Embed Size (px)
Citation preview

1
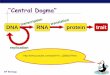
central dogma
tRNA
rRNA
sRNA
mRNA
ribosomal proteins
DNApRNAp
structural proteins
transporters
regulators
enzymesDNA
rpl
tsx tsl
ribosomes
+ regulation
• tsx initiation control by transcription factors (TF)• tsl initiation control by sRNA and RNA-binding proteins • tsx termination control by anti-terminators (eg. protein, sRNA)• control of mRNA and protein degradation
coupled toenvironmentalsignals
1
tsx initiation controlby σ-factors and TFs
tsx elongation
tsx termination controlby proteins and sRNA
tsl initiation controlby proteins and sRNA tsl elongation
mRNA stability controlmRNA stability control
post-tsl control: modification & proteolysis
genetic circuits utilize allthese modes of regulation!
2

2
Topic 3: Post-transcriptional controlA. Transcriptional elongation and termination
1. Basic models of tsx elongation and termination2. mechanisms of termination (intrinsic vs rho-dependent)
B. Control of termination (=anti-termination or AT)1. AT at a single termination site (various mechanisms)2. processive AT (Q, N, Nus)
C. Translational mechanisms (initiation, elongation, termination)D. Translational control
1. RNA-binding protein2. riboswitch3. small regulatory RNA
E. Protein degradation and post-translational control1. proteolytic machinery2. protein unfolding3. substrate selection
3
A. Transcriptional elongation and termination(post-tsx-initiation control)
• “normal” termination at end of an operon• premature termination within or even at the beginning of an operon→ control mechanism (antitermination)
4

3
1. Model of tsx elongation
transcript length NN N+1N-1
backtracked
forward-trackedtra
nscr
iptio
n m
ode m
1
2
0
-1
energetics of tsx elongationΔGN ,m = ΔGN ,m; DNA bubble
+ΔGN ,m; RNA-DNA hybrid + ΔGN ,m; RNAP binding
! bubble prefers A/T-richstretches of DNA(AT: weaker basepair, DNA:DNA stronger than DNA:RNA hybrid)
! forwardtrack: energetically costly! backtrack: energetically neutral
(even favored if tsx error occurs)
5
kinetics of tsx elongation:• elemental pause -- freq: 1 every 10s-- duration: ~1s
• longer pauses-- freq: 1 every 100s-- duration: 1~10s
Adelman et al. 2002
Landick 2006
typ. few-tens sec.
in vitro also arrest
• pauses can be stabilized (and prolonged) by several mechanisms:
6

4
kinetic model:
N N+1
actively transcribing
paused
terminated
kfv
kfvT
⋅+
≈
⋅++
=
ττ00 1
1
)11(1
1
k
f 1/t
v0
Yager & von Hippel 1992
N-1
Termination efficiency
‘normal’ site: v0~100/s, f~0.1/s, t ~1s, k<0.1/s
termination site: v0~f~100/s, t~(1-10) s, k~ 1-5/s
T < 10-4
T~ 0.2-1
7
optimal terminator: • hairpin with GC-rich stem
followed by runs of U’s• 7-9nt separating hairpin and U’s• downstream sequence inductive
to TEC pausing
2. Mechanisms of termination(a) intrinsic termination
E. coli: 50% of mRNA have intrinsic terminators at their end70% of non-coding RNA mutations
disrupting termination
8

5
Model of intrinsic termination
• destabilization of TEC requires simultaneous disruption of HBS and RBS– hairpin formation displaces RNA from UBS (stimulated by NusA, part of TEC)– stretch of U’s provides weakest HBS
• pausing at terminator promotes hairpin formation and is essential for termination (depends on downstream sequence)
[Nudler & Gottesman, 2002]
9
(b) rho-dependent termination
• rho binds to ~40nt stretch ofunstructured, C-rich mRNA ( = RUT)
• translocation of rho requires energy• actual termination site not well-defined
– up to 120nt distal to RUT– generally correlate with pause sites
10

6
• polarity: nonsense mutation affects the expression of downstream genes
• tsx-tsl coupling → no tsx of untranslated mRNA(quality check on translation?)
• similar effect may arise at on-set of starvation (a.a. shortage)or upon exposure to translation-inhibiting antibiotics?
11
• polarity: nonsense mutation affects the expression of downstream genes
• tsx-tsl coupling → no tsx of untranslated mRNA(quality check on translation?)
• similar effect may arise at on-set of starvation (a.a. shortage)or upon exposure to translation-inhibiting antibiotics?
antibiotics or sudden starvation
RNAP
stalled ribosome
RNAP
rho-dependent termination
12

7
Quantitative study of tsx processivityDai, Zhu, TH (Nat Microb 2019)
LacZP O
LacIT0:IPTG
LacZP O RNAp
T1
LacZP O RNAp RNApT2
A B C
A AA
A
timeT1 T2T0
mR
NA
abun
danc
e
BB
B
C
13
0 40 80 120 1600
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P2365 primer
P259 primer
P1540 primer
P860 primer
P1903 primer
P3069 primer
P1199 primer
Glucose (DT: 45 min)
0 50 100 150 2000
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P1903 primer
P3069 primer
P1199 primer
P2365 primer
P259 primer
P1540 primer
P860 primer
Glucose FA 0.25 µg/mL (DT:200 min)
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
P1903 primer
P3069 primerP2365 primer
P259 primer
P1540 primerP860 primer
Glucose FA 0.1µg/mL (DT:100 min)
P1199 primer
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Glucose 8µM Cm (DT:140 min)
P1903 primer
P3063 primerP2365 primer
P259 primer
P1540 primer
P860 primerP1199 primer
A B
C D
waiting time (s)
Quantitative study of tsx processivity
x-intercept: (~10s)tsx init time
slope: tsx speed(47.6 nt/s)
position-dependentwaiting time
slope: d[m]/t
Dai, Zhu, TH (Nat Microb 2019)
14

8
0 40 80 120 1600
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P2365 primer
P259 primer
P1540 primer
P860 primer
P1903 primer
P3069 primer
P1199 primer
Glucose (DT: 45 min)
0 50 100 150 2000
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P1903 primer
P3069 primer
P1199 primer
P2365 primer
P259 primer
P1540 primer
P860 primer
Glucose FA 0.25 µg/mL (DT:200 min)
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
P1903 primer
P3069 primerP2365 primer
P259 primer
P1540 primerP860 primer
Glucose FA 0.1µg/mL (DT:100 min)
P1199 primer
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Glucose 8µM Cm (DT:140 min)
P1903 primer
P3063 primerP2365 primer
P259 primer
P1540 primer
P860 primerP1199 primer
A B
C D
Quantitative study of tsx processivity
0 40 80 120 1600
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P2365 primer
P259 primer
P1540 primer
P860 primer
P1903 primer
P3069 primer
P1199 primer
Glucose (DT: 45 min)
0 50 100 150 2000
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P1903 primer
P3069 primer
P1199 primer
P2365 primer
P259 primer
P1540 primer
P860 primer
Glucose FA 0.25 µg/mL (DT:200 min)
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
P1903 primer
P3069 primerP2365 primer
P259 primer
P1540 primerP860 primer
Glucose FA 0.1µg/mL (DT:100 min)
P1199 primer
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Glucose 8µM Cm (DT:140 min)
P1903 primer
P3063 primerP2365 primer
P259 primer
P1540 primer
P860 primerP1199 primer
A B
C D
0 40 80 120 1600
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P2365 primer
P259 primer
P1540 primer
P860 primer
P1903 primer
P3069 primer
P1199 primer
Glucose (DT: 45 min)
0 50 100 150 2000
200
400
600
800
Induction time (s)
Lac
Z m
RN
A a
bu
nd
ance
P1903 primer
P3069 primer
P1199 primer
P2365 primer
P259 primer
P1540 primer
P860 primer
Glucose FA 0.25 µg/mL (DT:200 min)
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
P1903 primer
P3069 primerP2365 primer
P259 primer
P1540 primerP860 primer
Glucose FA 0.1µg/mL (DT:100 min)
P1199 primer
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Glucose 8µM Cm (DT:140 min)
P1903 primer
P3063 primerP2365 primer
P259 primer
P1540 primer
P860 primerP1199 primer
A B
C D
0 50 100 150 2000
500
1000
Time (s)
Rel
ativ
e m
RN
A le
vel
Glucose Erythromycin 90 (µg/mL) (DT:126 min)
P1903 primerP3069 primerP1199 primerP2365 primerP259 primerP1540 primerP860 primer
Dai, Zhu, TH (Nat Microb 2019)
15
Quantitative study of tsx processivity
0 1000 2000 3000 4000 50000.1250.2500.500
1248
16
Probe location
Slo
pe
Glucose FA 0.1FA 0.25Cm8C-Starvation
Z3' Y5' Y3' A5' A3'
• similar effects for different drugs and starvation• gradual loss of tsx processivity: 50% loss per 1-2 kb• abrupt drop (~4x) at the end of lacZ gene
Dai, Zhu, TH (Nat Microb 2019)
16

9
Quantitative study of tsx processivity
0 60 120 1800
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Glucose 8 µM Cm
P1903 primer
P3063 primerP2365 primer
P259 primer
P1540 primer
P860 primerP1199 primer
0 50 100 150 2000
200
400
600
800
1000
1200
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Nonsense mutation
0 50 100 150 2000
200
400
600
800
1000
Induction time (s)
Rel
ativ
e m
RN
A le
vel
Glucose Ery 90 (µg/mL)
0 800 1600 2400 32000.0
0.2
0.4
0.6
0.8
1.0
Location
Rel
ativ
e sl
op
e
No drugFA 0.3FA 1.2Cm8
Isoleucine starvationNonsense mutation
Ery90
A B
D
C
0 100 200 3000
50
100
150
200
250
Induction time(s)
Rel
ativ
e m
RN
A le
vel
Isoleucine starvation
E
• similar effects for nonsense mutation (no physiological perturbation)• 50% loss per 0.5 kb
0 500 1000 1500 2000 25000.1
0.2
0.4
0.8
1.6
glucose Cm 8stop codon
position
mR
NA
synt
hers
is ra
te
LacZP O
nonsense mutation Dai, Zhu, TH (Nat Microb 2019)
17
0 50 100 1500
200
400
600
800
1000
Induction time(s)
LacZ
mR
NA
abu
ndan
ce
Stop codon-croRP297 primerP636 primerP898+150 primerP1237+150 primerP1578+150 primerP1941�150 primerP2403+150 primer
0 500 1000 1500 2000 25000
5
10
15
Position (nt)
Slo
pe
Stop codon croR terminatorA B
Quantitative study of tsx processivity
LacZP O
nonsense mutationtsx pause site (croR)
! effective termination by rho requires tsx pause site following stop codon! intended function of rho: termination vs quality control
0 50 100 1500
200
400
600
800
1000
Induction time(s)
LacZ
mR
NA
abu
ndan
ce
Stop codon-croRP297 primerP636 primerP898+150 primerP1237+150 primerP1578+150 primerP1941�150 primerP2403+150 primer
0 500 1000 1500 2000 25000
5
10
15
Position (nt)
Slo
pe
Stop codon croR terminatorA B
Dai, Zhu, TH (Nat Microb 2019)
18

10
B. Control of termination: Anti-termination
• ~10% of genes in E. coli are controlled by AT• two types:-- control of a single termination site-- processive AT (controls many terminators)
• many different mechanisms
19
1. AT at a single terminator
positive feedback: increasesthe amplitude of fold-changes
BglG
alternative hairpin stabilizedby RNA-binding protein BglG
usually involves intrinsic terminator
(a) via protein-mRNA interaction (e.g., bgl operon)
alternative structure
20

11
(b) via sRNA-mRNA interaction
time window for sRNA action
anti-terminator
anti anti-terminator
[Gerhard Wagner lab]
with RNAIII
w/o RNAIII
21
(c) via small molecule-RNA interaction (riboswitch)
22

12
(d) via coupling to translation (= translational attenuation)
e.g., trp operon of E.coli (biosynthesis of Trp)
Trp level high: ribosome fast → termination
Trp level low: ribosome stalls → antitermination
23
translational attenuation
24

13
very different implementation of the same ‘idea’ in B. subtilis
trp RNA-binding attenuation proteinbound TRAP stabilizes terminator conformationno TRAP bound: AT more stable
25
2. Processive antitermination• requires special proteins (AT complex) which associate and travel with RNAP
• loading of ATC upstream of terminator• can read through multiple terminatorsover many thousands of bases
• well-studied examples:-- N and Q from phage lambda-- rRNA operons
N• N recruited by Nut site in RNA
Q
• Q recruited by qut site in DNA
• mechanism: stabilizes paused TEC
! Q and N necessary for transcribing long operons (Q: 23,000nt)
27

14
Q and N control long operons (Q: 23 kb !!!)
→ probability of spontaneous termination ?
estimate TEC stability using the model of Yager & von Hippel
• termination efficiency at normal site ~ 5 x 10-4
(possibly higher in vivo due to rho)
• normal operon (length < 5000 nt): 80 % of initiated transcripts reach end (if no internal termination sites)
• lambda late operon (23000 nt): 32 %
Q: 10-fold stabilization: restored to 90%
→ AT: control of operon + adaptation of TEC to operon length
28