Embed Size (px)
Citation preview

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 191
Part III
Organization of Cell Populations
Chapter 10Development and Differentiation
Since ancient times, people have wondered how organisms are formed during
the developmental process, and many researchers have worked tirelessly in
search of the answer. Although large amounts of data have been accumulated
along the way, a full understanding of the developmental phenomena has not yet
been achieved. However, with the development of techniques to analyze these
phenomena at the gene level, our knowledge in the area has rapidly increased
in recent years. Now, we are becoming increasingly aware that the seemingly
diverse biological developmental phenomena involved are in fact related to a
number of common basic mechanisms.
By referring to specific case examples, this chapter discusses the basic mechanisms
of biological development that have been revealed at the molecular level.
I . Oogenesis
In many animals other than mammals, the materials necessary for continued
development to a certain stage need to be pre-stored in the egg. These materials,
which are derived from the mother and therefore known as maternal factors, include
many substances that play important roles in the early stages of development. The main
constituents of maternal factors are mRNA and special proteins. These factors are
involved in many of the important events that occur during the early stages of
development, such as determining which way the embryo faces (e.g., head-tail
direction and dorsal-ventral-side direction), determining the fate of embryonic cells
(i.e., deciding which tissues or organs they will become), regulating cell growth, etc.
Many maternal factors are stored in the oocyte during oogenesis, and many are
stored unevenly in the cytoplasm of the oocyte (Fig. 10-1). The uneven distribution
of these factors is closely associated with the determination of the direction of the
embryo and the fate of embryonic cells. As an example, maternal factors
localized in the egg will become unevenly distributed over embryonic cells
(blastomeres) through the division of the egg (cleavage). As a result, they become
deeply involved in the determination of these variables.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 192
I I . Ferti l ization and Cleavage
Besides the important task of combining two sets of genes from the father and
mother into one, fertilization plays a number of other significant roles. As an
example, in frog embryos, as discussed later, the direction of the body is
determined by the site of the oocyte that the fertilizing sperm enters. The entry of
the sperm also sets off the process of cleavage.
The cell cycle of cleavage during the early stages of development is very rapid. As
an example, the average cell cycle in the cell division of frogs is approximately 16
hours, while early embryonic cells complete their cycle in 30 minutes. This is
because the cell cycle lacks phases equivalent to the extent of the G1 and G2
phases. In other words, cell division takes place through repetition of the DNA
synthesis period (S phase) and the mitotic period (M phase), making the cell cycle
very short (see Chapter 9). In the early stages of development, a certain number of
cells are thus created by the repetition of cell division within a short space of time.
Cleavage patterns are diverse, and include a type with unique cleavage
directions as well as one with uneven blastomere sizes (Fig. 10-2). This diversity
is due to the uneven distribution of nutrients in the embryo, or may reflect the need
to unevenly distribute maternal factors localized in the cytoplasm (see IV in Chapter
9). Among these, the uneven distribution of maternal factors plays a particularly
important role in the developmental process. This can be easily understood by
taking the destiny of the blastomeres in nematodes as an example (Fig. 10-3). In
this case, in each cell division, germ cell granules (maternal factors necessary for
the embryo to form germ cells) are unevenly allocated to just one of the two cells.
Those that receive the granules remain in the germ cell line and finally become
Figure 10-1 Oogenesis in fruit f l iesOogenesis in fruit flies occurs with the help of cells known as nurse cells. A germ cell divides four times to form sixteen cells, of which only one becomes an oocyte (an immature egg in the process of oogenesis). The rest become nurse cells and facilitate the creation of the oocyte. The cytoplasm of the oocyte and nurse cells is connected, and many proteins and mRNA (i.e., the mRNA of maternal factors such as nanos and bicoids) that are synthesized in nurse cells are transported through this pathway to the oocyte. Many of these materials are unevenly stored in the oocyte. → indicates the direction of transport of materials synthesized in nurse cells.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 193
germ cells. All others become somatic cells that form various tissues and organs.
Cell lineage describes which cells become the cells of which tissues during the
process of a fertilized egg becoming an adult. (see the Column on p.194).
I I I . Determination of Embryo Directionality
The direction of the body is determined immediately after fertilization in terms of,
for example, which side of the embryo becomes the head/tail, and maternal
factors pre-stored in the oocyte play a pivotal role in this process. Since the
importance of these factors in development has been studied in depth for fruit flies
and frogs, these organisms are used as examples in the discussions below.
Figure 10-3 Cleavage and the fate of blastomeres in nematodesIn each cleavage, germ cell granules are unevenly allocated to one of the two cells. Those that receive the granules remain the germ cell line and become germ cells, while all others become somatic cells.
Figure 10-2 Cleavage patternsThere are various types of cleavage, which are classified by the direction of division and the size of blastomeres.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 194
Cell lineage, like a family tree, is a line that shows the history of a fertilized
egg going through cleavage and subsequently being differentiated into
various parts of the body. Since humans consist of an enormous number of
cells (60 trillion in an adult), it is impossible to trace the history of all cells
from fertilized egg to adult body. However, in nematodes, which measure
only 1 mm in length, the number of cells in an adult body is only 959. The
initial number is 1,090, but 131 cells are programmed to die during the
developmental process (this is known as apoptosis; see Chapter 9). It is
therefore easy to create a cell lineage for nematodes, as they consist of such
small numbers of cells. Indeed, a cell lineage map for nematodes, from the
fertilized egg to all cells that make up a mature individual, has already been
created (Column Fig. 10-1). Furthermore, the complete DNA sequence of
nematodes (consisting of approx. 100 million base pairs) has been
determined, and it is estimated that approximately 17,500 genes are
distributed over six chromosomes. As multicellular organisms on an easily
manageable scale, nematodes have therefore been widely used as a
material suitable for analyzing the basic mechanisms of organisms at the
molecular level, including developmental mechanisms, functional analysis of
the neural network and the causes of diseases.
Column Cell Lineage of Nematodes
Column Figure 10-1 Cell l ineage of nematodesIn nematodes, the lineage of all cells – from the fertilized egg (zygote) to the 959 somatic cells that make up an adult individual – has been determined.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 195
Development of Fruit Flies
In fruit flies, as shown in Figure 10-1, a number of maternal factor types that
determine the directionality of the embryo (i.e., the head and tail sides) are
stored in the oocyte during oogenesis. Among these, the major factors are mRNA
for two protein – bicoid and nanos. These mRNAs are stored disproportionately
in both sides of the oocyte. After fertilization, they are translated and the
concentration gradient of bicoid and nanos is created along the lengthwise axis
of the embryo (Fig. 10-4).
In the early stages of development in fruit flies, only the nucleus divides, and the
plasma membrane that envelops it is not newly created; the embryo therefore takes
on the appearance of a multinucleated cell. At this stage, all the nuclei are in the
same cytoplasm. Under these circumstances, when the concentration gradient of
bicoid and nanos is created in the embryo, the germ cell nuclei are directly
exposed to the gradient. Since bicoid and nanos are factors that regulate gene
expression and protein synthesis, the expression of new genes and protein synthesis
take place in accordance with their concentration gradients (Fig. 10-4).
The expression of gap genes occurs in accordance with the concentration
gradient patterns of bicoids and nanos, and the proteins translated from gap
genes are known as transcription factors. As a result, based on the expression
pattern of the gap genes, pair-rule genes are expressed in a way that forms
seven stripes in the embryo (this mechanism is discussed later). The proteins
translated from the pair-rule genes are also transcription factors. Then, based on
the expression pattern, segment-polarity genes are expressed in a way that
forms 14 stripes in the embryo. As a result of the cascade expression of these
gene groups, 14 regions (or compartments) are formed in the embryo, which is
Figure 10-4 The process from determination of directionality in the fruit - f ly embryo to segmentation of the embryo into 14 partsThe gene expression pattern observed during the process of bicoid and nano protein expression and the subsequent segmentation of the embryo into 14 parts is shown here. See Column Figure 10-2 on homeobox genes.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 196
the origin of the 14 basic regions of fruit flies (i.e., the segmental structure of
insects). The cascade expression of these gene groups takes place in accordance
with the concentration gradient patterns of transcription factors (Fig. 10-5), which
are the proteins that regulate the expression of target genes by binding to their
regulatory regions (see Chapter 4). Such regulation may enhance or suppress the
expression of target genes. Figure 10-5A shows an example of the expression of
a target gene being enhanced only in regions where the transcription factor
concentration is within a certain range, and otherwise being suppressed. In this
case, the expression of target genes is induced when the concentration is within
a certain range. Figure 10-5B shows an example of the expression of a target
region occurring in an area where the action of enhancing transcription factors
dominates that of inhibitory transcription factors.
During the process from the formation of the concentration gradients of bicoids
and nanos to the expression of segment polarity genes, 14 regions are formed
in the embryo based on the expression pattern of the genes. As the next step, the
fate of the 14 regions (in terms of the organs to be created from each) must be
determined. A group of genes called homeobox genes play an important role in
this process (Column Fig. 10-2).
Figure 10-5 Patterns of a new gene being expressed in accordance with the transcription factor concentration gradient(A) An example of a gene being expressed only in a region where the transcription factor concentration is within a certain range. In this case, upper and lower limits (thresholds) of the transcription factor concentration causing gene expression exist. (B) An example of a gene being expressed in a region where the concentration balance of transcription factors that inhibit gene expression and those that enhance it is within a certain range. The enhancement of gene expression is based on the combined effects of the different promoting factors.
(A)
(B)

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 197
During the long history of research into fruit flies, many mutants with structural
abnormalities have been reported. Recent gene-level analysis has identified
a series of genes that cause structural abnormality in the body of fruit flies.
These are called homeotic gene complexes (HOM-C), and consist of a
group of eight genes (Column Fig. 10-2A). It has also been found that gene
groups equivalent to these exist in four clusters (HoxA-D) in mammals.
In addition to this correspondence in the sequence of genes in chromosomes
between HOM-C in fruit flies and Hox in vertebrates, their expression
patterns in the body are also similar. As an example, the expression pattern
of HOM-C in fruit flies (which is expressed in an anteroposterior direction
along the body) and that of Hox in vertebrates (which is expressed along the
head-tail axis) are very similar (Column Fig. 10-2B).
The series of genes contained in HOM-C and Hox are called homeobox
genes, as all proteins translated from such genes have a region consisting of
60 amino acids called a homeodomain. The homeodomain is a region
with a special tertiary structure for binding to DNA (Column Fig. 10-2C).
The proteins translated from homeobox genes are transcription factors that
regulate the expression of other genes by attaching to them.
It is believed that homeobox genes determine which region becomes which
organ after the rough regional division that takes place in the embryo.
Column Homeobox Genes
Column Figure 10-2 Homeobox genes(A) HOM-C in fruit flies and HoxA–D in mammals are shown here. ↔ indicates the correspondence between the two. (B) The similarity of gene expression between HOM-C in fruit flies and Hox in vertebrates. The homeobox genes in fruit flies are expressed in the body in accordance with the gene sequence on the chromosomes. This is also the case with homeobox genes in vertebrates. As an example in mice, a part of the homeobox genes expressed in the brain and the spinal cord is shown. The arrow shows the directionality of homeobox gene expression along the head-tail axis. (C) The tertiary structure of the region known as the homeodomain, which all proteins translated from homeobox genes have, is shown here. It has three α-helices with which it binds to DNA.
(A) (B)
(C)

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 198
Development of Frogs
Next, we discuss development of frog embryo as an example of vertebrate
development. The upper hemisphere of the frog egg, which is rich in pigment
and therefore dark, is called the animal pole, while the opposite hemisphere,
which is white, is called the vegetal pole. Before fertilization, the egg has only
animal-vegetal-pole directionality. However, upon fertilization, the future dorsal
and ventral sides are determined in the embryo. This is because the surface part
of the cytoplasm in the egg moves in a particular direction after fertilization. This
in turn causes the movement of the maternal factors that determine the dorsal side
of the embryo from the vegetal pole to near the equator on one side of the
embryo (Fig. 10-6). As a result, the side opposite the sperm entry point becomes
the future dorsal side. Thus, in the development of frogs too, maternal factors
stored locally in the egg play an important role in determining the directionality
of the embryo.
A protein called Dishevelled is among the important maternal factors that are
stored in the vegetal pole of the embryo and turn the region to which they move
into the dorsal side. This protein moves to a region where it causes the expression
of new genes, thereby inducing that side of the embryo to become the dorsal side
(Column Fig. 10-3). As a result, a region called the organizer, which determines
the dorsal side of the embryo, is created on the side that the Dishevelled moves
to. The basic structure of the frog’s body is formed around this region.
Figure 10-6 Determination of dorsal-ventral directionalityPost-fertilization movement in the surface part of the frog egg is shown here. Upon fertilization, part of the cytoplasm in the vegetal pole moves to a point near the equator on the side opposite the sperm entry point.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 199
Dishevelled suppresses the degradation of a transcription factor called
β–catenin. This means that β–catenin increases in the cytoplasm on the
side of the embryo to which Dishevelled moves. The increased amount of
β–catenin moves into the nucleus and causes the expression of its target,
the Siamois gene. Siamois (a transcription factor) causes the expression of
its target (the goosecoid gene) in cooperation with other transcription factors
known as Smads. The goosecoid gene has a strong effect in inducing the
formation of the dorsal side in the embryo.
IV. Cell Differentiation and Stem Cells
During the developmental process, cells derived from one fertilized egg become
units with various functions such as muscle cells, neurons and epithelial cells. This
phenomenon is called cell differentiation.
Embryonic cells in the early developmental stage have the potential to become
various cell types. These cells, which have not yet undergone differentiation, are
referred to as undifferentiated cells. However, as development proceeds, most
cells are differentiated into units with particular functions. Each differentiated cell
Column Figure 10-3 Process of Dishevelled determining the dorsal side of the embryoThe process of maternal factors determining the dorsal side of the embryo takes place mainly through the regulation of gene expression by transcription factors. During the process, maternal factors and extracellular signals collaborate.
Column Roles of Maternal Factors in Determining the Dorsal Side of the Frog Embryo

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 200
type shows the different gene expression pattern and level. (Fig. 10-7). In other
words, in differentiated cells, the expression of certain genes necessary to fulfill
their cellular functions is enhanced, whereas the expression of unnecessary genes
is suppressed.
In differentiated cells, the suppression of unnecessary genes is performed through
chemical modification of the genes themselves (or of proteins that bind to them)
or through the binding of special proteins to them. Such suppression may be
temporary or semi-permanent (see Chapter 4). If the suppression is unlocked –
thus allowing gene expression – the cells regain the potential to be transformed
into other cells.
Cells that retain the ability to differentiate into many cell types exist in the various
tissues of the human body. These are known as stem cells, and are believed to
be involved in the repair of damaged tissues. Attempts to collect, grow and
differentiate these cells in order to artificially create tissues and organs have
recently been made. The aim is to create tissues and organs using stem cells
obtained from patients and implant them back into the body. The method of
implanting the patient’s own tissues avoids immunological rejection and raises
fewer ethical issues than other methods (such as techniques that use human
embryonic cells).
Figure 10-7 Cell DifferentiationA model showing changes in the gene expression pattern and level during cell differentiation. The gene expression patterns occurring in three types of differentiated cell are schematically outlined. Differentiated cells have the same gene set as the fertilized egg, but the gene expression pattern and level differ.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 201
V. Induction and Morphogenetic Movement
Induction is a phenomenon that strongly influences the fate of adjacent cells and
tissues through its action on them. Induction occurs throughout the course of
embryonic development.
When induction (which is found in various phases of the developmental process)
is considered at the cellular level, a number of common mechanisms are found.
The three main methods by which one cell influences and changes the fate of
other cells are via secretory substances, molecules on the plasma membrane
surface, and gap junctions (Fig. 10-8). Whichever method is taken, one cell can
influence many adjacent cells.
In triploblastic animals, whose bodies consist of the three basic layers of
endoderm, mesoderm and ectoderm (including many types of animal from
planarians to humans), the embryo is roughly divided into three regions in the
early stages of development. This determines the fate of each region. By way of
example, the respiratory and digestive organs are formed from the endoderm,
muscle and connective tissues are formed from the mesoderm, and the central
nervous system and skin are formed from the ectoderm. Induction plays a role in
the differentiation into the three germ layers. This mechanism is discussed below
using mesoderm induction in frogs as an example.
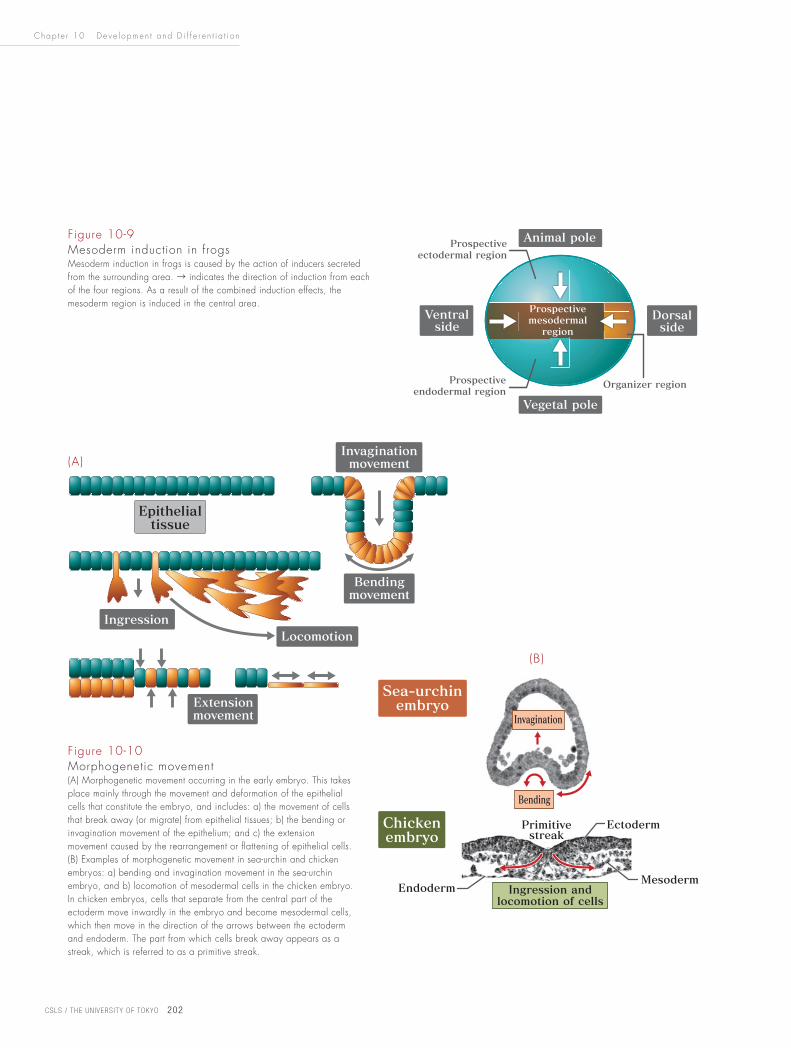
In mesoderm induction, the mesoderm is formed in a certain area of the embryo
(the equatorial region) as a result of influence from various other parts of the
embryo (Fig. 10-9). This induction is caused by many types of substance secreted
from the organizer on the dorsal side, the animal pole, the vegetal pole and
ventral side. Through their interaction, the mesoderm region is induced in the
intermediate area of the embryo (the equatorial region).
Morphogenetic Movement
After the determination of the rough arrangement of embryonic regions – such as
the prospective mesoderm and ectoderm areas – following the formation of the
organizer (the center of embryogenesis), major morphogenetic movement is seen.
First, the formation of three germ layers and morphogenetic movement occurs to
create archenterons, which later become the gastrointestinal tract. Second, further
morphogenetic movement occurs to create the neural tube. The distribution of
embryonic cells is rearranged through these morphogenetic movements, thereby
allowing the interaction of embryonic cells that previously existed separately.
Figure 10-8 Induction in adjacent cells(A) There are three methods of causing induction in adjacent cells: a) influence via secretory substances such as growth factors and hormones; b) influence through cell adhesion via proteins and carbohydrate chains that exist on the surface of the plasma membrane; and c) influence from the formation of gap junctions between the cell and adjacent cells. (B) Methods of transmitting induction to surrounding adjacent cells: a) through the diffusion of secretory substances; b) by extending cell protrusions and attaching them to surrounding cells; and c) intercellular transmission via gap junctions. → indicates the direction of induction.
(B)
(A)
Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 202
Figure 10-9Mesoderm induction in frogsMesoderm induction in frogs is caused by the action of inducers secreted from the surrounding area. → indicates the direction of induction from each of the four regions. As a result of the combined induction effects, the mesoderm region is induced in the central area.
Figure 10-10 Morphogenetic movement(A) Morphogenetic movement occurring in the early embryo. This takes place mainly through the movement and deformation of the epithelial cells that constitute the embryo, and includes: a) the movement of cells that break away (or migrate) from epithelial tissues; b) the bending or invagination movement of the epithelium; and c) the extension movement caused by the rearrangement or flattening of epithelial cells. (B) Examples of morphogenetic movement in sea-urchin and chicken embryos: a) bending and invagination movement in the sea-urchin embryo, and b) locomotion of mesodermal cells in the chicken embryo. In chicken embryos, cells that separate from the central part of the ectoderm move inwardly in the embryo and become mesodermal cells, which then move in the direction of the arrows between the ectoderm and endoderm. The part from which cells break away appears as a streak, which is referred to as a primitive streak.
(A)
(B)

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 203
Mesoderm formation, gastrulation and neurulation all take place through a number
of basic cell movements (Fig. 10-10). Since the early embryo is made of epithelial
tissue (see Chapter 11), morphogenetic movement during this stage mainly involves
the movement and deformation of epithelial cells. This includes the bending
movement caused by contraction on one side of epithelial cells, invagination
movement in which bent epithelial tissues extend inward in the embryo, locomotive
movement caused by cells separated from epithelial cells, and extension movement
caused by the flattening and rearrangement of epithelial cells.
Among these movement types, one that plays a particularly important role in
subsequent organogenesis is the large-scale movement of mesodermal cells. This
movement in vertebrates is so great that the cells move to the opposite side of the
embryo. Taking the chicken embryo as an example, cells that break away from
the epithelium of the ectoderm enter the embryo, form mesodermal cells and move
to the sides and the anterior part between the ectoderm and endoderm (Fig. 10-
11). The prospective heart mesoderm regions, which later become the heart, are
located at the tip of the moving mesoderm on both sides of the embryo.
Neural Induction
After mesoderm induction, neural induction occurs. By this process, the neural
tube (which later becomes the brain and the spinal cord) is created from the
ectoderm. Neural induction is an important event in the development of animals,
since the neural tube plays the central role in the construction of the body and
Figure 10-11Movement of mesodermal cells in the chicken embryoA schematic diagram showing the movement of mesodermal cells, seen from the dorsal side (the ectoderm is not shown). The prospective heart mesoderm region is located at the tip of the moving mesoderm. → indicates the direction of movement of mesodermal cells.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 204
forms the brain and the spinal cord – the most important components in the
bodies of animals.
Neural induction is brought about by the organizer and the mesoderm. First, the
organizer sends induction signals to the ectoderm. Then, induction signals are
sent from the mesoderm, which has moved below the ectoderm, to the ectoderm
itself. Both induction effects are exerted by substances secreted from cells. In
response to this induction, which destines the ectoderm to become nerve tissue,
the neural tube – the origin of the brain and the spinal cord – is formed from the
ectoderm. The brain is created from the anterior part of the tube, and the spinal
cord is created from the posterior part (Fig. 10-12).
Figure 10-12Neural induction in a frog embryoNeural induction exerted by the mesoderm to the ectoderm is shown here. Inducers are secreted from the anterior and posterior parts of the mesoderm that have moved inside the embryo, which respectively induce the formation of the brain and the spinal cord from the ectoderm.

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 205
As with animals, organogensis in plants is also regulated by various genes.
In this column, the formation of flower organs is discussed in connection with
homeotic genes. Generally, flowers consist of four organs: calyxes, petals,
stamens and pistils (carpels) (Column Fig. 10-A). These organs can be
depicted as a concentric ring structure of four layers (whirls) as in the floral
diagram (Column Fig. 10-4B). It has gradually become clear that the
formation of these organs is regulated by the combined expression of the
three homeotic gene groups of A, B and C (the ABC model, Column Fig.
10-4C). In the first whirl, only gene group A is expressed, forming calyxes.
In the second whirl, gene groups A and B are expressed, forming petals. In
the third whirl, gene groups B and C are expressed, forming stamens. In the
fourth whirl, only gene group C is expressed, forming pistils (carpels)
(Column Fig. 10-4C). Antagonism exists between gene groups A and C; if
the functions of gene group A are lost, the gene-group-C functions dominantly
over the entire flower, and if those of gene group C are lost, the gene-
group-A functions dominantly. Therefore, if the functions of gene group B are
lost, only gene group A is expressed in the first and second whirls, and only
gene group C is expressed in the third and fourth whirls, thereby forming a
flower consisting of calyxes, calyxes, carpels and carpels (Column Fig. 10-
4D). If gene C is lost, only gene group A is expressed in the first whirl, only
gene groups A and B are expressed in the second and third whirls, and only
gene group A is expressed in the fourth whirl, thereby forming a flower
consisting of calyxes, petals, petals and calyxes (Column Fig. 10-4E).
Column The Mechanism of Flower Organ Formation
Column Figure 10-4 The ABC model in f lowers(A) An Arabidopsis flower, and (B) an Arabidopsis floral diagram. The flower consists of four calyxes, four petals, six stamens and two carpels. (C) Different organs are formed through the combined effects of the three gene groups A, B and C. (D) A mutant that has lost the functions of gene group B. (E) A mutant that has lost the functions of gene group C.
(A)
(B)
(C)(D) (E)

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 206
VI. Organogenesis
The triploblastic structure is formed through a process in which the mesoderm
enters between the ectoderm and the endoderm. In concurrence with the
formation of the triploblastic structure, interaction between the germ layers occurs,
thereby initiating organogenesis. Induction phenomena like those shown in Figure
10-8 are seen when organs are formed through interaction between germ layers.
Here, organogenesis is discussed using the heart as an example.
The formation of the heart (the first of all organs to begin expressing its functions)
is initiated by mesoderm movement. This organ is formed from embryonic regions
collectively known as the prospective heart mesoderm. There are two such
regions located in the posterior part of the embryo. Together with the mesoderm,
these two regions move a long way from the posterior to the anterior part of the
embryo. They then merge in the anterior part, creating a tubular heart that
gradually changes shape to eventually form a heart consisting of atria and
ventricles (Fig. 10-13A).
While moving to the anterior part of the embryo, the prospective heart mesoderm
regions are influenced by inducers secreted from the endoderm and ectoderm,
thereby being induced to take the path to generate the heart. Through the
subsequent expression of homeobox genes, the regions then become destined to
form the heart.
A number of interesting findings have been made from the identification of the
inducers involved in heart formation and the genes that determine embryonic
regions to become the heart. Such findings include a structural similarity between
the homeobox gene that controls heart formation in vertebrates (the Nkx gene)
and the homeobox gene that controls the formation of dorsal blood vessels
(equivalent to the heart in mammals) in fruit flies (the Tinman gene) (Fig. 10-13B).
Another such finding is a similarity between the inducers (secretory substances)
that cause the expression of these genes in both fruit flies and vertebrates. These
findings indicate that the basic mechanism used in the formation of dorsal blood
vessels in fruit flies has been well conserved throughout evolution and is found in
heart formation in vertebrates. Similar cases have been confirmed in many
organs, including the formation of limbs and the nervous system.
Homeotic genes also play an important role in plant organogenesis. One such
example is the formation of flower organs (see the Column on p.205).

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 207
Figure 10-13 Cardiogenesis(A) The heart is formed by two prospective heart mesoderm regions moving from the left and right sides of the embryo and finally merging. First, a tubular heart is formed, which is then twisted clockwise in a sigmoid curve (this is reversed in the photo, since it was taken from the ventral side) to form the atria and ventricles, thus becoming the heart. The photo shows an amphibian (newt) heart consisting of two atria and one ventricle. (B) The similarities between vertebrates and fruit flies are shown with regard to inducers involved in cardiogenesis, the homeobox genes that determine heart formation, and the timing of expression for these genes. Bone morphogenetic factors and decapentaplegic, which induce the mesoderm to form the prospective heart mesoderm region, belong to the same growth factor group.
(B)
(A)

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
CSLS / THE UN IVERS ITY OF TOKYO 208
• While the developmental processes of organisms are diverse, a number
of common basic mechanisms are found.
• In the various phenomena that occur during the early stages of development
(such as the determination of embryo directionality and the destiny of
germ cells), substances derived from the mother (maternal factors) stored
in the egg play important roles.
• As development proceeds, the embryo is roughly compartmentalized into
regions. The fates of these regions (i.e., determination of which ones will
form the various tissues and organs) are then decided by the expression
of homeobox genes.
• The proteins translated from homeobox genes are transcription factors,
which have been well conserved in the evolutionary process. These play
an important role in forming the bodies of organisms.
• Throughout the development process, induction occurs in which cells in
certain regions influence those in adjacent regions. There are several
induction patterns, but in each case, induction causes the expression of
new genes in target cells, thereby determining the fate of those cells.
• Germ layers are formed in many animals, and interaction among these
layers causes the formation of tissues and organs. The large-scale
morphogenetic movement that occurs in the early stages of development
allows the formation of these germ layers and the subsequent interaction
among them.
• It is now possible to compare the genes involved in the development of
nematodes and fruit flies with those involved in human development. As a
result, it has been shown that genes with a similar structure play similar
functions in each. This indicates that the basic mechanisms forming the
bodies of organisms have been continuously passed on during the course of
evolution and function in a similar way in the formation of the human body.
Summary Chapter 10

Chap t e r 10 De v e l opmen t a nd D i f f e r e n t i a t i o n
10
CSLS / THE UN IVERS ITY OF TOKYO 209
[1]
The embryo of triploblastic organisms consists of three basic
structures. Name these structures and the organs and tissues
that develop from them.
[2]
Using fruit flies and frogs as examples, explain how the
anteroposterior axis of animals is determined in the early
stages of development.
[3]
Explain the concept of induction using examples.
[4]
Using the ABC model, explain how double flowers are
formed. Also outline which organs in a normal flower form
which organs in these cases.
Problems
(Answers on p.258)
![1 Historical-Comparative Methods CSLS/Boalt Miniseries on Empirical Research Methods, Workshop 2, Oct. 23, 2007 [See accompanying reference list] © 2007](https://img.pdfslide.us/doc/110x75/56649d6a5503460f94a483ff/1-historical-comparative-methods-cslsboalt-miniseries-on-empirical-research.jpg)




























