Embed Size (px)
Citation preview

182
Biochimica et Biophysics Acta, 664 (1981) 182-187 @ Elsevier/North-Holland Biomedical Press
BBA 57779
CHANGES IN THE AMOUNT OF LUNG AND AIRWAY PHOSPHATIDYLCHOLINE IN 0.5-12.5-DAY-OLD RABBITS
ALAN JOBE, MACHIKO IKEGAMI and HARRIS JACOBS
Fetal-Maternal Research Laboratories, Department of Pediatrics, Harbor-UCLA Medical Center, Torrance, CA 90509 (U.S.A.)
(Received October 27th, 1980)
Key words: Phosphatidylcholine;Surfactantphosphatidylcholine; Development: (Rabbit
lung)
Summary
Newborn rabbits delivered spontaneously at term and cared for by the mothers were studied from 0.5 to 12.5 days of age. Curves are constructed to describe the changes in weight, lung and alveolar wash phosphatidylcholine and saturated phosphatidylcholine, and lung protein. The curves are complex and non-linear. However, expressing the increases in lung and alveolar wash phosphatidylcholine and saturated phosphatidylcholine pool sizes relative to animals weight results in a decreasing linear relationship from 0.5 to 12.5 days of age. By 12.5 days the ratios of lung phosphatidylcholine and saturated phos- phatidylcholine to weight approximate the ratios measured for adult rabbits. The ratios of saturated to total phosphatidylcholine in the alveolar washes and lungs remained invarient throughout the study period.
Introduction
Fetal animals progressively accumulate surfactant phospholipids in lung tissue during the last 20% of term gestation [l]. The surfactant normally appears in the airways just before birth. Following birth and with the clearance of the fetal lung fluid, the amount of phospholipid recovered by lavage of new- born rabbit lungs increases lo-fold within the first 24 h of life [2]. The initial increase in surfactant in the airways is assumed to come from the surfactant stored in lamellar bodies of fetal lung type II pneumocytes. Radioactively labeled phospholipid precursors were used to pulse label newborn rabbit lung phosphatidylcholine and to study the labeling of lamellar body and alveolar wash fractions [ 31. These studies suggested that the surfactant phospholipids of

183
newborn rabbit were secreted and turned over more slowly than in adult rab- bits. However, changes in phospholipid pool sizes with growth in young rabbits need to be measured to better define how the lung grows in the immediate newborn period.
Materials and Methods
Animals. New Zealand White rabbits delivered, and the litters were cared for by the mothers. From approx. 12 h after birth to 12 days of age, one or two newborn rabbits were removed from each of 3-5 litters at 24-h intervals for study. Five rabbits were taken each day from healthy litters. Each animal was weighed and killed with intraperitoneal pentobarbital [3]. The aorta was severed and the chest and neck opened. An appropriately sized cannula was tied into the trachea, and the lungs were lavaged with physiologic saline at 20°C by fully inflating the lung as assessed visually. Six full inflation volumes were infused and withdrawn three times each, and the total recovered lavage volume was pooled. Lungs were removed and a 20% (w/v) homogenate in physiologic saline was prepared [ 31. Duplicates of each lavage and homogenate sample were taken for measurement of phosphatidylcholine, saturated phosphatidylcholine and protein contents.
The lungs of five adult rabbits weighing 1415 ? 36 g were processed similarly.
Phospholipid measurements. All samples were extracted twice according to the method of Bligh and Dyer [4] and the lipid phases pooled and dried under nitrogen. [ 14C] Dipalmitoyl phosphatidylcholine (Applied Sciences, State College, PA) was added to each lipid extract. The phospholipids from duplicate aliquots of each lipid extract were separated by thin-layer chromatography with CH,ClJCH,OH/acetic acid/H,0 (65 : 25 : 8 : 4, v/v) [ 31. The phosphati- dylcholine spots were analyzed for radioactivity [3] and phosphate by the method of Bartlett [ 51. Saturated phosphatidylcholine was recovered from the lipid extracts following oxidation with 0s04 and column chromatography on alumina [6]. The resulting solutions of saturated phosphatidylcholine were divided for measurements of radioactivity and phosphate.
Protein assay. Protein was measured according to the method of Lowry et al. [ 71 using bovine serum albumin as a standard.
Data analysis. All phosphatidylcholine and saturated phosphatidylcholine measurements were corrected for losses during analysis of the lipids using the percent recovery of [ 14C]dipalmitoyl phosphatidylcholine. The losses of phos- phatidylcholine were approx. 14% and of saturated phosphatidylcholine were approx. 10%. The phosphatidylcholine separated from other lipids by one- dimensional thin-layer chromatography cochromatographs with phosphatidyl- inositol; therefore, 6% of the lung and 7% of the alveolar wash phosphatidyl- choline represent phosphatidylinositol [8]. The estimated amounts of phos- phatidylinositol were subtracted from the phosphatidylcholine pool sizes. For each time point, measurements in duplicate from five rabbits were used to cal- culate mean +S.E. values. A least-squares regression program was used to derive equations to fit the mean values vs. age as linear, polynomial or exponential
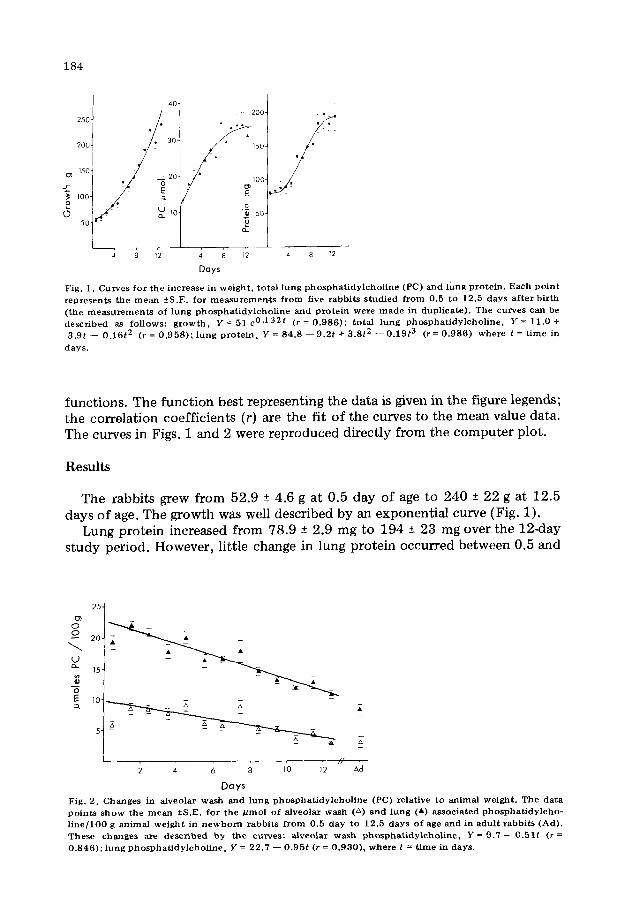
184
Fig. 1. Curves for the increase in weight, total lung phosphatidylcholine (PC) and lung protein. Each point represents the mean fS.E. for measurements from five rabbits studied from 0.5 to 12.5 days after birth (the measurements of lung phosphatidylcholine and protein were made in duplicate). The curves can be described as follows: growth, Y = 51 .0.132f (r = 0.986); total lung phosphatidylcholine. Y = 11.0 + 3.gt - 0.16$ (r = 0.958); lung protein, Y = 84.8 - 9.2t + 3.8t2 - 0.19t3 (r = 0.986) where t = time in
days.
functions. The function best representing the data is given in the figure legends; the correlation coefficients (r) are the fit of the curves to the mean value data. The curves in Figs. 1 and 2 were reproduced directly from the computer plot.
Results
The rabbits grew from 52.9 2 4.6 g at 0.5 day of age to 240 ?r 22 g at 12.5 days of age. The growth was well described by an exponential curve (Fig. 1).
Lung protein increased from 78.9 2 2.9 mg to 194 + 23 mg over the 12-day study period. However, little change in lung protein occurred between 0.5 and
- 2 4 6 8 IO 12 Ad
Days
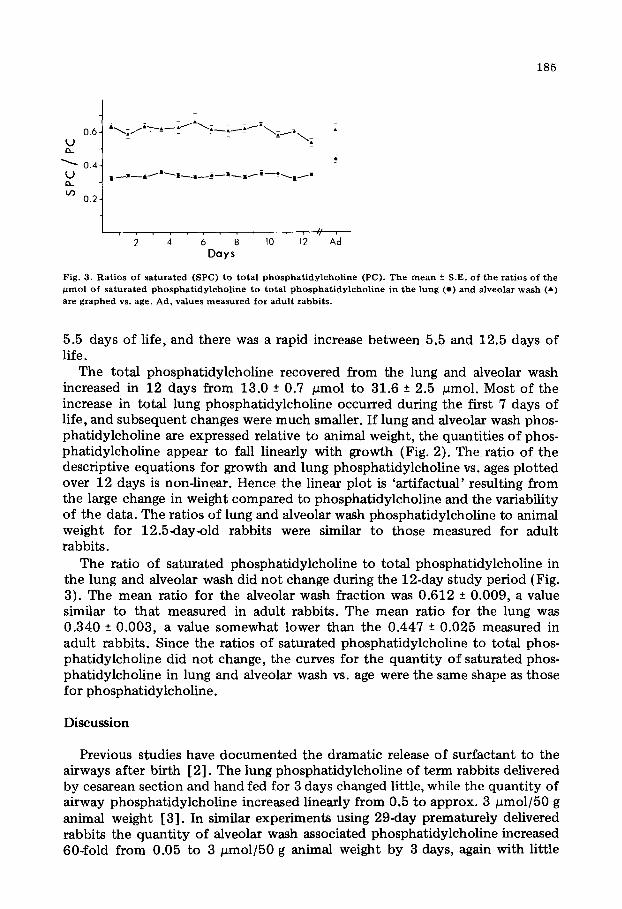
Fig. 2. Changes in alveolar wash and lung phosphatidylcholine (PC) relative to animal weight. The data points show the mean +S.E. for the firno of alveolar wash (A) and lung (A) associated phosphatidylcho- line/100 g animal weight in newborn rabbits from 0.5 day to 12.5 days of age and in adult rabbits (Ad). These changes are described by the curves: alveolar wash phosphatidylcholine, Y= 9.7 - 0.51t (r = 0.846): lung phosphatidylcholine. Y = 22.7 - 0.95t (r = 0.930), where t = time in days.
185
k?
. 0.4- ;
p-=--r’x--E_,__l--‘_x/_ ‘--I,i,*
ln 0.2-
I,,,,,,,,,..T,y, 2 4 6
Days8
IO 12 Ad
Fig. 3. Ratios of saturated (SPC) to total phosphatidylcholine (PC). The mean * SE. of the ratios of the pm01 of saturated phosphatidylcholine to total phosphatidylcholine in the lung (0) and alveolar wash (A) are graphed vs. age. Ad, values measured for adult rabbits.
5.5 days of life, and there was a rapid increase between 5.5 and 12.5 days of life.
The total phosphatidylcholine recovered from the lung and alveolar wash increased in 12 days from 13.0 2 0.7 pmol to 31.6 f 2.5 pmol. Most of the increase in total lung phosphatidylcholine occurred during the first 7 days of life, and subsequent changes were much smaller. If lung and alveolar wash phos- phatidylcholine are expressed relative to animal weight, the quantities of phos- phatidylcholine appear to fall linearly with growth (Fig. 2). The. ratio of the descriptive equations for growth and lung phosphatidylcholine vs. ages plotted over 12 days is non-linear. Hence the linear plot is ‘artifactual’ resulting from the large change in weight compared to phosphatidylcholine and the variability of the data. The ratios of lung and alveolar wash phosphatidylcholine to animal weight for 12.5day-old rabbits were similar to those measured for adult rabbits.
The ratio of saturated phosphatidylcholine to total phosphatidylcholine in the lung and alveolar wash did not change during the la-day study period (Fig. 3). The mean ratio for the alveolar wash fraction was 0.612 + 0.009, a value similar to that measured in adult rabbits. The mean ratio for the lung was 0.340 f 0.003, a value somewhat lower than the 0.447 + 0.025 measured in adult rabbits. Since the ratios of saturated phosphatidylcholine to total phos- phatidylcholine did not change, the curves for the quantity of saturated phos- phatidylcholine in lung and alveolar wash vs. age were the same shape as those for phosphatidylcholine.
Discussion
Previous studies have documented the dramatic release of surfactant to the airways after birth [2]. The lung phosphatidylcholine of term rabbits delivered by cesarean section and hand fed for 3 days changed little, while the quantity of airway phosphatidylcholine increased linearly from 0.5 to approx. 3 pmol/50 g animal weight [ 31. In similar experiments using 29day prematurely delivered rabbits the quantity of alveolar wash associated phosphatidylcholine increased 60-fold from 0.05 to 3 ~01/50 g animal weight by 3 days, again with little

186
change in the amount of lung phosphatidylcholine [ 91. These results in opera- tively delivered rabbits indicate continued entrance of phosphatidylcholine into the airway.
Gross et al. [lo] reported increased amounts of phosphatidylcholine in the alveolar wash of both hand-fed and fasted operatively delivered rabbits over the first 2 days of life. The hand-fed rabbits gained only 2.6 g over 3 days [lo]. The pups in the present study cared for by the mothers gained an average of 27.7 g over the same 3day period. The quantity of phosphatidylcholine recovered from the airways of these rapidly growing healthy newborn rabbits increased appreciably for 4-5 days, and then changed little during the remain- ing days of the study. Growth as assessed by weight gain increased more rapidly than the lung and alveolar wash phosphatidylcholine pools. Thus, 36-h-old rabbits have approx. 2.5-times more lupg and 3-times more airway phosphati- dylcholine relative to weight than do adult rabbits. However, airway and lung phosphatidylcholine relative to animal weight falls in the immediate newborn period to adult values by 12.5 days of age.
The constancy of the ratios of the quantity of saturated to total phosphati- dylcholine in both the lung and alveolar wash suggests in healthy, growing rabbits that the relative metabolism of saturated to total phosphatidylcholine is tightly regulated. The changing ratios typical of late fetal life are no longer apparent [ll]. The kinetics of secretion into the airways of radiolabeled saturated and total phosphatidylcholine studied with either radiolabeled palmitic acid or choline are parallel in both 3day-old and adult rabbits, although the time relations are different at the two ages [8,12]. These labeling studies also support the idea of tight coordination in the metabolism of saturated and total phosphatidylcholine.
Lung protein did not change until after approx. 5 days of age, followed by a rapid increase with continued growth. This pattern of change is clearly differ- ent from the pattern of change of lung phosphatidylcholine. Thus, for example, expressing changes in lung and airway phosphatidylcholine per mg protein may be deceptive. These experiments demonstrate the complex relationship between lung phospholipids, protein and growth in newborn rabbits. They also demonstrate the potential pitfall of assuming that in developing animals a parameter which changes linearly with age when measured with respect to a second parameter shows the same relationship when expressed as an absolute quantity with respect to age.
Acknowledgements
This work was supported by NIH grant HD-11932 and by Research Career Development Award HD-HL-00252 to A.J.
References
1 Clements, J.A. and TO&Y, W. (1977) in Development of the Lung (Hodson, W.A., ed.). pp. 349-
366, Marcel Dekker, New York
2 Roomy. S.A.. Gobran, L.I. and Wai-Lee. T.S. (1977) J. Clin. Invest. 60, 754-759
3 Jobe, A., Kirkpatrick. E. and Gluck. L. (1978) Pediatr. Res. 12, 669475
4 B&b, E.G. and Dyer, W.J. (1959) Can. J. Biochem. Physiol. 37.911-917

187
5 Bartlett. G.R. (1959) J. Biol. Chem. 234,466-468 6 Mason, R.J., Nellenbogen, J. and Clementa, J.A. (1976) J. Lipid Res. 17, 281-284 7 Lowry,P.H.. Rosebrough. N.J., Farr. A.L. and Randall. R.J. (1951) J. Biol. Chem. 193, 265-275 8 Jobe, A. (1980) Pediatr. Res. 14, 319-325
9 Jobe, A. and Gluck. L. (1979) Pediatr. Res. 13.635-640 10 Gross. I.. Wilson, C.M. and Rooney. S.A. (1978) Biochim. Biophys. Acta 528. 190-198 11 Roomy, S.A. and Gobran, L.I. (1977) Lipids 12,1050-1054 12 Jobe, A. (1977) Biochim. Biophys. Acta 489,440-453



![I i pase-cat al y zed hydrolysis phosphatidylcholine of ... · PDF filephosphatidylcholine of guinea pig very low density ... [ ''C]palm~toyl phosphatidylcholine and purified bovine](https://img.pdfslide.us/doc/110x75/5a83b0f17f8b9a682c8ef22c/i-i-pase-cat-al-y-zed-hydrolysis-phosphatidylcholine-of-of-guinea-pig-very-low.jpg)

























