Embed Size (px)
Citation preview


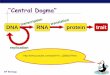
Central Dogma
Genetic information, stored in chromosomes and
transmitted to daughter cells via DNA replication is
expressed through transcription to RNA and
subsequent translation into polypeptide chains.
Central dogma: the flow of information from DNA to
RNA to protein.

Genetic Code
It is a dictionary that express the relation between
a sequence of nucleotides and a sequence of AAs.
Each individual word (codon) is composed of three
nucleotide and indicates an AA in the protein.
Codons are presented in mRNA language as
adenine (A), guanine (G), cytosine (C) & uracil (U).
Since 4 nucleotide bases are used to produce 3-base codons; that is (43) → 64 different
combinations of bases.

Term = chain terminator
codon.
AUG, codes for Met &
serves as the initiator
codon and also encodes
for internal Met in a
protein.
In mammalian
mitochondria,
AUA codes for Met
UGA for Trp
AGA and AGG serve as
chain terminators.
Genetic Code


Most AAs are coded by more than one codon?
Of the 64 codons, 61 code for the 20 AAs involved
in protein structure Val has 4 codons (GUU, GUG, GUC, GUA)
Phe has 2 codons (UUU, UUC)
Met has only one codon (AUG).
Termination (stop or nonsense) codons:
The 3 codons UAG, UGA, and UAA do not code for
amino acids, but are termination codons.
When one of these codons appears in an mRNA, it
signals that synthesis of the peptide chain coded for
by that mRNA is completed.
Degeneracy of codon

Consequences of altering the
nucleotide sequence
Point mutation: changing a single nucleotide base on the mRNA chain → one of three possible results
1. Silent mutation: The codon
containing the changed base
codes for the same amino acid.
2. Missense mutation: The codon
containing the changed base codes
for a different amino acid.
3. Nonsense mutation: The codon
containing the changed base
becomes a termination codon.

Possible effects of point mutations

Characteristics of genetic code
1- Specificity:
a specific codon always codes for the same AA.
2- Universality:
In all living organisms the genetic code is the same.
Exceptions???
3- Redundancy (degeneracy):
A given AA may have more than one triplet codon,
e.g. Arg & Leu each have six different codons.
4. Nonoverlapping and commaless:
the code is read from a fixed starting point as a
continuous sequence of bases, taken three at a time.

Reading frame

If one or two nucleotides
are either deleted from
or added to the interior of a message sequence →
frame- shift mutation
and the reading frame is
altered.
The resulting AA
sequence become
radically different from
this point.
Frame shift mutation

Components Required For Translation
(Protein Synthesis)
1. All the AAs that are found in the finished product.
2. The mRNA to be translated.
3. tRNAs.
4. Functional ribosomes.
5. Energy sources.
6. Enzymes & protein factors needed for the initiation,
elongation & termination of the polypeptide chain.
These gather in cytosol, like raw materials are
brought together in a factory, prior to turning on the
assembly line that creates a finished product.

Amino acids
All the AAs that eventually appear in the finished
protein must be present at the time of protein
synthesis.
If one AA is missing (e.g. the diet does not contain
an essential AA) that amino acid is therefore in
limited supply in the cell and translation stops at
the codon specifying that amino acid.
This demonstrates the importance of having all
essential AAs in sufficient quantities in the diet to
insure continued protein synthesis

Transfer RNA (tRNA)
At least one specific tRNA molecule is required per
AA.
In humans there are at least 50 different tRNA.
Because there are only 20 different AAs, some AAs
have more than one specific tRNA molecule.
This is particularly true of those amino acids that are
coded for by several codons.

1. Amino acid attachment site:
Each tRNA has an attachment site for a specific AA
at its 3'- end (-CCA-amino acid).
When a tRNA has a covalently attached AA it is said
to be charged.
when tRNA is not bound to an AA it is described as
being uncharged.
The AA that is attached to the tRNA molecule is said
to be activated.

2. Anticodon:
Three-base nucleotide
sequence that recognizes a
specific codon on mRNA.
This codon specifies the
insertion of the AA carried
by that tRNA into the
growing peptide chain .
tRNAs are known as
adaptor molecules??
Because they carry a
specific AA and recognize
the codon for that AA

Family of enzymes that attach AAs
to their corresponding tRNAs.
Each enzyme recognizes a specific
L-AA & the corresponding tRNA.
The enzyme catalyzes a two-step reaction → covalent attachment of
AA to the 3'-end of corresponding
tRNA.
The extreme specificity of the
synthetase in recognizing these
two structures is mainly
responsible for the high fidelity of
translation of the genetic message.
Aminoacyl-tRNA synthetases

Ribosomes are large complexes of
protein and rRNA.
They consist of two subunits-
one «large» and one «small» whose
relative sizes are given in terms of
their sedimentation coefficients,
or S (Svedberg) values.
Pro- and eukaryotic ribosomes are
similar in structure, and serve the
same function i.e. factories for the
synthesis of proteins.
Functionally competent
ribosomes

Protein Factors
1. Initiation,
2. Elongation
3. Termination (or release)
Some of these protein factors perform a
catalytic function,
whereas others stabilize the synthetic
machinery.

Sources of energy
ATP and GTP
Cleavage of four high-energy bonds is required for
adding one AA to the growing polypeptide chain:
A. Two from one ATP in aminoacyl-tRNA
synthetase reaction (one in removing PPi, and
one in the subsequent hydrolysis of the PPi to
inorganic phosphate by pyrophosphatase)
B. Two from two GTPs (one for binding the
aminoacyl-tRNA to the A site, and one for the
translocation step).

Codon Recognition by tRNA
Recognition of a particular codon in mRNA is
achieved by the anticodon sequence of the tRNA.
Some tRNAs can recognize more than one codon for a given amino acid.
A. Antiparallel binding between codon and
anticodon
B. "Wobble" hypothesis

A. Antiparallel binding between
codon and anticodon
Binding of tRNA anticodon to mRNA codon
follows the rules of complementary and antiparallel
binding:
the mRNA codon is read 5'→3' by an anticodon
pairing in the flipped (3'→5’) orientation.
When writing the sequences of both codons and
anticodons, the nucleotide sequence must
ALWAYS be listed in the 5'→3' order.

B. "Wobble" hypothesis
It describes mechanism by which
tRNAs can recognize more than
one codon for a specific AA.
The base at the 5'-end of the
anticodon (the «first base of the
anticodon) is not as spatially
defined as the other two bases.
Movement of that first base allows
nontraditional base pairing with the
3'-base of the codon (the «last»
base of the codon).

B. "Wobble" hypothesis
This movement is called «wobble» and allows
a single tRNA to recognize more than one
codon.
The result of this wobbling is ‘that there need
not be 61 different tRNA in order to read the
61 codons coding, for amino acids.

Steps in Protein Synthesis
Protein synthesis translates 3-letter alphabet
of nucleotide sequences on mRNA → 20-
letter alphabet of AAs.
mRNA is translated from its 5'- to 3'-end,
producing a protein synthesized from its
amino-terminal (N-terminal) to its
carboxyl-terminal (C-terminal).
Eukaryotic protein synthesis resembles that
of prokaryotes in most aspects.

Steps in Protein Synthesis
Each eukaryotic mRNA has only one coding
region: it is monocistronic.
In contrast, prokaryotic mRNAs often have
several coding regions: they are
polycistronic . Each coding region has its
own initiation and termination codon and
produces a separate type of polypeptide.
Translation is divided into 3 separate steps:
a- initiation. b- elongation. c- termination.




A) Initiation
It involves assembly of components of translation
system before peptide bond formation occurs. ???
1. two ribosomal subunits,
2. the mRNA to be translated,
3. the Met-tRNA
4. GTP (which provides energy for the process),
5. Initiation factors that facilitate the assembly of this
initiation complex

3.In prokaryotes, there are 3 initiation factors
(IF-1, IF-2, and IF-3), whereas in eukaryotes,
there are over ten (eIF to indicate eukaryotic
origin).






80S ribosome has 2 sites,
1. P-site contains the peptidyl-tRNA
2. A- site contains new amino acyl-tRNA.
Aminoacyl-tRNA for first codon: Met-tRNA enters at
P-site, leaving the A-site free and elongation cycle is
ready to start.

New aminoacyl-tRNA + GPT + EF-1 → complex that
enters site A, then EF-1 and GDP + Pi are released.
Peptidyl transferase catalyzes peptide bond
formation .
Uncharged tRNA moves from P to E site & then is
released.
The newly formed peptidyl- tRNA at the A-site
moves to P-site in the presence of GTP and EF-2
(translocase).
Now, the new aminoacyl-tRNA can come to A-site.

eEF-1 + GTP eEF-1 + GDP + Pi

It occurs when one of the chain terminating codons
(UAA, UAG or UGA) appears in the A-site.
No tRNA can recognize this codon, the releasing
factor + GTP+ peptidyl-tRNA cause hydrolysis of the
bond between the peptide and tRNA at the P-site.
80S ribosome is broken into 60S + 40S to finally
give separate peptide, tRNA, 40S, 60S, releasing
factor and GDP+Pi.


Polysomes (polyribosomes)
Multiple ribosomes on the same mRNA molecule
form a polyribosome, or «polysome» that can
translate the same mRNA molecule simultaneously.
Because of their relatively large size, the ribosome
particles cannot attach to an mRNA any closer than
35 nucleotides apart.



Post-translational Modification of
Polypeptide Chains
Many polypeptide chains are covalently modified,
either while they are still attached to the ribosome or
after their synthesis has been completed.
These modifications may include:
1. Removing part of the translated sequence.
2. Covalent addition of one or more chemical
groups required for protein activity.


Trimming
Many secretory proteins are initially made as large,
precursor molecules that are functionally inactive.
These precursor proteins are cleaved in the ER, the
Golgi apparatus, in developing secretory
vesicles (e.g. insulin), or after secretion.
e.g. Zymogens are inactive precursors of secreted
enzymes that become activated through cleavage
once they have reached their proper sites of action;
pepsinogen, becomes activated in the stomach.
The synthesis of enzymes as zymogens protects
the cell from being digested by its own products.

Covalent alterations
Proteins, both enzymatic and structural, may be
activated or inactivated by covalent attachment of a
variety of chemical groups:
1. Phosphorylation:
2. Glycosylation
3. hydroxylation

1. Phosphorylation:
It occurs on OH of serine,
threonine, or less frequently,
tyrosine residues in a
protein.
It is catalyzed by protein
kinases and may be
reversed by protein
phosphatases.
The phosphorylation may
increase or decrease the
functional activity of the
protein.

2. Glycosylation:
Proteins destined to become
part of a plasma membrane or
secreted from the cell have
carbohydrate chains attached
to serine or threonine OH (O-
linked) or asparagine (N-
linked).
The stepwise addition of sugars
occurs in the endoplasmic
reticulum and the Golgi
apparatus.

3. Hydroxylation
Proline and lysine of the α-chains of collagen are
extensively hydroxylated in the endoplasmic
reticulum.