Embed Size (px)
Citation preview

Central Innervation of the Rat Ependymaand Subcommissural Organ With SpecialReference to Ascending SerotoninergicProjections From the Raphe Nuclei
JENS D. MIKKELSEN,1,3* ANDERS HAY-SCHMIDT,2 AND PHILIP J. LARSEN1
1Department of Anatomy, University of Copenhagen, DK-2200 Copenhagen, Denmark2Department of Anatomy, Royal Veterinary andAgricultural University,
Copenhagen, DK-2000 F Denmark3Department of Neurobiology, H. Lundbeck A/S, DK-2500 Copenhagen-Valby, Denmark
ABSTRACTThe subcommissural organ (SCO) and the cerebral ependyma receive serotoninergic
innervation, but little is known about their origin in the raphe nuclei. Application of theretrograde tracer cholera toxin subunit B (ChB) in the third ventricle resulted in uptake inependymal axons and backfilling of perikarya in the dorsomedian part of the dorsal raphenucleus, immediately under the caudal aqueduct. By using dual staining with antiseraagainst serotonin and ChB, a portion of the retrogradely labeled neurons was observed toco-store serotonin. Phaseolus vulgaris–leucoagglutinin (PHA-L) was injected into differentraphe nuclei to fill the neurons in the same areas where the retrogradely labeled neurons werefound. PHA-L injection in the midline of the dorsal raphe nucleus gave rise to ascendingaxonal processes in the mesencephalic central gray, fromwhere they entered the periventricu-lar strata and the third ventricular ependyma. In the cerebral ependyma, large numbers ofpositive fibers were consistently found in the ventral part of the lateral ventricles and in thedorsal part of the third ventricle. A large number of PHA-L-immunoreactive fibers wereobserved in the hypendymal layer of the lateral part of the SCO. Terminal fibers near theependymal cells were also observed. In all cases, the PHA-L injections labeled innervatingfibers both within the ependyma and in the SCO, whereas injections into the median raphenucleus or in other raphe nuclei (i.e., the raphe pallidus and the raphe pontis) labeled fibersneither in the SCO nor in the ependyma. This study shows that a specific group ofpredominantly serotoninergic neurons innervates both the ependyma and the SCO and isprobably involved in cerebrospinal fluid regulation. J. Comp. Neurol. 384:556–568, 1997.r 1997 Wiley-Liss, Inc.
Indexing terms: cerebral ventricles; serotonin; rat; neural tracing
The subcommissural organ (SCO) occupies the dience-phalic roof and covers the posterior commissure, where itforms the lining of the roof of the cerebral aqueduct at itsjunction with the third ventricle. The SCO consists ofspecialized ependymal and ependyma-derived cells of neu-roepithelial origin (for reviews, see Oksche, 1969; Leon-hardt, 1980; Oksche et al., 1993). The SCO consists of twodistinct layers: an ependymal layer of ependymal cells anda hypendymal layer consisting of hypendymal cells, pro-cesses of the ependymal cells, glial cells, nerve fibers, andvascular elements (Oksche, 1969; Oksche et al., 1993).Even though variations among species in the shape andfine structural organization of its ependymal and hypepen-dymal formations have been described, the SCO and the
ependyma are highly constant structures in the vertebratebrain (Oksche et al., 1993). However, its role in brain orneuroendocrine function is almost obscure. The SCO se-cretes glycoproteins into the ventricle (Oksche, 1969;Collins and Woollam, 1979; Sterba et al., 1981; Meiniel et
Grant sponsor: Danish Medical Research Council; Grant number: 12-1642; Grant sponsor: Danish Medical Association; Grant sponsor: LægeEilif og hustru Ane Trier Hansens Legat; Grant sponsor: Ivan NielsensFoundation; Grant sponsor: NOVONordisk Foundation.*Correspondence to: Jens D. Mikkelsen, Department of Neurobiology, H.
LundbeckA/S, Ottiliavej 9, DK-2500 Valby-Copenhagen, Denmark. E-mail:[email protected] 23 May 1996; Revised 3 February 1997; Accepted 5 March 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 384:556–568 (1997)
r 1997 WILEY-LISS, INC.

al., 1988; Rodriguez et al., 1992). The close relationships ofthe secretory cells of the organ with both blood vessels andthe cerebrospinal fluid (CSF), as demonstrated by usingclassical neurosecretory stains or conventional electronmicroscopy, indicate that the SCO is involved in neurohu-moral regulation (Oksche, 1969; Tulsi, 1983; Rodriguez etal., 1984a,b) or in regulation of the flow of CSF (Cifuenteset al., 1994). Support for the latter opinion comes fromindirect evidence that the SCO is lacking or degeneratingin animal models with hydrocephalus (Takeuchi et al.,1987; Irogini et al., 1990).Ultrastructural analysis of the SCO has shown the
presence of synaptic contacts between axonal boutons andboth ependymal and hypendymal cells (Møllgaard andWiklund, 1979; Meiniel et al., 1988), indicating that theorgan is centrally innervated. This input is considered tobe involved in regulation of its secretory function.Histofluo-rescence, immunohistochemical, and autoradiographicstudies have shown that the SCO and the cerebral epen-dyma of some mammalian species contain a large numberof serotoninergic nerve fibers (Chan-Palay, 1976; Bouchaudand Arluison, 1977; Møllgaard and Wiklund, 1979; Takeu-chi and Sano, 1983). Serotonin-accumulating terminalsestablish asymmetrical synaptic contacts with both epen-dymal and hypendymal cells (Bouchaud, 1979; Møllgaardand Wiklund, 1979). These fibers in the SCO disappearafter electrolytic lesions of the dorsal and median raphenuclei of the mesencephalon and pons (Leger et al., 1983),perhaps the same neurons that innervate the ependymallining of the ventricle. The aim of this study was todetermine the precise location of the neurons of the raphesystem projecting to the SCO and to determine to whichdegree other parts of the ependyma were innervated.Attempts to inject retrograde tracer into the ventricular
lumen have been difficult to interpret because it is virtu-ally impossible to determine where in the ependymallining the uptake of tracer takes place. The SCO isdifferent from other circumventricular organs because itpossesses a blood–brain barrier (Krisch, 1993). Neverthe-less, the small size of the SCO in rodents has made the useof in vivo neuronal tracers difficult because substancescannot be delivered iontophoretically into the SCOwithoutlabeling the adjacent structures or the CSF. The sameholds true for tracing from specific parts of the cerebral
ependyma. Furthermore, the close relationship betweenthe SCO, the posterior commissure, and the deep pinealgland makes labeling of passing fibers and leakage oftracer into the blood circulation highly likely. However, toobtain a topography of the projecting neurons, choleratoxin subunit B (ChB) was microinjected into the thirdventricle. This tracer, when injected into the ventricle,should potentially allow labeling of the axons in contactwith the CSF. These experiments produced a large numberof densely labeled neurons in the dorsal raphe nucleus.The interpretational problems related to the topographicalorganization of the ascending projections to the SCO,resulting from the retrograde tracing experiments, wereovercome by the application of the anterograde tracerPhaseolus vulgaris–leucoagglutinin (PHA-L; Gerfen andSawchenko, 1984) into the same areas. This tracer pro-vides a high resolution of labeled axons, a crucial point inthe identification of projections to the circumventricularorgans (see Larsen et al., 1991). Therefore, to determinethe distribution of afferents in the SCO, we injected PHA-Linto several regions within the raphe system to identify thedistribution of projecting fibers in the ependymaand the SCO.
MATERIALS AND METHODS
Ninety adult male Wistar rats (180–250 g), kept understandard conditions with free access to food and water,were used for retrograde (20 animals) and anterograde (70animals) tracing experiments. The animals were exposedto a photoperiod of 12 hours light and 12 hours dark, wereanesthetized 4–6 hours after light onset with tribrometha-nol (0.25 g/kg body weight) dissolved in 9:1 saline:ethanol,and placed in a Kopf stereotaxic apparatus. A glass micro-electrode was broken to give a final tip diameter of 15–20µm and was filled with a solution of 2.5% PHA-L (Vector,Burlingame, CA) or ChB (Sigma, St. Louis, MO), whichwas dialyzed as described previously and applied accord-ing to the protocol of Larsen et al. (1994). For injections inthe raphe nuclei, injections were applied in the midline atdifferent anteroposterior levels, according to the rat brainatlas of Paxinos and Watson (1986). PHA-L and ChB wereapplied iontophoretically (Midgard, current source, Stoelt-ing Inc., Wood Dale, Illinois) by using positive currentpulses of 10 µA (7 seconds on, 7 seconds off) for 15 minutes.ChB was injected microiontophoretically in the midlineinto the third ventricle by using a similar approach.After a survival period of 7 days, the animal was
reanesthetized with tribromethanol and perfused throughthe vascular system for 3 minutes with 100 ml 0.05 Mphosphate buffered saline (PBS; pH 7.4), to which heparin(15,000 IU/l) was added, followed by 400ml 4% paraformal-dehyde dissolved in 0.1 M phosphate buffer (pH 7.4) for 15minutes. The brain was postfixed in the same fixative for4–16 hours and cryoprotected in a 20% sucrose–PBSsolution for at least 2 days at 4°C. Serial frontal cryostatsections of 40 µm thickness were collected in PBS forfree-floating immunohistochemistry.Sections immunoreacted with anti-PHA-L were pro-
cessed as previously described by using a polyclonal rabbitantiserum directed against PHA-L (DAKO, Copenhagen,Denmark) at a dilution of 1:2,000 in PBS containing 0.3%Triton X-100 and 1.0% bovine serum albumin (BSA) (Ger-fen and Sawchenko, 1984; Larsen et al., 1991). Thesections were stained by using the indirect avidin-biotinmethod and diaminobenzidine (DAB) as the chromogen
Abbreviations
3V third ventricle4V forth ventricleAq cerebral aqueductBDHC benzidine dihydrochlorideBSA bovine serum albumineChB cholera toxin, subunit BCG central grayCSF cerebrospinal fluidDAB diaminobenzidineDRn dorsal raphe nucleusDRvm ventromedian part of dorsal raphe nucleusDRdm dorsomedian part of the dorsal raphe nucleusDRl lateral part of the dorsal raphe nucleusDTgP dorsal tegmental nucleus, pericentral partLV lateral ventricleir immunoreactivemlf mediolateral fascicleMR median raphe nucleusPBS phosphate buffered salinePHA-L Phaseolus vulgaris–leucoagglutininSCO subcommissural organxscp decussatio cerebellaris superior
557NEURAL PROJECTIONS TO THE EPENDYMA

(Larsen et al., 1991). Sections to be immunoreacted forChB immunoreactivity were processed by using a poly-clonal goat antiserum directed against ChB (LIST Biologi-cals, Campbell, CA) at a dilution of 1:2,000 in PBScontaining 0.3% Triton X-100 and 1% BSA, as describedelsewhere (Mikkelsen, 1992).After the immunohistochemi-cal reaction, a series of sections was counterstained inthionin (Grubler & Co., Leipzig, Germany) for the demon-stration of cytoarchitecture.
Dual immunohistochemistry
In those cases where application of the ChB was re-stricted to the third ventricle, another series of sectionswas stained for both ChB and serotonin immunoreactivityaccording to the dual-immunohistochemical method previ-ously described (Levey et al., 1986; Larsen and Mikkelsen,1992; Mikkelsen and Larsen, 1993). First, the sectionswere tested for serotonin immunoreactivity by incubatingthe sections in a rabbit antiserumagainst serotonin (Immu-nonuclear, Stillwater) diluted 1:20,000 in PBS containing0.3% Triton X-100 and 1% BSA and treated with theavidin-biotin method as described above by using DAB asthe chromogen. The sections were then washed 6 3 10minutes in PBS and transferred to a solution of ChBantiserum diluted to 1:4,000 in PBS containing 0.3% TritonX-100 and 1% BSA. The immunoreaction was carried out asdescribed above, but the sectionswere stained in a solution ofbenzidine di-hydrochloride (BDHC; Sigma, St Louis, MO)rather than the DAB reaction. Sections prestained forserotonin immunoreactivity were washed several times inPBS, followed by washing for 2 3 10 minutes in 1 Macetate buffer (pH 6.5), before being incubated in freshlyprepared BDHC solution made by dissolving 40 mg BDHC(Sigma) in 90ml distilled water (40°C) for 10minutes. Thissolution was mixed with 10 ml 0.5 M acetate buffer (pH6.5) containing 80 mg sodium nitroprusside and aliquotedinto wells. The sections were placed in wells containing 1ml BDHC solution at room temperature, and the reactionwas started by adding 10 µl 3% H2O2 to each well. After 20minutes, the reaction was stopped by three washes inpostreaction buffer of 0.05 M sodium acetate buffered 0.9%saline (pH 6.5) at 4°C. The sections were washed in PBSbefore being mounted onto gelatin-coated glass slides. Thesections were then dried and coverslipped in Depext.
RESULTS
Retrograde tracing with ChB
Deposits of ChB were iontophoretically injected into thethird ventricle, and animals without labeled cell bodieswithin the periventricular strata were used for morecareful examination. In other cases, small deposits wereobserved in the periventricular zone of the hypothalamus,with leaks into the CSF.When ChB was injected into the ventricular system,
several characteristic features were observed. First, tracerwas taken up by the ependyma of the third ventricle, thelateral ventricles, and the cerebral aqueduct (Fig. 1A–D).Second, tanycytes were densely labeled, and their pro-cesses extended toward the external layer of the medianeminence (Fig. 2). Third, densely labeled neurons in a specificsubset of raphe neurons were identified (Fig. 3A–D).If the injection site involved the ependyma, the periven-
tricular area, or both, two other characteristic featureswere observed. First, retrogradely labeled neurons in the
medial amygdala were found, which were not detected ifthe tracer was delivered into the CSF. Second, if theinjection accidentally involved the ependyma, labeled neu-rons were found throughout the dorsal raphe nucleus andnot only within the distinct subregion of the nucleus.The uptake of ChB was heterogenously distributed in
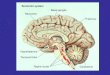
the ependyma. In the lateral ventricles, labeled intraepen-dymal processes were observed in all parts, although thedensest accumulation was observed in the ventral part ofthe lateral ventricles (Fig. 1A). The ependymal lining ofthe lateral septum and the medial striatum exhibitedmoderate labeling, whereas the ependyma under the cor-pus callosum was lightly labeled and contained only a fewimmunoreactive nerve fibers. Around the intraventricularforamen, an extensive network of ChB-immunoreactive(ir) nerve fibers was consistently found (Fig. 1B). Inaddition, in the network of fibers present in the ependymaof the third ventricle, some ependymal cells were alsolabeled (Fig. 1C). In the rostral part of the third ventricle, anetwork of intraependymal fibers was found (Fig. 1C–D),together with a population of ependymal cells. The numberof labeled intraependymal fibers and ependymocytes wasconsistently lower in the mediobasal hypothalamus thanin the rostral hypothalamus. The level of uptake in theependyma differed from case to case, even though theinjections were performed under identical conditions.In the caudoventral third ventricle, the uptake of tracer
was very dense. This part of the ependyma was character-ized by a dense homogeneous uptake of tracer in the epen-dyma and by many densely labeled smooth processes into theventral hypothalamus originating from labeled tanycytes (Fig.2). These processes, which were mostly smooth and possessedno varicosities, coursed through the arcuate nucleus to-ward the marginal zone of the median eminence.The ependyma covering the medial habenular nucleus,
the caudal third ventricle, and the aqueduct were denselylabeled.Distribution of ChB-ir cell bodies in the raphe sys-
tem. The labeled neurons of the raphe nuclei were de-tected in distinct subnuclei within the midline dorsalraphe subnuclei (Fig. 3). The cases used for analysis of thedistribution of raphe neurons satisfied the criteria forlabeling only in the ventricular lumen, and they allrevealed a similar distribution of backfilled neurons, al-though their numbers were slightly variable. The denselylabeled neurons were observed under the caudalmost partof the aqueduct and the rostral part of the fourth ventricle(Figs. 3–4). Theneuronswere found in the caudal dorsal raphenucleus at the level of the superior cerebellar decussatio. Fewneurons were observed in the median raphe nucleus.The cell bodies were 15–20 µm in diameter, usually with
two filled smooth processes (Fig. 4A–D). The neurons inthe dorsal raphe were round, ovoid, or bipolar in shape.The neurons in the ventromedian part of the dorsal raphenucleus (DRn) were round and smaller, often exhibitingprocesses toward the ependyma. Because this ependymawas filled with immunoreactive material, it was not pos-sible to determine whether the processes ended in theependyma or in the periventricular strata.The ChB-ir neurons were concentrated in the dorsome-
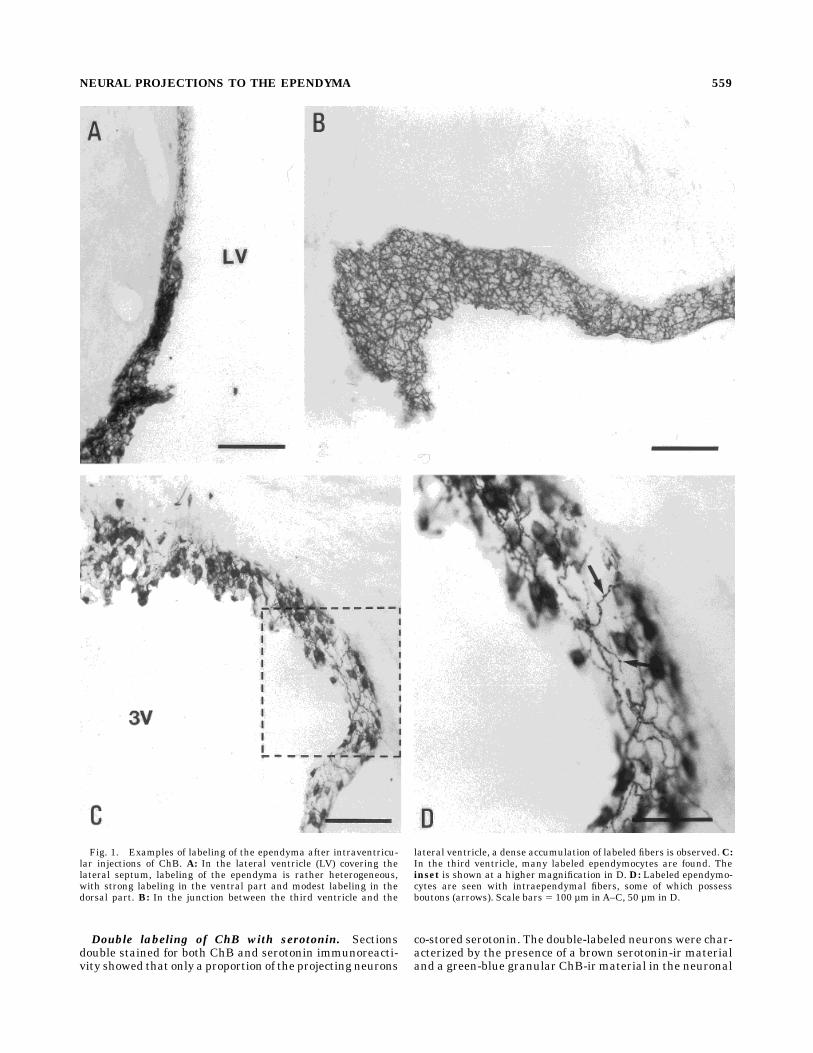
dial and the ventromedian dorsal raphe subnuclei (Fig.5A–C). No labeling was seen in neurons in the lateralsubportion of the DRn. The majority of the neurons wasfound in the dorsomedian part.
J.D. MIKKELSEN ET AL.558
Double labeling of ChB with serotonin. Sectionsdouble stained for both ChB and serotonin immunoreacti-vity showed that only a proportion of the projecting neurons
co-stored serotonin. The double-labeled neurons were char-acterized by the presence of a brown serotonin-ir materialand a green-blue granular ChB-ir material in the neuronal
Fig. 1. Examples of labeling of the ependyma after intraventricu-lar injections of ChB. A: In the lateral ventricle (LV) covering thelateral septum, labeling of the ependyma is rather heterogeneous,with strong labeling in the ventral part and modest labeling in thedorsal part. B: In the junction between the third ventricle and the
lateral ventricle, a dense accumulation of labeled fibers is observed.C:In the third ventricle, many labeled ependymocytes are found. Theinset is shown at a higher magnification in D. D: Labeled ependymo-cytes are seen with intraependymal fibers, some of which possessboutons (arrows). Scale bars 5 100 µm inA–C, 50 µm in D.
559NEURAL PROJECTIONS TO THE EPENDYMA

cytoplasm (Fig. 5D–G). The distribution and numbers ofChB-labeled cells identified by using BDHC as the chromo-gen were virtually identical with that identified withsimilar sections stained with DAB alone. Double-labeledcells were detected only in the dorsomedian and ventrome-dian subnuclei of the DRn. The number of ChB-labeledneurons in the median raphe nucleus were much too low toallow for a careful estimation of the degree of colocaliza-tion with serotonin.The ChB-labeled cells were observed exclusively inside
the raphe nuclei as defined by the distribution of serotonin-containing neurons. Often, ChB-labeled cells were ob-served not to co-store serotonin (Fig. 5D–G). Occasionally,due to the level of diffuse ChB staining of the plexus ofprocesses in the ependymal area of the dorsomediansubnucleus, detection of colocalization in certain neuronsin this region was not possible. However, the number ofdouble-labeled neurons were in the range of 50% of thetotal number of ChB-ir neurons in both the ventromedianand the dorsomedian subnuclei.
Retrogradely labeled neurons outside the raphe nu-
clei. Interestingly, the retrogradely labeled neurons de-tected outside the raphe system were relatively few andwere found exclusively in the diencephalon. Among ani-mals injected in the third ventricle, cells were distributedin the hypothalamic periventricular nucleus (Fig. 6A), themedial part of the hypothalamic paraventricular nucleus(Fig. 6B), the suprachiasmatic nucleus, the magnocellulartuberomammillary nucleus, and the lateral hypothalamicarea (not shown). The neurons in the periventricularhypothalamus were densely labeled, and the immunoreac-tivity was distributed homogeneously in their cytoplasm.Single, long, smooth dendrites of these neurons wereobserved to course both toward the ependyma and deeperinto the hypothalamus (Fig. 6A–B). In the lateral hypotha-lamic area, multipolar and bipolar neurons were observedmostly in a region around the fornix. Outside the hypothala-mus, single, lightly stained neurons were occasionallyobserved in the intergeniculate leaflet of the thalamus(Fig. 6C).
Anterograde tracing with PHA-L
The ascending projection from the raphe nuclei to theSCO and the ependyma was studied with PHA-L tracing.Injections of PHA-L were centered in different areas of theraphe nuclei, but only deposits in the midline DRn gaverise to PHA-L-labeled nerve fibers in the SCO and theependyma (Fig. 7). More specifically, the caudal mesence-phalic and pontine part of the DRn, where ChB-ir neuronswere observed, were the only cases giving rise to innerva-tion of the ependyma. Notably, deposits of tracer in thisarea presumably also leaked PHA-L into the ventricularsystem. Injections into the dorsolateral part of the DRn atthe same level as the injections into the rostral part of theDRn, the median raphe nucleus, or the raphe magnusproduced no labeling of fibers in the ependyma. Tracerdeposits delivered directly into the ventricular lumen didnot give rise to PHA-L-labeled fibers in the brain.Ascending pathways. The PHA-L-ir nerve fibers were
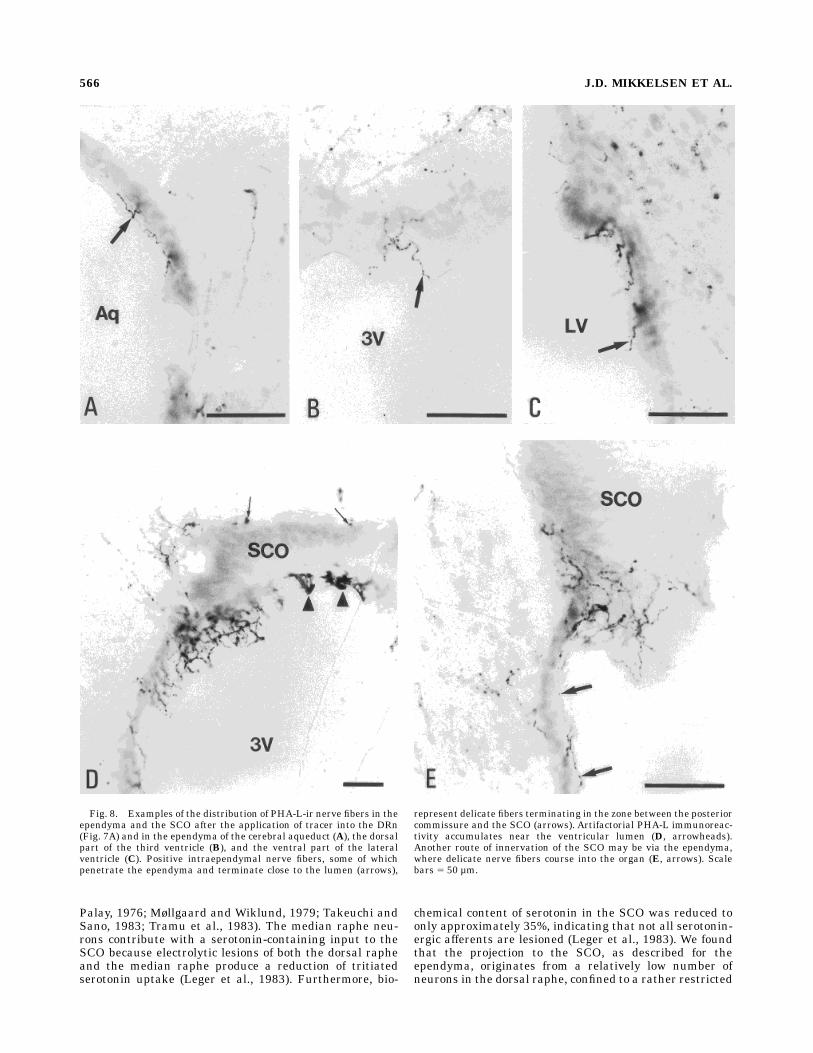
found in the ependyma covering the mesencephalic aque-duct and in the periventricular strata of the region (Fig.8A). Most ascending fibers accumulated in the dorsal partof the periaqueductal gray. The dense network of fibersfound in the dorsal part of the aqueductal ependyma couldbe followed directly to the SCO.Fibers were detected throughout the ependyma of the
third ventricle, except in the ventral part of the ventriclecovering the arcuate nucleus. In the lateral ventricleependyma, a number of positive fibers were seen. Thefibers in the third ventricle ependyma were usually deli-cate and endowed withmany boutons en passage (Fig. 8C).Often, the PHA-L nerve fibers penetrated the ependymaand terminated in close relation to the ventricular lumen(Fig. 8B–D). A number of PHA-L fibers were distributed inthe ventral part of the lateral ventricles (Fig. 8C), whereasthe ependyma covering the corpus callosum was lessdensely innervated.The same injections occasionally gave rise to positive
nerve fibers in the plexus choroideus (not shown).
SCO
A dense termination of PHA-L nerve fibers were ob-served in the SCO (Fig. 8D–E). The fibers entered the SCOfrom both the ependyma (Fig. 8E) and the posteriorcommissure (Fig. 8D). The group of fibers coursing either
Fig. 2. After intraventricular injection of ChB, a dense accumula-tion of tanycytes in the ventral part of the third ventricle (3V) near thearcuate nucleus is observed. Several smooth fibers course from theventricularwall toward themedian eminence (ME). Scale bar5 100 µm.
J.D. MIKKELSEN ET AL.560

along the posterior commissure or from the precommis-sural nucleus into the SCO mostly innervated the lateralpart of the SCO. Single fibers and boutons were also incontact with the proximal part of the central hypendymallayer of the SCO (Fig. 8E). A dense plexus of fibers withextensive arborization and termination was detected inthe lateral part of the SCO. No fibers were seen in theependymal cell layer. A relatively large number of termi-nals also were observed in the ependyma near the SCO,
and in some cases, the same fibers were innervated boththe ependyma and the SCO (Fig. 8E).In the central part of the SCO, where the organ contacts
the CSF, an accumulation of nonspecific immunoreactivity,probably representing binding to SCO glycoproteins (Debbageet al., 1993), was observed in animals injected with PHA-L(Fig. 8D). This accumulation alsowas consistently observed incases inwhich PHA-L injectionswere not centered in theDRnand thus were considered as an unspecific cross reaction.
Fig. 3. Schematic illustration showing the distribution of positive neurons in the dorsal raphe nucleiafter an iontophoretic injection of ChB in the third ventricle.
561NEURAL PROJECTIONS TO THE EPENDYMA

Fig. 4. A–C: Distribution of ChB-ir neurons in the raphe. Theneurons are densely accumulated in the dorsal raphe nucleus, underthe ependyma of the aqueduct (Aq). There is a dense plexus of smoothdendritic processes between the ependyma and the cell bodies.Agroup
of neurons is located ventrally in the medial subnucleus of the ventralpart of the dorsal raphe (arrows, A). At a higher magnification, largedensely labeled neurons with smooth processes (arrows) are observed(D). Scale bars 5 100 µm inA–C, 50 µm in D.
J.D. MIKKELSEN ET AL.562
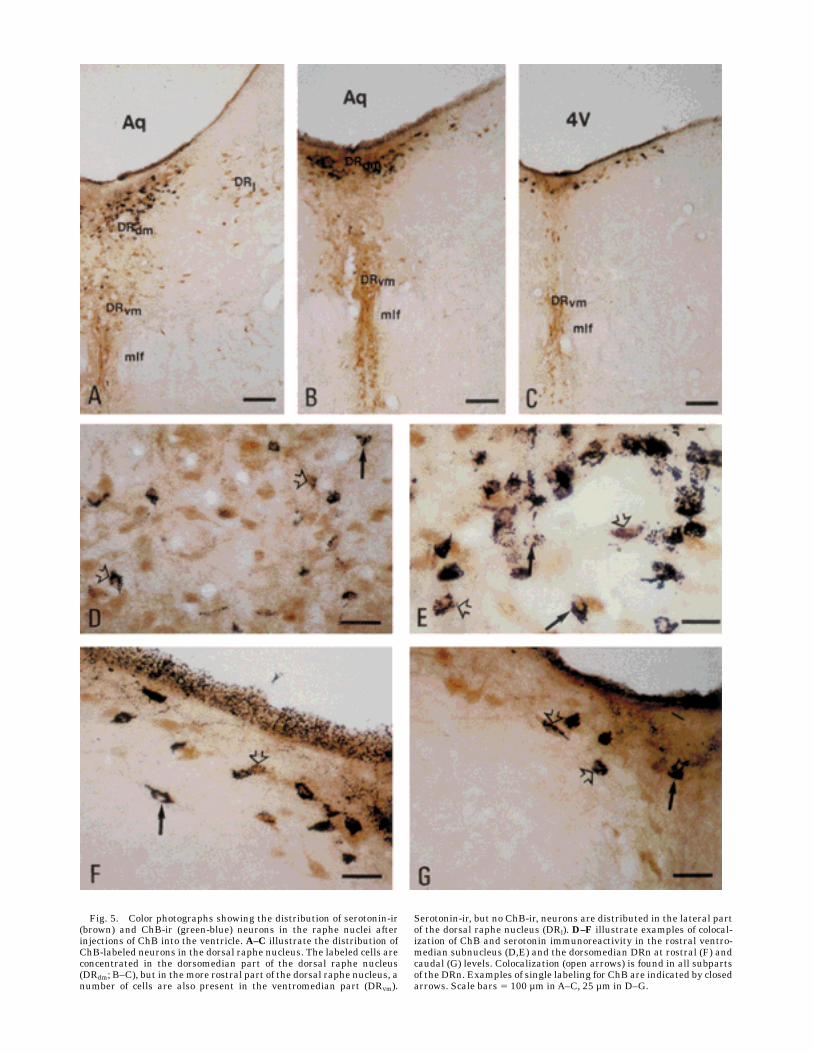
Fig. 5. Color photographs showing the distribution of serotonin-ir(brown) and ChB-ir (green-blue) neurons in the raphe nuclei afterinjections of ChB into the ventricle. A–C illustrate the distribution ofChB-labeled neurons in the dorsal raphe nucleus. The labeled cells areconcentrated in the dorsomedian part of the dorsal raphe nucleus(DRdm; B–C), but in themore rostral part of the dorsal raphe nucleus, anumber of cells are also present in the ventromedian part (DRvm).
Serotonin-ir, but no ChB-ir, neurons are distributed in the lateral partof the dorsal raphe nucleus (DRl). D–F illustrate examples of colocal-ization of ChB and serotonin immunoreactivity in the rostral ventro-median subnucleus (D,E) and the dorsomedian DRn at rostral (F) andcaudal (G) levels. Colocalization (open arrows) is found in all subpartsof the DRn. Examples of single labeling for ChB are indicated by closedarrows. Scale bars 5 100 µm inA–C, 25 µm in D–G.

DISCUSSION
The intraependymal nerve fiber plexus has been knownfor more than two decades (Vigh-Teichmann, 1974). Vigh-Teichmann (1974) described the mammalian infundibularfloor as containing (a) fine-calibered axonlike fibers synaps-ing with the ventricular surface of the ependymal cells, (b)nerve cells located intraependymally and subependymallyprotruding into the ventricular lumen, (c) large smoothintraventricular nerve fibers resembling dendrites contact-ing the CSF, and (d) large-calibered fibers with electron-dense granular vesicles. We injected ChB into the thirdventricle to produce the densest labeling of the sameportion of the ventricular system and the SCO as possible.The study revealed an overall extensive uptake of ChB inthe ependyma lining the third ventricle, especially bytanycytes adjacent to the hypothalamic arcuate nucleus.The concentration of tracer may have been higher in thethird ventricle than in the lateral ventricles. Conse-quently, the uptake by fibers in the ependyma of the lateralventricle is perhaps underscored. However, uptake tookplace only at those locations of the ventricle where PHA-L-positive fibers also were observed after injections into theraphe system. For example, dense termination was ob-served in the ventral part of the lateral ventricles, in thethird ventricle ependyma including the SCO, and in theaqueduct, areas where a dense accumulation of ChB wasfound after injections into the third ventricle. We assumethat the differences in the concentration is not sufficient toexplain the differences in the uptake of retrograde tracerin the third ventricle versus other cerebrospinal lumina.By far, the majority of ChB-labeled cell bodies were seen inthe raphe neurons, ependymal cells, or tanycytes. How-ever, a small number of positive cell bodies were observedin the medial hypothalamus, morphologically resemblingthose described as CSF-contacting neurons (Vigh-Teich-mann, 1974). Their processes were smooth, probably den-drites protruding into the ventricular lumen, which makethem good candidates as contributors to the supraependy-
mal plexuses. Some fibers with boutons resembled axons,which more likely originated from the raphe.With anterograde and retrograde tracing techniques,
the present study demonstrates that the ventricular epen-dyma, including the SCO, receives a direct neuronalinnervation from a distinct and specific subpart of midlinedorsal raphe neurons. The raphe system is a large neuro-nal system with extensive projections to virtually all partsof the central nervous system, and the major transmitterin this system is serotonin (Steinbusch, 1981; Jacobs andAzmitia, 1992). Retrograde tracing studies have revealedmajor widespread projections to a number of forebrainstructures, such as the lateral hypothalamic area aroundthe medial forebrain bundle (Takagi et al., 1980), differentnuclei of the thalamus (Cornwall and Phillipson, 1988;Groenewegen, 1988), striatum (van der Kooy and Hattori,1980a,b; Veening et al., 1980), hippocampus (Imai et al.,1986), and hypothalamus (Sawchenko et al., 1983; Larsenet al., 1996).Accordingly, serotonin-ir nerve fibers originat-ing from the raphe nuclei are found throughout the brain(Steinbusch, 1981; Jacobs andAzmitia, 1992).A number of studies have shown the presence of sero-
toninergic fibers in supraependymally located plexusesthroughout the ventricular system (Lorez and Richards,1973, 1975; Richards and Tranzer, 1974; Chan-Palay,1976; Møllgaard and Wiklund, 1979; Steinbusch, 1981;Takeuchi and Sano, 1983). Serotoninergic fibers are mostnumerous in the dorsal portion of the third ventricle,which is characterized by its lining of ependymycytes thatpossess long cilia on the luminal surface (Bruni et al.,1972). In contrast, the ventral portion of the third ven-tricle, which is characterized by its lining of nonciliatedependymal cells (Brightman et al., 1975), receives little orno serotoninergic innervation. Considering the imperme-able nature of the nonciliated ependyma having individualcells with tight junctions, it may be of functional signifi-cance that only readily permeable sites of the ependymallining receives serotoninergic innervation.
Fig. 6. Labeled neurons outside the raphe nucleus. Single ChB-ir neurons are found in theperiventricular area of the hypothalamus (A), the medial paraventricular nucleus of the hypothalamus(B), and the intergeniculate leaflet of the thalamus (C). Long processes coursing toward the ependyma areindicated by arrows. Scale bars 5 50 µm.
J.D. MIKKELSEN ET AL.564
The raphe system is topographically organized withprojections from the rostral raphe to the telencephalon andthe caudal raphe to the spinal cord and lower brainstem(Vertes, 1991). Except for a few neurons in the medianraphe nucleus, the neurons within the raphe system givingrise to ependymal axons were located in a very specificsubset of DRn, corresponding to the subependymal part ofthe dorsomedian dorsal raphe nucleus and the ventrome-dian dorsal raphe nucleus. These neurons appear to consti-tute a specific population of raphe neurons. Their locationin the midline raphe, their close relationship to theventricle by extensive dendritic arborization in the overly-ing ependyma, and their projections throughout the epen-dyma suggest that they are involved in the regulation ofCSF flow and/or homeostasis. This notion is further sup-ported by the direct connection between DRn and thechoroid plexus. The study of Chan-Palay (1976), however,has emphasized that the serotoninergic fibers do not formclassic synaptic contacts with ependymal cells, suggestingthat they release their putative neurotransmitters, includ-ing serotonin, into the CSF. Taken together, the rapheneurons may play a receptive role in CSF flow andcomposition, and their direct neuronal projection back tothe ependyma and the SCO is a direct feedback loop.Little is known about the functional role of serotonin in
the ependyma, but serotonin is probably the only transmit-ter of the ependyma found in such a high concentrationand density (Takeuchi and Sano, 1983). In the centralnervous system, serotonin is considered to play a modula-
tory role in neurotransmission (Jacobs andAzmitia, 1992).The PHA-L-labeled fibers in the ependyma form a denseplexus among the ependymocytes, and the role of serotoninmay be different in terms of a transmitter in neuron–gliainteraction, as suggested for the SCO.In many cases, PHA-L fibers and axonal terminals were
found in relation to the ventricular lumen. Accordingly,ChB is taken up by ependymocytes, specialized tanycytes,and the axons penetrating the ependyma. Thus, if seroto-nin or other transmitters contained in these axons areactively released to the CSF, they should play a modula-tory role throughout the neuraxis.The relatively low number of cell bodies projecting to the
entire ependyma indicate that these neurons constitute ahighly specialized group of neurons giving rise to anextensive collateralization. The ascending pathways inner-vating the ependyma are rather diffuse.Adistinct networkof ascending fibers is apparently present in the ependyma,whereas innervation via classical ascending pathways, i.e.,themedial forebrain bundle, ismore unlikely. Thewidespreadinnervation from the injection sites in the DRn suggests thatthese neurons are implicated in divergent functions. It wouldbe interesting to knowwhether the same cells projecting to theependyma also project to other forebrain structures.
Innervation of the SCO
Innervation of the SCO by serotoninergic nerve fibers iswell established (Aghajanian and Gallager, 1975; Chan-
Fig. 7. Two representative PHA-L injections into the dorsal raphenucleus. A: An injection into the midline under the aqueduct (Aq),where ChB-positive cells also are observed. This injection extensively
stains the SCO and the ependyma. B: In contrast, PHA-L-positivecells reside slightly laterally to the midline and to the ependyma, andno positive fibers are observed. Scale bar 5 100 µm.
565NEURAL PROJECTIONS TO THE EPENDYMA
Palay, 1976; Møllgaard and Wiklund, 1979; Takeuchi andSano, 1983; Tramu et al., 1983). The median raphe neu-rons contribute with a serotonin-containing input to theSCO because electrolytic lesions of both the dorsal rapheand the median raphe produce a reduction of tritiatedserotonin uptake (Leger et al., 1983). Furthermore, bio-
chemical content of serotonin in the SCO was reduced toonly approximately 35%, indicating that not all serotonin-ergic afferents are lesioned (Leger et al., 1983). We foundthat the projection to the SCO, as described for theependyma, originates from a relatively low number ofneurons in the dorsal raphe, confined to a rather restricted
Fig. 8. Examples of the distribution of PHA-L-ir nerve fibers in theependyma and the SCO after the application of tracer into the DRn(Fig. 7A) and in the ependyma of the cerebral aqueduct (A), the dorsalpart of the third ventricle (B), and the ventral part of the lateralventricle (C). Positive intraependymal nerve fibers, some of whichpenetrate the ependyma and terminate close to the lumen (arrows),
represent delicate fibers terminating in the zone between the posteriorcommissure and the SCO (arrows). Artifactorial PHA-L immunoreac-tivity accumulates near the ventricular lumen (D, arrowheads).Another route of innervation of the SCO may be via the ependyma,where delicate nerve fibers course into the organ (E, arrows). Scalebars 5 50 µm.
J.D. MIKKELSEN ET AL.566

area. The ascending projections from these cells extendedwithin or along the ependyma of the aqueduct, mostly inthe dorsal part of the aqueduct, directly into the region ofthe SCO. Electrolytic lesions of the median raphe nucleusmay affect the overlying dorsal raphe neurons and electro-lytic lesions, probably also involving the ependyma of theaqueduct, where the majority of the ascending projectionsto the SCO is located. Except for the serotoninergicafferents, little is known about the origin of SCO innerva-tion. A serotoninergic innervation has not been shown inall species examined, such as the rabbit (Møllgård, 1979),so it is uncertain as to whether afferents from the rapheexist in this species. It would be interesting to knowwhether there are nonserotoninergic afferents originatingfrom the raphe cells in the rabbit and in other species andwhether this input subserves any functional role. Themajor population of projecting neurons was located in themediodorsal part of the caudalmost dorsal raphe nucleus,extending rather extensively in the rostrocaudal extent,and this small rim of cells contained both serotoninergicand nonserotoninergic neurons.The function of the SCO is considered to be related to
neurohumoral processes (Oksche et al., 1993). The un-known secretory product of the SCO is secerned to theCSF, and serotonin inhibits this secretory processes. Otherresults have shown that the serotoninergic afferents,which do not contain gamma-aminobutyric acid (GABA),innervates the SCO ependymocytes (Voutsinos et al.,1994). These afferents likely originate from the neurons inthe DRn described in the present study, although otherpossibilities exist (Mikkelsen, 1994; present study). Graftedmidbrain neurons projecting to the adult SCO identified byserotonin uptake innervates the glia cells (Voutsinos et al.,1994), indicating that theremay be a neuronal–glia connec-tion in the SCO. The fact that the ependymocytes and thespecialized SCO ependymocytes are modified glia cells andhave direct innervation (Møllgaard and Wiklund, 1979)suggests specific relationships between cells of neural andglial origin. Because the serotoninergic cells are born early,they are perhaps involved in the development of glia cells(Lieth et al., 1990; Voutsinos et al., 1994).Considering the dense serotoninergic innervation of the
ependyma, it may be surprising that a substantial numberof fibers originating from themidbrain are nonserotoniner-gic, although a subportion of neurons belonging to theraphe system are nonserotoninergic. Colocalization stud-ies have shown that GABAergic neurons are present in theraphe nuclei (Belin et al., 1983), but it remains to bedetermined whether some of the neurons projecting to theSCO and the ependyma are GABAergic. However, supra-ependymal fibers contain both serotonin and GABA (Har-randi et al., 1986). Notably, serotonin is co-stored andcodistributed in the caudal raphe nuclei with substance P,thyrotropin releasing hormone, and galanin (Johnson etal., 1993). These peptides are located mainly in the rapheobscurus, and none of these peptides have been found inthe SCO or in the ependyma.
ACKNOWLEDGMENTS
We thank Gitte Sørensen and Tine Ø. Gerlen for skillfultechnical assistance and Grazyna Hahn for assistance inthe preparation of photomicrographs.
LITERATURE CITED
Aghajanian, G.K., and D.W. Gallager (1975) Raphe origin of serotonergicnerves terminating in the cerebral ventricles. Brain Res. 88:221–231.
Belin, M.F., D.W. Nanopoulos, M. Didier, M. Aguera, H. Steinbusch, A.Verhofstad, M. Maitre, and J.F. Pujol (1983) Immunohistochemicalevidence for the presence of gamma-aminobutyric acid and serotonin inone nerve cell. A study on the raphe nuclei using antibodies toglutamate decarboxylase and serotonin. Brain Res. 275:329–339.
Bouchaud, C. (1979) Evidence for amultiple innervation of subcommissuralependymocytes in the rat. Neurosci. Lett. 12:253–258.
Bouchaud, C., and M. Arluison (1977) Serotoninergic innervation of theependymal cells in the rat subcommissural organ. A fluorescence andelectron microscopic and autoradiographic study. Biol. Cell 30:61–64.
Brightman,M.W., L. Prescott, and T.S. Reese (1975) The ventricular systemin neuroendocrine mechanisms. In K.M. Knigge (ed): Brain–EndocrineInteraction. Basel: Karger, pp. 146–165.
Bruni, J.E., D.G. Montemurro, R.E. Cluttenburg, and R.P. Singh (1972)Scanning electron microscopic study of the ependymal surface of thethird ventricle of the rabbit, rat, mouse, and human brain. Anat. Res.174:407–420.
Chan-Palay, V. (1976) Serotonin axons in the supra- and subependymalplexuses and in the leptomeninges; their roles in local alterations ofcerebrospinal fluid and vasomotor activity. Brain Res. 102:103–130.
Cifuentes, M., S. Rodriguez, J. Perez, J.M. Grondona, E.M. Rodriguez, andL.P. Fernandez (1994) Decreased cerebrospinal fluid flow through thecentral canal of the spinal cord of rats immunologically deprived ofReissner’s fibre. Exp. Brain Res. 98:431–440.
Collins, P., and D.H. Woollam (1979) The ventricular surface of thesubcommissural organ: A scanning and transmission electron micro-scopic study. J. Anat. 129:623–631.
Cornwall, J., and O.T. Phillipson (1988) Afferent projections to the parafas-cicular thalamic nuclei of the rat, as shown by retrograde transport ofwheat germ agglutinin. Brain Res. Bull. 20:139–150.
Debbage, P., W. Lehmann, U.K. Hanisch, and W.W. Naumann (1993)Immunological crossreactivities between proteins secreted by the sub-commissural organ, and plant lectins. Acta Histochem. 94:131–140.
Gerfen, C.R., and P.E. Sawchenko (1984) An anterograde neuroanatomicaltracing method that shows the detailed morphology of neurons, axonsand terminals: Immunohistochemical localization of an axonally trans-ported plant lectin, Phaseolus vulgaris leucoagglutinin. Brain Res.209:283–309.
Groenewegen, H.J. (1988) Organization of the afferent connection of themediodorsal thalamic nucleus in the rat, related to the mediodorsal–prefrontal topography. Neuroscience 24:379–431.
Harrandi, M., M. Didier, M. Aguera, A. Calas, and M.F. Belin (1986) GABAand serotonin (5HT) pattern in the supraependymal fibers of the ratepithalamus: Combined radioautographic and immunocytochemicalstudies. Effect of 5HT content on 3H GABA accumulation. Brain Res.370:241–249.
Imai, H., D.A. Steindler, and S.T. Kitai (1986) The organization of divergentaxonal projections from the midbrain raphe nuclei in the rat. J. Comp.Neurol. 243:363–380.
Irogini, C., E.M. Rodriguez, M. Heinrichs, K. Frese, S. Herzog, A. Oksche,and R. Rott (1990) Immunocytochemical study of the subcommissuralorgan of rats with postnatal induced hydrocephalus. Exp. Brain Res.82:384–392.
Jacobs, B.L., and E.C. Azmitia (1992) Structure and function of the brainserotonin system. Physiol. Rev. 72:165–229.
Johnson, H., B. Ulfhake, A. Dagerlind, G.W. Bennett, K.C. Fone, and T.Hokfelt (1993) The serotoninergic bulbospinal system and brainstem-spinal cord content of serotonin, TRH-, and substance P-like immunore-activity in the aged rat with special reference to the spinal cord motornucleus. Synapse 15:63–89.
Krisch, B. (1993) Blood- and cerebrospinal fluid-dominated compartmentsof the rat brain. In A. Oksche, E.M. Rodrıguez, and P. Fernandez-Llebrez (eds): The Subcommissural Organ.An Ependymal Brain Gland.Berlin: Springer-Verlag, pp. 151–161.
Larsen, P.J., and J.D. Mikkelsen (1992) Vasoactive intestinal peptide (VIP)in magnocellular neurons of the hypothalamo-neurohypophysial sys-tem of the mink (Mustela vision) is co-localized with vasopressin oroxytocin. J. Comp. Neurol. 326:180–192.
Larsen, P.J., M.Møller, and J.D.Mikkelsen (1991) Efferent projections fromthe periventricular and medial parvocellular subnuclei of the hypotha-lamic paraventricular nucleus to the circumventricular organs of therat. A Phaseolus vulgaris leucoagglutinin (PHA-L) tracing study. J.Comp. Neurol. 306:463–479.
567NEURAL PROJECTIONS TO THE EPENDYMA

Larsen, P.J., A. Hay-Schmidt, and J.D. Mikkelsen (1994) Efferent connec-tions of the lateral hypothalamic area to the hypothalamic nucleus inthe rat. J. Comp. Neurol. 342:299–319.
Larsen, P.J., A. Hay-Schmidt, N. Vrang, and J.D. Mikkelsen (1996) Originof projections from the midbrain raphe nuclei to the hypothalamicparaventricular nucleus in the rat: A combined retrograde and antero-grade tracing study. Neuroscience 70:963–988.
Leger, L., A. Degueurce, J.J. Lundberg, J.F. Pujol, and K. Møllgard (1983)Origin and influence of the serotoninergic innervation of the subcommis-sural organ in the rat. Neuroscience 10:411–423.
Leonhardt, H. (1980) Ependym und Circumventriculare Organe. In A.Oksche and L. Vollrath (eds): Neuroglia I. Handbuch der Mikrosko-pischen Anatomie des Menschen. Heidelberg: Springer-Verlag, pp.177–665.
Levey, A.I., J.P. Bolam, D.B. Rye, A.E. Hallanger, R.M. Demuth, M.M.Mesulam, and B.H. Wainer (1986) A light and electronmicroscopicprocedure for sequential double staining using diaminobenzidine dihy-drochloride. J. Histochem. Cytochem. 34:1449–1457.
Lieth, E., D.R. McClay, and J.M. Lauder (1990) Neuronal–glia interactions:Complexity of neurite outgrowth correlates with substrate adhevisity ofserotonergic neurons. Glia 3:169–179.
Lorez, H.P., and J.G. Richards (1973) Distribution of indolealkylaminenerve terminals in the ventricles of the rat brain. Z. Zellforsch.144:511–522.
Lorez, H.P., and J.G. Richards (1975) 5-HT nerve terminals in the fourthventricle of the rat brain: Their identification and distribution studiedby fluorescence histochemistry and electron microscopy. Cell TissueRes. 165:37–48.
Meiniel, A., J.L. Molat, and R. Manual (1988) Complex-type glycoproteinssynthesized in the subcommissural organ of mammals. Light- andelectron-microscopic investigations by use of lectins. Cell Tissue Res.253:383–395.
Mikkelsen, J.D. (1992) Visualization of the efferent projections by immuno-histochemical identification of cholera toxin B (CHB). Brain Res. Bull.28:619–623.
Mikkelsen, J.D. (1994) A direct neuronal projection from the lateralgeniculate nucleus to the subcommissural organ of the rat. Cell TissueRes. 277:437–445.
Mikkelsen, J.D., and P.J. Larsen (1993) Substance P in the suprachiasmaticnucleus of the rat: An immunohistochemical and in situ hybridizationstudy. Histochemistry 100:316.
Møllgaard, K., and L. Wiklund (1979) Serotoninergic synapses on ependy-mal and hypendymal cells of the rat subcommissural organ. J. Neurocy-tol. 8:445–467.
Møllgård, K. (1979) Serotoninergic regulation of secretory activity of thesubcommissural organ in some rodents. Thesis, University of Lund,Lund, Sweden.
Oksche, A. (1969) The subcommissural organ. J. Neurovisc. Relat. Suppl.9:111–139.
Oksche, A., E.M. Rodrıguez, and P. Fernandez Llebrez (1993) The Subcom-misural Organ. An Ependymal Brain Gland. New York: Springer, pp.1–333.
Paxinos, G., and C. Watson (1986) The Rat Brain in Stereotaxic Coordi-nates. San Diego: Academic Press.
Richards, J.G., and J.P. Tranzer (1974) Ultrastructural evidence for thelocalization of an indolealkylamine in supra-ependymal nerves fromcombined cytochemistry and pharmacology. Experientia 30:287–289.
Rodriguez, E.M., A. Oksche, S. Hein, S. Rodriguez, and R. Yulis (1984a)Comparative immunocytochemical study of the subcommissural organ.Cell Tissue Res. 237:427–441.
Rodriguez, E.M., A. Oksche, S. Hein, S. Rodriguez, and R. Yulis (1984b)Spatial and structural interrelationships between secretory cells of thesubcommissural organ and blood vessels. An immunocytochemicalstudy. Cell Tissue Res. 237:443–449.
Rodriguez, E.M., A. Oksche, S. Hein, and C.R. Yulis (1992) Cell biology ofthe subcommissural organ. Int. Rev. Cytol. 135:39–121.
Sawchenko, P.E., L.W. Swanson, H.W.M. Steinbusch, andA.A.J. Verhofstad(1983) The distribution and cells of origin of serotonergic inputs to theparaventricular and supraoptic nuclei of the rat. Brain Res. 277:355–360.
Steinbusch, H.W.M. (1981) Distribution of serotonin-immunoreactivity inthe central nervous system of the rat—Cell bodies and terminals.Neuroscience 4:557–618.
Sterba, G., I. Kleim, W. Naumann, and H. Petter (1981) Immunocytochemi-cal investigation of the subcommissural organ in the rat. Cell TissueRes. 218:659–662.
Takagi, H., S. Shiosaki, M. Tohyama, E. Senba, and M. Sakanaka (1980)Ascending components of the medial forebrain bundle from the lowerbrain stem in the rat, with special reference to raphe and catecholaminecell groups. A study by the HRPmethod. Brain Res. 193:315–337.
Takeuchi, I.K., R. Kimura, M. Matsuda, and R. Shoji (1987) Absence ofsubcommissural organ in the cerebral aquaduct of congenital hydro-cephalus spontaneously occuring in MT/HokIdr mice. Acta Neuro-pathol. 73:320–322.
Takeuchi, Y., and Y. Sano (1983) Serotonin distribution in the circumven-tricular organs of the rat. An immunohistochemical study. Anat.Embryol. 167:311–319.
Tramu, G., A. Pillez, and J. Leonardelli (1983) Serotonin axons of theependyma and circumventricular organs in the forebrain of the guineapig. An immunohistochemical study. Cell Tissue Res. 228:297–311.
Tulsi, R.S. (1983) Neural elements associated with the subcommissuralorgan of the brushtailed possum (Trichosurus vulpecula). Cell TissueRes. 234:335–350.
van der Kooy, D., and T. Hattori (1980a) Bilaterally situated dorsal raphecell bodies have only unilateral forebrain projections in the rat. BrainRes. 192:1–5.
van der Kooy, D., and T. Hattori (1980b) Dorsal raphe cells with collateralprojections to caudate-putamen and substantia nigra: A fluorescentretrograde double labeling study in the rat. Brain Res. 186:1–7.
Veening, J. G., F. M. Cornelissen, and P. A. J. M. Lieven (1980) Thetopographical organization of the afferents to the caudatoputamen ofthe rat. Neuroscience 5:1253–1268.
Vertes, R.P. (1991) A PHA-L analysis of ascending projections of the dorsalraphe nucleus in the rat. J. Comp. Neurol. 313:643–668.
Vigh-Teichmann, I. (1974) The infundibular cerebrospinal-fluid-contactingneurons. Adv. Embryol. Cell Biol. 50:7–91.
Voutsinos, B., L. Chouaf, P. Mertens, F.P. Ruiz, Y. Joubert, M.F. Belin, andB.M. Didier (1994) Tropism of serotonergic neurons towards glialtargets in the rat ependyma. Neuroscience 59:663–672.
568 J.D. MIKKELSEN ET AL.





![ReviewArticle - Hindawi Publishing Corporation · PDF file · 2014-03-26ReviewArticle ... overlying the pterygomandibular raphe medially and the ... ramus [8]. This is especially](https://img.pdfslide.us/doc/110x75/5aaac3787f8b9a9a188ea657/reviewarticle-hindawi-publishing-corporation-overlying-the-pterygomandibular.jpg)