-
COPYRIGHT 2008 BY THE JOURNAL OF BONE AND JOINT SURGERY,
INCORPORATED
111
Cellular Strategies for Enhancement of Fracture Repair
By Thomas E. Patterson, PhD, Ken Kumagai, MD, PhD, Linda
Griffith, PhD, and George F. Muschler, MD
Tissue engineering seeks to translate scientific knowledge into
tangible products to advance the repair, replace-ment, or
regeneration of organs and tissues. Current tissue engineering
strategies have progressed recentlyfrom a historical approach that
is based primarily on biomaterials to a cell and tissue-based
approach that in-cludes understanding of cell-sourcing and
bioactive stimuli. New options include methods for harvest and
trans-plantation of tissue-forming cells, bioactive matrix
materials that act as tissue scaffolds, and delivery ofbioactive
molecules within scaffolds. These strategies are already benefiting
patients, and they place increasingdemands on orthopaedic surgeons
to have a solid foundation in the contemporary concepts and
principles ofcell-based tissue engineering.
Essentially all orthopaedic tissue engineering strategies can be
distilled to a strategy or combination of strategiesthat seek to
increase the number or relative performance of bone-forming cells.
The global term connective tissueprogenitors has been used to
define the heterogeneous populations of stem and progenitor cells
that are found innative tissue and that are capable of
differentiating into one or more connective tissue phenotypes.
These stem orprogenitor populations are found in various tissue
sources, with varying degrees of ability to differentiate
alongconnective tissue lineages. Available cell-based strategies
include targeting local cells with use of scaffolds or bio-active
factors, or transplantation of autogenous connective tissue
progenitor cells derived from bone marrow orother tissues, with or
without processing to change their concentration or prevalence. The
future may includemeans of homing circulating connective tissue
progenitor cells with use of intrinsic chemokine systems, or
modify-ing the biological performance of connective tissue
progenitor cells by means of genetic modifications.
Stem and Progenitor Cells in Musculoskeletal Tissues
tem and progenitor cells are present in all adult tissuesand are
critical to tissue health, maintenance, and re-sponse to injury or
disease throughout life. Stem cells
give rise to progenitor cells and are distinguished from themby
their capacity for self-renewal by a process of asymmetriccell
division. Progenitor cells, by definition, have finite limitson
their capacity for self-renewal and generally progress togive rise
to one or more differentiated phenotypes1,2.
Stem and progenitor cell populations are the upstreamcomponents
of continuous systems of cell renewal in virtuallyall human
tissues. This turnover is most evident in tissues thatremodel
rapidly, such as the lining cells of the gastrointestinaltract
(replaced every three days) or dermis (replaced every twoweeks). In
bone, turnover is much slower. Osteocytes or bone-
lining cells, the differentiated cells that define bone tissue,
maysurvive for twenty years in human cortical bone.
However,continuous remodeling requires the formation of many
newosteoblasts. Osteoblasts, in turn, are continuously derivedfrom
a much smaller number of preosteoblasts and upstreamprogenitor
cells. The number of true stem cells needed to sup-port this
process may be very small (on the average, less thanone in 20,000
nucleated cells in native marrow). The activa-tion of stem cells
and the proliferation of progenitor cells toform new osteoblasts
are vastly accelerated as a result oftrauma, such as
fractures2.
In the 1960s, Burwell showed that implantation of can-cellous
bone grafts induced bone formation, which could betraced to the
activity of primitive osteogenic cells in bonemarrow3.
Friedenstein4 showed that new bone was formed byproliferating
fibroblast-like marrow cells and that the number
S
Disclosure: In support of their research for or preparation of
this work, one or more of the authors received, in any one year,
outside funding orgrants in excess of $10,000 from the National
Institutes of Health (the National Institute of Arthritis and
Musculoskeletal and Skin Diseases[NIAMS] and the National Institute
of General Medical Sciences [NIGMS]) and Therics, Inc. One or more
of the authors, or a member of his or herimmediate family,
received, in any one year, payments or other benefits or a
commitment or agreement to provide such benefits from
commercialentities in excess of $10,000 (DePuy, Synthes, and
Therics, Inc.) and less than $10,000 (Orthofix). No commercial
entity paid or directed, oragreed to pay or direct, any benefits to
any research fund, foundation, division, center, clinical practice,
or other charitable or nonprofit organizationwith which the
authors, or a member of their immediate families, are affiliated or
associated.
J Bone Joint Surg Am. 2008;90(Suppl 1):111-9
doi:10.2106/JBJS.G.01572
Patterson.fm Page 111 Friday, January 11, 2008 10:38 AM
-
112
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R S
of these proliferating cells could be assayed by counting
thenumber of fibroblastic colony-forming units in vitro. An
in-triguing feature of many tissues, including
musculoskeletaltissues, is that upstream progenitor cells are often
multipotent.Stem cells derived from bone, bone marrow, and
peritrabecu-lar tissues in cancellous bone, periosteum, cartilage,
muscle,fat, and vascular pericytes are capable of differentiation
intomultiple phenotypes, including bone, cartilage, tendon,
liga-ment, fat, muscle, and nerve5-18. This has important
implica-tions with regard to the design of tissue engineering
strategies,in that cells derived from one tissue might be useful in
form-ing other tissue types. Harvest of these tissues varies with
re-spect to the associated host morbidity19-25. Aspiration of
bonemarrow is associated with the least morbidity and provides
asingle-cell suspension that can be readily processed
intraoper-atively for immediate implantation13. Fat has also been
pro-posed as a low-morbidity tissue source, although it
requiresgreater processing11,26. Many names have been used to
describethe colony-forming cells found in bone marrow,
periosteum,or trabecular bone, in addition to fibroblastic
colony-formingunits. These terms include mechanocytes, bone marrow
stro-mal cells, and mesenchymal stem cells, although the
precisedefinition and biological capabilities ascribed by these
termsare not entirely synonymous.
For purposes of describing experimental and clinical re-sults
with use of primary cells isolated from human tissue andgrown
without culture expansion, we have defined the termconnective
tissue progenitor as the entire heterogeneous po-pulation of stem
and progenitor cells that are capable of dif-ferentiating into one
or more connective tissue phenotypes,including bone, fat,
cartilage, blood, and fibrous tissues1. Thenumber of connective
tissue progenitor cells in various tissuesis most often estimated
with use of the colony-forming unitassays, that is, cells that give
rise to a colony of proliferatingprogenitor cells in vitro under
conditions that are selected topromote activation and proliferation
of one or more fractionsof the connective tissue progenitor
population. In these assays,each colony represents the progeny of a
founding stem or pro-genitor cell. Functional biological
differences between thecolony-founding cells are revealed by
differences in the prolif-eration rate, the morphology, the
migration, and the differen-tiation of their progeny in each colony
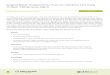
(Fig. 1).
The term connective tissue progenitor recognizes thatthese
tissue-derived cells are not a pure or uniform popula-tion and may
be derived from more than one pool of stemcells and progenitor
cells in native tissues. These cells may in-clude true quiescent,
multipotent stem cells that become acti-vated after harvest and are
capable of self-renewal. However,colonies may also be formed by
cells that are already prolifer-ating in vivocells that lack
self-renewal capabilities andmay exhibit intrinsic commitment to
various stages of diverselineages17,27,28. This diversity can be a
source of frustration forthose looking for homogeneous purified
populations of cells,but in practice this diversity should be
expected, given themultifunctional nature of the bone marrow
environment.Therefore, this diversity can also be viewed as a
source of
valuable information that can be dissected experimentally
tounderstand the prevalence and kinetics of various
connectivetissue stem-cell populations and to understand the ways
inwhich these populations change according to age, gender, dis-ease
states, pharmacological intervention, and tissue engi-neering
strategies1,13,29.
Another experimental and tissue engineering approachto study
stem and progenitor populations from bone marrowor other tissue
sources involves the in vitro expansion of cellsderived from
connective tissue progenitor cells. Various nameshave been given to
culture-expanded populations of cells thathave been selected under
different culture conditions, includ-ing bone marrow stromal
cells15, mesenchymal stem cells30,and adult multipotential
progenitor cells31,32. These terms arenot synonymous1, but they all
denote that progenitor cells canbe isolated and expanded under
appropriate conditions andthat these cells can retain the capacity
to differentiate into avariety of musculoskeletal
phenotypes15,17,30. The term mesen-chymal stem cell has acquired a
particularly narrow definitionof bone marrow-derived,
culture-expanded cells that are iso-lated and expanded according to
the methods pioneered byCaplan33.
Culture-expanded cell populations differ dramaticallyfrom the
heterogeneous population of connective tissue pro-genitor cells
that are present in native tissue. Under in vitroculture
conditions, the heterogeneous population rapidly be-comes more
homogeneous. When cells are grown in vitro,clones of cells that
divide most rapidly and those that have thegreatest capacity for
continued proliferation have a competi-tive advantage. In vitro
expansion therefore produces a strongselective pressure favoring
these traits.
All tissues vary substantially with respect to cellularityand
the prevalence of connective tissue progenitor cells. As-pirated
bone marrow is the best characterized source ofconnective tissue
progenitor cells, containing a mean of ap-proximately forty million
nucleated cells and approximately2000 connective tissue progenitor
cells per milliliter. How-ever, the yield of connective tissue
progenitor cells can varywidely between individuals, aspiration
sites, and even be-tween individual aspirations13,25,29. In
contrast, fat is far lesscellular (approximately six million cells
per cubic centimeterof tissue) and the prevalence of connective
tissue progenitorcells is greater (as high as one per 4000 cells),
but fat-derivedconnective tissue progenitor cells exhibit different
patterns ofproliferation, migration, and differentiation than do
bonemarrow-derived connective tissue progenitor cells.
Differences between connective tissue progenitor cellsharvested
from different individuals and from various tissuesources are
likely a function of the health and histologicalcharacteristics of
the local tissues and reflect the underlyingkinetics of stem cell
function in that tissue. These variables arein turn influenced by
age, gender, and both local and systemicdisease1,13,29,34. For
example, bone marrow cellularity declineswith age. There is also an
age-related decline in the prevalenceof connective tissue
progenitor cells, at least in women29. How-ever, age and gender
account for only a small fraction of the
Patterson.fm Page 112 Friday, January 11, 2008 10:38 AM
-
113
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R
variation in the concentration and prevalence of
connectivetissue progenitor cells among patients13,25,29. As a
result, osteo-genic connective tissue progenitor cells can be
harvested withuse of bone marrow aspiration in patients of all
ages.
Differences in biological potential among connectivetissue
progenitor cells derived from various tissues can haveimportant
practical implications with regard to the selectionof cell sources
for tissue engineering. Individual bone marrow-derived connective
tissue progenitor cells can be capable ofdifferentiating into a
broad range of phenotypes, includingbone, fibrous tissue, fat,
muscle, cartilage, and perhaps evenneural tissue, liver, and
cardiac muscle1,2,15,27,30,35,36. Connectivetissue progenitor cells
derived from muscle, fat, and cartilagealso have a broad repertoire
of intrinsic differentia-tion11,12,20,21,24,37. Some studies have
suggested that fat-derivedand bone marrow-derived cells are
similar38, but others havedemonstrated a decreased osteogenic
potential in fat-derived
cells39 and the absence of surface markers characteristic of
os-teoblastic progenitor cells40.
Potential Strategies for Use of Autogenous Connective Tissue
Progenitor Cells in Therapeutic Applications
here are five major types of cell-based tissue engineering:(1)
local targeting of connective tissue progenitor cells
where new tissue is needed, (2) homing of connective
tissueprogenitor cells into areas where they may not currently
re-side, (3) physically transplanting autogenous connective
tissueprogenitor cells to augment the local population, (4)
trans-planting culture-expanded or modified connective tissue
pro-genitor cells, and (5) transplanting fully formed tissue.
Targeting Connective Tissue Progenitors in SituLocal targeting
strategies are designed to promote desired tis-
T
Fig. 1
Heterogeneity of connective tissue progenitor cells. The
colonies shown in this image were cul-
tured from human bone marrow for nine days and then stained for
alkaline phosphatase activity,
a marker of early bone formation. The image illustrates that
colonies differ in size, cell density,
and the extent and distribution of alkaline phosphatase
activity. These morphologic differences
are manifestations of intrinsic differences among connective
tissue progenitor cells at the time
that they were harvested from bone marrow and placed into
culture. (Reprinted from: Muschler
GF, Nakamoto C, Griffith LG. Current concepts review.
Engineering principles of clinical cell-based
tissue engineering. J Bone Joint Surg Am. 2004;86:1541-58.)
Patterson.fm Page 113 Friday, January 11, 2008 10:38 AM
-
114
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R S
sue formation by stimulating the activation, migration,
pro-liferation, and/or differentiation of local connective
tissueprogenitor cells. The strategy relies on a sufficient local
popula-tion of connective tissue progenitor cells. Tissue scaffolds
pro-vide a surface on which cells and connective tissue
progenitorcells can attach, proliferate, migrate, and
differentiate. Thesescaffolds also prevent the encroachment of
adjacent tissues intoan area where new tissue is desired. Examples
of targeting in-clude implantation of acellular tissue scaffolds
(e.g., allograftbone); locally delivered growth factors (e.g., bone
morphoge-netic proteins); biophysical stimulation, such as
mechanicalloading41-43, electromagnetic stimulation44-46, or
ultrasound47;and systemic pharmacological strategies (e.g.,
parathyroid hor-mone for the treatment of osteoporosis48-53 or
systemic growthhormones to increase muscle mass in the
elderly54,55).
Homing of Connective Tissue Progenitor CellsHoming generally
refers to the recruitment of cells from thesystemic circulation.
Homing and the underlying mechanismsof homing are well established
in hematopoietic connectivetissue progenitor cells. Several studies
have suggested that os-teogenic connective tissue progenitor cells
may travel throughthe systemic circulation56, although the extent
to which circu-lating osteogenic cells contribute to normal
fracture repair isonly beginning to be characterized.
With use of a mouse parabiosis model, Kumagai et al.have
recently shown that circulating cells do home to the siteof a
fibular fracture57. By two weeks after fracture, roughly 6%to 12%
of cells that express alkaline phosphatase in the frac-ture sites
(interpreted as osteoblasts) were found to be derivedfrom
circulating cells57. These and other data suggest that stemcell
homing may be a normal biological process that may be-come the
target of new therapies to enhance the arrival of os-teogenic
connective tissue progenitor cells at sites of bonerepair.
Transplantation of Connective Tissue Progenitor CellsIn many
clinical wound-healing scenarios, new tissue forma-tion may be
hampered by a local deficiency or suboptimal lo-cal population of
connective tissue progenitor cells. This isparticularly true in
regions of previous trauma, infection, irra-diation, scar, or
compromised vascularity. Many studies haveshown that
transplantation of connective tissue progenitorcells into a
bone-healing site can improve the outcome of bothconductive and
inductive grafts, even in sites that are sur-rounded by nondiseased
tissues1,58. This suggests that manyand perhaps all situations of
normal tissue repair may be lim-ited by the population of
connective tissue progenitor cells inlocal tissues.
Autogenous cancellous bone-grafting has long been themost
prevalent and effective example of cell transplantation.Several
clinical studies have suggested that transplantation ofconnective
tissue progenitor cells in aspirated bone marrowhas value in
bone-healing applications3,10,59-65. Additionally,concentration of
bone marrow cells by centrifugation could
increase osteogenesis further. Many surgeons now use bonemarrow
because of its biological value and low risk.
The aspiration technique is important. Muschler et al.found that
limiting the volume of the aspirate to 2 mL persite reduces
dilution with peripheral blood and increases theconcentration of
marrow-derived connective tissue progenitorcells13. Furthermore,
the efficacy of a bone marrow graft canbe enhanced by the use of
certain porous implantable materi-als to selectively concentrate
and select marrow-derived con-nective tissue progenitor cells from
bone marrow, a processknown as selective retention58. Selective
retention of connectivetissue progenitor cells can be used to
rapidly enrich the popu-lation of marrow-derived connective tissue
progenitor cells byremoving red blood cells, serum, and most other
cells in mar-row and contaminating cells from peripheral blood.
Grafts en-riched in this way have improved the results of
bone-graftingin a canine spinal-fusion model58. The use of a
centrifuge toconcentrate low-density cells from bone marrow
(buffy-coatcells) for transplantation has also been described by
bothConnolly66,67 and Hernigou et al.68.
Transplantation of Culture-Expanded CellsCulture-expanded cells
from muscle, fat, and bone marrowmay be useful in regeneration of
bone, cartilage, muscle, andtendon tissue6,35,69-75. In vitro
expansion can generate a largenumber of progenitor cells; however,
it also adds substantialcost and some risks, such as contamination
with bacteria orviruses or depletion of proliferative
capacity76-78. This strategyis already applied clinically in the
area of cartilage repair79-81. Invitro selection of the most
rapidly proliferating cells may alsoselect cells with mutations or
epigenetic changes that mightconfer a tumor-forming potential,
although we are not awareof any reports of human tumors formed by
culture-expandedcells and the risk of tumor formation appears to be
very low.
Transplantation of Genetically Modified Cells and Their
ProgenyThe intrinsic biological potential and performance of
connec-tive tissue progenitor cells and their progeny can be
geneticallymodified by either transiently or permanently altering
the ex-pression of one or more genes82. Advances in genetic
engineer-ing techniques facilitate the efficient engineering of
cells thatsecrete factors (e.g., bone morphogenetic protein-2) that
ef-fect a change in local tissue formation83. Although
transplanta-tion of genetically modified cells may not play a role
in electiveclinical tissue engineering in the near future, it has
substantialpotential value, particularly in the setting of
inherited geneticdefects (e.g., osteogenesis imperfecta84) and for
tissues (such ascartilage) that consist of relatively homogeneous
long-livedcells and in which stable phenotypic expression may be a
cur-rent limitation in biological outcome85,86.
Mass Transport Limitations and Metabolic Demand
n all cell-transplantation settings, access to substrate
mole-cules and clearance of metabolic products are critical to
cellI
Patterson.fm Page 114 Friday, January 11, 2008 10:38 AM
-
115
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R
survival. Due to high demand and slow diffusion rates for
ox-ygen, few cells tolerate diffusion distances in excess of 0.2
mm.At a graft site in which the radius of the graft is
approximately5 mm, diffusion of oxygen is able to support only a
limitednumber of transplanted cells, resulting in central
necrosis.This limitation highlights the need to design bone grafts
that(1) limit the total number of cells and (2) increase the
fractionof transplanted cells that will contribute to tissue
repair, eitherby positive selection or by negative selection
(removal of non-contributing, inhibitory, or competing cells).
Design and Selection of Scaffolds for Tissue Engineering
hree-dimensional porous scaffolds can be designed withspecific
architectures at the nano, micro, and macro scale
(i.e., molecular, cellular, and tissue-length scales,
respectively).Desirable scaffold features include (1) preservation
of a tis-sue volume for the formation of new tissue; (2)
nanoporos-ity and microporosity that allows effective mass
transport tosupport transplanted cells; (3) a connected
microporosity andmacroporosity that allows for contiguous tissue
and vascularingrowth; (4) a surface chemistry and texture that
enhancesthe attachment, migration, proliferation, and
differentiationof osteogenic connective tissue progenitor cells;
and (5) degra-dation properties that are consistent with
preservation of thehealth of local tissues and effective ongoing
remodeling. Cur-rent scaffold options include tissue-derived
materials, biologi-cal polymers, ceramics or mineral-based
matrices, and metalsas well as composites of two or more materials.
The overallmechanical properties of a scaffold (strength, modulus,
andtoughness) are determined both by the material properties ofthe
bulk material and by its three-dimensional structure.Matching the
mechanical properties of a scaffold to the graftenvironment is
critically important so that progression oftissue-healing is not
limited by mechanical failure of the scaf-fold prior to successful
tissue regeneration. Rapidly evolvingthree-dimensional fabrication
methods (e.g., three-dimen-sional printing and three-dimensional
stereolithography) aswell as the development of new materials
(e.g., polycarbo-nates87-89 and polypropylene fumarates90-93)
provide highlypromising platforms for future development.
The surface chemistry defines much of the environ-ment that
cells will experience soon after implantation andhas a profound
effect on the attachment and survival of cellsfollowing
implantation as well as on early proliferation,migration, and
differentiation. Implanted materials rapidlybecome coated with
proteins and lipids, which are the prin-cipal mediators of the
cellular response to most materials. Ithas been speculated that
hydroxyapatite and some other ce-ramics may preferentially
sequester bioactive molecules thatare important for bone
regeneration. Indeed, hydroxyapa-tite and tricalcium phosphate
materials perform successfullyas depot delivery vehicles for bone
morphogenetic proteinsboth in animals94 and humans95,96. Protein
adsorption can,however, induce a conformational change, which may
hideor expose sites that interact with cell-surface receptors.
For
example, fibronectin is a more active adhesion moleculeon
hydrophilic surfaces (e.g., glass) than on hydrophobicsurfaces
(e.g., polytetrafluoroethylene [Teflon] or polyethyl-ene)97-100.
The attachment, survival, proliferation, and dif-ferentiation of
stem and progenitor cells can be modulatedin vitro if scaffold
surfaces are precoated with selected bioac-tive proteins, including
bone morphogenetic protein-2 andbone morphogenetic
protein-7101-105.
Proteins and small bioactive peptides can also be selec-tively
concentrated and presented by covalently linking themto a
surface96,106,107. This provides more control over conforma-tion, a
slower rate of release from the surface, and longer re-tention.
Presentation of growth factors in a matrix-boundfashion may better
mimic the native physiology of many sol-uble signaling molecules,
including most proteins. Tetheringmay not be appropriate, however,
for signaling moleculesthat need to be internalized (e.g., steroid
hormones)108-111.This strategy may also be particularly well suited
for the de-sign of matrices with selective affinity for specific
cells or setsof cells (e.g., connective tissue progenitor cells,
endothelialcells, and platelets).
The pharmacokinetics of delivery of bone morphoge-netic proteins
has been shown to be an important clinicalvariable in a variety of
materials, including degradablepolymers112-119, type-I
collagen120-122, and calcium phosphateceramics123,124. The
retention time of implanted bone morpho-genetic protein correlates
with biological efficacy, presumablybecause the longer a bone
morphogenetic protein is retained,the higher the probability that
it will act on an appropriatetarget cell. Retention time has been
related both to solubility125
and to isoelectric point94.Current clinical strategies for
protein delivery are tech-
nically simple but require that the protein be delivered in
ahigh concentration in order to diffuse into adjacent tissuesand
act on local connective tissue progenitor cells. The disad-vantage
of these strategies is that they provide relatively littlecontrol
over the rate of delivery, conformation, presentation,clearance, or
degradation of the delivered protein. Althoughcurrent strategies
for delivery of bone morphogenetic proteinscan be effective, the
vast majority of the massively supraphysi-ological doses of bone
morphogenetic proteins that are cur-rently required are likely
wasted, and only a small fractionactually elicits a
receptor-mediated signal that enhances newbone formation. These
methods, therefore, leave substantialroom for improvement in
delivery kinetics and distribution ofbioactive proteins.
A scaffold designed to deliver viable cells must providean
environment with physiologic pH and osmolarity. The de-sign of
scaffold bulk material must also consider the effects ofdegradation
products. The degradation of many polyester-based matrices, such as
polylactides and polyglycolides, pro-duces acidic degradation
products (lactic acid and glycolicacid), and therefore those
matrices are not ideal for cell trans-plantation and tissue
regeneration. Similarly, materials thatresult in an early
hyperosmolar environment, such as glycerol(used to improve handling
of many bone pastes) and calcium
T
Patterson.fm Page 115 Friday, January 11, 2008 10:38 AM
-
116
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R S
sulfate (both acidic and hyperosmolar), are likely not
appro-priate materials for use in a cell-transplantation
environment.
Opportunities for Rational Design of Future Materials, Devices,
and StrategiesOptimizing combinations of cells, matrices, and
locally andsystemically active stimuli will remain a complex
process char-acterized by a highly interdependent set of variables
with analmost infinite range of possible combinations. Future
tissueengineering strategies that are particularly rich in
opportunityinclude (1) improved methods for intraoperative harvest,
con-centration, and selection of stem and progenitor cells; (2)
cell-delivery systems that enhance cell survival by managing
thebalance of mass transfer and metabolic demand; (3)
three-dimensional scaffolds with architectural and
mechanicalfeatures that are customized for specific clinical
applications;(4) chemically defined surfaces that present
covalently teth-ered, biologically active molecules; (5) defined
microtexturedsurfaces to elicit desired cell attachment, migration,
differenti-ation, and survival; (6) scaffold materials for which
degrada-tion delivers biologically inert or even bioactive
molecules,minimizing the toxicity associated with degradation
prod-ucts; and (7) delivery systems for soluble molecule (e.g.,
bonemorphogenetic proteins and other protein growth
factors)delivery systems that ensure both a biologically active
con-formation and provide a local concentration profile that is
ap-propriate for the target cell population, minimizing the
totaldose of bioactive agent that is required and the attendant
riskof unwanted collateral effects.
These developments in cell-based approaches to im-
prove fracture repair must also be informed by a combinationof
clinical experience, knowledge of basic biological princi-ples,
medical necessity, and commercial practicality. The re-sponsibility
for rational development is shared by the entireorthopaedic
community (developers, vendors, and physi-cians) and must be
focused as much as possible on objectiveand systematic assessment
and reporting. This challenge ismade particularly urgent by the
recent rapid addition of manynew clinical options that often have
narrow and nonoverlap-ping regulatory approval but that also are
rapidly applied inoff-label applications by clinicians earnestly
seeking the bestpossible care for their patients. Prospective,
randomized pre-clinical and clinical trials will continue to play a
critical role inthe initial evaluation of new materials for
specific indications.In addition, prospective cohort studies will
remain critical as ameans of demonstrating that controlled studies
can be gener-alized to the broader orthopaedic community.
Prospectiveregistries related to trauma care will also play an
importantrole by objectively defining settings in which current
practicefalls short of reported or desired outcomes. Such
registriescould be applied directly to define opportunities for
neededprospective trial and identify settings in which
randomizationis impractical due to insufficient power or unethical
due to ex-isting evidence of success or failure in specific
settings.
Corresponding author:George F. Muschler, MDDepartments of
Orthopaedic Surgery and Biomedical Engineering (ND-20), Cleveland
Clinic, 9500 Euclid Avenue, Cleveland, OH 44195. E-mail address:
[email protected]
References
1. Muschler GF, Midura RJ. Connective tissue progenitors:
practical concepts for clinical applications. Clin Orthop Relat
Res. 2002;395:66-80.
2. Muschler GF, Midura RJ, Nakamoto C. Practical modeling
concepts for connec-tive tissue stem cell and progenitor
compartment kinetics. J Biomed Biotechnol. 2003;3:170-93.
3. Burwell RG. Studies in the transplantation of bone. VII. The
fresh composite homograft-autograft of cancellous bone. An analysis
of factors leading to osteo-genesis in marrow transplants and
marrow-containing bone grafts. J Bone Joint Surg Br.
1964;46:110-40.
4. Friedenstein AJ. Precursor cells of mechanocytes. Int Rev
Cytol. 1976;47:327-59.
5. Brighton CT, Lorich DG, Kupcha R, Reilly TM, Jones AR,
Woodbury RA 2nd. The pericyte as a possible osteoblast progenitor
cell. Clin Orthop Relat Res. 1992;275:287-99.
6. Caplan AI, Bruder SP. Mesenchymal stem cells: building blocks
for molecular medicine in the 21st century. Trends Mol Med.
2001;7:259-64.
7. Connolly J, Guse R, Lippiello L, Dehne R. Development of an
osteogenic bone-marrow preparation. J Bone Joint Surg Am.
1989;71:684-91.
8. Dore-Duffy P, Katychev A, Wang X, Van Buren E. CNS
microvascular pericytes exhibit multipotential stem cell activity.
J Cereb Blood Flow Metab. 2006;26:613-24.
9. Friedenstein AJ, Petrakova KV, Kurolesova AI, Frolova GP.
Heterotopic of bone marrow. Analysis of precursor cells for
osteogenic and hematopoietic tissues. Transplantation.
1968;6:230-47.
10. Garg NK, Gaur S. Percutaneous autogenous bone-marrow
grafting in congeni-tal tibial pseudarthrosis. J Bone Joint Surg
Br. 1995;77:830-1. Erratum in: J Bone Joint Surg Br.
1996;78:683-4.
11. Gimble JM, Robinson CE, Wu X, Kelly KA. The function of
adipocytes in the
bone marrow stroma: an update. Bone. 1996;19:421-8.
12. Huard C, Moisset PA, Dicaire A, Merly F, Tardif F, Asselin
I, Tremblay JP. Trans-plantation of dermal fibroblasts expressing
MyoD1 in mouse muscles. Biochem Biophys Res Commun.
1998;248:648-54.
13. Muschler GF, Boehm C, Easley K. Aspiration to obtain
osteoblast progenitor cells from human bone marrow: the influence
of aspiration volume. J Bone Joint Surg Am. 1997;79:1699-709.
Erratum in: J Bone Joint Surg Am. 1998;80:302.
14. ODriscoll SW. Articular cartilage regeneration using
periosteum. Clin Orthop Relat Res. 1999;367 Suppl:S186-203.
15. Owen M, Friedenstein AJ. Stromal stem cells: marrow-derived
osteogenic pre-cursors. Ciba Found Symp. 1988;136:42-60.
16. Peng H, Huard J. Muscle-derived stem cells for
musculoskeletal tissue regen-eration and repair. Transpl Immunol.
2004;12:311-9.
17. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R,
Mosca JD, Moor-man MA, Simonetti DW, Craig S, Marshak DR.
Multilineage potential of adult hu-man mesenchymal stem cells.
Science. 1999;284:143-7.
18. Shi S, Gronthos S. Perivascular niche of postnatal
mesenchymal stem cells in human bone marrow and dental pulp. J Bone
Miner Res. 2003;18:696-704.
19. Bahrami S, Stratmann U, Wiesmann HP, Mokrys K, Bruckner P,
Szuwart T. Peri-osteally derived osteoblast-like cells
differentiate into chondrocytes in suspen-sion culture in agarose.
Anat Rec. 2000;259:124-30.
20. Bosch P, Musgrave D, Ghivizzani S, Latterman C, Day CS,
Huard J. The effi-ciency of muscle-derived cell-mediated bone
formation. Cell Transplant. 2000;9:463-70.
21. Bosch P, Musgrave DS, Lee JY, Cummins J, Shuler T,
Ghivizzani TC, Evans T, Robbins TD, Huard J. Osteoprogenitor cells
within skeletal muscle. J Orthop Res. 2000;18:933-44.
22. Bradham DM, Horton WE Jr. In vivo cartilage formation from
growth factor
Patterson.fm Page 116 Friday, January 11, 2008 10:38 AM
-
117
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R
modulated articular chondrocytes. Clin Orthop Relat Res.
1998;352:239-49.
23. Caterson EJ, Nesti LJ, Albert T, Danielson K, Tuan R.
Application of mesenchy-mal stem cells in the regeneration of
musculoskeletal tissues. MedGenMed. 2001;E1.
24. Halvorsen YC, Wilkison WO, Gimble JM. Adipose-derived
stromal cellstheir utility and potential in bone formation. Int J
Obes Relat Metab Disord. 2000;24 Suppl 4:S41-4.
25. Majors AK, Boehm CA, Nitto H, Midura RJ, Muschler GF.
Characterization of human bone marrow stromal cells with respect to
osteoblastic differentiation. J Orthop Res. 1997;15:546-57.
26. Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ,
Benhaim P, Lorenz HP, Hedrick MH. Multilineage cells from human
adipose tissue: implications for cell-based therapies. Tissue Eng.
2001;7:211-28.
27. Aubin JE. Advances in the osteoblast lineage. Biochem Cell
Biol. 1998;76:899-910.
28. Aubin JE. Osteoprogenitor cell frequency in rat bone marrow
stromal popula-tions: role for heterotypic cell-cell interactions
in osteoblast differentiation. J Cell Biochem. 1999;72:396-410.
29. Muschler GF, Nitto H, Boehm CA, Easley KA. Age- and
gender-related changes in the cellularity of human bone marrow and
the prevalence of osteoblastic pro-genitors. J Orthop Res.
2001;19:117-25.
30. Caplan AI. Mesenchymal stem cells. J Orthop Res.
1991;9:641-50.
31. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD,
Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J,
Aldrich S, Lisberg A, Low WC, Largaespada DA, Verfaillie CM.
Pluripotency of mesenchymal stem cells derived from adult marrow.
Nature. 2002;418:41-9. Erratum in: Nature. 2007;447:879-80.
32. Reyes M, Verfaillie CM. Characterization of multipotent
adult progenitor cells, a subpopulation of mesenchymal stem cells.
Ann N Y Acad Sci. 2001;938:231-5.
33. Caplan AI. Review: mesenchymal stem cells: cell-based
reconstructive ther-apy in orthopedics. Tissue Eng.
2005;11:1198-211.
34. DIppolito G, Schiller PC, Ricordi C, Roos BA, Howard GA.
Age-related osteo-genic potential of mesenchymal stromal stem cells
from human vertebral bone marrow. J Bone Miner Res.
1999;14:1115-22.
35. Bruder SP, Fox BS. Tissue engineering of bone. Cell based
strategies. Clin Or-thop Relat Res. 1999;367 Suppl:S68-83.
36. Pittenger MF, Mosca JD, McIntosh KR. Human mesenchymal stem
cells: pro-genitor cells for cartilage, bone, fat and stroma. Curr
Top Microbiol Immunol. 2000;251:3-11.
37. Tallheden T, Dennis JE, Lennon DP, Sjgren-Jansson E, Caplan
AI, Lindahl A. Phenotypic plasticity of human articular
chondrocytes. J Bone Joint Surg Am. 2003;85 Suppl 2:93-100.
38. De Ugarte DA, Morizono K, Elbarbary A, Alfonso Z, Zuk PA,
Zhu M, Dragoo JL, Ashjian P, Thomas B, Benhaim P, Chen I, Fraser J,
Hedrick MH. Comparison of multi-lineage cells from human adipose
tissue and bone marrow. Cells Tissues Organs. 2003;174:101-9.
39. Winter A, Breit S, Parsch D, Benz K, Steck E, Hauner H,
Weber RM, Ewerbeck V, Richter W. Cartilage-like gene expression in
differentiated human stem cell spheroids: a comparison of bone
marrow-derived and adipose tissue-derived stro-mal cells. Arthritis
Rheum. 2003;48:418-29.
40. Gronthos S, Franklin DM, Leddy HA, Robey PG, Storms RW,
Gimble JM. Sur-face protein characterization of human adipose
tissue-derived stromal cells. J Cell Physiol. 2001;189:54-63.
41. Carter DR, Beaupr GS, Giori NJ, Helms JA. Mechanobiology of
skeletal re-generation. Clin Orthop Relat Res. 1998;355
Suppl:S41-55.
42. Klein-Nulend J, Roelofsen J, Sterck JG, Semeins CM, Burger
EH. Mechanical loading stimulates the release of transforming
growth factor-beta activity by cul-tured mouse calvariae and
periosteal cells. J Cell Physiol. 1995;163:115-9.
43. Turner CH, Owan I, Alvey T, Hulman J, Hock JM. Recruitment
and proliferative responses of osteoblasts after mechanical loading
in vivo determined using sustained-release bromodeoxyuridine. Bone.
1998;22:463-9.
44. Aaron RK, Ciombor DM. Acceleration of experimental
endochondral ossifica-tion by biophysical stimulation of the
progenitor cell pool. J Orthop Res. 1996;14:582-9.
45. Brighton CT, Wang W, Seldes R, Zhang G, Pollack SR. Signal
transduction in electrically stimulated bone cells. J Bone Joint
Surg Am. 2001;83:1514-23.
46. Patterson TE, Sakai Y, Grabiner MD, Ibiwoye M, Midura RJ,
Zborowski M, Wolfman A. Exposure of murine cells to pulsed
electromagnetic fields rapidly
activates the mTOR signaling pathway. Bioelectromagnetics.
2006;27:535-44.
47. Rubin C, Bolander M, Ryaby JP, Hadjiargyrou M. The use of
low-intensity ultra-sound to accelerate the healing of fractures. J
Bone Joint Surg Am. 2001;83:259-70.
48. Cosman F, Nieves J, Woelfert L, Formica C, Gordon S, Shen V,
Lindsay R. Par-athyroid hormone added to established hormone
therapy: effects on vertebral fracture and maintenance of bone mass
after parathyroid hormone withdrawal. J Bone Miner Res.
2001;16:925-31.
49. Finkelstein JS, Klibanski A, Arnold AL, Toth TL, Hornstein
MD, Neer RM. Prevention of estrogen deficiency-related bone loss
with human parathyroid hormone-(1-34): a randomized controlled
trial. JAMA. 1998;280:1067-73.
50. Lane NE, Sanchez S, Modin GW, Genant HK, Pierini E, Arnaud
CD. Parathyroid hormone treatment can reverse
corticosteroid-induced osteoporosis. Results of a randomized
controlled clinical trial. J Clin Invest. 1998;102:1627-33.
51. Morley P, Whitfield JF, Willick GE. Parathyroid hormone: an
anabolic treatment for osteoporosis. Curr Pharm Des.
2001;7:671-87.
52. Reeve J, Meunier PJ, Parsons JA, Bernat M, Bijvoet OL,
Courpron P, Edouard C, Klenerman L, Neer RM, Renier JC, Slovik D,
Vismans FJ, Potts JT Jr. Anabolic effect of human parathyroid
hormone fragment on trabecular bone in involutional osteoporosis: a
multicentre trial. Br Med J. 1980;280:1340-4.
53. Slovik DM, Rosenthal DI, Doppelt SH, Potts JT Jr, Daly MA,
Campbell JA, Neer RM. Restoration of spinal bone in osteoporotic
men by treatment with human par-athyroid hormone (1-34) and
1,25-dihydroxyvitamin D. J Bone Miner Res. 1986;1:377-81.
54. Hennessey JV, Chromiak JA, DellaVentura S, Reinert SE, Puhl
J, Kiel DP, Rosen CJ, Vandenburgh H, MacLean DB. Growth hormone
administration and ex-ercise effects on muscle fiber type and
diameter in moderately frail older people. J Am Geriatr Soc.
2001;49:852-8.
55. Liu PY, Wishart SM, Handelsman DJ. A double-blind,
placebo-controlled, ran-domized clinical trial of recombinant human
chorionic gonadotropin on muscle strength and physical function and
activity in older men with partial age-related androgen deficiency.
J Clin Endocrinol Metab. 2002;87:3125-35.
56. Khosla S, Eghbali-Fatourechi GZ. Circulating cells with
osteogenic potential. Ann N Y Acad Sci. 2006;1068:489-97.
57. Kumagai K, Vasanji A, Drazba JA, Butler RS, Muschler GF.
Circulating cells with osteogenic potential are physiologically
mobilized into the fracture healing site in the parabiotic mice
model. J Orthop Res. 2007 Aug 29 [Epub ahead of print].
58. Muschler GF, Nitto H, Matsukura Y, Boehm C, Valdevit A,
Kambic H, Davros W, Powell K, Easley K. Spine fusion using cell
matrix composites enriched in bone marrow-derived cells. Clin
Orthop Relat Res. 2003;407:102-18.
59. Burwell RG. The fate of bone grafts. In: Apley AG, editor.
Recent advances in orthopaedics. London: Churchill; 1969. p
115-207.
60. Burwell RG. Studies in the transplantation of bone. 8.
Treated composite homograft-autografts of cancellous bone: an
analysis of inductive mechanisms in bone transplantation. J Bone
Joint Surg Br. 1966;48:532-66.
61. Burwell RG. The function of bone marrow in the incorporation
of a bone graft. Clin Orthop Relat Res. 1985;200:125-41.
62. Connolly JF, Guse R, Tiedeman J, Dehne R. Autologous marrow
injection as a substitute for operative grafting of tibial
nonunions. Clin Orthop Relat Res. 1991;266:259-70.
63. Connolly JF, Guse R, Tiedeman J, Dehne R. Autologous marrow
injection for delayed unions of the tibia: a preliminary report. J
Orthop Trauma. 1989;3:276-82.
64. Garg NK, Gaur S, Sharma S. Percutaneous autogenous bone
marrow grafting in 20 cases of ununited fracture. Acta Orthop
Scand. 1993;64:671-2.
65. Healey JH, Zimmerman PA, McDonnell JM, Lane JM. Percutaneous
bone mar-row grafting of delayed union and nonunion in cancer
patients. Clin Orthop Relat Res. 1990;256:280-5.
66. Connolly JF. Clinical use of marrow osteoprogenitor cells to
stimulate osteo-genesis. Clin Orthop Relat Res. 1998;355
Suppl:S257-66.
67. Connolly JF. Injectable bone marrow preparations to
stimulate osteogenic re-pair. Clin Orthop Relat Res.
1995;313:8-18.
68. Hernigou P, Poignard A, Beaujean F, Rouard H. Percutaneous
autologous bone-marrow grafting for nonunions. Influence of the
number and concentration of progenitor cells. J Bone Joint Surg Am.
2005;87:1430-7.
69. Bittira B, Kuang JQ, Al-Khaldi A, Shum-Tim D, Chiu RC. In
vitro preprogram-
Patterson.fm Page 117 Friday, January 11, 2008 10:38 AM
-
118
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R S
ming of marrow stromal cells for myocardial regeneration. Ann
Thorac Surg. 2002;74:1154-60.
70. Glimm H, Eaves CJ. Direct evidence for multiple self-renewal
divisions of hu-man in vivo repopulating hematopoietic cells in
short-term culture. Blood. 1999;94:2161-8.
71. Kadiyala S, Young RG, Thiede MA, Bruder SP. Culture expanded
canine mes-enchymal stem cells possess osteochondrogenic potential
in vivo and in vitro. Cell Transplant. 1997;6:125-34.
72. McKay R. Stem cellshype and hope. Nature.
2000;406:361-4.
73. McNiece I, Briddell R. Ex vivo expansion of hematopoietic
progenitor cells and mature cells. Exp Hematol. 2001;29:3-11.
74. Quarto R, Mastrogiacomo M, Cancedda R, Kutepov SM, Mukhachev
V, Lavroukov A, Kon E, Marcacci M. Repair of large bone defects
with the use of au-tologous bone marrow stromal cells. N Engl J
Med. 2001;344:385-6.
75. Ringe J, Kaps C, Burmester GR, Sittinger M. Stem cells for
regenerative med-icine: advances in the engineering of tissues and
organs. Naturwissenschaften. 2002;89:338-51.
76. Hayflick L, Moorhead PS. The serial cultivation of human
diploid cell strains. Exp Cell Res. 1961;25:585-621.
77. Shi S, Gronthos S, Chen S, Reddi A, Counter CM, Robey PG,
Wang CY. Bone formation by human postnatal bone marrow stromal stem
cells is enhanced by te-lomerase expression. Nat Biotechnol.
2002;20:587-91.
78. Simonsen JL, Rosada C, Serakinci N, Justesen J, Stenderup K,
Rattan SI, Jensen TG, Kassem M. Telomerase expression extends the
proliferative life-span and maintains the osteogenic potential of
human bone marrow stromal cells. Nat Biotechnol. 2002;20:592-6.
79. Bentley G, Biant LC, Carrington RW, Akmal M, Goldberg A,
Williams AM, Skin-ner JA, Pringle J. A prospective, randomised
comparison of autologous chondro-cyte implantation versus
mosaicplasty for osteochondral defects in the knee. J Bone Joint
Surg Br. 2003;85:223-30.
80. Horas U, Pelinkovic D, Herr G, Aigner T, Schnettler R.
Autologous chondrocyte implantation and osteochondral cylinder
transplantation in cartilage repair of the knee joint. A
prospective, comparative trial. J Bone Joint Surg Am.
2003;85:185-92.
81. Knutsen G, Engebretsen L, Ludvigsen TC, Drogset JO, Grntvedt
T, Solheim E, Strand T, Roberts S, Isaksen V, Johansen O.
Autologous chondrocyte implantation compared with microfracture in
the knee. A randomized trial. J Bone Joint Surg Am.
2004;86:455-64.
82. Hannallah D, Peterson B, Lieberman JR, Fu FH, Huard J. Gene
therapy in or-thopaedic surgery. Instr Course Lect.
2003;52:753-68.
83. Bonadio J. Tissue engineering via local gene delivery. J Mol
Med. 2000;78:303-11.
84. Prockop DJ. Marrow stromal cells as stem cells for
nonhematopoietic tis-sues. Science. 1997;276:71-4.
85. Hunziker EB. Articular cartilage repair: basic science and
clinical progress. A review of the current status and prospects.
Osteoarthritis Cartilage. 2002;10:432-63.
86. Peterson L. Articular cartilage injuries treated with
autologous chondrocyte transplantation in the human knee. Acta
Orthop Belg. 1996;62 Suppl 1:196-200.
87. Choueka J, Charvet JL, Koval KJ, Alexander H, James KS,
Hooper KA, Kohn J. Canine bone response to tyrosine-derived
polycarbonates and poly(L-lactic acid). J Biomed Mater Res.
1996;31:35-41.
88. Ertel SI, Kohn J, Zimmerman MC, Parsons JR. Evaluation of
poly(DTH carbon-ate), a tyrosine-derived degradable polymer, for
orthopedic applications. J Biomed Mater Res. 1995;29:1337-48.
89. Meechaisue C, Dubin R, Supaphol P, Hoven VP, Kohn J.
Electrospun mat of tyrosine-derived polycarbonate fibers for
potential use as tissue scaffolding mate-rial. J Biomater Sci Polym
Ed. 2006;17:1039-56.
90. Payne RG, McGonigle JS, Yaszemski MJ, Yasko AW, Mikos AG.
Development of an injectable, in situ crosslinkable, degradable
polymeric carrier for osteogenic cell populations. Part 2.
Viability of encapsulated marrow stromal osteoblasts cultured on
crosslinking poly(propylene fumarate). Biomaterials.
2002;23:4373-80.
91. Payne RG, McGonigle JS, Yaszemski MJ, Yasko AW, Mikos AG.
Development of an injectable, in situ crosslinkable, degradable
polymeric carrier for osteogenic cell populations. Part 3.
Proliferation and differentiation of encapsulated marrow stromal
osteoblasts cultured on crosslinking poly(propylene fumarate).
Biomateri-als. 2002;23:4381-7.
92. Payne RG, Yaszemski MJ, Yasko AW, Mikos AG. Development of
an injectable, in situ crosslinkable, degradable polymeric carrier
for osteogenic cell popula-tions. Part 1. Encapsulation of marrow
stromal osteoblasts in surface crosslinked gelatin microparticles.
Biomaterials. 2002;23:4359-71.
93. Wang S, Lu L, Yaszemski MJ. Bone-tissue-engineering material
poly(propylene fumarate): correlation between molecular weight,
chain dimensions, and physical properties. Biomacromolecules.
2006;7:1976-82.
94. Uludag H, DAugusta D, Golden J, Li J, Timony G, Reidel R,
Wozney JM. Im-plantation of human recombinant bone morphogenetic
proteins with biomaterial carriers: a correlation between protein
pharmacokinetics and osteoinduction in the rat ectopic model. J
Biomed Mater Res. 2000;50:227-38.
95. Boden SD, Kang J, Sandhu H, Heller JG. Use of recombinant
human bone morphogenetic protein-2 to achieve posterolateral lumbar
spine fusion in hu-mans: a prospective, randomized clinical pilot
trial. Spine. 2002;27:2662-73.
96. Griffith LG. Polymeric biomaterials. Acta Mater.
2000;48:263-77.
97. Altankov G, Thom V, Groth T, Jankova K, Jonsson G, Ulbricht
M. Modulating the biocompatibility of polymer surfaces with
poly(ethylene glycol): effect of fi-bronectin. J Biomed Mater Res.
2000;52:219-30.
98. Groth T, Altankov G. Fibroblast spreading and proliferation
on hydrophilic and hydrophobic surfaces is related to tyrosine
phosphorylation in focal contacts. J Biomater Sci Polym Ed.
1995;7:297-305.
99. Groth T, Altankov G, Kostadinova A, Krasteva N, Albrecht W,
Paul D. Altered vitronectin receptor (alphav integrin) function in
fibroblasts adhering on hydropho-bic glass. J Biomed Mater Res.
1999;44:341-51.
100. Lewandowska K, Balachander N, Sukenik CN, Culp LA.
Modulation of fi-bronectin adhesive functions for fibroblasts and
neural cells by chemically deriva-tized substrata. J Cell Physiol.
1989;141:334-45.
101. Burkus JK, Heim SE, Gornet MF, Zdeblick TA. Is INFUSE bone
graft superior to autograft bone? An integrated analysis of
clinical trials using the LT-CAGE lum-bar tapered fusion device. J
Spinal Disord Tech. 2003;16:113-22.
102. Burkus JK, Transfeldt EE, Kitchel SH, Watkins RG,
Balderston RA. Clinical and radiographic outcomes of anterior
lumbar interbody fusion using recombinant human bone morphogenetic
protein-2. Spine. 2002;27:2396-408.
103. Dennis JE, Caplan AI. Porous ceramic vehicles for
rat-marrow-derived (Rat-tus norvegicus) osteogenic cell delivery:
effects of pre-treatment with fibronectin or laminin. J Oral
Implantol. 1993;19:106-15,136-7.
104. Dennis JE, Haynesworth SE, Young RG, Caplan AI.
Osteogenesis in marrow-derived mesenchymal cell porous ceramic
composites transplanted subcutane-ously: effect of fibronectin and
laminin on cell retention and rate of osteogenic expression. Cell
Transplant. 1992;1:23-32.
105. Johnsson R, Strmqvist B, Aspenberg P. Randomized
radiostereometric study comparing osteogenic protein-1 (BMP-7) and
autograft bone in human non-instrumented posterolateral lumbar
fusion. Spine. 2002;27:2654-61.
106. Griffith LG. Emerging design principles in biomaterials and
scaffolds for tis-sue engineering. Ann N Y Acad Sci.
2002;961:83-95.
107. Griffith LG, Naughton GK. Tissue engineeringcurrent
challenges and ex-panding opportunities. Science.
2002;295:1009-14.
108. Kuhl PR, Griffith-Cima LG. Tethered epidermal growth factor
as a paradigm for growth factor-induced stimulation from the solid
phase. Nat Med. 1996;2:1022-27. Erratum in: Nat Med. 1997;3:93.
109. Mann BK, Schmedlen RH, West JL. Tethered-TGF-beta increases
extracellu-lar matrix production of vascular smooth muscle cells.
Biomaterials. 2001;22:439-44.
110. Swindle CS, Tran KT, Johnson TD, Banerjee P, Mayes AM,
Griffith L, Wells A. Epidermal growth factor (EGF)-like repeats of
human tenascin-C as ligands for EGF receptor. J Cell Biol.
2001;154:459-68.
111. Zisch AH, Schenk U, Schense JC, Sakiyama-Elbert SE, Hubbell
JA. Co-valently conjugated VEGFfibrin matrices for
endothelialization. J Contr Rel. 2001;72:101-13.
112. Fischgrund JS, James SB, Chabot MC, Hankin R, Herkowitz HN,
Wozney JM, Shirkhoda A. Augmentation of autograft using rhBMP-2 and
different carrier me-dia in the canine spinal fusion model. J
Spinal Disord. 1997;10:467-72.
113. Isobe M, Yamazaki Y, Mori M, Amagasa T. Bone regeneration
produced in rat femur defects by polymer capsules containing
recombinant human bone morpho-genetic protein-2. J Oral Maxillofac
Surg. 1999;57:695-9.
114. Isobe M, Yamazaki Y, Mori M, Ishihara K, Nakabayashi N,
Amagasa T. The role of recombinant human bone morphogenetic
protein-2 in PLGA capsules at an extraskeletal site of the rat. J
Biomed Mater Res. 1999;45:36-41.
Patterson.fm Page 118 Friday, January 11, 2008 10:38 AM
-
119
THE JOU R N A L OF BO N E & JO I N T SU RG ER Y JB JS .ORGVO
LUM E 90-A SU P P L E M E N T 1 2008
CE L LUL A R ST R A TE G I E S F O R EN H AN CE M E N T OF FR A
C TU RE RE P AI R
115. Kandziora F, Bail H, Schmidmaier G, Schollmeier G, Scholz
M, Knispel C, Hiller T, Pflugmacher R, Mittlmeier T, Raschke M,
Haas NP. Bone morphogenetic protein-2 application by a
poly(D,L-lactide)-coated interbody cage: in vivo results of a new
carrier for growth factors. J Neurosurg. 2002;97(1 Suppl):40-8.
116. Kenley R, Marden L, Turek T, Jin L, Ron E, Hollinger JO.
Osseous regenera-tion in the rat calvarium using novel delivery
systems for recombinant human bone morphogenetic protein-2
(rhBMP-2). J Biomed Mater Res. 1994;28:1139-47.
117. Mayer M, Hollinger J, Ron E, Wozney J. Maxillary alveolar
cleft repair in dogs using recombinant human bone morphogenetic
protein-2 and a polymer carrier. Plast Reconstr Surg.
1996;98:247-59.
118. Weber FE, Eyrich G, Grtz KW, Maly FE, Sailer HF. Slow and
continuous appli-cation of human recombinant bone morphogenetic
protein via biodegradable poly(lactide-co-glycolide) foamspheres.
Int J Oral Maxillofac Surg. 2002;31:60-5.
119. Woo BH, Fink BF, Page R, Schrier JA, Jo YW, Jiang G, DeLuca
M, Vasconez HC, DeLuca PP. Enhancement of bone growth by sustained
delivery of recombi-nant human bone morphogenetic protein-2 in a
polymeric matrix. Pharm Res. 2001;18:1747-53. Erratum in: Pharm
Res. 2003;20:334.
120. Hecht BP, Fischgrund JS, Herkowitz HN, Penman L, Toth JM,
Shirkhoda A.
The use of recombinant human bone morphogenetic protein 2
(rhBMP-2) to pro-mote spinal fusion in a nonhuman primate anterior
interbody fusion model. Spine. 1999;24:629-36.
121. Hollinger JO, Schmitt JM, Buck DC, Shannon R, Joh SP,
Zegzula HD, Wozney J. Recombinant human bone morphogenetic
protein-2 and collagen for bone re-generation. J Biomed Mater Res.
1998;43:356-64.
122. Marden LJ, Hollinger JO, Chaudhari A, Turek T, Schaub RG,
Ron E. Recombi-nant human bone morphogenetic protein-2 is superior
to demineralized bone ma-trix in repairing craniotomy defects in
rats. J Biomed Mater Res. 1994;28:1127-38.
123. Magin MN, Delling G. Improved lumbar vertebral interbody
fusion using rhOP-1: a comparison of autogenous bone graft, bovine
hydroxylapatite (Bio-Oss), and BMP-7 (rhOP-1) in sheep. Spine.
2001;26:469-78.
124. Ripamonti U, Ramoshebi LN, Matsaba T, Tasker J, Crooks J,
Teare J. Bone induction by BMPs/OPs and related family members in
primates. J Bone Joint Surg Am. 2001;83 Suppl 1:S116-27.
125. Brekke JH, Toth JM. Principles of tissue engineering
applied to programma-ble osteogenesis. J Biomed Mater Res.
1998;43:380-98. Erratum in: J Biomed Mater Res. 1999;48:95.
Patterson.fm Page 119 Friday, January 11, 2008 10:38 AM