.
21 75 52.36 4.4-fold
2.4.7 PCR-based Random Amplified Polymorphic DNA analysis of plants obtained
from different in vitro culture conditions
A set of 20 primers (MAP01 to MAP20) with the sequences described earlier were used to
study DNA polymorphism for determining the similarity index (degree of similarity)
among the 14 randomly selected in vitro raised plants of C. borivilianum along with parent
plant. These randomly selected plants were obtained from different experimental
conditions i.e. plants obtained from cultures maintained on semi-solid or liquid media.
Plants were grown and maintained in glass house conditions along with control parent
plant.
PCR amplification by using MAP 01 to MAP 20 primers were carried out for the
isolated genomic DNA of all the 14 sample plants obtained from different in vitro
conditions along with parent plant (Plate-5a-d). Among the 20 primers, 5 primers i.e.,
MAP 03, MAP 05, MAP 06, MAP 07 and MAP 11 accounting to 25% of the total primers
used in the present investigation, did not exhibit any amplification. The remaining 15
primers i.e. MAP 01, MAP 02, MAP 04, MAP 08, MAP 09, MAP 10, MAP 12, MAP 13,
MAP 14, MAP 15, MAP 16, MAP 17, MAP 18, MAP 19 and MAP 20, which accounted
for 75% of the total primers used in the present investigation exhibited amplification.
MAP 01, MAP 04, MAP 08, MAP 09, MAP 14, MAP 15, MAP 17, MAP 18, MAP 19,
MAP 20 exhibited clear fragmentation of DNA isolated from selected plants. A total of 36
RAPD bands were produced through amplification out of which 26 bands were found to
be monomorphic and 10 were polymorphic in nature. The polymorphic amplification
products were produced by primers MAP 01, MAP 04, MAP 08, MAP 15, MAP 17 and
MAP 20. Thus, about 77.30% of monomorphism was exhibited by the randomly selected
in vitro raised plants of C. borivilianum obtained from different experimental conditions in

113
the present study (Table-20). The similarity matrix based on Nei and Li’s (1979) principle
was used to generate a graphic phenogram through UPGMA using NTSYSpc version
2.02j to find out the similarity between the plants of C. borivilianum obtained from
different in vitro experimental conditions as mentioned above along with control parent
plant. The similarity coefficients of all the 15 sample plants are presented in Table-21.
Similarity coefficients among the 15 sample plants ranged from 87% to 100%. The most
diversed pairs showing 87% similarity were sample 8 and 13 and another diverse pair
were sample 8 and 15 showing 89% similarity. The rest of the plants showed much
narrower variability with similarity coefficients of 90% to100%. Thus, plants obtained
from two different experimental conditions i.e. semi-solid and liquid culture conditions
exhibited slight variability among themselves. While samples of semi-solid or liquid
medium origin did not exhibited much variability with each other.
Table-20: Monomorphic and polymorphic bands as observed with different primers
Primers Total number of
bands
Monomorphism Polymorphism
Number of bands
Percentage (%)
Number of bands
Percentage (%)
MAP01 MAP04 MAP08 MAP09 MAP14 MAP15 MAP17 MAP18 MAP19 MAP20
4 7 5 3 3 4 3 2 1 4
1 5 3 3 3 3 2 2 1 3
25.00 71.00 60.00
100.00 100.00
75.00 67.00
100.00 100.00
75.00
3 2 2 - - 1 1 - - 1
75.00 29.00 40.00
- -
25.00 33.00
- -
25.00 Total 36 26 77.30 10 22.70
In order to analyze the relatedness among all the in vitro raised plants along with
control plant (parent plant), the UPGMA based dendrogram was constructed using paired
matrix values. It is evident from dendrogram (Fig.-11) that in vitro raised plant numbered
13 (liquid medium origin) constituted a separate major cluster from the other in vitro
raised plants. In the other major sub-cluster, in vitro raised plant numbered 15 (liquid
medium origin) was more distinct than the rest of the plants. In general the average matrix
of indices calculated on the basis of analysis of 20 different decamer primers vs. in vitro
raised plants showed that all sampled plants had a narrow genetic base. However the
unique polymorphic profiles of DNA markers were able to distinguish between plants of
different origin.

114
Table-21: Similarity coefficients of 14 randomly selected C. borivilianum plants
grown under different in vitro conditions along with parent plant
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
1 1 2 0.93 1 3 0.93 1 1 4 0.93 1 1 1 5 0.95 0.98 0.98 0.98 1 6 0.97 0.96 0.96 0.96 0.93 1 7 0.93 1 1 1 0.98 0.96 1 8 0.94 0.95 0.95 0.95 0.97 0.94 0.95 1 9 0.95 0.98 0.98 0.98 1 0.97 0.98 0.96 1 10 0.91 0.95 0.95 0.95 0.93 0.94 0.95 0.93 0.96 1 11 0.91 0.95 0.95 0.95 0.96 0.94 0.95 0.93 0.96 0.94 1 12 0.95 0.98 0.98 0.98 1 0.98 0.98 0.97 1 0.96 0.96 1 13 0.94 0.9 0.9 0.9 0.91 0.93 0.9 0.87 0.96 0.94 0.94 0.96 1 14 0.91 0.95 0.95 0.95 0.96 0.94 0.95 0.93 0.96 0.93 0.93 0.96 0.93 1 15 0.92 0.94 0.94 0.94 0.95 0.93 0.94 0.89 0.95 0.92 0.92 0.95 0.89 0.95 1
Sample1: Parent plant (control); sample 2-8: plants obtained from semi-solid culture conditions; sample 9-
15: plants obtained from liquid culture condition
Coefficient0.75 0.81 0.88 0.94 1.00
1
6
2
3
4
7
5
9
12
8
14
11
10
15
13
Fig.-11: UPGMA based dendrogram showing the relationship among 14 in vitro raised plants of C.
borivilianum obtained from different culture conditions along with parent plant

115
2.5 DISCUSSION
Chlorophytum borivilianum (Family-Liliaceae) commonly known as safed musli is an
endangered medicinal herb valued for its dried fasciculated storage roots which possess
immunomodulatory and aphrodisiac properties. The roots of safed musli (C. borivilianum)
are of immense commercial importance because they are source of steroidal saponins and
glycosides which are of great pharmaceutical value. Tuberous roots of plants are widely
used for general health purpose as tonics and aphrodisiac due to the prescence of steroidal
saponins viz. neotigogenin, neohecogenin, stigmasterol and tokorogenin (Tandon and
Shukla, 1995b). They form an important ingredient in a large number of Herbal, Unani,
Homeopathic and Allopathic formulations. The main active principles of roots, saponins
are known as stimulants and metabolic enhancers and are also shown to posses anti-
tumour activity. The extract of dried root tubers of C. borivilianum acts as
psychostimulant and has a beneficial effect on the brain and human body by increasing
alertness, mental ability, intelligence and sexual characters. Demand for dried roots of the
plant due to its therapeutic activity and diversified uses is increasing in Indian and
International market. It is estimated that the overall demand for safed musli’s dried tubers
in Indian and foreign markets is more than 35,000 tonnes per year while the supply is only
a meager 500 tonnes per year. Of the available quantum of safed musli, over 95% of it
comes through wild harvesting, an act expressly prohibited by the various state
governments. The cultivation of safed musli is also limited in the country. Due to its
significant demand and supply gap there is an urgent need to conserve and multiply the
plant in bulk and its domestication to meet the present demand. Therefore, commercial
cultivation of this species is highly desired. In nature the plant is propagated by seeds and
has been enlisted as an endangered species (Nayar and Shastri, 1988) due to low seed set,
seed viability and germination (11-24%) associated with overexploitation from the natural

116
habitats and insufficient attempts either to allow its replenishment or its cultivation
(Bordia et al., 1995). This has prompted National Medicinal Plant Board to categorize it
amongst the prioritized plant species for promoting its conservation and commercial
cultivation. Due to low regeneration rate through seedlings, vegetative method of
propagation is the generally adopted mode of regeneration at present for its commercial
cultivation. It is propagated vegetatively by root tubers. Poor seed setting and germination
and slow propagation rate through vegetative propagation are major constraints in the
large-scale commercial cultivation of safed musli. Because of these inherent problems of
regeneration and non-availability of elite planting material and agrotechnological
practices, large-scale commercial cultivation of safed musli has not succeded so far and a
large amount of annual internal requirement is met through wild collections. Keeping in
view the economic importance of this species, it would be highly desirable to develop in
vitro procedures for regeneration in which plant tissue culture offers the possibility of
rapid clonal propagation and conservation of germplasm of this species. Micropropagation
offers many advantages over conventional propagation methods. True-to-type
multiplication through micropropagation provides genetically identical (George, 1993)
disease free plants within shorter time and comparatively smaller space independent of
seasonal and other environmental conditions (Debergh and Zimmerman, 1991). It
represents one way of possible recovery for some endangered and over exploited
medicinal plant species as well as a method for providing raw plant materials for
extraction of medicinally important compounds known as secondary metabolites.
The present investigation deals with the standardization of culture conditions
conducive for rapid multiplication of safed musli (C. borivilianum) via multiple shoot
regeneration. In this context, nutritional requirements for multiple shoot regeneration,
growth and rooting of in vitro regenerated shoots were optimized after establishing axenic
cultures of C. borivilianum through explants obtained from C. borivilianum plants
maintained in glass house. Subsequently shoot growth and multiplication response and

117
biomass production were compared on semi-solid and liquid culture medium for achieving
a cost-effective and efficient method of multiple shoot production of C. borivilianum.
Liquid medium was optimized for multiple shoot regeneration and growth by testing
different levels of sucrose and pH. For achieving further cost-effectiveness and efficiency
in the present protocol for micropropagation of C. borivilianum, experiments were
conducted for optimizing the inoculum density and volume of culture medium at shake
flask level in liquid culture medium and further experiments were also performed to
compare micropropagation in shake flask and a bench top stirred tank bioreactor with a
view to employ bioreactors for up-scaling large-scale propagation of this endangered yet
commercially important plant. Subsequently plants regenerated in vitro in semi-solid and
liquid culture conditions were established in soil and genetic stability and fidelity of these
plants was analyzed through Random Amplified Polymorphic DNA (RAPD) approach, a
prerequisite for large-scale commercial production of this plant.
Shoot base explants of C. borivilianum, cultured on semi-solid MS medium
supplemented with various levels of cytokinins either individually or in combinations,
exhibited different frequency of morphogenetic response and thereby the number of shoots
regenerated varied. Growth parameters such as number of shoots/explant, length of shoots
(cms) and number of roots/explant in the regenerated shoots were recorded after 30 days
of culture in the present study. Amongst all cytokinins or their combinations viz. Kn,
BAP, Kn+BAP, 2iP, Z, AS or TDZ tested only BAP or TDZ were effective and produced
a maximum of 11.4 and 18.4 shoots/explant after 30 days of culture period respectively
thereby producing highest number of shoots/explant as compared to other cytokinins or
their combinations.
Amongst all cytokinins tested, it was observed that BAP at 1.0-7.5 mgl-1 levels
exhibited best response. TDZ showed better response than BAP for shoot regeneration but
shoot growth and length were comparatively more in BAP supplemented medium. Similar
observations have been recorded earlier in medicinal herbs like Lavandula stoechas (Jose,
1996), Rauvolfia tetraphylla L. (Faisal et al., 2005) and Curcuma longa L. (Prathanturarug
et al., 2005), where shoots regenerated on the medium containing TDZ showed stunted
shoot growth and exhibited symptoms of hyperhydricity upon prolonged exposure to the
TDZ. As observed earlier (Fratini and Ruiz, 2002), TDZ inhibited root development as
compared to other hormones especially BAP. Observations revealed that Kn at 5.0 mgl-1
level supported shoot elongation better than BAP but induced regeneration of lesser
number of shoots/explant (7 shoots/explant as against 11.4 shoots/explant on medium

118
fortified with 5.0 mgl-1 BAP). 2-iP also induced multiple shoot formation as observed on
kinetin supplemented medium although shoot length was comparatively lower than on Kn
and BAP supplemented medium. In order to improve upon the shoot growth and length,
Kn and BAP were tested in equal proportions. The combinations supported shoot growth
but shoot multiplication response was low compared to the medium supplemented with
BAP alone. Besides, callusing response from base of regenerated shoots was observed on
BAP and kinetin supplemented medium. Higher levels of cytokinins tested (>7.5 mgl-1)
were less effective. Amongst all the cytokinins, AS and Z were least responsive. MS basal
medium without any hormone (control medium) in the present study also supported shoot
growth but shoot regeneration was not comparable to that observed on MS medium
fortified with 5.0 mgl-1 BAP. Thus, MS basal medium supplemented with 5.0 mgl-1 BAP
was found to be most suitable for growth and elongation of regenerated shoots without
inducing the callusing response.
Amongst all the five auxins (IAA, IBA, NAA, 2,4-D or P) tested, none of the
auxins could induce multiple shoots in cultured explants however IAA, IBA and NAA
supported shoot growth. IBA and NAA were effective in root induction and growth from
basal portion of in vitro regenerated shoots. Amongst all auxins tested, NAA at 0.25 mgl-1
level induced optimal root regeneration (an average number of 19.6 roots per explant) but
also exhibited callusing. IBA (2.0 mgl-1) supplemented to MS basal medium induced 16.6
roots per explant without any intervening callus phase. Picloram (P) or 2,4-D were least
effective amongst all auxins tested. Callusing was also observed at the basal portion of
shoots cultured on 2,4-D supplemented media.
Different strengths of MS basal medium were also tested to standardize best
nutritional requirement for root induction and growth and it was observed that three-fourth
strength of MS salts supplemented with 2.0 mgl-1 IBA gave best response and an average
number of 15 roots per shoot with an average length of 1.5 cms were obtained.
In vitro clonal micropropagation of C. borivilianum via shoot buds on stem disc
and shoot base culture using semi-solid medium has been reported earlier (Purohit et al.,
1994a; Dave et al., 2003a; Pudake and Dhumale, 2003; Sharma and Mohan 2006). In
these reports like the present investigation in vitro cultures were initiated from stem discs
possessing shoot buds and young shoot bases obtained from field-grown plants. But later
on shoot bases were used as a source of initial explants because of low contamination rate
compared to in vitro cultures initiated from stem discs. In these studies best response of

119
shoot multiplication was observed on MS medium supplemented with 5.0 mgl-1 BAP and
3% sucrose. Purohit et al. (1994a) also observed callusing on Kn or NAA containing
medium employed for shoot regeneration or root induction experiments respectively.
Pudake and Dhumale (2003) found MS medium supplemented with 2.0 mgl-1 BAP
optimal for shoot multiplication. Amongst different combinations of IBA or NAA tested
for rooting, 3.0 mgl-1 IBA was found to be optimal for rhizogenesis. Sharma and Mohan
(2006) also recorded maximum frequency and number of shoots regenerated on MS
medium containing 2.0 mgl-1 BAP. In vitro regenerated shoots showed 86.7% rooting
(fasciculated roots) on Knops medium containing iron and vitamins of MS medium with
2.0 mg l-1 IBA and 0.1% activated charcoal while in the present study, three-fourth
strength MS medium supplemented with 2.0 mgl-1 IBA was found optimal for
rhizogenesis.
Reduced strength of composition of MS medium has been reported to be better
than full strength in many other plant species also (Jha et al., 2002; Ebrahim et al., 2007)
and employed for lowering callusing tendency. IBA responded better than other auxins for
rhizogenesis in many plant species (Verma et al., 2002; Amany et al., 2007)
The present study revealed that the clonal propagation through multiple shoots
obtained from shoot base explants is a viable strategy for the multiplication of C.
borivilianum plants for commercial purposes. Through a rapid increase in the number of
shoots which can ultimately give rise to rooted plants (without callus formation thus
avoiding incidence of genetic variability), this protocol can be useful for conservation and
multiplication of this valuable plant.
A shoot base with shoot bud primordia treated to be the basic culturable unit, could
give rise to well developed plantlet in the present study. Therefore attention was focussed
to maximize the production of such propagable units which will consequently result in low
cost of plant production. Many tissue culture nurseries throughout the world use shake
flasks or bottles containing agar medium for commercial cultivation but efficiency is not
high enough because of high labour costs and time involved.
In order to improve efficiency, the use of shake flask culture techniques for mass
propagation of plants has been reported earlier (Takayama and Misawa, 1981). In the
present study attempts were made to elucidate the stepwise procedure in establishing an
efficient liquid culture system by improving upon the existing micropropagation
prortocols on safed musli which employed semi-solid culture medium (Purohit et al,

120
1994a; Dave et al., 2003a; Pudake and Dhumale, 2003; Sharma and Mohan 2006). In
these reports the tissue culture studies in safed musli (Chlorophytum borivilianum) were
carried out using agar-solidified medium only while in the present study MS basal medium
supplemented with 5.0 mgl-1 BAP with or without agar (0.8%, w/v) was used to analyze
the comparative growth performance of C. borivilianum shoot base explants on semi-solid
and liquid culture medium for achieving a cost-effective method for micropropagation of
C. borivilianum through multiple shoot production. This is first report on
micropropagation in C. borivilianum through multiple shoot proliferation employing
liquid culture medium.
Liquid cultures supported better shoot growth and multiplication of regenerated
shoots than semi-solid medium. An average of 15 shoots/flask (7.5-fold increase over 2
shoots/flask inoculum density) could be obtained in liquid medium against 9 shoots/ flask
(4.5-fold increase) on agar-solidified medium during same culture duration. Purohit et al.
(1994a) earlier exhibited that 5.0 mgl-1 BAP concentration in semi-solid culture medium
was best for shoot multiplication and reported four-fold increase in shoot multiplication
rate. Similarly Dave et al. (2003a) have reported only 3.5-fold increase in shoot
multiplication on BDH brand (British Drug House, India) agar-gelled MS medium
containing 5.0 mgl-1 BAP while other gelling agents did not supported shoot
multiplication. In the present study the average length of shoots recorded on liquid
medium was 6.5 cms against 2.8 cms on semi-solid medium. Therefore, in the present
study, a significant enhancement in shoot multiplication rate was observed employing
liquid culture medium. Liquid culture medium not only supported multiplication of shoots
but shoot growth was also better. It was observed that agitation in liquid cultures
supported shoot growth and multiplication in a better way compared to stationary liquid
cultures. Liquid culture medium also supported better root induction and growth and
rhizogenesis was observed from basal region of regenerated shoots.
Higher cost of plant production has always restricted the use and exploitation of
micropropagation technique at industrial level (Kozai et al., 1997). To overcome these
limitations certain cost-reduction strategies have been employed (Levin and Vasil, 1989).
For scale-up of cultures, bioreactors using liquid culture medium have been recommended
(Kim et al., 2004; Paek et al., 2005).
The physical state of the culture medium and its composition affect the in vitro
growth of plants to a great extent. In micropropagation, culture medium gelled with agar is

121
commonly used as a support matrix for the explant growth. It significantly adds upto 65%
of the cost of the culture medium (Mitra et al., 1998). Besides, it also results in local
accumulation of heat and hinders the availability of dissolved oxygen to the cultured cells
(Kohlenbach and Wernicke, 1978). There have been several reports related to adverse
culture effects of use of agar for gelling of medium (Debergh, 1983; Bornman and
Vogelmann, 1984; Conner and Meredith, 1984). On the other hand by employing liquid
culture medium, reduction in plant production cost and increased growth response can be
achieved (Paque et al., 1992; Bhagyalakshmi and Singh, 1995). Use of liquid culture
medium resulted in better growth and multiplication of regenerated shoots because of
proper contact of tissue in liquid medium resulting in increased nutrient uptake compared
to semi-solid medium. This resulted in reduction of the plant production cost because of
omission of agar from the culture medium, which is one of the costliest components of
culture medium. Liquid medium can be employed in bioreactors for large-scale
multiplication of plants. The response of cultured tissue to media manipulation and
selection pressure is more rapid in liquid culture medium. The advantages of liquid culture
medium for enhancing shoot proliferation and growth have been reported in several plant
species. In Rhododendron shoot production on the liquid medium was 10-fold higher than
with the agar-solidified medium (Douglas, 1984). The potential of liquid culture for the
enhanced bud regeneration from aspen roots is also well documented (Vinocur et al.,
2000). An efficient system for clonal propagation in liquid culture was well established for
oranamental gentian (Hosokawa et al., 1998). Maene and Debergh (1985) reported
elongation and root induction in Cordyline, Philodendron, Magnolia and Spathiphyllum
where liquid media was added to establish the exhausted cultures instead of subculturing
the tissue to fresh medium. Kim et al. (2003) also reported enhanced shoot multiplication
and bulblet formation of Allium sativum in liquid cultures. Chen et al. (2005) observed the
use of liquid cultures for mass proliferation of meristematic clusters of daylily
(Hemerocallis spp.) followed by regeneration of adventitious shoots on semi-solid agar
culture medium as an efficient system for large-scale micropropagation. When shoot buds
are shake cultured in liquid medium, quite different growth responses were observed
between species or genera. Some plant species grew fairly well in shake cultures while
others were inhibited. The growth characteristics of cultures depend on the nature of the
genera, species or cultivars. Begonia x hiemalis grew well in liquid medium but
Saintpaulia and strawberry grew fairly well on agar medium and the growth was
suppressed in liquid culture medium (Takayama, 1991). The optimal composition of the

122
liquid medium used for a particular plant species may be different from that of the agar
medium. Contrary to this, in the present study the same medium composition (MS medium
containing 5.0 mgl-1 BAP) supported optimal growth of cultured shoot base explants of C.
borivilianum, both in agar gelled and liquid media.
Sucrose as a carbon and energy source has significant effect on the overall growth
of plants, therefore effect of different sucrose levels on shoot multiplication and growth
from in vitro cultured shoot bases of C. borivilianum was observed in the present study.
Observations revealed that maximum number of shoots/explant were obtained on 4%
sucrose level closely followed by 3% sucrose level. Growth response (shoot regeneration
and growth) was lower on all other levels tested and higher levels resulted in severe
reduction in most of the growth parameters tested. Amongst 3% and 4% concentrations of
sucrose, 3% sucrose concentration was preferred due to the the higher cost-efficiency.
Sucrose is the most common carbon source used in plant cell, tissue and organ
culture media. Studies have shown that sucrose also influences secondary metabolism in
cell and organ culture (Fowler, 1983; Merillon et al., 1984). In many plant systems
independent growth could not be achieved on a medium without sucrose (Grout and Price,
1987). Singh and Shyamal (2001) evaluated different requirements of sucrose for in vitro
rooting of hybrid tea rose cultivar Sonia and Super Star. The difference in their
requirements might be due to genotypic effect. Alleviated sucrose levels favoured rooting
and root quality but a slight declining trend was noted with the highest (4%) level tested.
Like any other morphognetic response, rooting is an energy consuming process and
sucrose is needed as an energy source. In apple root initiation decreased proportionately
with decreasing sucrose level (Lane, 1978) and shoots cultured on media without sucrose
did not survive. High frequency in vitro shoot multiplication in Plumbago indica was
achieved in a medium containing 3% sucrose (Chetia and Handique, 2000). In our studies
also 3% sucrose level supported high rates of shoot growth and multiplication. In gerbera
also induction of multiple shoots was accomplished on MS medium supplemented with
3% sucrose and other phytohormones (Aswath and Choudhury, 2002). While Mehta et al.
(2000) reported that increase in sucrose concentration from 2% to 4% in the medium
increased caullogenic response in the tamarind plants and increase of sucrose
concentration to 6% induced browning of media which proved detrimental for the growth
of the shoots. On the other hand Misra and Dutta (2001) reported that liquid medium
having 9% sucrose and other phytohormones was suitable for growth of bulblets in
isolated unrooted shoots. Takayama and Misawa (1979) suggested that the effect of

123
sucrose on the bulb growth of Lilium auratum could be mediated through changes in
osmotic potential. Apparently high sucrose levels were more stressful for shoots exhibiting
poor development. In our studies explants inoculated on media containing high sucrose
levels (beyond 8%) also exhibited somewhat similar growth response. Sugars are reported
to have an osmotic role and also act as a source of energy and carbon in inducing shoot
regeneration in tobacco callus (Brown et al., 1979) and from leaf pieces of Solanum
melongena (Mukherjee et al., 1991).
In studies to examine the effect of different pH levels on multiple shoot
regeneration and growth, it was observed that pH level 5.86 (control) gave better response
in terms of growth and development of proapagable units (shoots of which shoot base part
is utilized) and roots than pH levels lower or higher than the control medium. Highest
biomass and root length were also recorded on control medium. However, little work has
been carried out to determine the effect of medium pH on plant growth in vitro either in
solid or liquid medium, although different plant species growing in vivo were shown to
have distinct pH optima for growth ranging from 4.2 to 7.4 (Scheffer and Schachtschabel,
1979; Mengal, 1984). In natural field conditions the pH affected availability of nutrients,
uptake of nutrients by the plant and the survival of soil borne plant pathogens in the
rhizosphere (Smiley, 1978; Scheffer and Schachtschabel, 1979; Mengal, 1984).
Differences in initial pH values have been found to affect the pattern of in vitro growth
and multiplication of explants and the induction of roots in many plant species (Leifert et
al., 1992). It was observed that ability to maintain high growth rates on a medium with
initially low pH such as 3.5 appears to depend on the ability of plants to raise the pH of the
medium. While in Hemerocallis, Hosta and Delphenium shoot multiplication rates were
equal to that obtained in control medium (pH 5.6), Choisya and Daphne had reduced
multiplication rates. In our studies pH levels higher or lower than control (5.86) medium
negatively affected the shoot development and multiplication. Development and
multiplication of sugarcane plants was not affected and they resisted micropropagation in
acidic pH values (Isabela and Gluseppina, 2003). The pH of the medium below 4.5 was
shown to affect in vitro morphogenesis (Rastogi and Sawhney, 1987; Smith and Krikorian,
1990a).
In vitro growth reductions observed may be due to low NH4+ availability with
decreasing pH (Martin and Rose, 1975). Uptake of nutrients from the medium is inhibited
since ‘active uptake’ of nutrients i.e. potassium is thought to rely on proton gradients

124
across biological membranes (Hodges, 1973) and biological membranes are known to be
affected by the low medium pH values of less than 4.0 (Mengal, 1984).
In another study, the effect of inocuum density on shoot regeneration and growth
was recorded which ultimately affected the biomass (f. wt.) production in the culture
vessel. About 7.0-7.5 fold increase in number of shoots/flask was observed in two shoot
base explants/culture flask inoculum density after 40 days of culture while higher
inoculum densities resulted in decrease in multiplication response. This is probably due to
higher number of shoots regenerated on the low inoculum densities compared to those
obtained on the higher inoculum densities.
In the shake flasks some of the explants were fully submerged while some others
were partially submerged or floating. This situation resulted in differential morphogenetic
response of inoculated shoot base explants. Submerged explants were less responsive than
partially submerged or floating explants. It is likely that dissolved oxygen (DO) plays a
critical role in such situations. Although many plant species have been cultured using
liquid cultures, vitrification and physiological disorder of tissue cultured plants, tissues
exhibiting translucency and hyperhydric transformations have been reported for many
crops cultured directly in liquid media (Debergh et al., 1981; Pierik, 1991). Therefore,
many techniques i.e. providing support to plants by using rock wool, poly-urethane and
cellulose plugs etc. in stationary liquid media (Debergh and Maene, 1981; Conner and
Meredith, 1984) have been explored. To avoid virtification, a two stage culture method
(15-20 days on solid medium followed by 15-20 days on liquid medium) has also been
suggested (Barve et al., 1986). The common use of the two-phase (a liquid phase over a
solid phase) medium has led to a high multiplication ratio combined with low sensitivity
to vitrification (Pierik, 1987).
In order to record the minimal amount of medium required for the growth and
multiplication of explants to overcome the problem of their submergence and
hyperhydricity, different volumes of liquid media (10-50 ml/flask) were tested under
agitated conditions for 40 days of the culture in the present study. The shoot multiplication
response at 30 or 40 ml of medium volume was almost similar. An average of 15.4
shoots/flask were obtained at 40 ml of medium volume against 15 shoots/flask containing
30 ml medium. 30 ml volume of medium was preferred over 40 ml because of better shoot
growth response and utilization of lower volume of culture medium resulting towards
cost-efficiency. However, higher (50 ml) and lower (10 ml) volume of medium adversely

125
affected the shoot multiplication and growth, in higher (50 ml) volume possibly as a result
of lower oxygen availability and submergence of the explants while lower volume of the
medium also resulted in lower production of shoots/culture due to the rapid depletion of
the medium. Shoot multiplication response to different volumes of liquid media under
static and agitated conditions in tea (Camellia sinensis) has also been reported earlier
(Sandal et al., 2001). Chu et al. (1993) reported that shoots grown in 30 ml or even higher
volume of liquid medium had a higher multiplication ratio than those grown in lower (10
ml or 20 ml) as also observed in the present study. They also showed that miniature rose
can be grown and multiplied successfully without any support in liquid substrate as was
also observed in the present study and higher multiplication rate in liquid medium as
compared to semi-solid medium may be due to greater availability of hormones and other
nutrients in the liquid medium. Longer culture period in lower volume of liquid medium
yielded the best multiplication results in rose (Chu et al., 1993). On the other hand Sandal
et al. (2001) observed that even more lower volume (20 ml) was most effective for tea
shoot proliferation and that at higher volume (50 ml) hyperhdricity symptoms were
induced as a result of lower availability of oxygen and submergence of the explant.
Contrary to the adverse effect of the high volume of the liquid medium on the shoot
growth as observed in the present study in C. borivilianum, a linear increase in the growth
of Begonia plantlets was observed (Takayama and Misawa, 1981). Begonia plantlets had
luxuriant growth in flasks with 100 ml or 150 ml of the medium compared to the growth in
30 ml of culture medium dispensed in 300 ml Erlenmeyr flasks with agitation speed of 180
rpm. Nadgauda and Mascarenhas (1996) reported good shoot proliferation in bamboo in
static liquid cultures. In present studies agitated liquid cultures were more suitable than the
static liquid cultures for shoot regeneration and growth.
In the present study by employing liquid medium, while on one hand lesser amount
of medium is consumed, on the other hand shoot growth and multiplication response were
better than that observed on semi-solid medium. Simultaneously in liquid medium by
excluding agar, shoot production cost is also reduced. Role of liquid versus agar-gelled
medium in mass propagation and ex vitro survival of banana has been well documented
(Bhagyalakshmi and Singh, 1995). Recently the physical form of the culture medium has
received much attention and use of liquid medium for in vitro micropropagation is
advocated as one of the strategies for reducing the cost of in vitro regenerated plantlets.
Further, handling of liquid medium is comparatively easier than solid medium, which

126
saves labour and energy. Thus, due to higher response of shoot growth and multiplication,
liquid culture medium can be useful for large-scale multiplication of C. borivilianum.
Thus, results in the present study at shake flask level clearly revealed that the MS
basal medium (30 ml) fortified with 5.0 mgl-1 BAP at pH 5.86 dispensed in 250 ml
Erlenmeyer flask supported the maximum production of propagable units and plantlets
with developed roots showed high (78-80%) survival rate when transplanted to soil. This
efficient micropropagation method can be a suitable alternative to the previously
developed methods for micropropagation of C. borivilianum and can also be applied for
mass propagation of plants using the liquid culture medium in bioreactors which may
further reduce plant production cost.
Plant propagation by tissue culture techniques is encumbered by intensive labour
requirement for the multiplication process, thus scaling-up systems and automation of the
unit operations are necessary to cut down the productin costs (Aitken-Christie, 1991;
Vasil, 1991a). The major disadvantage of the conventional micropropagation techniques is
the difficulty in controlling the chemical/physical parameters in the culture vessel.
Therefore it is important to develop the new propagation strategies to overcome these
limitations of conventional micropropagation techniques. Adopting bioreactors/fermentors
with liquid media for micropropagation is favourable due to the ease for scaling-up (Preil,
1991) resulting in low production costs. Adoption of bioreactors may be beneficial for the
development of new regeneration systems that are free from physiological disorders and
year round production of the useful plants can be achieved (Paek et al., 2001). Large-scale
culture of plants regenerating via organogenesis is rather limited and has been reported for
ferns (Ziv and Hadar, 1991), Philodendron (Ziv and Ariel, 1991), Begonia, Lilium
(Takayama and Misawa, 1981, 1982; Takayama et al., 1991) and Gladiolus (Ziv, 1989,
1990). Only a few bioreactor configurations are presently used in practice for plant
propagation (Takayama and Akita, 1994; Akita and Ohta, 1998). This is mainly due to the
lack of systematic and factorial experiments on propagation in bioreactors which are
needed to reveal the complex interactions between the plant physiolgy and physical
parameters of bioreactors.
During the present course of study, mechanically agitated bench top stirred tank
reactor, one of the most popular industrial-scale bioreactor system was employed for
scaling-up culture of C. borivilianum. The culture vessel had a basic configuration and
was provided with two air spargers, one above and another below the autoclavable nylon

127
mesh and an agitator working at 75 rpm. It had advantages in terms of effectiveness in
mixing and breaking up the bubbles and enhanced oxygenation. On the other hand, it had
some disadvantages such as complicated configuration, greater exposure to contaminats,
difficulty in optimizing variable conditions and high-energy consumption. In the present
experiment provision of nylon mesh as a septum in culture vessel just below the medium
surface provided support to growing shoot base explants and avoided vitrification. This
also prevented tissue injury by avoiding direct contact with impeller. About 75% of the
cultured shoot base segments exhibited growth response and about 4.4-fold increase in the
shoot biomass over the initial inoculum weight was recorded after 21 days of culture
period. The effects of number and position of the impeller and vessel configuration may
affect shoot proliferation and further needs to be determined to ensure optimum conditions
(Lee, 1997).
Another important aspect of micropropagation protocols for commercial purposes
is the genetic integrity of plants. In vitro raised plants are usually susceptible to genetic
changes due to culture stress (Dunstan and Thorpe, 1986; Cecchini et al., 1992; Rani and
Raina, 1998). Long term multiplication may lead to somaclonal or epigenetic variation in
the micropropagated plants questioning the fidelity of their clonal nature.
The exact cause of somaclonal variation in in vitro cultures are still unknown,
although it is believed that alterations in auxin-cytokinin concentrations and their ratio,
duration of in vitro culture, in vitro stress due to unnatural conditions (Modgil et al., 2005)
together or independently are responsible in some cases. Cultured plant tissues are also
known to undergo high levels of oxidative stress due to reactive oxygen species formed
within the cells and the latter is known to cause DNA damage, including that of
microsatellite instability (Jackson et al., 1998). Therefore it is necessary to establish
genetic uniformity of micropropagated plants. Several techniques such as cytological,
isozymes and molecular markers have been employed to detect variation if any or to
confirm the genetic stability of micropropagated plants (Gupta and Varshney, 1999).
Utility of RAPD as a means of molecular analysis of in vitro regenerated plants has
been very well documented by many workers (Williams et al., 1990; Brown et al., 1993;
Rani et al., 1995; Rani and Raina, 1998; Gupta and Varshney, 1999). The detection of
subtle somclonal variation at the phenotypic, cytological, biochemical and molecular
levels among the microropagated plants in many taxa (Armstrong and Phillips, 1988;
Amberger et al., 1992; Rival et al., 1998; Rani and Raina, 2000, 2002) have however

128
pointed out the validity of the concept that complete genetic stability is retained in cultures
derived through organized meristem cultures but the occurrence of cryptic genetic effects
arising via somaclonal variation in the regenerants seriously limits the broader utility of
micropropagation system. Genetic fidelity of micropropagated plants has immense
practical utility and commercial implications. Therefore, it is important to first establish
the suitability of the micropropagation protocol developed with respect to the production
of genetically identical and stable plants. In the present study an efficient regeneration
system via multiple shoot bud induction from shoot base explants of C. borivilianum is
reported and subsequently genetic fidelity of plants regenerated under different culture
conditions i.e. semi-solid and liquid was assessed by RAPD technique. In this study PCR
amplifications by using a set of 20 different primers was carried for 14 randomly selected
in vitro raised plants obtained from different culture conditions and their RAPD band
pattern was compared with that of explant donar mother plant. In vitro raised plants of C.
borivilianum obtained from different culture conditions in the present study revealed about
77.30% monomorphism. Out of total 36 RAPD bands produced through amplification, 26
bands were monomorphic, while only 10 bands were polymorphic. Similarity coefficients
amongst the 15 accessions ranged between 87% to 100%. It was evident from the RAPD
analysis that in the present study plants obtained from two different experimental
conditions i.e. semi-solid and liquid culture conditions exhibited slight variability
compared to each other. On the other hand samples of semi-solid or liquid medium origin
did not exhibited much variability when compared with other samples obtained from same
culture conditions. Most of the plants showed much narrower variability with similarity
coefficients of 90% to 100%. In general the average matrix of indices calculated on the
basis of analysis of 20 different decamer primers vs. in vitro raised plants showed that all
sampled plants had a narrow genetic base. However the unique polymorphic profiles of
DNA markers were able to distinguish between plants of different origin.
In the present investigation, the monomorphic banding pattern in micropropagated
plants and the in vivo maintained mother plant of C. borivilianum obtained after screening
primers that are common for higher plants shows that organized multiple shoot bud
differentiation from shoot base explants without intervening callus phase is least
vulnerable to genetic changes. This establishes the suitability of protocol developed for the
micropropagation of C. borivilianum. These results were further supported by earlier
reports where RAPD markers have indicated the maintenance of genetic integrity amongst
meristem culture derived plants in Pinus thunberguii (Goto et al., 1998), Phoenix

129
dactylifera (Javouhey et al., 2000), Eucalyptus tereticornis and E. camaldulensis (Rani
and Raina, 2000, 2002), chestnut root stock hybrids (Carvalho et al., 2004) and almond
(Martins et al., 2004).
In contrast RAPD markers have also been shown to detect somaclonal variations
among the micropropagaed plants (Isabel et al., 1996; Rival et al., 1998; Rani et al., 2000;
Palombi and Damiano, 2002). A species/genotype specific response to genetic
stability/instability in tissue culture conditions is not uncommon and has been reported in
many plant taxa (Rani and Raina, 2000, 2002). The results of the present study in
conjunction with the earlier reports (Lattoo et al., 2006) has convincingly emphasized that
plants derived through multiple shoot proliferation from shoot base culture explants using
liquid medium as a culture substrate exhibit high genetic stability during in vitro
propagation and therefore protocol can be adapted at commercial level.
High multiplication frequency, molecular and phenotypic stability ensures the
efficiency of the protocol developed for production and conservation of this commercially
important endangered medicinal herb. Efficient regeneration process optimized here also
has implications for genetic transformation of C. borivilianum.





























