Embed Size (px)
Citation preview

1
•One of Mendel’s express aims was to understand how first generation traits (parents) would disappear in the second generation (F1 or hybrid) and reappear in the third generation (F2)
•Gregor Mendel conducted experiments with peas between 1857 and 1863
lPresented results in 1865
lPoorly received/understood: Karl Wilhelm von Nageli, a noted Swiss botanist. Nageli, whose thinking sometimes veered from science to mysticism, dismissed Mendel's work
Note: Correns, one of three re-discoverers in 1900, was a nephew of Nageli!
Slide 1 of Chapter 1R. Ward: Spring 2001
Chapter 1: Mendel’s breakthrough: patterns, particles and principles of
heredity
please read pages 10 through 13

2
Slide 2 of Chapter 1R. Ward: Spring 2001
Plant life cycles
From: Plant Life Cycles and Angiosperm Development: Susan R. Singer from Embryology: Constructing the Organism (S.F. Gilbert and A. M. Raunio, eds.) 1997. SinauerAssociates, Sunderland, MA.
Sperm and eggs= gametes (N)
Seed
Maternal tissue:
seed coat=ovule wall
Pod/fruit wall= ovary wall
First 2N cell=zygote
Endosperm is 3N2 copies of egg
chromosomes, 1 of sperm
•Review the life cycles of plants and be familiar with these terms:lSporophytic or maternal tissue
lGametes (sperm and eggs)
lpollination and Fertilization
lZygote

3
Slide 3 of Chapter 1R. Ward: Spring 2001
Mendel’s Experimental Organism
•experimental organism was the garden pea: advantages-
Ønaturally self pollinated which generates “true breeding” plants (later we’ll call these homozygous plants)
Øeasily cross fertilized or cross-pollinated
Øreadily available clear-cut alternative forms of traits
Øseed traits (color, shape) allowed for easy generation of large data sets

4
Slide 4 of Chapter 1R. Ward: Spring 2001
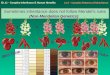
Mendel’s traits
ltwo of Mendel’s traits exhibited Xenia
•XENIA: genetic differences among pollen grains can manifest as phenotypic differences in seed set on female parent in P generation.
lXenia genes expressed in embryo (including cotyledons) or endosperm.
But not seed coat or pod attributes
lgenotypes of both egg and pollen determine phenotype
lphenotype of F1 generation manifest in F1 seeds produced on true breeding female parent.
•Phenotype could vary within a pod
•seed shape: round vs wrinkled – embryo/cotyledon trait
•seed color: yellow vs green
lMendel referred to this as “albumen” color
lactually is color of cotyledon, which is embyronic tissue
lfive of Mendel’s traits manifest in flowers, pods or stem morphology
•phenotype of F1 generation manifest in plants grown from seeds produced by cross pollination
•pod shape: inflated vs constricted
lNote: P generation pods contain F1 seed, and F1 pods contain F2 seeds, etc.!
lTherefore, parental generation pods have same genotype and phenotype irrespective of pollen genotype
•pod color: green vs yellow
•flower/pod position: axillary vs terminal
•stem length: standard vs dwarf

5
Slide 5 of Chapter 1R. Ward: Spring 2001
Fn seed vs. Fn plants
P
plant
P
plant
pollen
Male
(pollen source)
Female
(emasculated)
F1 seed
F1
F2 seed
F2 plants
P seed P seed
•P generation fruit tissue (ovary walls=pericarp, ovule walls=seed coat) is derived from cells that are direct descendents of the parental mitotic divisions of the zygote/embryo in the seed from which the P generation plant arose. All such tissue is therefore genetically identical on a single plant, even if the P generation plant was pollinated/fertilized by a genetically different plant.
•Seeds on P generation plants used as females in a cross-fertilization carry F1 generation embryos.
•Seeds can have three distinct tissues-
Øembryo, including cotyledons (can be different from mother plant)
Øseed coat (derived from maternal tissue just like the fruit wall or pericarp is)
Øendosperm- derived from union of two clones of the egg and one clone of the sperm
•The embryo represents the progeny, so fruits and seeds are bi-generational.
•This is important when considering mendel’s experiments since some traits are expressed in embryonic tissue which is enclosed within ovule walls (seed coats) and ovary walls (pericarp)

6
Slide 6 of Chapter 1R. Ward: Spring 2001
Xenia in Pea
P1 male•purple flower•yellow seeds•yellow pod
P1 female•white flower•green seeds•green pod
F1 plant•purple flower only••green pod only
F1 seed•all seed on female in the cross will have yellow seed (since seed color is an embryonic trait, and yellow is dominant to green)•but pods are maternal tissue and not affected by the pollen genotype
true breeding(all progeny from self pollination
identical to mother plant)
•segregation for xenia traits can be viewed among seeds within pods of F1 plants.
•segregation for non-xenia traits only manifest on plants derived from the seed on the F1 plants (i.e., F2 seed).

7
• Background: the historical puzzle of inheritance
•Genetics involves:
Øcareful observation of populations over generations
Øanalysis of carefully acquired data on individuals
Ødevelopment and testing of theoretical frameworks
•Mendel was the first person to put these three elements together to form a theory of inheritance
Øa great deal of observation took place in the time before Mendel
-huge efforts at cataloging species was conducted (Linnaeus, Darwin)
-cross fertilization within and among species was induced and observed
•Artificial selection was the first applied genetic practice
Øplants and animals were domesticated through selection of individuals that exhibited favorable traits (e.g., non-aggressiveness in the progenitors of domesticated dogs, horses, cattle etc.; longer seed retention in cereals, etc.)
•The puzzle of passing on desirable traits
Øby Mendel’s birth (1822), plant and animal breeders were able to generate individuals with new and valued combinations of traits by controlled matings
Øbut the value of these progeny as parents was unpredictable (i.e., the value of an individuals own progeny was unpredictable)
•Sheep breeding was a particularly important activity in Moravia, and at a 1837 conference of the Moravian Sheep Breeders Society, the Abbott Cyril Napp proposed that breeders needed to discover three things:
Øwhat is inherited
Øhow is it inherited, and
Øwhat is the role of chance in inheritance?
•before Mendel, blending, preformation (e.g., the idea of a homunculus) and other erroneous theories were proposed to explain the contradiction between 1) the obvious reality that members of a species had progeny who were of the same species; and 2) the equally obvious reality that variation of individual phenotype (appearance) exists within and among families
•Mendel’s work laid the foundation for our current view of heredity
•Key features of his approach include:
Øfocus on a single species, the naturally self-pollinated but easily cross-pollinated annual pea (Pisum sativum)
Øfocus on traits that had distinct, mutually exclusive (“antagoni
-furthermore- Mendel worked with only two forms of each of the seven traits he studied
Øselection and propagation of “pure-breeding lines” which upon self-pollination produce progeny that are collectively identical in form to their parents and to each other. Pure-breeding lines in a self-pollinated crop can be maintained in a pure state throughout multiple generations simply by isolating their progeny from other lines.
Øcareful control of matings between plants of different pure-lines, with attention to which parent was used as the female.
Øgeneration of large datasets consisting of observations on individual progeny plants
-note that the word ‘plant’ here is inclusive of the embryo within a seed.
Øcomparison of such results with predictions based on hypothetical models
Slide 7 of Chapter 1R. Ward: Spring 2001
Gregor Mendel: context and tools
Fig. 1.6 from Hartwell et al

8
Slide 8 of Chapter 1R. Ward: Spring 2001
Creating a monohybrid
P1 male•yellow seeds
P1 female•green seeds
true breeding(all progeny from self pollination
identical to mother plant)
F1 seed•all seed on female in the cross will have yellow seed (since seed color is an embryonic trait, and yellow is dominant to green)
•Genetic Analysis According to Mendel
•The embryos (i.e., progeny plants) that develop within the seeds of a plant used as a female in a cross (or hybridization) are deemed an “F1” generation if the parents differed in at least one trait.
•analyses of the progeny of plants grown from the F1 seed was the basis of Mendel’s work
•Mendel began his work by focusing on one trait at a time
Øanalysis of the frequencies of alternate forms of a single trait in the P, F1, subsequent generations of one cross is called “Monohybrid analyses”
Øin reality, the parents used in Mendel’s monohybrid crosses must have differed for other traits.
Øthe key is that Mendel ignored the other traits and focused on only one at a time (at first)

9
Slide 9 of Chapter 1R. Ward: Spring 2001
P, F1, and F2 generations
•Figure 1.9: Monohybrid analysis of seed color in pea (an embryonic trait)
•Mendel took pollen from a yellow seeded pure breeding line and placed it on stigmas of emasculated flowers of plants from a green seeded pure breeding line
•The F1 seeds (which matured in the pods on the female parent), were all yellow
Øthe reciprocal cross was also made (using the yellow seeded pure breeding line as the male parent)
Øagain, all of the F1 seeds were yellow
•Mendel planted the F1 seeds and allowed them to self pollinate and develop mature seed.
•The F2 seeds (which matured on the F1 plants) included both yellow and green forms of the seed color trait, in a ratio of approximately 3 yellow for every 1 green seed.
•These results refuted the concept of blending, since the green form of the trait in the P generation reappeared in the F2 generation after disappearing in the F1 generation.
•Mendel observed that there are two kinds of behavior of yellow pea derived plants:
Øsome breed true (have only yellow seed progeny upon self pollination)
Øsome generate both yellow and green seed upon self pollination
•Mendel hypothesized that
Ø1) for each trait, each plant carries two discrete copies of a unit of inheritance (which we now call genes)
Ø2) these genes come in two forms, which we now call “alleles”
Ø3) one allele is dominant and the other is recessive
-the yellow allele of the seed color gene in pea is dominant to the green allele of the same gene.
•The P generation plants in Fig. 1.9 either carry two dominant alleles (the yellow parent), or two recessive alleles (the green parent)
•F1 generation seeds in Figure 1.9 carry one yellow and one green allele.
•end
•end

10
Slide 10 of Chapter 1R. Ward: Spring 2001
The Law of Segregation
•Figure 1.10: The law of segregation is two part
Ø1) gametes (sperm and eggs) contain only one allele of a given gene;
-pure breeding yellow peas form gametes that all have the yellow allele, and
-pure breeding green peas form gametes that all have the green allele
-F1 generation plants form both kinds of gametes in equal quantities ( ½ with the yellow allele, and ½ with the green allele).
-in other words, both alleles can reside in and pass through an F1 generation without any change in their fundamental behavior
-so alleles are SEGREGATED from each other in gamete formation
Ø2) Individual progeny are created from the random union of one male gamete and one female gamete.

11
Slide 11 of Chapter 1R. Ward: Spring 2001
Self pollination of the F1 Generation
•gametes have only one allele (Y or y)
•the two alleles of an F1 appear in equal frequencies in that F1’s gametes
•since the frequency of ‘Y’ equals the frequency of ‘y’ in both eggs and pollen of an F1,
•the probability of a random male gamete carrying ‘Y’ = 0.5, which equals the probability of it carrying the ‘y’ allele
•gametic union in self fertilization is random, leading to four possible ways to form a zygote:
Ø (egg-derived written first): Yy, YY, yY, and yy
•each of those outcomes has a probability of ¼, by two lines of reasoning:
Ønone of the four is more likely than the other, so they are equally probable
Øthe “Law of the Product” states that the probability of two (or more) independent events occurring simultaneously ( or sequentially) equals the product of the probabilities of the individual events.

12
Slide 12 of Chapter 1R. Ward: Spring 2001
The outcomes of two “events” are independent if they satisfy two constraints:
v1) the outcome of one event doesn’t influence the outcome of the other event; and v2) they can occur together (i.e., they are not
mutually exclusive)vexamples: Øtwo consecutive coin tosses- one coin doesn’t
influence the other; and it’s possible to doØa ‘Y’ egg combining with a ‘y’ sperm- the
probability of the egg being ‘Y’ is not affected by the identity of the sperm; and zygotes always derive from one sperm and one egg

13
Slide 13 of Chapter 1R. Ward: Spring 2001
Law of the Sum
v if an event can be achieved by two or more if an event can be achieved by two or more mutually exclusive routes, the probability of the mutually exclusive routes, the probability of the event equals the sum of all possible routes.event equals the sum of all possible routes.
v the �the �YyYy� genotype in the F2 generation depicted in � genotype in the F2 generation depicted in both Figures 1.9 and 1.11 can occur in one of two both Figures 1.9 and 1.11 can occur in one of two mutually exclusive routes:mutually exclusive routes:Ø
�Y� from the male�Y� from the malevSince the probability of both routes equals 0.5, and Since the probability of both routes equals 0.5, and
they are mutually exclusive, we add the they are mutually exclusive, we add the probabilities to determine the probability of a probabilities to determine the probability of a heterozygous F2 plant.heterozygous F2 plant.
•“events” are in the eye of the beholder
•possible events in the context of genetics:
Øthe genotype of a gamete randomly selected from one plant
Øthe phenotype of a plant randomly selected from a population of plants
Øa zygote is formed by union of an egg carrying ‘Y’ and a sperm c
Øa zygote is formed by union of an egg carrying ‘Y’ and a sperm c

14
Slide 14 of Chapter 1R. Ward: Spring 2001
Laws of probability in Action
two mutually exclusive ways to achieve Yy
just one way to form YY or yy

15
Slide 15 of Chapter 1R. Ward: Spring 2001
Frequencies and ratios
vFrequencies are used in genetics to represent the proportion that one class of individuals represents of a whole population.
vA list of the frequencies of all classes is a “frequency distribution”. A complete frequency distribution must sum to 1.0 .
vWe often speak of “ratios” in genetics. Frequency distributions are converted to ratios by using the numerator of the fractional frequencies after application of a common denominator.

16
Slide 16 of Chapter 1R. Ward: Spring 2001
Frequencies and corresponding ratios: examples
vBelow are examples of frequency distributions and their corresponding ratios
Frequency distribution Ratio0.75 yellow, 0.25 green 3 yellow : 1 green0.25 YY, 0.5Yy, 0.25 yy 1 YY : 2 Yy : 1 yy0.8 tall, 0.2 short 4 tall : 1 short

17
Slide 17 of Chapter 1R. Ward: Spring 2001
Inter-converting frequencies and ratiosv Assume you have 25 blue, 50 green, and 25 black marbles for a
total of 100 marbles.v The Frequency distribution of marble colors is 1/4 blue : 1/2 green
: 1/4 black .v The ratio of marble colors is 1 : 2 : 1 (blue/green/black)v To convert from fractional frequencies to ratios,
Ø apply a common denominator to each frequency, (in this case the common denominator is 4, so the freq. dist. becomes 1/4 blue : 2/4 green : 1/4 black).
Ø Then use the numerators to represent the ratios: 1 blue : 2 green : 1 black.
v To convert a ratio to frequencies, Ø sum each of the values in the ratio expression (in this case 1, 2 and
1, which sums to 4) ;Ø and use that as the denominator under each of the ratio values. So
a ratio of 1 : 2 :1 translates into a frequency distribution of 1/4 : 2/4 : 1/4.

18
Slide 18 of Chapter 1R. Ward: Spring 2001
Ratios and Frequencies of a subset of classes
v Ratios among classes remain constant even if one or more classesis discarded
v If we discard all of the black marbles from the slide above, theratio of blue to green marbles is unchanged and remains:Ø 1 blue : 2 green
v But the frequencies in an original population do not equate withthe correct frequencies in a subset of classes. We had 1/4 blue and 1/2 green when we had the black marbles included, but that cannot be a complete frequency distribution since it does not sum to 1.0 .
v To derive the correct frequencies of a subset of classes, first write down the ratios (1 : 2), and then convert that to a frequency distribution. The sum of ratio values is 1+2=3, so using 3 as the denominator, the ratio of 1 : 2 converts to a frequency distribution of 1/3 and 2/3.

19
Slide 19 of Chapter 1R. Ward: Spring 2001
Frequencies and Ratios: an application (1 of 2)
¼ ¼ ¼ ¼
2/4 Yy1/4 YY
•Ratio of homozygous to heterozygous Yellow F2 Progeny: 1 YY : 2 Yy; •Which is equivalent to a frequency distribution of 1/3 YY and 2/3 Yy
1/4 yy
From fig. 1.12, Hartwell
•When we look only at the yellow progeny we are
•If you know the ratio of the occurrences of two events (for instance 1 YY for every 2 Yy), you can convert the ratio into frequency distribution (frequency of each type of event- 1/4, 1/2, etc.) by summing the values in the ratio ( in this case 2+1) and using that sum as a denominator for each events ratio value.

20
Slide 20 of Chapter 1R. Ward: Spring 2001
Frequencies and Ratios: an application (2 of 2)
YY Yy
2/3 Yy1/3 YY
Yellow F2’s
Self pollination
2/3 * [1/4 YY 2/4 Yy 1/4 yy]= 2/12 YY 4/12 Yy 2/12 yy
1/3 * [1/1 YY]=4/12 YY
F3
Yellow F3’s
Ratio 6(=4+2) YY : 4 Yy :2 yy
Ratio 6 YY : 4 YyFrequencies 6/10 YY, 4/10 Yy; or 3/5 YY, 2/5 Yy
•Why multiply the frequencies of genotypes derived from Yy by 2/3?
•Because two independent events are required for a given genotype to occur in the progeny of a Yy F2 plant.
Øfirst, the F2 had to be Yy. That probability is 2/3
Øsecond, the probability of, say, YY is 1/4 in the progeny of a Yy F2
•Since these two events are independent, we multiply the two probabilities: 2/3 * 1/4 = 2/12

21
Slide 21 of Chapter 1R. Ward: Spring 2001
Some Definitions
•Phenotype- appearance
•Genotype- Alleles present in cells of a plant for one or more genes
•Homozygous- both alleles of a gene are identical
•Heterozygous- the two alleles are different

22
Slide 22 of Chapter 1R. Ward: Spring 2001