Embed Size (px)
Citation preview

Chap. 7 Transcriptional Control of Gene Expression (Part C)
Topics• Molecular Mechanisms of Transcription Repression and
Activation
• Regulation of Transcription Factor Activity
Goals• Learn how activators &
repressors control chromatin structure and pre-initiation complex assembly.
• Learn how the activities of activators and repressors are controlled.
Transcriptionally active polytene chromosomes
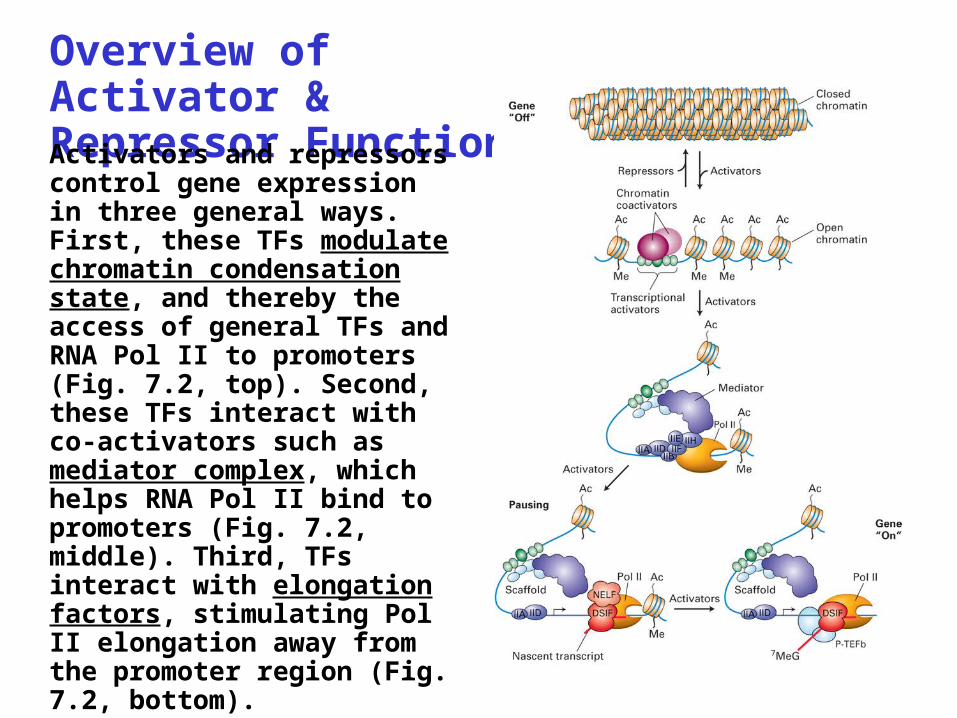
Overview of Activator & Repressor FunctionActivators and repressors control gene expression in three general ways. First, these TFs modulate chromatin condensation state, and thereby the access of general TFs and RNA Pol II to promoters (Fig. 7.2, top). Second, these TFs interact with co-activators such as mediator complex, which helps RNA Pol II bind to promoters (Fig. 7.2, middle). Third, TFs interact with elongation factors, stimulating Pol II elongation away from the promoter region (Fig. 7.2, bottom).

Co-activators & Co-repressorsThe distinction between an activator/co-activator and repressor/co-repressor is based on whether or not the protein binds specifically to DNA. Namely, activators/repressors have DNA binding domains that allow them to bind to DNA. Co-activators/co-repressors typically don’t bind to specific sequences in DNA. They typically exert their effects on transcription initiation via protein-protein interactions within transcription initiation complexes at promoters, or by modifying histone tails. Most of our discussion about activator and repressor functions will center on yeast as less currently is known about their functions in higher eukaryotes.

Interphase ChromatinInterphase chromatin exists in two different condensation states (Fig. 6.33a). Heterochromatin is a condensed form that has a condensation state similar to chromatin found in metaphase chromosomes. Euchromatin is considerably less condensed. Heterochromatin typically is found at centromere and telomere regions, which remain relatively condensed during interphase. The inactivated copy of the X-chromosome (Barr body) that occurs in cells in females also occurs as heterochromatin. In contrast, most transcribed genes are located in regions of euchromatin. Common modifications occurring in histone H3 in hetero- and euchromatin are illustrated in Fig. 6.33b.
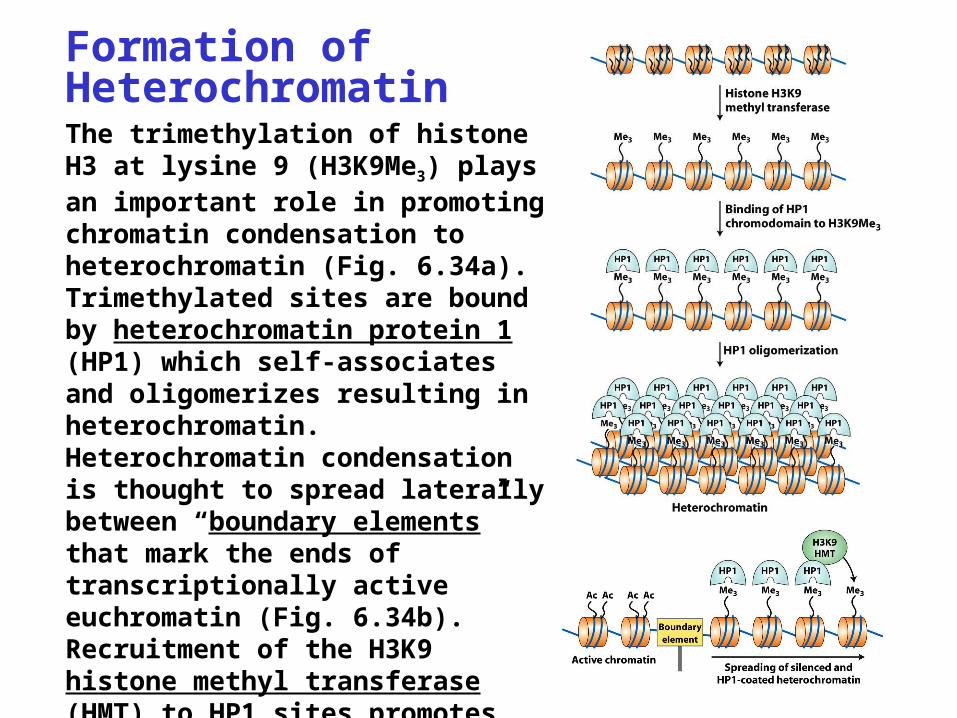
Formation of HeterochromatinThe trimethylation of histone H3 at lysine 9 (H3K9Me3) plays an important role in promoting chromatin condensation to heterochromatin (Fig. 6.34a). Trimethylated sites are bound by heterochromatin protein 1 (HP1) which self-associates and oligomerizes resulting in heterochromatin. Heterochromatin condensation is thought to spread laterally between “boundary elements” that mark the ends of transcriptionally active euchromatin (Fig. 6.34b). Recruitment of the H3K9 histone methyl transferase (HMT) to HP1 sites promotes heterochromatin spreading by catalyzing H3 methylation.
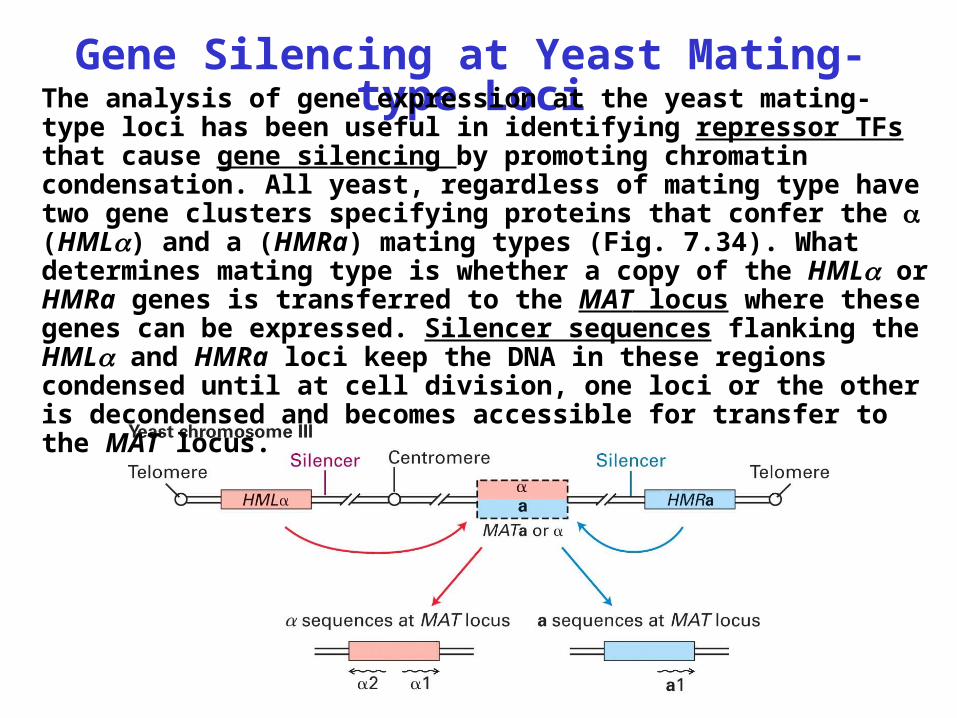
Gene Silencing at Yeast Mating-type LociThe analysis of gene expression at the yeast mating-type
loci has been useful in identifying repressor TFs that cause gene silencing by promoting chromatin condensation. All yeast, regardless of mating type have two gene clusters specifying proteins that confer the (HML) and a (HMRa) mating types (Fig. 7.34). What determines mating type is whether a copy of the HML or HMRa genes is transferred to the MAT locus where these genes can be expressed. Silencer sequences flanking the HML and HMRa loci keep the DNA in these regions condensed until at cell division, one loci or the other is decondensed and becomes accessible for transfer to the MAT locus.
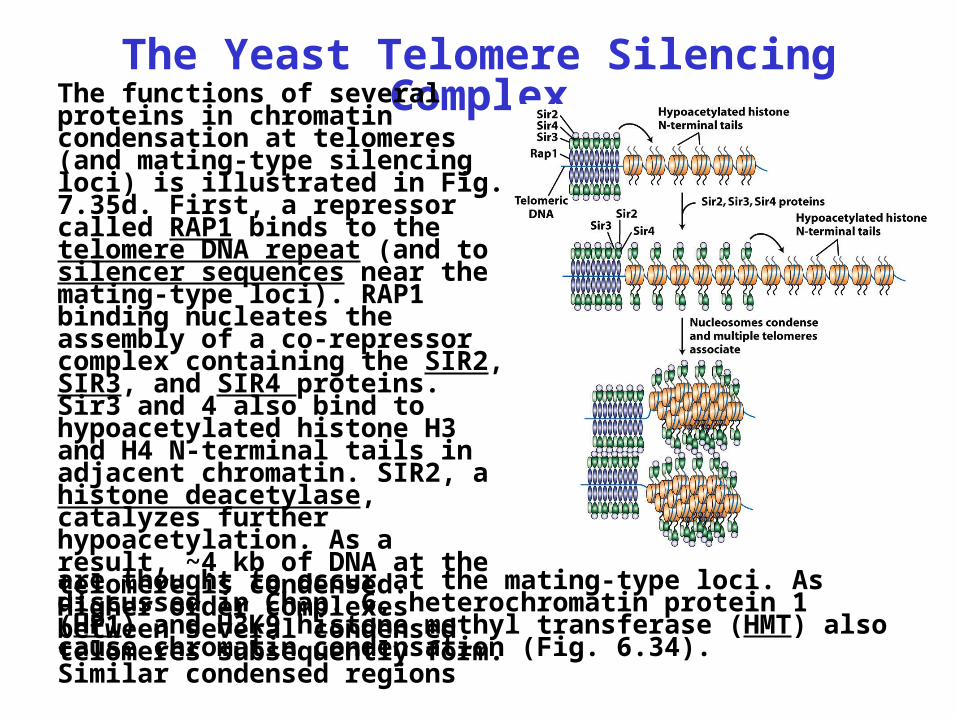
The Yeast Telomere Silencing ComplexThe functions of several
proteins in chromatin condensation at telomeres (and mating-type silencing loci) is illustrated in Fig. 7.35d. First, a repressor called RAP1 binds to the telomere DNA repeat (and to silencer sequences near the mating-type loci). RAP1 binding nucleates the assembly of a co-repressor complex containing the SIR2, SIR3, and SIR4 proteins. Sir3 and 4 also bind to hypoacetylated histone H3 and H4 N-terminal tails in adjacent chromatin. SIR2, a histone deacetylase, catalyzes further hypoacetylation. As a result, ~4 kb of DNA at the telomere is condensed. Higher order complexes between several condensed telomeres subsequently form. Similar condensed regions
are thought to occur at the mating-type loci. As discussed in Chap. 6, heterochromatin protein 1 (HP1) and H3K9 histone methyl transferase (HMT) also cause chromatin condensation (Fig. 6.34).
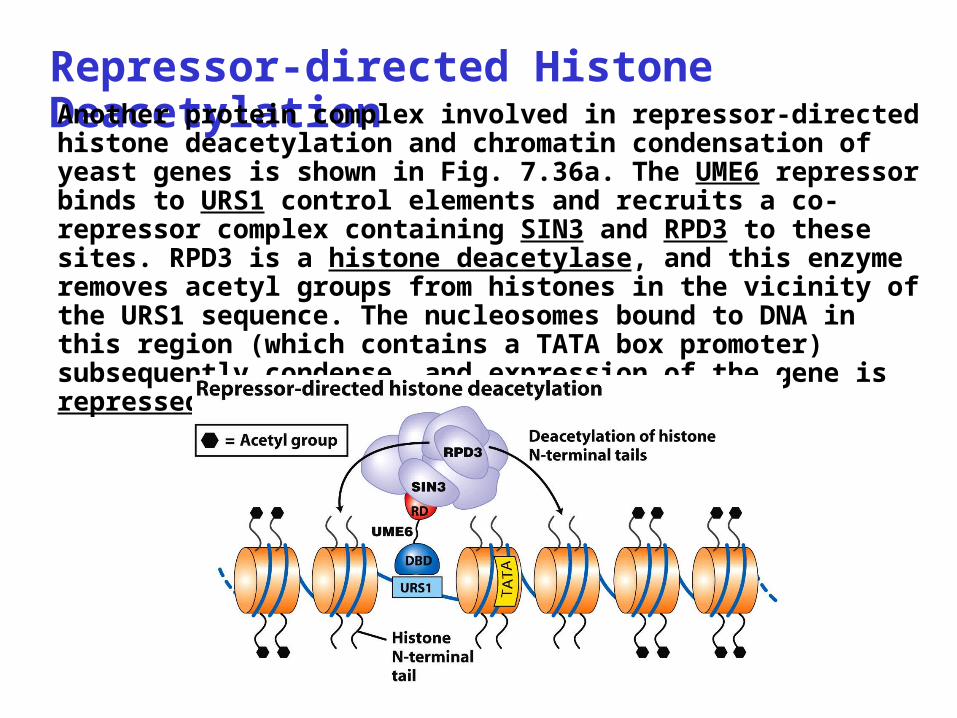
Repressor-directed Histone DeacetylationAnother protein complex involved in repressor-directed histone deacetylation and chromatin condensation of yeast genes is shown in Fig. 7.36a. The UME6 repressor binds to URS1 control elements and recruits a co-repressor complex containing SIN3 and RPD3 to these sites. RPD3 is a histone deacetylase, and this enzyme removes acetyl groups from histones in the vicinity of the URS1 sequence. The nucleosomes bound to DNA in this region (which contains a TATA box promoter) subsequently condense, and expression of the gene is repressed.

Activator-directed Histone HyperacetylationYeast genes can be turned on by histone acetylation and
chromatin decondensation (Fig. 7.36b). In the example shown, the GCN4 activator first binds to its UAS upstream of the TATA box of a regulated gene. GCN4-UAS binding recruits a co-activator complex containing the GCN5 histone acetylase to the site. Through the activity of GCN5, histone N-terminal tails are hyperacetylated leading to chromatin decondensation and formation of beads-on-a string type chromatin. General TFs and RNA Pol II are then able to interact with the promoter, and the gene is transcribed. Multisubunit proteins called chromatin remodeling complexes also are important in decondensing chromatin.
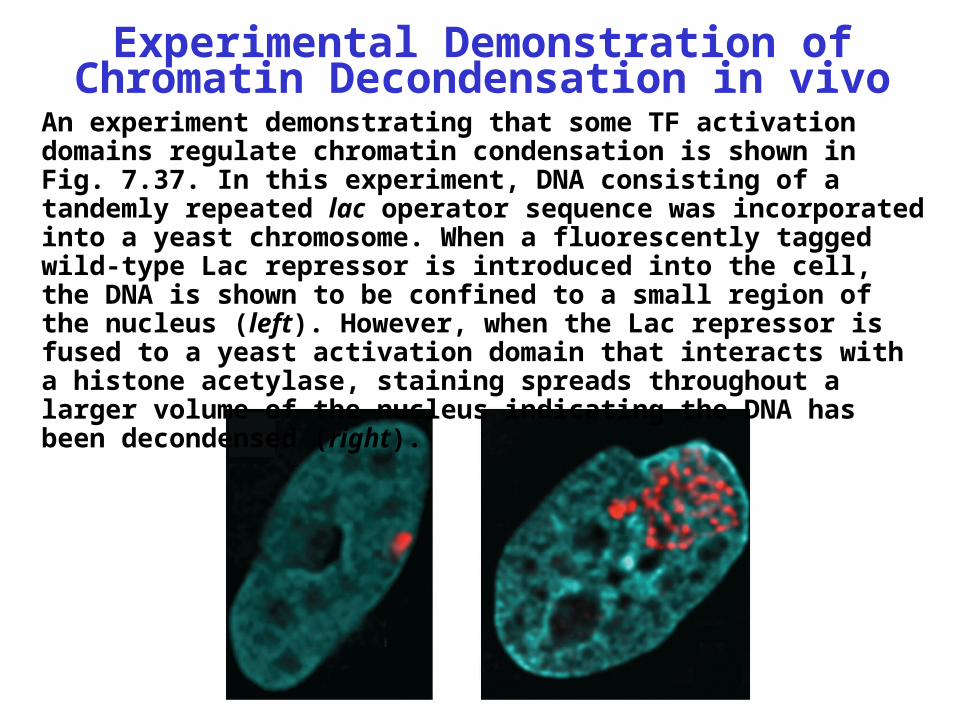
Experimental Demonstration of Chromatin Decondensation in vivo
An experiment demonstrating that some TF activation domains regulate chromatin condensation is shown in Fig. 7.37. In this experiment, DNA consisting of a tandemly repeated lac operator sequence was incorporated into a yeast chromosome. When a fluorescently tagged wild-type Lac repressor is introduced into the cell, the DNA is shown to be confined to a small region of the nucleus (left). However, when the Lac repressor is fused to a yeast activation domain that interacts with a histone acetylase, staining spreads throughout a larger volume of the nucleus indicating the DNA has been decondensed (right).

Mediator ComplexMediator complex is a multisubunit co-activator that helps load RNA Pol II onto promoters. Some mediator subunits bind to RNA Pol II and others bind to TF activation domains. The structure of yeast mediator bound to RNA Pol II is shown in Fig. 7.38a. The subunit composition of human mediator complex is shown in Fig. 7.38c. Some mediator subunits exhibit histone acetylase activity.
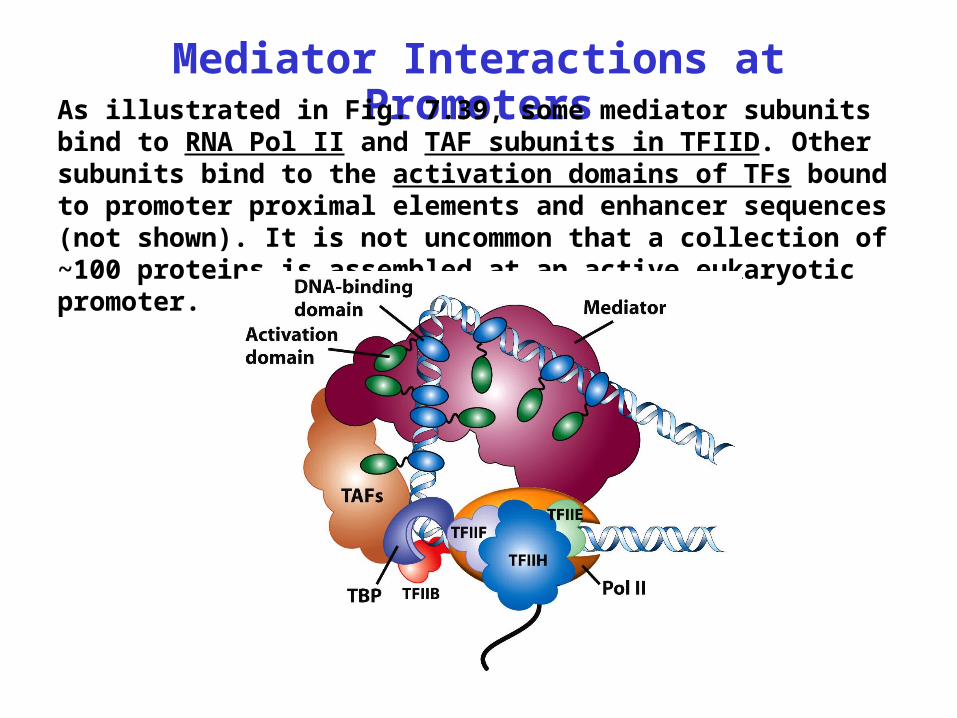
Mediator Interactions at PromotersAs illustrated in Fig. 7.39, some mediator subunits bind to RNA Pol II and TAF subunits in TFIID. Other subunits bind to the activation domains of TFs bound to promoter proximal elements and enhancer sequences (not shown). It is not uncommon that a collection of ~100 proteins is assembled at an active eukaryotic promoter.

Modular Structure of Activators IIFunctional domains in activators are joined by flexible protein linker sequences (Fig. 7.27). Due to the presence of linkers, the spacing and location of DNA control elements often can be shifted without interfering with DNA binding and regulation of promoters. The evolution of gene control regions through shuffling of DNA binding sequences between genes may have been favored due to the lack of strong requirements for control element spacing and location. Furthermore, the evolution of new activator protein genes through domain swapping has probably also been facilitated by linker sequences.

Yeast Two-hybrid System (I)The flexibility of activator-mediated gene transcription in eukaryotes is exploited in the widely used yeast two-hybrid system, which is an in vivo system for identification of interacting pairs of proteins. In this method, genes encoding hybrid proteins containing yeast DNA-binding and activation domains are first constructed (Fig. 7.40a). The "bait hybrid" contains the GAL4 DNA binding domain fused to the "bait domain" which is one of the proteins being tested. The "fish hybrid" contains the second protein, the "fish domain”, fused to a GAL4 activation domain. If the bait and fish domains can interact, then the two hybrid proteins can activate transcription of a reporter gene (e.g., HIS) (Fig. 7.40b).
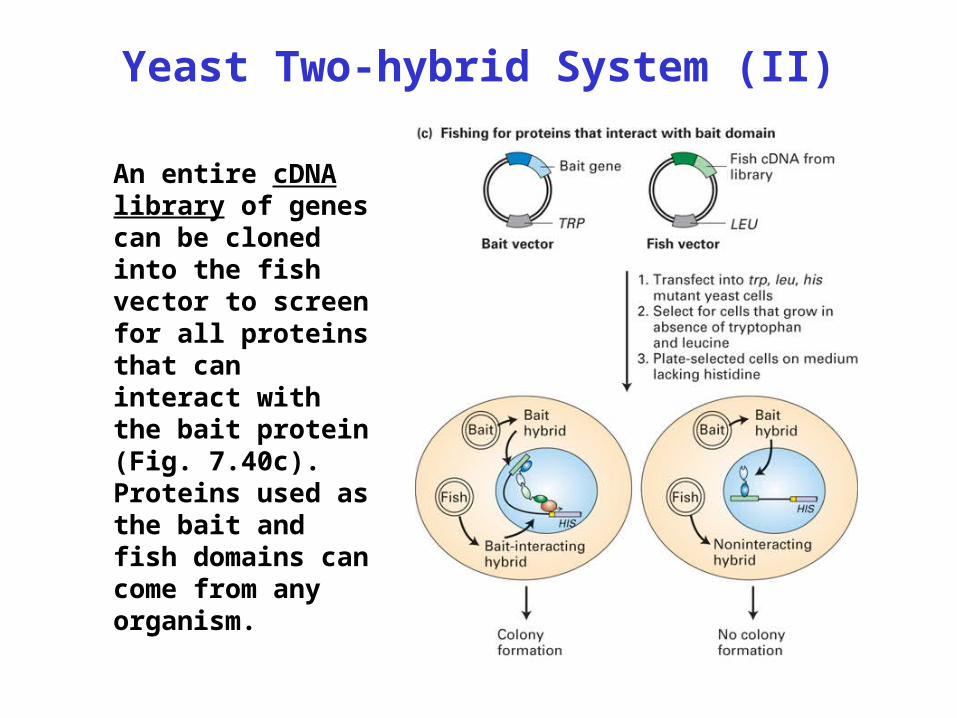
Yeast Two-hybrid System (II)
An entire cDNA library of genes can be cloned into the fish vector to screen for all proteins that can interact with the bait protein (Fig. 7.40c). Proteins used as the bait and fish domains can come from any organism.
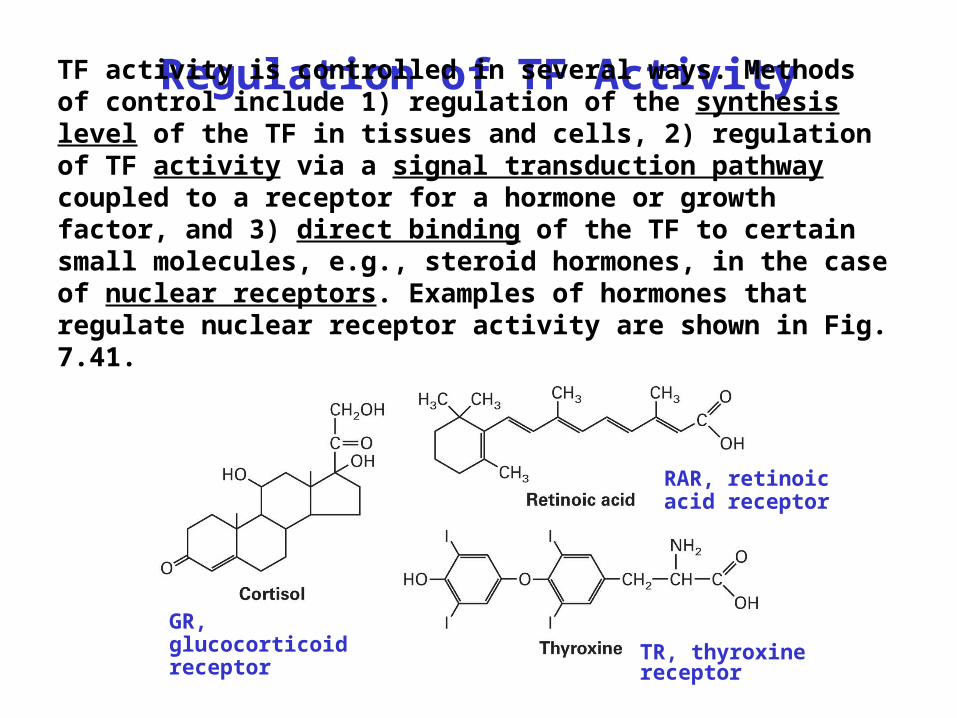
Regulation of TF ActivityTF activity is controlled in several ways. Methods of control include 1) regulation of the synthesis level of the TF in tissues and cells, 2) regulation of TF activity via a signal transduction pathway coupled to a receptor for a hormone or growth factor, and 3) direct binding of the TF to certain small molecules, e.g., steroid hormones, in the case of nuclear receptors. Examples of hormones that regulate nuclear receptor activity are shown in Fig. 7.41.
GR, glucocorticoid receptor
RAR, retinoic acid receptor
TR, thyroxine receptor
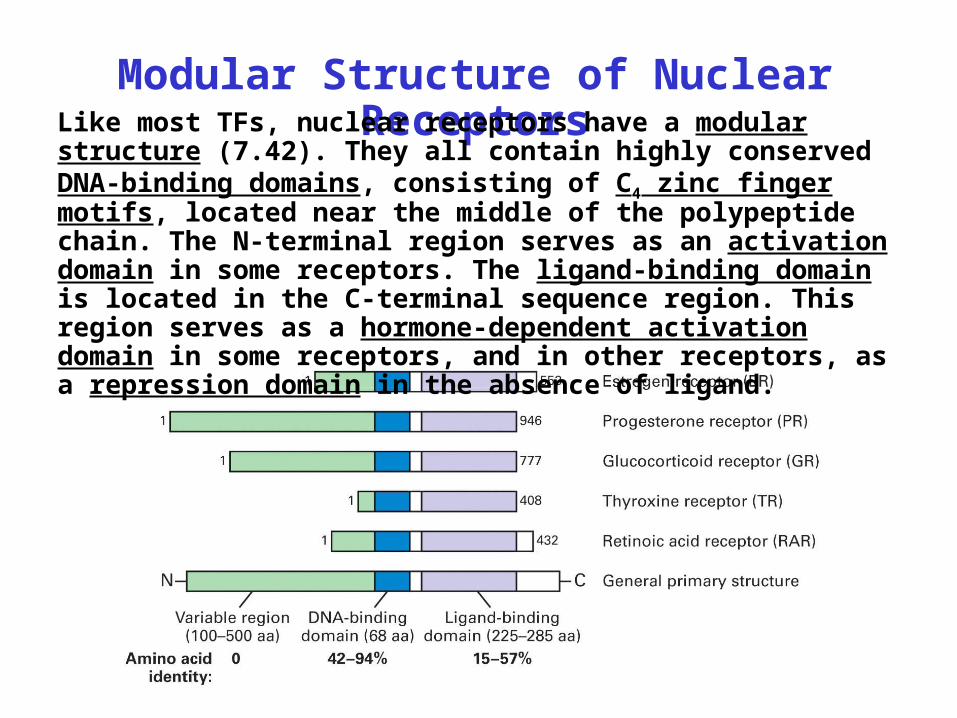
Modular Structure of Nuclear ReceptorsLike most TFs, nuclear receptors have a modular
structure (7.42). They all contain highly conserved DNA-binding domains, consisting of C4 zinc finger motifs, located near the middle of the polypeptide chain. The N-terminal region serves as an activation domain in some receptors. The ligand-binding domain is located in the C-terminal sequence region. This region serves as a hormone-dependent activation domain in some receptors, and in other receptors, as a repression domain in the absence of ligand.

Nuclear Receptor DNA Response ElementsThe DNA sequence elements bound by some nuclear
receptors are shown in Fig. 7.43. The conservation of these sequences is reflected in the structural conservation of the DNA binding domains. The GR and ER receptors bind DNA as homodimers with a two-fold rotational symmetry. Thus, their binding sites are organized as inverted repeats on the two DNA strands. The VDR, TR, and RAR receptors bind DNA as heterodimers in which one subunit is always the RXR TF. The DNA sequence elements recognized by these heterodimers is a AGGTCA direct repeat. The spacing of the repeats is important in establishing binding specificity.
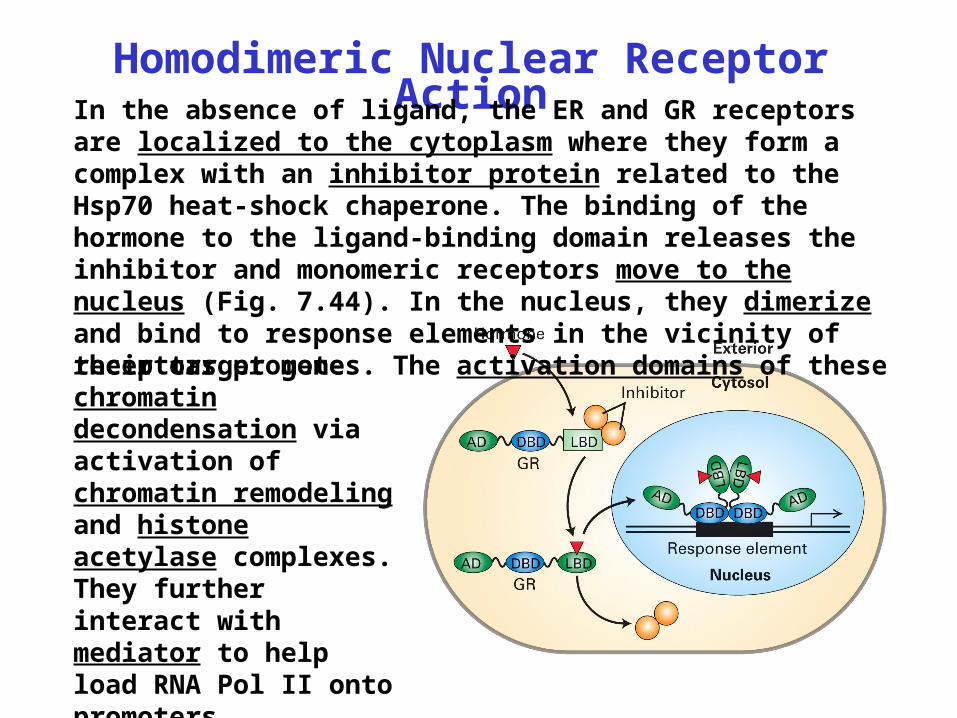
Homodimeric Nuclear Receptor ActionIn the absence of ligand, the ER and GR receptors are
localized to the cytoplasm where they form a complex with an inhibitor protein related to the Hsp70 heat-shock chaperone. The binding of the hormone to the ligand-binding domain releases the inhibitor and monomeric receptors move to the nucleus (Fig. 7.44). In the nucleus, they dimerize and bind to response elements in the vicinity of their target genes. The activation domains of thesereceptors promote chromatin decondensation via activation of chromatin remodeling and histone acetylase complexes. They further interact with mediator to help load RNA Pol II onto promoters.

Heterodimeric Nuclear Receptor Action
Heterodimeric nuclear receptors, e.g., RXR-VDR, RXR-TR, and RXR-RAR, are localized exclusively in the nucleus. They bind to DNA in the absence of ligand, and when bound, cause chromatin to condense via activation of histone deacetylases. Upon ligand binding, the ligand-binding domain undergoes a conformational change. The ligand-bound conformation stimulates the activity of histone acetylases and also can bind mediator, stimulating pre-initiation complex assembly at promoters.







![AP- ˘ ˇˆ˙ˆ˝˛˚˜ ! Transcriptional Repression of Human ... · Transcriptional Repression of Human Immunodeficiency Virus Type + by AP-. Kenichi IMAI ... LTR ^ ]](https://img.pdfslide.us/doc/110x75/5f0eb7647e708231d44096ea/ap-oe-transcriptional-repression-of-human-transcriptional.jpg)





















