Embed Size (px)
Citation preview

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �1
Module 2
Cell Signalling Pathways
Synopsis
Cells use a large number of clearly defined signalling pathways to regulate their activity. In this module,attention is focused on the ON mechanisms responsible for transmitting information into the cell. Thesesignalling pathways fall into two main groups depending on how they are activated. Most of them areactivated by external stimuli and function to transfer information from the cell surface to internal effectorsystems. However, some of the signalling systems respond to information generated from within thecell, usually in the form of metabolic messengers. For all of these signalling pathways, information isconveyed either through protein–protein interactions or it is transmitted by diffusible elements usuallyreferred to as second messengers. Cells often employ a number of these signalling pathways, andcross-talk between them is an important feature. In this section, attention is focused on theproperties of the major intracellular signalling pathways operating in cells to regulate their cellularactivity.
During the processes of development, specific cell types select out those signalling systems that aresuitable to control their particular functions. One of the aims of this website is to understand how theseunique cell-specific signalsomes function to regulate different mammalian cell types.
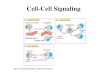
Intracellular signalling pathwaysThere are a large number of intracellular signalling path-ways responsible for transmitting information within thecell. They fall into two main categories. The majority re-spond to external stimuli arriving at the cell surface, usu-ally in the form of a chemical signal (neurotransmitter,hormone or growth factor), which is received by recept-ors at the cell periphery that function as molecular an-tennae embedded in the plasma membrane. These recept-ors then transfer information across the membrane usinga variety of transducers and amplifiers that engage a di-verse repertoire of intracellular signalling pathways (Path-ways 1–16 in Module 2: Figure cell signalling pathways).The phosphoinositide signalling and Ca2+ signalling sys-tems (Pathways 2–6) have been grouped together becausethey contain a number of related signalling pathways thatoften interact with each other. The other categories arethe pathways that are activated by signals generated fromwithin the cell (Pathways 17 and 18). There are a numberof metabolic messengers that act from within the cell toinitiate a variety of signalling pathways.
All of these signalling pathways generate an internalmessenger that is responsible for relaying information tothe sensors that then engage the effectors that activate cel-lular responses. The main features of the signalling path-ways summarized in Module 2: Figure cell signalling path-ways are outlined below:
1. Cyclic AMP signalling pathway. One of the first sig-nalling systems to be characterized was the cyclic
Green text indicates links to content within this module; blue text indicates linksto content in other modules.
AMP signalling pathway, which led to the second mes-senger concept that applies to many other signallingsystems. The idea is that the external stimulus arrivingat the cell surface is the first messenger, which is thentransformed at the cell surface by adenylyl cyclase(AC) into a second messenger, cyclic AMP, which ispart of the signalling cascade that then activates down-stream effectors.
2. Cyclic ADP-ribose (cADPR) signalling and nicotinicacid–adenine dinucleotide phosphate (NAADP) sig-nalling systems function in Ca2+ signalling. An ADP–ribosyl cyclase (ADP-RC) is responsible for generat-ing these two second messengers.
3. Voltage-operated channels (VOCs) contribute toCa2+ signals by controlling the entry of external Ca2+
in excitable cells.4. Receptor-operated channels (ROCs) contribute to
Ca2+ signals by controlling Ca2+ entry in both ex-citable and non-excitable cells.
5. Stimuli that activate phospholipase C (PLC) to hy-drolyse PtdIns4,5P2 (also known as PIP2) generate anumber of signalling pathways:• Inositol 1,4,5-trisphosphate (InsP3)/Ca2+ signalling
cassette• Diacylglycerol (DAG)/protein kinase C (PKC) sig-
nalling cassette• PtdIns4,5P2 signalling cassette• Multipurpose inositol polyphosphate signalling
pathway6. PtdIns 3-kinase signalling is activated by stimuli that
stimulate PtdIns 3-kinase to phosphorylate PIP2 toform the lipid second messenger PtdIns3,4,5P3 (PIP3).
C©2009 Portland Press Limited www.cellsignallingbiology.orgLicensed copy. Copying is not permitted, except with prior permission and as allowed by law.

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �2
Module 2: Figure cell signalling pathways
Wnt catenin β-
Hh GLI
Notch NICD
Ras Erk1/2
Smad TGF-β
NOX O + H O2 22
JAK STAT
CEL
LULA
R RE
SPO
NSE
S
PI 3-K PIP
PLD PA
p38
NF- BκI Bsκ
JNK
SMase ceramide S1P/
PLC
DAG
PIP2
IP3
VOC
ROC
ADP-RC
cADPR
NAADP
Ca2+
3
AC cAMP
Metabolism AMP AMPK
NOS NONitrosylation
cGMP
REC
EPTO
RS &
TRA
NSD
UC
ERS
STIM
ULI
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
18
19
17
ATF6, PERK
SCAP, bHLHER
EF
FE
CTO
RS
SE
NS
OR
S
Summary of the major signalling pathways used by cells to regulate cellular processes.Cells have a number of signalling systems that are capable of responding either to external stimuli or to internal stimuli. In the case of the former, externalstimuli acting on cell-surface receptors are coupled to transducers to relay information into the cell using a number of different signalling pathways(Pathways 1–17). Internal stimuli derived from the endoplasmic reticulum (ER) or from metabolism activate signalling pathways independently ofexternal signals (Pathways 18 and 19). All of these pathways generate an internal messenger that then acts through an internal sensor to stimulatethe effectors that bring about different cellular responses. As described in the text, the names of these signalling pathways usually reflect a majorcomponent(s) of the pathway.
7. Nitric oxide (NO)/cyclic GMP signalling pathway.Nitric oxide (NO) synthase (NOS) generates the gasNO that acts both through cyclic GMP and nitrosyla-tion reactions. NO has a particularly important rolein modulating the activity of other pathways such asCa2+ signalling.
8. Redox signalling. Many receptors act throughNADPH oxidase (NOX) to form reactive oxygenspecies, such as the superoxide radical (O2
−•) and hy-drogen peroxide (H2O2), which act to regulate theactivity of specific signalling proteins such as tyrosinephosphatases, transcription factors and ion channels.The O2
−• participates in the nitrosylation reaction inPathway 7.
9. Mitogen-activated protein kinase (MAPK) signalling.This is a classical example of a protein phosphorylationcascade that often begins with Ras and consists of anumber of parallel pathways that function to control
many cellular processes and particularly those relatedto cell proliferation, cell stress and apoptosis.
10. Nuclear factor κB (NF-κB) signalling pathway. Thissignalling system has a multitude of functions. It isparticularly important in initiating inflammatory re-sponses in macrophages and neutrophils as part of aninnate immune response to invading pathogens.
11. Phospholipase D (PLD) signalling pathway. This is alipid-based signalling system that depends upon thehydrolysis of phosphatidylcholine by phospholipaseD (PLD) to give phosphatidic acid (PA), which func-tions as a second messenger to control a variety ofcellular processes.
12. Sphingomyelin signalling pathway. Certain growthfactors and cytokines hydrolyse sphingomyelin togenerate two second messengers that appear to haveopposing effects in the cell. Ceramide seems to pro-mote apoptosis, whereas sphingosine 1-phosphate
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �3
(S1P) stimulates cell proliferation. S1P may also re-lease Ca2+ from internal stores. The action of S1P iscomplicated in that it is released from the cell, whereit can act as a hormone to stimulate external receptors.
13. Janus kinase (JAK)/signal transducer and activator oftranscription (STAT) signalling pathway. This is a fast-track signal transduction pathway for transferring in-formation from cell-surface receptors into the nucleus.The Janus kinases (JAKs) are tyrosine kinases thatphosphorylate the signal transducers and activatorsof transcription (STATs), which carry the informationinto the nucleus.
14. Smad signalling pathway. This pathway mediates theaction of the transforming growth factor β (TGF-β)superfamily, which controls transcription through theSmad transcription factors.
15. Wnt signalling pathways. These pathways play an im-portant role in both development and cell prolifera-tion.
16. Hedgehog signalling pathway. This pathway re-sembles the Wnt signalling pathway in that it alsofunctions to regulate development and cell prolifer-ation. The ligand Hedgehog (Hh) acts through thetranscription factor GLI.
17. Notch signalling pathway. This is a highly conservedsignalling system that functions in developmental pro-cesses related to cell-fate determination particularly instem cells. The notch receptor generates the transcrip-tion factor NICD (Notch intracellular domain).
18. Endoplasmic reticulum (ER) stress signalling. The en-doplasmic reticulum (ER) stress signalling pathwayconcerns the mechanisms used by the ER to transmitinformation to the nucleus about the state of proteinprocessing within the lumen of the ER.
19. AMP signalling pathway. This pathway is regulated byadenosine monophosphate (AMP), which functions asa metabolic messenger to activate an important path-way for the control of cell proliferation.
Not included in Module 2: Figure cell signalling path-ways are some additional signalling pathways that havespecific functions in regulating various aspects of cell meta-bolism, such as sterol sensing and cholesterol biosynthesis,that control the level of cholesterol in cell membranes. An-other example is found in the NAD signalling pathways,where NAD+ functions to regulate a number of cellularprocesses, including energy metabolism, gene transcrip-tion, DNA repair and perhaps ageing as well.
These cassettes then engage a variety of effectors thatare responsible for activating cellular responses. All ofthese mechanisms (Module 1: Figure signal transmissionmechanisms) depend upon information transfer mechan-isms whereby information is transferred along an orderlysequence of events to activate the internal effectors re-sponsible for inducing a great variety of cellular responses.
Cyclic AMP signalling pathwayCyclic AMP is a ubiquitous second messenger that regu-lates a multitude of cellular responses. Cyclic AMP form-
ation usually depends upon the activation of G protein-coupled receptors (GPCRs) that use heterotrimeric G pro-teins to activate the amplifier adenylyl cyclase (AC), whichis a large family of isoforms that differ considerably in boththeir cellular distribution and the way they are activated.There are a number of cyclic AMP signalling effectorssuch as protein kinase A (PKA), the exchange proteins ac-tivated by cyclic AMP (EPACs) that activate the smallGTP-binding protein Rap1 and the cyclic nucleotide-gated channels (CNGCs). These various effectors are thenresponsible for carrying out the cyclic AMP signallingfunctions that include control of metabolism, gene tran-scription and ion channel activity. In many cases, thesefunctions are modulatory in that cyclic AMP often actsto adjust the activity of other signalling pathways andthus has a central role to play in the cross-talk betweensignalling pathways. This modulatory function is particu-larly evident in the case of Ca2+ signalling in both neur-onal and muscle cells. Many of the functions of cyclicAMP depend upon the precise location of PKA relativeto both its upstream and downstream effectors. A familyof A-kinase-anchoring proteins (AKAPs) determines thiscellular localization of PKA as well as a number of othersignalling components. The OFF reactions responsible forremoving cyclic AMP are carried out by either cyclic AMPhydrolysis or by cyclic AMP efflux from the cell.
Cyclic AMP formationThe formation of cyclic AMP can be activated by a verylarge number of cell stimuli, mainly neurotransmittersand hormones (Module 1: Figure stimuli for cyclic AMPsignalling). All these stimuli are detected by G protein-coupled receptors (GPCRs) that use heterotrimeric G pro-teins, which are the transducers that are responsible foreither activating or inhibiting the enzyme adenylyl cy-clase (AC) (Module 2: Figure heterotrimeric G proteinsignalling). In the case of AC stimulation, the externalstimulus binds to the GPCR that functions as a guaninenucleotide exchange factor (GEF) to replace GDP withGTP, which dissociates the heterotrimeric complex intotheir Gβγ and Gα subunits (Module 2: Figure cyclic AMPsignalling). The GαS·GTP complex activates AC, whereasGαi·GTP inhibits AC. The Gα subunits have GTPaseactivity that hydrolyses GTP to GDP, thus terminatingtheir effects on AC. The endogenous GTPase of GαS·GTPis inhibited by cholera toxin and this causes the persistentactivation of the intestinal fluid secretion that results in thesymptoms of cholera.
Adenylyl cyclase (AC)The adenylyl cyclase (AC) family is composed of ten iso-forms: nine of them are membrane-bound (AC1–AC9),while one of them is soluble (AC10) (Module 2: Tableadenylyl cyclases). The domain structure of AC1–AC9 ischaracterized by having two regions where there are sixtransmembrane regions (Module 2: Figure adenylyl cyc-lase structure). The large cytoplasmic domains C1 and C2,which contain the catalytic region, form a heterodimer
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �4
Module 2: Table adenylyl cyclasesRegulatory properties and distribution of adenylyl cyclase.
Adenylyl cyclase(AC) isoform Gα Gβγ Gαi or Gαio
Modulation by Ca2+, calmodulin(CaM), Ca2+/CaM kinase II (CaMKII),protein kinase C (PKC), proteinkinase A (PKA) Tissue distribution
AC1 ↑ ↓ ↓ Gαo ↑ CaM and PKC Brain, adrenal medulla↓ CaMKII
AC2 ↑ ↑ – ↑ PKC Brain, skeletal and cardiac muscle, lungAC3 ↑ ↑ CaM and PKC Brain, olfactory epithelium
↓ CaMKIIAC4 ↑ ↑ ↑ PKC Brain, heart, kidney, liverAC5 ↑ ↓ ↓ ↓ Ca2+ and PKCα Brain, heart, kidney, liver, lung, adrenal
↓ PKAAC6 ↑ ↓ ↓ ↑ PKC Ubiquitous
↓ Ca2+ and PKAAC7 ↑ ↑ ↑ PKC Ubiquitous, high in brainAC8 ↑ ↑ CaM Brain, lung, heart, adrenalAC9 ↑ Brain, skeletal muscleAC10 – – – Activated by HCO3
− Testis
The membrane-bound adenylyl cyclases (AC1–AC9) are widely distributed. They are particularly rich in brain, but are also expressed inmany other cell types. The soluble AC10 is restricted to the testis. The primary regulation of the AC1–AC9 isoforms is exerted throughcomponents of the heterotrimeric G proteins, which are dissociated upon activation of neurotransmitter and hormonal receptors into theirα and βγ subunits. They are all activated by Gαs, but only some of the isoforms are inhibited by Gαi. The Gβγ subunit is able to activatesome isoforms, but inhibits others. The isoforms also differ in the way they are modulated by components of other signalling pathwayssuch as Ca2+ and PKC. Some of the others are inhibited by PKA, which thus sets up a negative-feedback loop whereby cyclic AMP caninhibit its own production. Modified from Table I in Whorton and Sunahara 2003. Reproduced from Handbook of Cell Signaling, Volume 2(edited by R.A. Bradshaw and E.A. Dennis), Whorton, M.R. and Sunahara, R.K., Adenylyl cyclases, pp. 419–426. Copyright (2003), withpermission from Elsevier.
Module 2: Figure adenylyl cyclase structure
TM 1-6 TM 7-12
C1
C1
C1
N N
N N
NH NH
N N
N N
O O
O
O
O
O
O
O
O
O
O
O
O
O
O
O
O O
O
O
O
O
O
P
P
P
P
P
P
OHOH OH
CH HH H
C
2 2
C2
C2
AC 1 - 9
AC 10
ATP
Pyrophosphate
Plasma membrane
Cyclic AMP
1 72 83 94 105 116 12
C2
Domain structure of adenylyl cyclase (AC).The nine membrane-bound adenylyl cyclases (AC1–AC9) have a similar domain structure. The single polypeptide has a tandem repeat of sixtransmembrane domains (TM) with TM1–TM6 in one repeat and TM7–TM12 in the other. Each TM cassette is followed by large cytoplasmic domains(C1 and C2), which contain the catalytic regions that convert ATP into cyclic AMP. As shown in the lower panel, the C1 and C2 domains come togetherto form a heterodimer. The ATP-binding site is located at the interface between these two domains. The soluble AC10 isoform lacks the transmembraneregions, but it retains the C1 and C2 domains that are responsible for catalysis.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �5
Module 2: Figure cyclic AMP signalling
β β
β
γ γγ
α
α α
αS
S i
i
Cyclic AMP
PKA
GTP
GDP
Cholera toxin
GDP
GTP
5’AMP
Cyclic GMP
5’GMP
P
PP
P
PP
P
P
P
P
P
Rap1
EPAC
DAG
CREB
Phosphorylase kinase
Lipase
Ca2+
Ca2++
+
+
+
+ + + ++
+++
+
-- +
Soluble adenylyl cyclase PDE
cGMP PDE
AMPAR
CFTR
PLNER/SRSERCA
RYRCa 1.1
Ca 1.2V
V
3HCO
F-2,6-P phosphatase
2
CNGC
Adenylylcyclase
Stimulatory agonists Inhibitory agonists
InsP3
PLCε
ABCC4
Organization and function of the cyclic AMP signalling pathway.Cyclic AMP is formed both by membrane-bound adenylyl cyclase and by the bicarbonate-sensitive soluble adenylyl cyclase. The former is regulatedby both stimulatory agonists that act through the αS subunit or through inhibitory agonists that act through either the αi or the βγ subunits. Theincrease in cyclic AMP then acts through three different effector systems. It acts through the exchange protein activated by cyclic AMP (EPAC), whichfunctions to activate Rap1. It can open cyclic nucleotide-gated channels (CNGCs). The main action of cyclic AMP is to activate protein kinase A (PKA)to phosphorylate a large number of downstream targets. Some of these drive specific processes such as gene transcription through phosphorylationof cyclic AMP response element-binding protein (CREB), and activation of ion channels [e.g. AMPA receptors and cystic fibrosis transmembraneconductance regulator (CFTR)] and various enzymes that control metabolism [e.g. fructose 2,6-bisphosphate (F-2,6-P2) 2-phosphatase, lipase andphosphorylase kinase]. Other downstream targets are components of other signalling pathways such as the cyclic GMP phosphodiesterase (cGMPPDE), the phospholamban (PLN) that controls the sarco/endo-plasmic reticulum Ca2+-ATPase (SERCA), the ryanodine receptor (RYR) and the Ca2+
channels CaV1.1 and CaV1.2.
and co-operate with each other to convert ATP into cyclicAMP.
Cyclic AMP signalling effectorsCyclic AMP is a highly versatile intracellular messen-ger capable of activating a number of different effectors(Module 2: Figure cyclic AMP signalling). An example ofsuch effectors is the exchange proteins activated by cyc-lic AMP (EPACs), which act to stimulate Rap. Anothergroup of effectors are the cyclic nucleotide-gated channels(CNGCs) (Module 3: Figure Ca2+ entry mechanisms) thatplay a particularly important role in the sensory systemsresponsible for smell and taste. Most of the actions of cyclicAMP are carried out by protein kinase A (PKA).
Protein kinase A (PKA)Many of the actions of cyclic AMP are carried out by pro-tein kinase A (PKA), which phosphorylates specific siteson downstream effector processes (Module 2: Figure cyc-lic AMP signalling). PKA is composed of two regulatory(R) subunits and two catalytic (C) subunits. The way inwhich cyclic AMP activates PKA is to bind to the R sub-units, which then enables the C subunits to phosphorylatea wide range of different substrates [Module 2: Figure pro-tein kinase A (PKA)]. Of the two types of PKA, proteinkinase A (PKA) I is found mainly free in the cytoplasm
and has a high affinity for cyclic AMP, whereas proteinkinase A (PKA) II has a much more precise location by be-ing coupled to the A-kinase-anchoring proteins (AKAPs).The AKAPs are examples of the scaffolding proteins thatfunction in the spatial organization of signalling pathwaysby bringing PKA into contact with its many substrates.The scaffolding function of the AKAPs is carried out byvarious domains such as the conserved PKA-anchoringdomain [yellow region in Module 2: Figure protein kinaseA (PKA)], which is a hydrophobic surface that binds toan extended hydrophobic surface on the N-terminal di-merization region of the R subunits. At the other end ofthe molecule, there are unique targeting domains (blue)that determine the way AKAPs identify and bind specificcellular targets in discrete regions of the cell.
Protein kinase A (PKA) IType I protein kinase A (PKA) associates with the RI iso-forms. As for all isoforms, the R subunits form dimersthrough their N-terminal dimerization/docking domains[yellow bar in Module 2: Figure protein kinase A (PKA)].In addition to holding two R subunits together, this N-terminal region is also responsible for docking to the A-kinase-anchoring proteins (AKAPs), as occurs for PKAII. However, the RI isoforms have a very low affinity forthe AKAPs and are thus mainly soluble. Cyclic AMP acts
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �6
Module 2: Figure protein kinase A (PKA)
C
C
C Tandem cyclic AMP-binding domains
RI
PKA I PKA II
RI
RI RI
RIIRII
RIIRII
C
C
C
CP
P P
Cyclic AMP
Substrate SubstrateSubstrate
Substrate
Substrate
Substrate
Cyclic AMP
AK
AP
AK
AP
Cellular target
Cellular target
The functional organization of protein kinase A (PKA).There are two types of PKA, type I PKA (PKA I) and type II PKA (PKA II), which differ primarily in the type of R subunits that associate with the Csubunits. There are four R subunit isoforms (RIα, RIβ, RIIα and RIIβ), which have somewhat different properties with regard to their affinity for cyclicAMP and their ability to associate with the A-kinase-anchoring proteins (AKAPs). It is these different R subunits that define the properties of the twotypes of PKA.
by binding to the tandem cyclic AMP-binding domains torelease active C subunits that then phosphorylate specificsubstrates. Since the RI subunits have a higher cyclic AMP-binding affinity, PKA I will be able to respond to the lowercyclic AMP concentrations found globally within the bulkcytoplasm.
Protein kinase A (PKA) IIA characteristic feature of Type II protein kinase A (PKA)is that the regulatory dimer is made up of RII subunits.Since this RII subunit has a much higher affinity for theA-kinase-anchoring proteins (AKAPs), PKA II is usuallydocked to this scaffolding protein and thus has a muchmore precise localization to specific cellular targets.
The substrates phosphorylated by cyclic AMP (Mod-ule 2: Figure cyclic AMP signalling) fall into two maingroups: the cyclic AMP substrates that regulate specificcellular processes and the cyclic AMP substrates that arecomponents of other signalling systems.
Cyclic AMP substrates that regulate specific cellularprocesses:
• In neurons, cyclic AMP acts through PKA to phos-phorylate Ser-845 on the AMPA receptor (Module 3:Figure AMPA receptor phosphorylation).
• Cyclic AMP acting through PKA stimulates thefructose-2,6-bisphosphatase component to lower thelevel of fructose 2,6-bisphosphate, which reduces glyco-lysis and promotes gluconeogenesis (Module 2: FigureAMPK control of metabolism).
• In insulin-secreting β-cells, the salt-inducible kinase 2(SIK2) that phosphorylates transducer of regulated cyc-lic AMP response element-binding protein (TORC)is inhibited by a cyclic AMP/PKA-dependent phos-phorylation (Module 7: Figure β-cell signalling).
• PKA phosphorylates the hormone-sensitive lipase(HLS) that initiates the hydrolysis of triacylglycerolto free fatty acids and glycerol in both white fat cells(Module 7: Figure lipolysis and lipogenesis) and inbrown fat cells (Module 7: Figure brown fat cell).
• PKA phosphorylates the phosphorylase kinase thatconverts inactive phosphorylase b into active phos-phorylase a in skeletal muscle (Module 7: Figure skeletalmuscle E-C coupling) and in liver cells (Module 7: Fig-ure glycogenolysis and gluconeogenesis).
• PKA activates the transcription factor cyclic AMP re-sponse element-binding protein (CREB) (Module 4:Figure CREB activation). This activation is a criticalevent in the induction of gluconeogenesis in liver cells(Module 7: Figure liver cell signalling).
• PKA inhibits the salt-inducible kinase 2 (SIK2) that nor-mally acts to phosphorylate TORC2, thereby prevent-ing it from entering the nucleus to facilitate the activityof CREB (Module 7: Figure liver cell signalling).
• PKA phosphorylates inhibitor 1 (I1), which assists theprotein phosphorylation process by inactivating proteinphosphatase 1 (PP1).
• PKA contributes to the translocation and fusion of ves-icles with the apical membrane during the onset of acid
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �7
secretion by parietal cells (Module 7: Figure HCl secre-tion).
• PKA phosphorylates the regulatory (R) domain on thecystic fibrosis transmembrane conductance regulator(CFTR) to enable it to function as an anion channel(Module 3: Figure CFTR channel).
• In the small intestine, PKA phosphorylates the cysticfibrosis transmembrane conductance regulator (CFTR)channel (Module 3: Figure CFTR channel) that is re-sponsible for activating fluid secretion (Module 7: Fig-ure intestinal secretion). Uncontrolled activation of cyc-lic AMP formation by cholera toxin results in cholera.
• In kidney collecting ducts, cyclic AMP acts throughPKA to phosphorylate Ser-256 on the C-terminal cyto-plasmic tail of aquaporin 2 (AQP2), enabling this waterchannel to fuse with the apical membrane to allow wa-ter to enter the cell (Module 7: Figure collecting ductfunction).
• In blood platelets, cyclic AMP activates thephosphorylation of vasodilator-stimulated phos-phoprotein (VASP), which is a member of theEna/vasodilator-stimulated phosphoprotein (VASP)family resulting in a decrease in the actin-dependentprocesses associated with clotting (Module 11: Figureplatelet activation).
Cyclic AMP substrates that are components of othersignalling systems:
• Phosphorylation of the cyclic GMP phosphodiesterasePDE1A by PKA results in a decrease in its sensitivityto Ca2+ activation.
• Entry of Ca2+ through the L-type CaV1.1 channel(Module 3: Figure CaV1.1 L-type channel) and the L-type CaV1.2 channel (Module 3: Figure CaV1.2 L-typechannel) is enhanced through PKA-dependent phos-phorylation.
• PKA-dependent phosphorylation of dopamine- andcyclic AMP-regulated phosphoprotein of apparent mo-lecular mass 32 kDa (DARPP-32) functions as a molecu-lar switch to regulate the activity of protein phosphatase1 (PP1).
• The ryanodine receptor 2 (RYR2) is modulated by phos-phorylation through PKA, which is associated with thecytoplasmic head through an AKAP (Module 3: Figureryanodine receptor structure).
• Sarco/endo-plasmic reticulum Ca2+-ATPase 2a(SERCA2a) increases its Ca2+-pumping activity whenthe inhibitory effect of phospholamban (PLN) isremoved following its phosphorylation by PKA onSer-16 (Module 5: Figure phospholamban mode ofaction).
Salt-inducible kinase 2 (SIK2)The salt-inducible kinase 2 (SIK2) functions to phos-phorylate the transducer of regulated CREB (TORC2),thereby preventing it from entering the nucleus to facilitatethe activity of the transcriptional factor CREB (Module 4:Figure CREB activation). It functions to regulate TORC2in liver cells (Module 7: Figure liver cell signalling) and
in insulin-secreting β-cells (Module 7: Figure β-cell sig-nalling).
Exchange proteins activated by cyclic AMP(EPACs)Other targets for the second messenger cyclic AMP are theexchange proteins activated by cAMP (EPACs). One of thefunctions of EPACs is to activate Rap1, which has manyfunctions in cells, many of which are related to controllingactin dynamics. In addition, the EPAC/Rap pathway canactivate phospholipase Cε (PLCε).
Cyclic AMP signalling functionsThe cyclic AMP signalling pathway functions in the con-trol of a wide range of cellular processes:
• Cyclic AMP suppresses spontaneous Ca2+ oscillationsduring oocyte maturation.
• Cyclic AMP has a potent anti-inflammatory action byinhibiting the activity of macrophages (Module 11: Fig-ure macrophage signalling) and mast cells (Module 11:Figure mast cell inhibitory signalling).
• Melanocortin 4 receptors (MC4Rs) on second-orderneurons use the cyclic AMP signalling pathwayto induce the hypothalamic transcription factorSingle-minded 1 (Sim1) to decrease food intake andweight loss (Module 7: Figure control of food intake).
• Cyclic AMP mediates the action of lipolytic hormonesin white fat cells by stimulating a hormone-sensitivelipase (Module 7: Figure lipolysis and lipogenesis).
• Heat production by brown fat cells is controlled bynoradrenaline acting through cyclic AMP (Module 7:Figure brown fat cells).
• The phosphorylation of DARPP-32 co-ordinates theactivity of the dopamine and glutamate signalling path-ways in medium spiny neurons (Module 10: Figure me-dium spiny neuron signalling).
• Adrenaline-induced glycogenolysis in skeletal musclecells depends upon a cyclic AMP-dependent phos-phorylation of phosphorylase kinase (Module 7: Figureskeletal muscle E-C coupling).
• Activation of the cyclic AMP-dependent transcriptionfactor cyclic AMP response element-binding protein(CREB) contributes to the regulation of glucagon bio-synthesis in glucagon-secreting α-cells (Module 7: Fig-ure α-cell signalling).
Cyclic AMP hydrolysisThere are two OFF reactions of the cyclic AMP sig-nalling pathway, cyclic AMP efflux from the cell and cyclicAMP hydrolysis. The latter is carried out by a family ofphosphodiesterase enzymes that hydrolyse cyclic AMP toAMP (Module 2: Figure cyclic AMP signalling).
Cyclic AMP effluxThere are two OFF reactions for the cyclic AMP sig-nalling pathway, cyclic AMP hydrolysis and cyclic AMPefflux from the cell (Module 2: Figure cyclic AMP sig-nalling). The latter is carried out by ABCC4, which is one
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �8
Module 2: Figure G protein binary switching
GDP
Gprotein
Gprotein
GTP
GDP
Ion channel modulation
GTP
Cyclic AMP signalling
3InsP /DAG signalling
Phosphate
Signalling responsesGEFs
Input from cell surface receptors
Cytoskeletal remodelling
Redox signalling
PtdIns 3-kinase signalling
MAP kinase signalling
PLD signalling
GAPs
GTPase-activating factors
Guanine nucleotide exchange factors
Signal transduction through G protein binary switching.All GTP-binding proteins (G proteins) function through a binary switching mechanism that is driven by the binding of GTP. The G protein is inactivewhen bound to GDP. When this GDP is exchanged for GTP, the G protein is activated and can stimulate a number of different signalling responses.The rapid switching to the active state is facilitated by the guanine nucleotide exchange factors (GEFs) that receive the information coming in fromthe receptors on the cell surface. By contrast, the GTPase-activating proteins (GAPs) accelerate the OFF reaction by enhancing the hydrolysis of GTPto GDP.
of the ATP-binding cassette (ABC) transporters (Mod-ule 3: Table ABC transporters).
GTP-binding proteinsThere are a large number of GTP-binding proteins, whichusually are referred to as G proteins, which play a cent-ral role in cell signalling as molecular switches (Module 2:Figure G protein binary switching). These G proteins arealso GTPases, and the switching is driven by both thebinding of GTP and its hydrolysis to GDP. The G proteinis inactive when bound to GDP, but when the GDP is ex-changed for GTP, the G protein/GTP complex is active andtransfers information down the signalling pathway untilthe endogenous GTPase activity hydrolyses GTP to GDP.The G proteins belong to two groups, the heterotrimericG proteins and the monomeric G proteins, which haveseparate, but overlapping, functions.
Heterotrimeric G proteinsThe heterotrimeric G proteins function as transducers forthe G protein-coupled receptors (GPCRs) that activate anumber of cell signalling pathways (Module 1: Table Gprotein coupled receptors). These G proteins are made upfrom 16 Gα, five Gβ and 11 Gγ genes (Module 2: Tableheterotrimeric G proteins). These different subunits arecharacterized by having various lipid modifications thatserve to insert them into the plasma membrane, where theyare positioned to detect information coming in from the
GPCRs and to relay it to various amplifiers. The Gα sub-units are either palmitoylated or myristoylated near theN-terminus, whereas the Gβγ subunits are prenylated.The heterotrimeric G proteins are extremely versatile sig-nalling elements, and both the Gα subunit and the Gβγ
subunit are able to relay information to downstream com-ponents (Module 2: Figure heterotrimeric G protein sig-nalling).
These heterotrimeric G proteins exist in two states.When the Gα subunit is bound to GDP, it forms a com-plex with Gβγ subunits to form the inactive heterotri-meric complex. When the GPCRs, which are sensitive to awide range of stimuli (Module 1: Figure stimuli for cyclicAMP signalling), are activated, they function as a guan-ine nucleotide exchange factor (GEF) to replace the GDPwith GTP. The binding of GTP not only activates theGα subunit, but also liberates the Gβγ subunit, both ofwhich can activate a range of signalling systems. This signaltransduction process is switched off when the endogen-ous GTPase activity of the Gα subunit hydrolyses GTPto GDP and the Gα/GDP complex recombines with theGβγ subunit to reform the inactive heterotrimeric com-plex. The normal rate of GTPase activity is very low (fourto eight conversions per s), which means that the two sub-units have a long time to find their targets. However, thereare two mechanisms for speeding up the GTPase activity.Firstly, some of the targets can act to accelerate the GT-Pase activity. Secondly, a family of regulators of G proteinsignalling (RGS) proteins function as GTPase-activating
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �9
Module 2: Table heterotrimeric G proteinsThe heterotrimeric proteins are assembled from subunits taken from the G protein α (Gα), G protein β (Gβ) and G protein γ (Gγ) families.Heterotrimeric G protein FunctionG protein α (Gα) subunits
Gαs Stimulate adenylyl cyclaseGαolf Stimulate adenylyl cyclaseGαi1 Inhibit adenylyl cyclaseGαi2 Inhibit adenylyl cyclaseGαi3 Inhibit adenylyl cyclaseGαo1 Inhibit adenylyl cyclaseGαo2 Inhibit adenylyl cyclaseGαt1 Stimulate cyclic GMP phosphodiesterase in rod photoreceptorsGαt2 Stimulate cyclic GMP phosphodiesterase in rod photoreceptorsGz Close K+ channels. Inhibits exocytosis (see Module 10: Figure lactotroph regulation)Gαgust Stimulate phospholipase Cβ (PLCβ) (see Module 7: Figure L cell)Gαq Stimulate phospholipase Cβ (PLCβ)Gα11 Stimulate phospholipase Cβ (PLCβ)Gα14 Stimulate phospholipase Cβ (PLCβ)Gα15 Stimulate phospholipase Cβ (PLCβ)Gα16 Stimulate phospholipase Cβ (PLCβ)Gα12 Stimulate RhoGEFs to activate Rho (Module 2: Figure Rho signalling)Gα13 Stimulate RhoGEFs to activate Rho (Module 2: Figure Rho signalling)
G protein β (Gβ) subunits; β1–β5 These β subunits combine with γ subunits to form βγ dimers that have a number of control functions(for details see Module 2: Figure heterotrimeric G protein signalling)
G protein γ (Gγ) subunits; γ1–γ11 These γ subunits combine with β subunits to form βγ dimers that have a number of control functions(for details see Module 2: Figure heterotrimeric G protein signalling)
proteins (GAPs) that accelerate the Gα subunit GTPaseactivity more than 1000-fold.
There are G protein receptor kinases (GRKs) such asβ-adrenergic receptor kinase 1 (βARK1), which phos-phorylate active receptors to provide binding sites forarrestin that prevent the heterotrimeric proteins frombinding the receptor and this leads to receptor desensit-ization (Module 1: Figure homologous desensitization).
The active Gα/GTP and Gβγ subunits are able to re-lay information to a large number of signalling pathways,which are described in more detail for the different sig-nalling pathways:
• The cyclic AMP signalling pathway (Module 2: Figurecyclic AMP signalling).
• The activation of phospholipase Cβ (PLCβ) in theinositol 1,4,5-trisphosphate (InsP3)/Ca2+ signalling cas-sette (Module 2: Figure PLC structure and function).
• Modulation of the CaV2 family of N-type and P/Q-typechannels (Module 3: Figure CaV2 channel family).
• Activation of the PtdIns 3-kinase signalling pathway(Module 2: Figure PtdIns 3-kinase signalling).
• Activation of phosphodiesterase 6 (PDE6) during pho-totransduction in photoreceptors (Module 10: Figurephototransduction).
• G-αolf functions in sperm motility and chemotaxis.• G-αgust, which is also known as gustudin, functions in
taste cells and in the L cells that detect food componentsin the lumen of the intestine (Module 7: Figure L cells).
Regulators of G protein signalling (RGS)The regulators of G protein signalling (RGS) are a largefamily of approximately 30 proteins that function asGTPase-activating proteins (GAPs) for the heterotrimericG proteins (Module 2: Figure heterotrimeric G protein sig-nalling). The Gα subunit has a low intrinsic GTPase activ-ity and this is greatly increased by the RGS proteins. RGSstructure is defined by an RGS-box region that is respons-
ible for binding to Gα/GTP. However, many of the RGSproteins contain a number of other protein–protein in-teraction domains, such asPDZ, phosphotyrosine-binding(PTB), pleckstrin homology (PH) and phox homology(PX) domains, indicating that, in addition to their GAPactivity, they may have other functions. One of these mightbe the regulation of the G protein-activated inwardly rec-tifying K+ (GIRK) channel.
The function of RGS proteins has been clearly definedin phototransduction, where RGS9 acts to accelerate GTPhydrolysis by Gαt (step 6 in Module 10: Figure photo-transduction).
Monomeric G proteinsThe monomeric GTP-binding proteins (G proteins) be-long to a large family of approximately 150 members. Raswas the founding member, and the family is often referredto as the Ras family of small G proteins. Within this family,it is possible to recognize five subfamilies: Ras, Rho, Rab,Ran and ADP-ribosylation factor (Arf) (Module 2: Tablemonomeric G protein toolkit). The Ran family plays a rolein nuclear transport, whereas the large Rab family func-tions in membrane trafficking. The Ras and Rho family areprimarily involved in cell signalling, where they function asbinary switches to control a number of cell signalling sys-tems. This binary switch is driven by the binding of GTP,which represents the ON reaction, and is followed by thehydrolysis of the GTP by the endogenous GTPase activ-ity. Most attention has focused on a small number of thesesignal transducers, and the following will be described indetail to illustrate their role in cell signalling:
• Ras signalling mechanisms• Rho signalling mechanisms• Rac signalling mechanisms• Cdc42 signalling mechanisms
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �10
Module 2: Figure heterotrimeric G protein signalling
Cyclic AMP
Cyclic AMP
Cyclic AMP
InsP Ca
InsP Ca
Cyclic GMP
DAG
+
Cytoskeleton
Cytoskeleton
Cytoskeleton
DAG
γγ
ββ
α
αα
α
GIRK
Ca 1.2V
AC
AC
AC
3
3
2+
2+
2+
GDP
GDPGTP
GRK
Arrestins
αs
αi
αq
12/13
t
GTP
PI 3-K
Ca
∆V
PIP3
PLC
G protein
Rho A
Cdc42
Rac
PDE6
PLC
GPCR
Stimulus
G responsesα
G responsesβγ
RGSproteins
P
Heterotrimeric G proteins function as transducers to activate many signalling pathways.External stimuli that bind to G protein-coupled receptors (GPCRs) act as guanine nucleotide exchange factors (GEFs) for the heterotrimeric G proteins.When the GDP on the Gα subunit is replaced with GTP, the complex dissociates into Gα/GTP and Gβγ subunits that are then capable of activatingor inhibiting a wide range of signalling systems. Most of the actions are stimulatory, but some are inhibitory, as illustrated by the yellow arrows. Theregulators of G protein signalling (RGS) proteins function as GTPase-activating proteins (GAPs) to facilitate the GTPase activity of the Gα subunit,which is the OFF reaction that terminates signalling. G protein receptor kinase (GRK) phosphorylates active receptors and provides binding sites forarrestin that result in receptor desensitization by preventing the heterotrimeric G proteins from binding the receptor.
Ras signalling mechanismsRas is a small (21 kDa) GTPase that plays a central roleas a signal transducer for a number of signalling sys-tems. It was first identified as a transducer for the tyr-osine kinase-linked receptors (Module 1: Figure stimulifor enzyme-linked receptors), where it functions to re-lay information to the mitogen-activated protein kinase(MAPK) signalling pathway. It is now known that ac-tivated Ras is able to relay information to a number ofother signalling pathways. Like other G proteins (Mod-ule 2: Figure G protein binary switching), Ras functions asa binary switch that is activated by binding GTP. Thereare thus two critical aspects to Ras action: how is theswitch controlled and how is information relayed out todifferent signalling pathways (Module 2: Figure Ras sig-nalling)? With regard to the first question, the ON reac-tion of the binary switch is controlled by a number of Rasguanine nucleotide exchange factors (RasGEFs) such asSon-of-sevenless (SoS), Ras guanine nucleotide release-in-ducing factors (RasGRFs) and Ras guanine nucleotidereleasing proteins (RasGRPs) (Module 2: Table mono-meric G protein toolkit). This conversion of Ras·GDP intoRas·GTP is inhibited by the tumour suppressor proteinmerlin. Neurofibromatosis type 2 is caused by mutationsin merlin.
There are a number of Ras GTPase-activating proteins(RasGAPs) that accelerate the OFF reaction by speed-
ing up the hydrolysis of GTP to GDP by Ras. Examplesof such RasGAPs include p120 RasGAP, neurofibromin,SynGAP, Ca2+-promoted Ras inactivator (CAPRI) andRas GTPase-activating-like (RASAL). The function ofsome of these RasGAPs such as CAPRI and RASAL areattracting considerable attention because they are Ca2+-sensitive proteins that rapidly translocate to the mem-brane through their C2 domains whenever there is an in-crease in the level of Ca2+. SynGAP, which is expressedmainly in brain, is also activated by Ca2+ following itsphosphorylation by Ca2+/calmodulin-dependent proteinkinase II (CaMKII). The role of Ca2+ in modulating theactivity of both the RasGEFs and RasGAPs indicates animportant feedback interaction between the Ca2+ and Rassignalling pathways.
The Ras signalling mechanism relays information to anumber of signalling targets:
• It activates Raf to initiate the MAPK signalling pathway(Module 2: Figure ERK signalling).
• It activates phospholipase Cε (PLCε) (Module 2: FigurePLC structure and function).
• It activates the PtdIns 3-kinase signalling pathway.• Ras can activate a family of RalGEFs such as Ral-
GDP dissociation stimulator that then activates the Ralproteins (RalA and RalB) (Module 2: Figure Ras sig-nalling). One of the functions of RalA is to stimulate
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �11
Module 2: Figure Ras signalling
2+
2+
2+
2+
GDP
Ras RasGTP
GDPGTP
DAGNMDAR
Glutamate Growthfactors
RASALNEUROFIBROMIN
CAPRI SynGAP
CaMKII
p120Ras GAP
PI 3-K
Ral-GDS PLD1
Ca
Ca
Ca
Ca
PIP3
ERK1/2Raf
PLCε
Ral
TK
SOS
TK
PTKR
GAPs+
+
+
+ +
+
Ras responses
RasGRFRasGRP
InsP Ca
DAG
32+
CaM
Function of the monomeric Ras G protein in cell signal transduction.Ras plays a role as a signal transducer to relay information from various external stimuli to a range of different Ras-dependent responses. When boundto GDP, Ras is inactive. Cell stimuli act through different guanine nucleotide exchange factors (GEFs) such as Son-of-sevenless (SoS), RasGRF andRasGRP to facilitate an exchange of GDP for GTP to create the activated Ras/GTP complex that can relay information through a number of signallingpathways. The action of Ras is terminated by a variety of GTPase-activating proteins (GAPs) that accelerate the ability of Ras to hydrolyse GTP backto GDP.
the phospholipase D signalling pathway (Module 2: Fig-ure PLD signalling).
Son-of-sevenless (SoS)Son-of-sevenless (SoS) is a classical Ras guanine nucle-otide exchange factor (RasGEF). Its mode of action isevident in the way the platelet-derived growth factor re-ceptor (PDGFR) is linked to the mitogen-activated pro-tein kinase (MAPK) signalling pathway (Module 1: Fig-ure PDGFR activation). SoS binds to the Src homology 3(SH3)-containing adaptor growth factor receptor-boundprotein 2 (Grb2), which is attached to the phosphotyr-osine residues of the activated receptor. Once it is associ-ated with the receptor, SoS comes into contact with Rasand can begin to facilitate the exchange of GDP for GTP,thus creating the active Ras/GTP complex that begins tostimulate the MAPK signalling pathway (for further de-tails see Module 2: Figure ERK signalling).
Ras guanine nucleotide release-inducing factors(RasGRFs)The Ras guanine nucleotide release-inducing factors(RasGRFs) and Ras guanine nucleotide releasing proteins(RasGRPs) are also important GEFs that can relay inform-ation to Ras. The RasGRFs, which are strongly expressedin brain, are particularly important for the activation ofthe MAPK signalling pathway in neurons, where they re-spond to activation of N-methyl-d-aspartate (NMDA) re-ceptors. One of the functions of the latter is to gate Ca2+,
which then acts through calmodulin (CaM) to stimulateRasGRF. This is a highly localized signalling event, be-cause the RasGRF is associated with the NMDA receptorand thus responds to the microdomain of Ca2+ near themouth of the channel. This signalling mechanism func-tions in neuronal gene transcription (Module 10: Figureneuronal gene transcription).
Ras guanine nucleotide releasing proteins (RasGRPs)The Ras guanine nucleotide releasing proteins (RasGRPs)are mainly expressed in haematopoietic cells (Module 2:Table monomeric G protein toolkit). These GRPs are alsoknown as CalDAG-GEFs because they are sensitive to di-acylglycerol (DAG) and Ca2+. They contain a C1 domainthat binds DAG and they also have a pair of Ca2+-bindingEF-hands. The sensitivity to both DAG and Ca2+ sug-gests that these RasGRPs may couple phosphoinositidesignalling to the activation of the various pathways thatare linked to Ras (Module 2: Figure Ras signalling).
Rac signalling mechanismsThe Rac signalling pathway has an important role in ac-tivating a number of signalling pathways (Module 2: Fig-ure Rac signalling). Like other G proteins, Rac functionsas a binary switch. It is inactive when bound to GDP,but when this GDP is exchanged for GTP, the Rac/GTPcomplex becomes active. External stimuli can activate thisswitch using different guanine nucleotide exchange factors
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �12
Module 2: Figure Rac signalling
GDP
Rac RacGTP
GDPGTP
NOX
IRS p53 WAVE
ROS signalling
Actinpolymerization
PAK1
Arp2/3
TIAM
PIP3
Vav1
3BP-1
SosCaMKIIP-Rex GEFs
Ca2+
GAPs
PTKR Rac responses
Actin-Profilin Profilin
P
Actin assembly
Cofilin
Cofilin
LIM-K1
GPCR
StimulusStimulus
γβ
TK
TK
Kalirin
Function of the Rac monomeric G protein in signal transduction.Rac is inactive when bound to GDP, but switches into an active form when this GDP is exchanged for GTP. This GTP for GDP exchange is facilitated bya number of guanine nucleotide exchange factors (GEFs) such as Tiam, Kalirin, Vav, SoS and P-Rex, which are sensitive to various messengers suchas Ca2+, PtdIns3,4,5P3 (PIP3) or Gβγ subunits of heterotrimeric G proteins. The activated Rac/GTP then relays information out to different signallingpathways as described in the text. Further details concerning the role of Wiskott–Aldrich syndrome protein (WASP) verprolin homologous (WAVE) areshown in Module 4: Figure actin remodelling.
(GEFs) such as T cell lymphoma invasion and metastasis(Tiam), Kalirin, Vav, SoS and P-Rex. Kalirin functions inthe Ephrin (Eph) receptor signalling pathway, where itlinks the EphB receptor to Rac and actin assembly (Mod-ule 1: Figure Eph receptor signalling). The lipid secondmessenger PtdIns3,4,5P3, which is produced by the PtdIns3-kinase signalling pathway, is particularly effective instimulating many of these GEFs.
The activated Rac/GTP complex has a number ofimportant actions:
• Rac is responsible for mediating the action of the PtdIns3-kinase signalling pathway in stimulating the NADPHoxidase to initiate the reactive oxygen species (ROS)signalling pathway (Module 2: Figure plasma membraneROS formation).
• It stimulates the JNK (c-Jun N-terminal kinase) sig-nalling pathway (Module 2: Figure JNK signalling).
• It promotes actin stability by stimulating p21-activatedkinase (PAK) to phosphorylate LIM kinase, whichphosphorylates cofilin to inhibit its ability to cut actin.
• It promotes actin remodelling by acting on in-sulin receptor substrate p53 (IRSp53), which activatesWiskott-Aldrich syndrome protein (WASP) verprolinhomologous (WAVE) to co-ordinate the activity of theactin-related protein 2/3 complex (Arp2/3 complex) thatinitiates actin polymerization (Module 4: Figure actinremodelling). Such a role is evident in neurons duringspine morphogenesis (Module 10: Figure postsynapticdensity) and during the Ca2+ and synaptic plasticity
changes that occur during long-term potentiation (LTP)(Module 10: Figure Ca2+-induced synaptic plasticity).
Rho signalling mechanismsThe Rho signalling pathway has an important role in reg-ulating a number of systems, in particular the operationof the cytoskeleton. It regulates the function of actin intwo main ways. Firstly, it controls some of the processesthat function in actin assembly and it also operates one ofthe control mechanisms that regulate the myosin II–actininteraction responsible for contraction both in smoothmuscle cells and non-muscle cells (Module 2: Figure Rhosignalling). Like other G proteins, Rho functions as a bin-ary switch. It is inactive when bound to GDP, but whenthis GDP is exchanged for GTP, the Rho/GTP complexbecomes active. External stimuli can activate this switchusing different Rho guanine nucleotide exchange factors(RhoGEFs). There are a large number of such RhoGEFsthat are characterized by having a Dbl homology (DH) do-main. The term Dbl comes from diffuse B-cell lymphoma,which is the cell line where the first RhoGEF was identi-fied. The family of Dbl-containing GEFs has now expan-ded to 69 members, some of which are shown in Mod-ule 2: Table monomeric G protein toolkit. These GEFs arenot necessarily specific for Rho: some are rather promis-cuous and will also function as Rac and Cdc42 GEFs.In the case of Rho, ephexin appears to be specific forcoupling ephrin receptors to Rho (Module 2: Figure Rhosignalling). Also leukaemia-associated RhoGEF (LARG),p115-RhoGEF and PDZ-RhoGEF are specific for
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �13
Module 2: Figure Rho signalling
GDP
Rho RhoGTP
GDPGTP
mDia
PtdIns4,5P
PtdIns4P
ROK
Ephexin
PIP3
2
GAPsDLC1 p190Rho
GAP
Rho responses
Actin-ProfilinProfilin
P
P
P
P
Actin assembly
Contraction
Cofilin
Cofilin
LIM-K1
MLCK
MLC
CaM
MYPT1
MYPT1
PPI
PPI
Myosin II
GPCR
Net1
StimulusStimulus
γβ
PDZ-RhoGEF
LARG
α12/13T
K
TK
PTKR
p115-RhoGEF
Actinpolymerization
PtdIns4P5-kinase
Function of the Rho monomeric G protein in cell signal transduction.Rho is a typical G protein that is activated when GDP is exchanged for GTP. This exchange is facilitated by a number of Rho guanine nucleotide exchangefactors (RhoGEFs). For example, the ephexins mediate the action of protein tyrosine-linked receptors (PTKRs), such as the ephrin receptors, whereasthe G protein-coupled receptors (GPCRs) use the α12/13 subunit of heterotrimeric G proteins to activate leukaemia-associated RhoGEF (LARG),p115-RhoGEF or PDZ-RhoGEF. The activated Rho/GTP complex then activates a number of signalling systems as outlined in the text.
coupling G protein-coupled receptors (GPCRs) to Rhoactivation. Net-1, which appears to be sensitive toPtdIns3,4,5P3 (PIP3), is normally found in the nucleus,but translocates to the cytoplasm, where it functions toactivate RhoA. Another important GEF is Ect2, which isalso found in the nucleus during interphase, but then as-sociates with microtubules during mitosis, where it stim-ulates the activation of Rho cytokinesis (Module 9: Figurecytokinesis).
The activated Rho/GTP complex is able to stimulate anumber of signalling processes, many of which are dir-ected towards remodelling of actin and its contraction.Many of the actions of Rho are carried out through a Rhokinase (ROK). Diaphanous-related forming 1 (Dia1) is oneof the downstream effectors that responds to Rho/GTPby interacting with profilin to increase actin polymeriz-ation.
The following are some examples of the function of Rhosignalling:
• Activation of smooth muscle contraction (Module 7:Figure smooth muscle cell E-C coupling).
• Assembly of the actomyosin contractile ring andactivation of contraction during cytokinesis (Module9: Figure cytokinesis).
• Endothelial cell contraction to open the permeabilitybarrier (Module 7: Figure endothelial cell contraction).
• Assembly of the actin ring in osteoclast podosomes(Module 7: Figure osteoclast podosome).
• Eph receptor-induced growth cone collapse in develop-ing neurons (Module 1: Figure Eph receptor signalling).
• Activation of uropod contraction during neutrophilchemotaxis (Module 11: Figure neutrophil chemotacticsignalling).
• Physical interactions between endothelial cells andthe extracellular matrix (ECM) activate p190RhoGAPthat is responsible for the mechanosensitive control ofVEGF receptor expression.
Cdc42 signalling mechanismsThe Cdc42 signalling mechanism has an important rolein controlling the actin cytoskeleton. It regulates some ofthe processes that function in actin assembly (Module 2:Figure Cdc42 signalling). Like other G proteins, Cdc42functions as a binary switch. It is inactive when boundto GDP, but when this GDP is exchanged for GTP, theCdc42/GTP complex becomes active. External stimuli ac-tivate this switch using different guanine nucleotide ex-change factors (GEFs), such as intersectin-long (ITSN-L) and PAK-interacting exchange factor α (α-Pix). Justhow these Cdc42 RhoGEFs are activated is still not prop-erly understood. In the case of the Ephrin (Eph) receptorsignalling pathway, intersectin is activated by binding tothe EphA receptor (Module 1: Figure Eph receptor sig-nalling). The activated Cdc42/GTP complex then stimu-lates various elements that regulate actin polymerization.It promotes actin stability by stimulating p21-activatedkinase (PAK) to phosphorylate LIM kinase, which phos-phorylates cofilin to inhibit its ability to cut actin. Oneof its main actions is to stimulate the Wiskott-Aldrichsyndrome protein (WASP) that acts on the actin-relatedprotein 2/3 complex (Arp2/3 complex) to initiate actin
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �14
Module 2: Table monomeric G protein toolkitSummary of the monomeric G proteins with their guanine nucleotide exchange factors (GEFs) and GTPase-activating proteins (GAPs).
Monomeric G protein, GEF or GAP CommentsMonomeric G proteins (approximately 150 members)
Ras family (36 members) Contribute to the activation of multiple signalling pathways (Module 2: Figure Rassignalling)
H-RasK-RasN-RasR-RasRalARalBRap1Rap2
Rho family (22 members)Rho Primary function is cytoskeletal remodelling (Module 2: Figure Rho signalling)
RhoARhoDRhoERhoFRhoHRhoVMito RhoRho BTB
Rac Cytoskeletal remodelling and ROS signalling (Module 2: Figure Rac signalling)Rac1Rac2Rac3Cdc42 Primary function is cytoskeletal remodelling (Module 2: Figure Cdc42 signalling)
Rab family A large family of GTPases that function in membrane traffickingRab1–Rab35
Arf familyArf1–Arf6 ADP-ribosylation factors function in membrane trafficking and can activate
phospholipase D (Module 2: Figure PLD signalling)Ran A small GTPase that functions in nucleocytoplasmic transport
Guanine nucleotide exchange factors (GEFs)RasGEFs
SoS1 Son-of-sevenless-1SoS2 Son-of-sevenless-2RasGRF1 Expressed in brain, particularly hippocampusRasGRF2RasGRF3 Expressed in brainRasGRP1 Expressed in T cellsRasGRP2 Expressed in neutrophils and plateletsRasGRP3 Expressed in B cellsRasGRP4 Expressed in mast cells
Rho family GEFsThe Dbl family The 69 family members have a Dbl homology domain (DH)
Dbl Diffuse B-cell lymphomaDbs Dbl’s big sisterEct2 Epithelial-cell transforming gene-2 that functions in cytokinesis(Module 9: Figure
cytokinesis)Ephexin Functions in growth cone collapse mediated by EphA receptors (Module 1: Eph
receptor signalling)Fgd1 Facial genital dysplasiaITSN-L Intersectin-long controls Cdc42 activity during EphB receptor signalling (Module 1:
Eph receptor signalling)Kalirin Functions to activate Rac during the action of Eph receptors on dendritic spine
morphogenesis (Module 1: Figure Eph receptor signalling)LARG Leukaemia-associated RhoGEFNET1 Neuroepithelioma transforming gene 1p115-RhoGEF Links G protein-coupled receptors to RhoPDZ-RhoGEF Links G protein-coupled receptors to RhoP-Rex1 PtdIns3,4,5P3-dependent Rac exchanger 1α-Pix PAK-interacting exchange factor α
β-Pix PAK-interacting exchange factor β
Tiam1 T cell lymphoma invasion and metastasis-1Tiam2 T cell lymphoma invasion and metastasis-2TrioVav1Vav2Vav3
Arf GEFsCytohesin 1Cytohesin 4ARNO/Geα2
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �15
Module 2: Table continuedMonomeric G protein, GEF or GAP CommentsGTPase-activating proteins (GAPs)
Ras GAPsp120RasGAPNeurofibromin This tumour suppressor is lost in the inherited disorder neurofibromatosis type 1SynGAPGAP1mGAP1IP4BP
Ins1,3,4,5P4 binds to this protein, causing it to dissociate from the plasma membraneCAPRI Ca2+-promoted Ras inactivatorRASAL Ras GTPase-activating-likeDAB2IPMiro Mitochondrial Rho-GTPase functions in mitochondrial motility (Module 5: Figure
mitochondrial motility)Rac GAPs
3BP-1 3BP-1 inactivates Rac.GTP by enhancing GTP hydrolysis (Module 2: Figure Racsignalling)
Rho GAPSDLC1 (p122RhoGAP in mice) Deleted in liver cancer 1 is altered in many tumours. It regulates actin formation in
adhesion complexes (Module 6: Figure integrin signalling)p190-RhoGAP Inhibits Rho activity during neutrophil chemotaxis (Module 11: Figure neutrophil
chemotactic signalling)
Since some of the families are very large, only representative members have been included.
Module 2: Figure Cdc42 signalling
GDP
Cdc42 Cdc42GTP
GDPGTP
IRS p53
WASP
PAK4
PIP3
Dbsα-Pix GEFs
?
GAPs
Cdc42 responses
Actin-ProfilinProfilin
P
Actin assembly
Cofilin
Cofilin
LIM-K1
Actinpolymerization
Arp2/3
GPCR
StimulusStimulus
γβ
TK
TK
PTKR
ITSN-L
Function of the Cdc42 monomeric G protein in signal transduction.When bound to GDP, Cdc42 is inactive, but it is activated when the GDP is exchanged for GTP. This exchange is accelerated by guanine nucleotideexchange factors (GEFs), but how these are activated is still somewhat of a mystery. The primary action of the Cdc42/GTP complex is to stimulateactin assembly by inhibiting the action of cofilin, by promoting actin polymerization by acting on actin/profilin and Wiskott–Aldrich syndrome protein(WASP). Further details on the role of WASP are shown in Module 4: Figure actin remodelling.
polymerization (Module 4: Figure actin remodelling). Thisaction is facilitated by Cdc42 acting on profilin through in-sulin receptor substrate p53 (IRSp53) and Mena.
p21-activated kinase (PAK)The p21-activated kinases (PAKs) are some of the majordownstream targets of Rac and Rho (Module 2: FigureRho-regulated kinases). They have been implicated in alarge number of processes such as actin remodelling, cell
cycle control, transcription and apoptosis. There are sixisoforms (PAK1–PAK6) with slightly different functions.Rac acts through LIM kinase 1 (LIMK1) to phosphorylatecofilin (Module 2: Figure Rac signalling). Rho acts throughPAK4 to phosphorylate the same LIMK1 (Module 2: Fig-ure Cdc42 signalling). Another function of PAK1 is tophosphorylate myosin light chain kinase (MLCK), res-ulting in a decrease in the activity of the actin–myosincontractile system.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �16
Module 2: Figure Rho-regulated kinases
Kinase
Kinase
Kinase
Kinase
Kinase
Kinase
PH
PH
PH
PH
CH
CH
CC1
CC1
CRD
CRD
CC
CC
CC2
CC2
3
3
PBD
PBD
PBD
PBD
PBD
PBD
Myotonic dystrophy kinase-related Cdc42-binding kinase (MRCK)
Rho kinase (ROK)
P21-activated kinase (PAK)
LIMK1/2 MLCK
MYPT1
LIMK1/2 FilGAP
Rac
Rho
GTP
GTP
Cdc42
Cdc42
GTP
GTP
Structure of Rho GTPase-regulated kinases.The Rho family exert many of their actions by stimulating protein kinases such as PAK, MRCK and ROK. These kinases exist as homodimers withPAK being arranged in a head-to-tail manner, whereas MRCK and ROK have a parallel alignment. The GTP-bound form of these p21 monomericG proteins bind to a p21-binding domain (PBD). CC, coiled-coil domain: CH, citron homology domain; CRD, cysteine-rich domain; FilGap, filaminGTPase-activating protein; MLCK, myosin light chain kinase; PH, pleckstrin homology domain.
The fragile X mental retardation protein 1 RMRP1),which is mutated in fragile X syndrome (FXS), mayact to inhibit the function of PAK particularly duringactin remodelling in neuronal spines (Module 10: FigureCa2+-dependent synaptic plasticity).
Myotonic dystrophy kinase-related Cdc42-bindingkinase (MRCK)Myotonic dystrophy kinase-related Cdc42-binding kinase(MRCK) has a number of functional domains and be-longs to the same family as Rho kinase (ROK) (Module 2:Figure Rho-regulated kinases). MRCK, which is activatedby Cdc42·GTP, may play a role in actin formation dur-ing the growth of neurites. One of the downstream tar-gets of MRCK is the MYPT1 subunit (Module 5: TablePP1 regulatory and inhibitory subunits and proteins) thatbinds protein phosphatase 1δ (PP1δ) and which dephos-phorylates the myosin light chain (MLC).
Rho kinase (ROK)Rho kinase (ROK) is a serine/threonine kinase that phos-phorylates key regulators of actin and myosin function(Module 2: Figure Rho-regulated kinases). ROK acts tostabilize actin by phosphorylating LIM, which then phos-phorylates cofilin to prevent it from severing actin. Rhoalso acts on diaphanous-related formin protein (Dia),which belongs to a formin-related protein family that in-teracts with profilin to promote actin polymerization.
The other major function of ROK is to control contrac-tion by activating myosin II in smooth muscle cells and
in non-muscle cells. The activity of myosin II is regulatedby the phosphorylation status of the myosin light chain(MLC) that is associated with the myosin head. WhenMLC is dephosphorylated, myosin is inactive, but when itis phosphorylated by the Ca2+-sensitive myosin light chainkinase (MLCK), myosin can begin to interact with actin toinduce contraction. ROK can influence the phosphoryla-tion of MLC by inhibiting the protein phosphatase 1 (PP1)by phosphorylating the scaffolding protein myosin phos-phatase targeting subunit 1 (MYPT1) (Module 5: TablePP1 regulatory, targeting and inhibitory subunits).
Diaphanous-related formin 1 (Dia1)There are two diaphanous-related formin proteins (Dia1and Dia2) that belong to a formin-related family. They con-tain three formin homology domains that are used to bindto various effectors. Dia1 is activated by the Rho signallingmechanism and functions to control actin polymarizationby binding to profilin (Module 2: Figure Rho signalling).Dia1 has also been implicated in the control of poly-cystin 2.
Ca2+ signallingCa2+ signalling is one of the major signalling systems incells (Module 2: Figure cell signalling pathways). It func-tions to regulate many different cellular processes through-out their life history. It triggers new life at the time offertilization. It controls many processes during develop-ment, and once cells have differentiated, it governs the
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �17
Module 2: Figure basic Ca2+ signalling mechanism
Ca2+ Ca2+
Exocytosis
Contraction
Gene transcription
ProliferationFertilization
100 nM 500nM
STIMULUS
Metabolism
µs
ms
sec
hr
min
Timedomain
Hypertrophy
ON REACTIONS
OFF REACTIONS
The basic mechanism of Ca2+ signalling.The concentration of Ca2+ in cells at rest is approximately 100 nM, but this increases to 500 nM or more following a stimulus that activates the Ca2+
ON reactions. When the stimulus is removed, the Ca2+ OFF reactions return the concentration of Ca2+ to its resting level. Ca2+ is a universal signalcapable of activating many different cellular processes operating over a very wide time domain.
activity of most cellular processes, effectively determininghow we metabolize, secrete, move and think. There also isa darker side to its action, because larger than normal elev-ations can cause cell death, either in the controlled mannerof programmed cell death (apoptosis) or in the more cata-strophic necrotic changes that occur during processes suchas stroke or cardiac ischaemia.
The basic mechanism of Ca2+ signalling is relativelysimple in that it depends upon an increase in the intracel-lular concentration of this ion. The Ca2+ concentration islow when cells are at rest, but when an appropriate stimu-lus arrives, there is a sudden elevation, which is responsiblefor a change in cellular activity. However, there are multiplevariations of this relatively simple theme. The versatilityof Ca2+ signalling is achieved by having an extensive Ca2+
toolkit from which a large number of Ca2+ signalling sig-nalsomes are assembled. This large toolkit contains manydifferent components that can be mixed and matched tocreate many different Ca2+ signalling modules. There areCa2+ entry channels, which control the entry of Ca2+ fromthe outside. There are Ca2+ release channels, which controlthe release of Ca2+ from internal stores. The Ca2+ buffersensure that the concentration of Ca2+ remains within itsoperation range and does not rise to levels that can inducecell death. Once Ca2+ has carried out its signalling func-tion, there are Ca2+ pumps and exchangers that removeit from the cytoplasm by either extruding it from the cellor returning it to the internal stores. Ca2+ signalling func-tions are carried out by various Ca2+ sensors and Ca2+
effectors that are responsible for translating Ca2+ signalsinto a change in cellular activity.
Basic mechanism of Ca2+ signallingCells at rest maintain a low intracellular concentrationof Ca2+ (approximately 100 nM), but this increases rap-idly into the micromolar range when cells are stimulated(Module 2: Figure basic Ca2+ signalling mechanism). Thisincrease in intracellular Ca2+ can operate over a very widetime domain (e.g. microseconds to hours) to regulate manydifferent cellular processes. This very wide temporal rangeof Ca2+ signalling is an intrinsic property of the Ca2+ sig-nalling modules.
An important feature of Ca2+ signalling is its dynamicnature, as exemplified by the fact that Ca2+ signals in-variably appear as a brief transient. The rising phase ofeach transient is produced by the ON reactions, whereasthe falling phase depends on the OFF reactions (Module2: Figure Ca2+ signalling dynamics). At any moment, thelevel of Ca2+ is determined by the balance between theCa2+ ON reactions that increase Ca2+ and the Ca2+ OFFreactions that remove it from the cytosol. An import-ant aspect of the ON and OFF reactions is their spa-tial location. An example of this spatial organization isthe ER/mitochondrial Ca2+ shuttle (Module 5: FigureER/mitochondrial shuttle), where events at the ER areclosely linked to those in the mitochondria. Another im-portant spatial aspect is that the ON reactions are oftenclosely associated with the effector systems that respond
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �18
Module 2: Figure Ca2+ signalling dynamics
ER
Mitochondria
Buffers
[Ca ]2+Resting
Activated[Ca ]2+
[ Ca ]2+
Ca 2+
Na +Na +
ON REACTIONS OFF REACTIONS
STIMULUS
+ +
2ndmessengers
SERCA
PMCA
Effectors
The dynamics of Ca2+ signalling.The dynamics of Ca2+ signalling are governed by an interplay between the ON and OFF reactions that control the fluxes of Ca2+ across both theplasma membrane and the internal organelles such as the endoplasmic reticulum (ER) and mitochondria. External stimuli activate the ON reactions,which introduce Ca2+ into the cytoplasm either through channels in the plasma membrane or from internal stores such as the ER. Most cells make useof both sources, but there are examples of cells using either external or internal sources to control specific processes. Most of the Ca2+ that entersthe cytoplasm is adsorbed on to buffers, while a much smaller proportion activates the effectors to stimulate cellular processes. The OFF reactionsremove Ca2+ from the cytoplasm using a combination of mitochondria and different pumping mechanisms. When cells are at rest, these OFF reactionskeep the concentration low, but these are temporarily overwhelmed when external stimuli activate the ON reactions. Sequential activation of the ONand OFF reactions gives rise to the Ca2+ transients (Module 2: Figure Ca2+ transient mechanisms), which are such a characteristic feature of Ca2+
signalling systems. PMCA, plasma membrane Ca2+-ATPase; SERCA, sarco/endo-plasmic reticulum Ca2+-ATPase.
to Ca2+. For example, voltage-operated channels (VOCs)in presynaptic endings are associated with the synapticvesicles, thus producing a highly localized puff of Ca2+ totrigger exocytosis (Module 4: Figure Ca2+-induced mem-brane fusion). Similarly, the type 2 ryanodine receptors(RYR2s) of cardiac cells are lined up close to the contract-ile filaments to ensure that Ca2+ will rapidly stimulatecontraction.
In cases where cells need to be stimulated over a longtime, these transients are repeated at set intervals to set upCa2+ oscillations. These oscillations are part of the spati-otemporal aspects of Ca2+ signalling.
Ca2+ signalling signalsomeCells have access to a large Ca2+ signalling toolkit (Mod-ule 2: Table Ca2+ signalling toolkit). Many of the toolkitcomponents have similar functions (Module 2: Figure Ca2+
signalling toolkit), which represents a generic Ca2+ sig-nalling system. In reality, however, each cell type has aclearly defined subset of toolkit components that will bereferred to as a Ca2+ signalling signalsome. These cell-specific signalsomes are put in place during developmentwhen a process of signalsome expression enables each dif-ferentiating cell to select out those signalling components
it will require to control its particular functions (Mod-ule 8: Figure signalsome expression). There are an enorm-ous number of cell-specific Ca2+ signalsomes (Module2: Figure cell-specific Ca2+ signalsomes). The importantpoint is that each signalsome generates a cell-specific Ca2+
signal with characteristic spatial and temporal properties.A signalsome is defined here as the collection of sig-
nalling components that make up each cell-specific sig-nalling system. The Ca2+ signalsomes of different cell typesoften display recurring themes in the form of Ca2+ sig-nalling modules (Module 2: Figure Ca2+ modules).
Ca2+ signalling modulesThe Ca2+ signalling system in specific cell types is oftennot a single entity, but is made up of distinct modules,which are mixed and matched to produce the cell-specificsystems found in different cell types. Some of the mainmodules used by cells are summarized in Module 2: FigureCa2+ modules:
1. Agonists such as the neurotransmitters glutamateand ATP act directly on receptor-operated channels(ROCs) in the plasma membrane to allow external Ca2+
to enter the cell.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �19
Module 2: Table Ca2+ signalling toolkitThe Ca2+ signalling toolkit.
Component CommentsRECEPTORS AND TRANSDUCERSG protein-coupled receptors (GPCRs) See Module 1: Table G protein-coupled receptors for a list of those
GPCRs that act by stimulating phospholipase Cβ
Tyrosine-kinase-linked receptors See Module 1: Figure tyrosine kinase-linked receptors. (The receptorsshown opposite can activate phospholipase Cγ)
Platelet-derived growth factor receptor (PDGFR) See Module 1: Figure PDGFR activationPDGFRα
PDGFRβ
Epidermal growth factor receptor (EGFR)ERBB1–ERBB4
Vascular endothelial growth factor receptor (VEGFR) See Module 9: Figure VEGF-induced proliferationVEGFR1–VEGFR3
G ProteinsGαq, Gα11, Gα14, Gα15, Gα16, Gβγ These G protein subunits activate phospholipase C (Module 2: Figure
heterotrimeric G protein signalling)Guanine nucleotide exchange factorsRasGRF1 A Ca2+-activated GEF expressed in brain (Module 2: Figure Ras
signalling)Regulators of G protein signalling (RGS)
RGS1, RGS2, RGS4, RGS16Phospholipase C (PLC)
PLCβ1–4, PLCγ1–2, PLCδ1–4, PLCε, PLCζ PLC hydrolyses PtdIns4,5P2 to form InsP3 and DAG (Module 2: FigurePLC structure and function)
ADP-ribosyl cyclase Functions to generate cADPR and NAADP (Module 2: FigurecADPR/NAADP function)
CD38CHANNELSPlasma membrane channels
Voltage-operated channels (VOCs) See Module 3: Table VOC classification for further detailsCaV1.1 (L-type)CaV1.2 (L-type)CaV1.3 (L-type)CaV1.4 (L-type)CaV2.1 (P/Q-type)CaV2.2 (N-type)CaV2.3 (R-type)CaV3.1 (T-type)CaV3.2 (T-type)CaV3.3 (T-type)
Ca2+-sensitive ion channelsCa2+-activated K+ channels See Module 3: Table properties of Ca2+-sensitive K+ channels for
further informationSK (small conductance Ca2+-sensitive channel)IK (intermediate conductance Ca2+-sensitive channel)BK (large conductance Ca2+-sensitive channel)
Ca2+-activated chloride channelHCLCA1 (human chloride channel, Ca2+-activated)
Receptor-operated channels (ROCs) See Module 3: Table receptor-operated channel toolkit for furtherdetails)
Nicotinic acetylcholine receptors5-HT3 5-Hydroxytryptamine receptorAMPA receptors α-Amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptorNMDA receptors N-methyl-D-aspartate receptorP2X receptors
Second messenger-operated channels (SMOCs) Module 3: Figure Ca2+ entry mechanismsArachidonate-regulated Ca2+ channel (IARC)
Cyclic nucleotide-gated channels (CNGs) Module 3: Figure cyclic nucleotide-gated channelsCNGA1–CNGA4, CNGB1, CNGB3
Transient receptor potential (TRP) ion channel family Module 3: Figure TRP channel phylogenyTRPC1–TRPC7 CanonicalTRPV1–TRPV6 VanilloidTRPM1–TRPM8 Melastatin
Polycystins Module 3: Figure polycystin domain structurePC1PC2
Endoplasmic reticulum (ER)/sarcoplasmic reticulum (SR) Ca2+release channelsInsP3 receptors (InsP3Rs) Module 3: Figure InsP3R structure
InsP3R1InsP3R2InsP3R3
Ryanodine receptors (RYRs) Module 3: Figure ryanodine receptor structureRYR1RYR2RYR3
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �20
Module 2: Table continuedComponent CommentsChannel regulators
TriadinJunctinSorcin EF-hand protein that can bind to annexin 7FKBP12 FK506-binding protein 12 kDaFKBP12.6 FK506-binding protein 12.6 kDaPhospholamban Module 5: Figure phospholamban mode of action
Ca2+ BUFFERSCytosolic buffers
Calbindin D-28kCalretininParvalbumin
Endoplasmic reticulum (ER)/sarcoplasmicreticulum (SR) buffers and chaperonesCalnexinCalreticulinCalsequestrinGRP78 Glucose-regulatory protein of 78 kDa. Also known as BiPGRP94 Glucose-regulatory protein of 94 kDa. Also known as endoplasmin
Ca2+ SENSORSEF-hand Ca2+-binding proteins
Calmodulin (CaM) A ubiquitous Ca2+ sensorCalcineurin B (CaNB) A sensor that resembles CaM and controls the function of calcineurin
(Module 4: Figure calcineurin)Troponin C (TnC) The Ca2+ sensor in striated muscleMiro Mitochondrial Rho-GTPase functions in mitochondrial motility (Module
5: Figure mitochondrial motility)DAG kinase α A Ca2+-sensitive enzyme that phosphorylates diacylglycerol (DAG)
(Module 2: Figure InsP3/DAG recycling)Stromal interaction molecule (STIM) Senses Ca2+ in the ER lumen (Module 3: Figure STIM-induced Ca2+
entry)S100 proteins
S100 protein clustered on chromosome 1 S100A1-S100A14 are all clustered on 1q21 (Module 4: Figure S100phylogenetic tree)
S100A1S100A2S100A3S100A4S100A5S100A6S100A7S100A8S100A9S100A10S100A11S100A12S100A13S100A14
S100 protein clustered on other chromosomesS100BS100CS100P
Neuronal Ca2+ sensor proteins (NCS) A family of 14 EF-hand Ca2+-binding proteinsNCS-1 Brain and retinaHippocalcin Restricted to hippocampal neuronsNeurocalcin δ Brain and retinaRecoverin Retina
Visinin-like proteins (VILIPs)VILIP-1 Brain and retinaVILIP-2 RetinaVILIP-3 Retina and cerebellar Purkinje cells
Guanylyl cyclase-activating proteins (GCAPs) Module 10: Figure phototransductionGCAP1 RetinaGCAP2 RetinaGCAP3 Retina
Kv-channel-interacting proteins (KChIPs)KChIP1 BrainKChIP2 BrainKChIP3/downstream regulatory element modulator(DREAM)/calsenilin
This protein has three names which are currently in use
KChIP4 Brain
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �21
Module 2: Table continuedComponent CommentsCa2+-binding proteins (CaBPs)
Caldendrin Brain and retinaL-CaBP1 A long splice variant of caldendrinS-CaBP1 A short splice variant of caldendrinCaBP2 RetinaCaBP3 RetinaCaBP4 Brain and retinaCaBP5 Retina
C2 domain Ca2+-binding proteinsSynaptotagmins The Ca2+ sensors that control exocytosis (Module 4: Figure
Ca2+-induced membrane fusion)Synaptotagmin ISynaptotagmin IISynaptotagmin III
Annexins Ca2+-dependent phospholipid-binding proteins (Module 4: Figureannexin structure)
Annexin A1Annexin A2Annexin A3Annexin A4Annexin A5Annexin A6Annexin A7Annexin A8Annexin A9Annexin A10Annexin A11Annexin A13
Ca2+-SENSITIVE ENZYMES AND PROCESSESCa2+-regulated enzymes
Ca2+-dependent protein kinases (CaMKs)CaMKICaMKIICaMKIIICaMKIVMyosin light chain kinase (MLCK)Phosphorylase kinaseIns1,4,5P3 3-kinase
Proline-rich tyrosine kinase 2 (Pyk2) A Ca2+-sensitive tyrosine kinase that functions in osteoclast podosomes(Module 7: Figure osteoclast podosome)
Lipid kinasehVps34 See Module 9: Figure target of rapamycin signalling
PKC-αPKC-βIPKC-βIIPKC-γ
PhosphodiesterasesCyclic GMP phosphodiesterase (PDE) Some of the phosphodiesterases are sensitive to Ca2+ see Module 5:
Table PDE family propertiesPDE1APDE1BPDE1C
Adenylyl cyclases (ACs) See Module 2: Table adenylyl cyclases for details of theseCa2+-sensitive enzymes
AC-1AC-IIIAC-VIIIAC-VAC-VI
Dual oxidases (DUOX1–2) See Module 2: Table redox signalling componentsNitric oxide synthase (NOS)
Module 2: Figure NO and cyclic GMP signallingEndothelial NOS (eNOS)Neural NOS (nNOS)
Ca2+-activated proteasesCalpain ICalpain II
Transcription factorsNuclear factor of activated T cells (NFAT) Module 4: Figure NFAT activation
NFATc1NFATc2NFATc3NFATc4
Cyclic AMP response element-binding protein (CREB) Module 4: Figure CREB activationDownstream regulatory element modulator (DREAM)CREB-binding protein (CBP)
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �22
Module 2: Table continuedComponent CommentsCa2+ PUMPS AND EXCHANGERSNa+/Ca2+ exchanger (NCX) See Module 5: Table Ca2+ pumping toolkit for further details
NCX1NCX2NCX3
Na+/Ca2+/-K+ exchanger (NCKX)NCKX1NCKX2NCKX3NCKX4
Mitochondrial channels and exchangers Module 5: Figure mitochondrial Ca2+ signallingPermeability transition pore (PMT)Na+/Ca2+ exchangerCa2+ uniporter
Plasma membrane Ca2+-ATPases (PMCAs) See Module 5: Table Ca2+ pumping toolkit for further detailsPMCA1PMCA2PMCA3PMCA4
Sarco/endo-plasmic reticulum Ca2+-ATPases (SERCAs) See Module 5: Table Ca2+ pumping toolkit for further detailsSERCA1SERCA2SERCA3
Secretory-pathway Ca2+-ATPase (SPCA) pumpsSPCA1 Ubiquitous; located in the GolgiSPCA2
The Ca2+ signalling system has a very large toolkit of signalling components. The ways in which the major components are organized aresummarized in Module 2: Figure Ca2+ signalling toolkit. Components from this toolkit can be mixed and matched to create a diversearray of cell-specific Ca2+ signalsomes (Module 2: Figure cell-specific Ca2+ signalsomes) that are capable of delivering Ca2+ signalswith very different spatial and temporal properties.
Module 2: Figure cell-specific Ca2+ signalsomes
SKELETALMUSCLE CELL
CARDIACATRIAL CELL
Ca1NEURON T CELL
Receptors
PLC
Na /Caexchanger
Buffers
Sensors
mGluR1 M1
TCR
Orai1
PLCβ PLCβ PLCγ1Ca 1.1V Ca 1.2V Ca 1.2/ Ca 2.1
Ca 2.2/ NMDARVVV
ET-1R/ α1R AngIIR
PMCAs
SERCAs
RYR1
+ 2+
RYR2InsP R2
RYR2InsP R2 33 InsP R13
PMCA1a,1c,1d
PMCA1c,1d,2a
PMCA1a, 2a3a
PMCA4b
SERCA1a, 1b SERCA2a SECA2b, 3 SERCA2b, 3
NCX NCX1 NCX1, 3 –
Parvalbumin ParvalbuminCalbindin 28K
Troponin cCalmodulin
Troponin cCalmodulin
Calmodulin Calmodulin
Entrychannels
Releasechannels
Some examples of cell-specific Ca2+ signalling signalsomes.The four cell types represented here generate Ca2+ signals with very different spatial and temporal properties. For example, the skeletal musclesignalsome selects out those components specialized to deliver rapid pulses of Ca2+ to activate contraction, whereas the T cell signalsome hasdifferent components that generate the much slower repetitive pulses of Ca2+ necessary to stimulate cell proliferation.
2. Second messengers such as diacylglycerol (DAG),cyclic AMP, cyclic GMP and arachidonic acid act-ing from the cytoplasmic side open second mes-senger-operated channels (SMOCs) in the plasmamembrane.
3. Membrane depolarization (�V) activates voltage-operated channels (VOCs) in the plasma membrane toallow a rapid influx of external Ca2+.
4. Membrane depolarization (�V) activates a specificVOC isoform, the CaV1.1 L-type channel, that
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �23
Module 2 : Figure Ca2+ signalling toolkit
Stimulus
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
CaBP
Cel
lula
r res
pons
es
VOC ROC SOC
Sensors
Effectors
ER/SR
Mitochondrion
InsP3
InsP3R
Receptors
G proteins
PLC
PMCA SERCA
NCX
RYR
SMOC
Uniporter
Summary of the major components that contribute to the Ca2+ signalling signalsome.A generic Ca2+ signalling signalsome is made up from components of the extensive Ca2+ signalling toolkit. The duplication of the individualcomponents represents the fact that there are numerous isoforms that further enhance the diversity of the Ca2+ signalling systems. The yellow arrowsillustrate the ON reactions that introduce Ca2+ into the cell, and the blue arrows depict the OFF reactions that pump Ca2+ either out of the cell or backinto the endoplasmic reticulum (ER). During its passage through the cytoplasm, Ca2+ resides temporarily on the buffers or within the mitochondria.To carry out its signalling function, Ca2+ binds to sensors that then employ a range of effectors to stimulate cellular processes. These differentcomponents are mixed and matched to construct Ca2+ modules (Module 2: Figure Ca2+ modules) that are then assembled to produce cell-specificsignalsomes. CaBP, Ca2+-binding protein; InsP3R, inositol 1,4,5-trisphosphate receptor; NCX, Na+/Ca2+ exchanger; PLC, phospholipase C; PMCA,plasma membrane Ca2+-ATPase; ROC, receptor-operated channel; RYR, ryanodine receptor; SERCA, sarco/endo-plasmic reticulum Ca2+-ATPase;SMOC, second messenger-operated channel; SOC, store-operated channel; VOC, voltage-operated channel.
activates the ryanodine receptor 1 (RYR1) in skeletalmuscle through a direct conformational-couplingmechanism.
5. Membrane depolarization (�V) activates voltage-operated channels (VOCs) in the plasma membrane toallow a rapid influx of external Ca2+ (see Module 3)to provide a Ca2+ trigger that then activates theryanodine receptor 2 (RYR2) to release Ca2+ storedin the sarcoplasmic reticulum (SR) through a process ofCa2+-induced Ca2+ release (CICR). This mechanism isfound in cardiac muscle and neurons.
6. Agonists acting on cell-surface receptors generate inos-itol 1,4,5-trisphosphate (InsP3) (Module 2: Figure InsP3
and DAG formation), which then diffuses into the cellto activate the InsP3 receptor (InsP3R) to release Ca2+
from the ER.
Ca2+ ON reactionsIn response to external stimuli, channels in the plasmamembrane or ER are opened, and Ca2+ flows into thecytoplasm to bring about the elevation of cytosolic Ca2+
responsible for cell activation (Module 2: Figure Ca2+ sig-nalling dynamics). During these ON reactions, the cellemploys a variety of both Ca2+ entry channels and Ca2+
release channels to create Ca2+ signals with markedly dif-ferent spatial and temporal properties.
The entry of Ca2+ across the plasma membrane is carriedout by many different channels whose names indicate howthey are opened:
• Voltage-operated channels (VOCs)• Agonist-operated channels (AOCs)• Receptor-operated channels (ROCs)• Second messenger-operated channels (SMOCs)• Store-operated channels (SOCs)
Release of Ca2+ from internal stores is carried out bydifferent types of channels and control mechanisms:
• Ryanodine receptors (RYRs)• InsP3 receptors (InsP3Rs)• NAADP control of Ca2+ release
One of the major problems in Ca2+ signalling has beento determine how stimuli arriving at the cell surface gainaccess to these internal stores. Two main mechanisms havebeen identified:
1. Conformational coupling through a protein–proteininteraction. This is a very fast mechanism that depends
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �24
Module 2: Figure Ca2+ modules
InsP3InsP R3
RYR1
RYR2
AGONIST
AGONIST
ROC
SMOC
VOC
Ca
Ca 2+
Ca 2+
Ca 2+
Ca 2+
Ca 2+
Ca 2+
2+
∆V
∆V
∆V
2
3
4
5
6
1
Second messengers
SR
SR
ER
Ca2+ signalling modules.Some of the main Ca2+ signalling modules that cells employ to generate Ca2+ signals are depicted. Ca2+ is either derived from the external mediumor released from internal stores such as the sarcoplasmic reticulum (SR) in muscle or the endoplasmic reticulum (ER) in other cell types. The ionchannels responsible for gating Ca2+ in these different modules are explained in the text.
upon a sensor in the plasma membrane interacting dir-ectly with an internal release channel. The receptor onthe cell surface is the CaV1.1 L-type channel (a voltagesensor), which is coupled to the type 1 ryanodinereceptor (RYR1)(Ca2+ module 4 in Module 2: Fig-ure Ca2+ modules). Information is transferred througha process of conformational coupling. This mechan-ism is restricted to skeletal muscle (Module 7: Figureskeletal muscle E-C coupling) and perhaps also to someneurons.
2. Generation of diffusible second messengers. Activa-tion of receptors or channels on the cell surface gen-erate second messengers that then diffuse into the cellto activate release channels. One of the most signific-
ant Ca2+-mobilizing messengers is Ca2+ itself, whichis a potent activator of the two main internal re-lease channels, the ryanodine receptors (RYRs) and theinositol 1,4,5-trisphosphate receptors (InsP3Rs). ThisCa2+-induced Ca2+ release (CICR) mechanism has theunique property of being autocatalytic and plays acentral role in generating those Ca2+ signals that ap-pear as regenerative Ca2+ waves (Module 2: FigureCa2+-induced Ca2+ release).
Another classical process for releasing internal Ca2+
is the inositol 1,4,5-trisphosphate (InsP3)/Ca2+ signallingcassette (Ca2+ module 6 in Module 2: Figure Ca2+
modules). Other Ca2+-mobilizing messengers have been
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �25
Module 2: Figure Ca2+-induced Ca2+ release
+
+
+++ +
+
+
+
Intracellular calcium wave
InsP3
Ca2+
Ca2+
Stimulus
Spark
CICR
CICR
CICR
Puff
VOC ROC
R
ER
RR
Blink
I II+ + + ++
+
The role of Ca2+-induced Ca2+ release (CICR) in mobilizing Ca2+ from internal stores.The Ca2+-sensitive channels are the ryanodine receptors (R) and the InsP3 receptors (I) located on the endoplasmic reticulum (ER). CICR has twoimportant functions. Firstly, it provides a mechanism for transferring information from the plasma membrane to these internal release channels. Theprimary mechanism is based on the voltage-operated channels (VOCs) that open in response to membrane depolarization to allow a small amountof Ca2+ to enter, which then diffuses into the cell to activate either R or I. The other important function of CICR is to link together these intracellularchannels so that the Ca2+ being released from one channel diffuses across to neighbouring channels that are excited to release further Ca2+, therebysetting up regenerative waves (yellow arrows).
described such as sphingosine 1-phosphate (S1P), cyclicADP ribose (cADPR) and nicotinic acid–adenine dinuc-leotide phosphate (NAADP).
The channels responsible for these Ca2+ ON reactionsusually have powerful inactivation mechanisms that rap-idly curtail the entry or release processes to prevent the cellbeing swamped with Ca2+, which can result in cell stressand apoptosis. Once the ON reactions have been curtailed,the Ca 2+ OFF reactions rapidly take over to return theactivated level of Ca2+ back to its resting level.
Ca2+-induced Ca2+ release (CICR)A process of Ca2+-induced Ca2+ release (CICR) plays acentral role in the way Ca2+ signals are generated. Thispositive-feedback mechanism whereby Ca2+ triggers itsown release has two important functions in cells. It en-ables Ca2+ entering across the plasma membrane to func-tion as a messenger to release Ca2+ from the internal store(Module 2: Figure Ca2+-induced Ca2+ release). This func-tion of CICR was first described in cardiac cells, where theCaV1.2 L-type channel provides an influx of trigger Ca2+
that then diffuses into the cell to activate the ryanodine re-ceptor 2 (RYR2) (Ca2+ module 5 in Module 2: Figure Ca2+
modules). A similar interaction is particularly evident forneuronal Ca2+ entry and release channels.
The other main function of CICR is to set upintracellular Ca2+ waves where an elevated level of Ca2+
in one region of the cell (the initiation site) propagatesthroughout the rest of the cytoplasm as a regenerative Ca2+
wave. Waves can progress by recruiting either RYRs or in-ositol 1,4,5-trisphosphate receptors (InsP3Rs) (Module 2:Figure Ca2+-induced Ca2+ release). These Ca2+ waves aremade up of elementary Ca2+ events such as the sparks andpuffs produced by the RYRs and InsP3Rs respectively. Itis these unitary events that are used to generate the regen-erative waves that make up global Ca2+ signals.
Ca2+ OFF reactionsCells use a variety of mechanisms to remove Ca2+ fromthe cytoplasm (Module 2: Figure Ca2+ signalling dynam-ics). The introduction of Ca2+ into the cell during theCa2+ ON reactions usually occurs for a relatively briefperiod during which there is a rapid increase in the in-tracellular concentration of Ca2+. In fact, the increase infree Ca2+ that can be measured in the cytoplasm usingaequorin or fluorescent indicators is a very small propor-tion of the total amount of Ca2+ that enters during theON reactions. Much of this Ca2+ is rapidly bound by thecytosolic buffers or is taken up by the mitochondria. Asthe Ca2+ concentration returns to its resting level, Ca2+
leaves the buffers and the mitochondria and is returned tothe ER or is pumped out of the cell resulting in a brief Ca2+
transient.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �26
Module 2: Figure Ca2+ transient mechanisms
ONReactions
OFFReactions
CALCIUM TRANSIENT
Ca entry
Ca buffers
Ca pumps (SERCA/PMCA)
Ca release
2+
2+
2+
2++
2+
Mitochondria
Na /Ca exchanger
The sequence of ON and OFF reactions during the generation of a typical Ca2+ transient.The rising phase of the Ca2+ spike results from the activation of Ca2+ entry and release mechanisms (yellow bars), which are then terminated byinactivation processes. Once the ON reactions have been inactivated, a series of OFF reactions operate in a sequential manner to restore Ca2+ toits resting level (blue bars). During the rising phase of the Ca2+ transient, large amounts of Ca2+ are rapidly bound to the Ca2+ buffers (calbindinD-28k and parvalbumin) and are taken up by the mitochondria. The mitochondria and cytosolic buffers help to shape the Ca2+ signal by reducing theimpact of the ON reactions. In effect, they enable the cell to generate very fast transients without running the risk of being overwhelmed by Ca2+.
The recovery process thus depends upon a complex in-terplay between cytosolic Ca2+ buffers, mitochondria andCa2+ pumps and exchangers on the internal stores and onthe plasma membrane (Module 2: Figure Ca2+ signallingdynamics). These Ca2+ OFF reactions operate at differentstages during the recovery phase of a typical Ca2+ spike.The buffers and mitochondria operate early, and the Ca2+
pumps and exchangers are responsible for restoring thestatus quo by pumping Ca2+ out of the cell or back into theER. These pumps and exchangers operate at different timesduring the recovery process. The Na+/Ca2+ exchangershave low affinities for Ca2+, but have very high capacitiesand this enables them to function at the beginning of the re-covery process to rapidly remove large quantities of Ca2+.On the other hand, the plasma membrane Ca2+-ATPase(PMCA) and sarco/endo-plasmic reticulum Ca2+-ATPase(SERCA) pumps have lower capacities, but their higher af-finities mean that they can complete the recovery processand can continue to pump at lower Ca2+ levels, thus en-abling them to maintain the internal stores and the restinglevel (Module 5: Figure Ca2+ uptake and extrusion).
Some of these OFF reactions interact with each otherduring the recovery period, and this is particularly evidentin the case of the ER/mitochondrial Ca2+ shuttle (Module5: Figure ER/mitochondrial shuttle). When Ca2+ is re-leased from the ER, Ca2+ is rapidly taken up by the mito-chondria and this is then released slowly back to the ERonce Ca2+ has returned to the resting level.
In addition to its role of returning the level of Ca2+ to itsresting level following a stimulus, the Ca2+ OFF reactionsare in constant operation to maintain the resting level of
Ca2+ within a fairly narrow range. This calciostat is a dy-namic system in that the OFF reactions are operating con-tinuously to reverse the constant basal rate of Ca2+ entrythrough various ‘Ca2+ leak pathways’ that remain poorlydefined. There is increasing evidence that presenilins mayfunction as such a leak channel and this has been incorpor-ated into the calcium hypothesis of Alzheimer’s disease (seestep 10 in Module 12: Figure amyloid cascade hypothesis).The latter can result in an elevation of cytosolic Ca2+ if theOFF reactions are inhibited. For example, agents such asthapsigargin, which inhibit the SERCA pumps on the ER,will result in an increase in Ca2+ concentration. Indeed,the rate at which the Ca2+ concentration rises followingpump inhibition provides a measure of the activity of theseleak pathways.
In pathological situations, the lack of oxygen reducesthe supply of energy, thus compromising the function ofthe OFF reactions, resulting in the rise of Ca2+ that isso damaging during stroke or cardiac ischaemia. Undernormal circumstances, however, the fully energized OFFreactions can rapidly reduce the pulse of Ca2+ introducedby the ON mechanisms, thus generating a brief Ca2+ tran-sient (Module 2: Figure Ca2+ transient mechanisms). Suchbrief pulses of Ca2+ are a characteristic feature of the Ca2+
signalling pathway and contribute to the spatiotemporalaspects of Ca 2+ signalling.
Ca2+ buffersCells express a large number of Ca2+-binding proteins,which fall into two main groups: Ca2+ sensors andCa2+ buffers. The Ca2+ sensors respond to changes in
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �27
intracellular Ca2+ by activating some downstream effectorprocess. In a sense, all proteins capable of binding Ca2+ willact as a buffer, and this applies to the sensors. However, theconcentration of these sensors is usually rather low, so theyhave little buffering capacity. The role of Ca2+ bufferingis carried out by the other major group of Ca2+-bindingproteins. Cells have a number of buffers (Module 2: TableCa2+ signalling toolkit) that are capable of binding to Ca2+
both in the cytoplasm and within the lumen of the endo-plasmic reticulum (ER). The sole function of these Ca2+
buffers is to bind Ca2+, and this has an important func-tion in shaping both the spatial and temporal properties ofCa2+ signals.
The major cytosolic buffers in cells are parvalbumin(PV) and calbindin D-28k (CB). The major buffers that op-erate within the lumen of the ER are calsequestrin (CSQ) inthe sarcoplasmic reticulum of muscle cells and calreticulin(CRT) in the ER of non-muscle cells. The latter is unusualin that it functions both as a cytosolic and a luminal buffer.The role of calsequestrin is discussed elsewhere and herewe concentrate on the function of the cytosolic buffers.The latter have subtly different Ca2+-binding propertiesand are expressed in cells in differing combinations andconcentrations to create Ca2+ signals that are tailored tocarry out different functions. For example, neurons suchas Purkinje cells express large amounts of PV and CB.As a consequence, Purkinje cells have a large endogenousCa2+ buffering capacity, e.g. their buffers bind approxim-ately 2000 Ca2+ ions for each free ion. Lower capacitiesof 50–100:1 are found in other cells. Motor neurons havea very low buffering capacity and consequently have largeCa2+ signals in both the soma and dendrites during normalphysiological responses, and this makes them particularlysusceptible to neurodegeneration. Buffer concentration isone of the important parameters in determining buffer ca-pacity. The other key parameters include affinity for Ca2+
and other metal ions, the kinetics of Ca2+ binding, releaseand mobility.
Alterations in Ca2+ buffers have been linked toschizophrenia.
Parvalbumin (PV)Parvalbumin (PV) is a slow-onset buffer. It has relativelylow on and off rates, which means that it cannot respondto the rapid onset of most Ca2+ signals. However, PVcan soak up Ca2+ once the signal has appeared and thusinfluences the rate at which Ca2+ recovers. It is stronglyexpressed in skeletal muscle, where it plays an importantrole in facilitating the rate of relaxation. In PV−/ − mice,there is a slowing in the recovery of the Ca2+ transient.Some of the effects of removing PV are compensated forby an increase in the volume of mitochondria that hasa similar ability to PV of removing free Ca2+ from thecytoplasm during the recovery phase.
Calbindin D-28k (CB)Calbindin D-28k (CB) is one of the major cytosolic buf-fers, particularly in neurons. It is a fast buffer that can havea major effect on both the spatial and temporal propertiesof Ca2+ transients. CB thus plays a major role in restricting
the size of the elementary Ca2+ events that form aroundCa2+ channels. CB is thus of central importance for theability of neurons to create the highly localized Ca2+ eventsthat occur in spines. The existence of these buffers has en-abled neurons to increase the numbers of their synapticconnections, and this neuronal miniaturization greatly en-hances the signal processing capacity of the brain. Somesupport for such a notion emerged from the finding of a re-markable compensatory mechanism whereby the volumeof the spines increases markedly in neurons when thesebuffers are knocked out. In effect, the lack of these buffersreduces the capacity of neurons to miniaturize the Ca2+
component of their signalling systems.The ability of CB to buffer internal Ca2+ may play an
important role in facilitating Ca2+ reabsorption by the kid-ney tubule (Module 7: Figure kidney Ca2+ reabsorption).In the case of Ca2+ reabsorption by the intestine, the fluxof Ca2+ is facilitated by calbindin D-9k (Module 7: Figureintestinal Ca2+ reabsorption).
An important aspect of the calcium hypothesis ofAlzheimer’s disease is that there is a decrease in the ex-pression of CB, which increases the sensitivity of neuronsto the enhanced Ca2+ signals that arise during the onset ofAlzheimer’s disease (Module 12: Figure amyloid cascadehypothesis).
Calreticulin (CRT)Calreticulin (CRT) is a low-affinity Ca2+-binding proteinthat is located within the lumen of the endoplasmic retic-ulum (ER). It has also been detected in the nucleus andcytoplasm and it may also be secreted into the extracellu-lar environment during periods of cell stress. Its primarylocation, however, is within the lumen of the ER, whereit functions both as a chaperone protein and as the majorER Ca2+ buffer. CRT has three main domains; there isa globular N-domain of unknown function followed bya proline-rich P-domain, which has sequences unique toCRT and its homologous proteins calnexin and calmegin.Finally, there is a highly acidic C-terminal domain, whichhas a large number of low-affinity Ca2+-binding sites cap-able of binding 20–30 mol of Ca2+/mol of protein. ThisC-terminal region terminates in the KDEL sequence re-sponsible for retaining CRT in the lumen of the ER.
The chaperone function of CRT is carried out inconjunction with a related chaperone calnexin. A cal-nexin/calreticulin cycle ensures the correct folding andsubunit assembly of glycoproteins and is thus essentialfor protein trafficking and secretion. The proper foldingand assembly of proteins is very dependent on a constantlevel of Ca2+ within the ER lumen and the chaperones.The ability of CRT to bind large amounts of Ca2+ en-ables it to function as an ER Ca2+ buffer to help main-tain this constancy of ER Ca2+. In addition to functioningas a passive buffer, CRT may play a more direct role inmaintaining homoeostatic control of their working envir-onment by modulating the activity of the Ca2+ channelsand pumps. When the Ca2+ level within the lumen gets toohigh, CRT and calnexin inhibit the sarco/endo-plasmic re-ticulum Ca2+-ATPase (SERCA) pump and, when Ca2+ isreleased, this inhibition is relieved and the SERCA pumps
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �28
Module 2: Figure temporal aspects
ONReaction
OFFReaction
CALCIUM TRANSIENT
Periodic stimulation Continuous stimulation
Fertilization, liver metabolism, fluid secretion, smooth muscle contraction, c ell proliferation,
Skeletal and cardiac cell contraction, synaptictransmission
Temporal aspects of Ca2+ signalling.In almost every example where Ca2+ is used as a signal, it is presented as a brief transient, which is the digital signal used to set up complex temporalpatterns of Ca2+ signalling. In some cells, these pulses are produced on demand in that they are generated in response to periodic stimulation (bluearrows) as occurs in muscle contraction where a brief burst of Ca2+ activates the contractile machinery, which then recovers when the Ca2+ signal isremoved. Likewise, the release of neurotransmitters from nerve terminals is triggered by a brief localized pulse of Ca2+. In many other tissues, whichreceive a continuous stimulation over a prolonged period (blue bar), the Ca2+ signal is again presented as brief spikes that are produced rhythmicallyto give highly regular Ca2+ oscillations whose frequencies vary with the level of cell stimulation.
can restore Ca2+ to its normal level. In addition to its roleas an ER buffer, CRT may play an active role in ensuringthat the Ca2+ concentration within the ER lumen is main-tained at the optimal level for protein folding to occur.
Ca2+ signalling functionThe function of Ca2+ as an intracellular second messen-ger is carried out by a combination of Ca2+ sensors andCa2+ effectors. The major sensors are the EF-hand pro-teins troponin C (TnC), calmodulin (CaM), neuronal Ca2+
sensor proteins (NCS) and the S100 proteins. However,there are a number of other sensors (Module 2: Table Ca2+
signalling toolkit).These sensors are then responsible for relaying inform-
ation through a range of effectors:
• Ca2+-sensitive K+ channels• Ca2+-sensitive Cl− channels (CLCAs)• Ca2+/calmodulin-dependent protein kinases (CaMKs)• Calcineurin• Phosphorylase kinase• Myosin light chain kinase (MLCK)• Ca2+-promoted Ras inactivator (CAPRI) (Module 2:
Figure Ras signalling)
Spatiotemporal aspects of Ca2+ signallingThe use of Ca2+ as a universal signal for cell regulation issomewhat paradoxical because this ion can be very toxic
to cells if its level remains high for a prolonged period.Such toxicity is avoided by presenting Ca2+ signals in apulsatile manner (Module 2: Figure temporal aspects). Inaddition to this temporal aspect, the Ca2+ signal is alsohighly organized in space. The elementary and global as-pects of Ca2+ signalling greatly increase the versatility ofthe Ca2+ signalling system in that it can act either locallyor globally. For example, muscle contraction is activatedby a global elevation in Ca2+, whereas the release of neur-otransmitters results from a minute punctate pulse of Ca2+
delivered directly to the docked vesicle by a Ca2+ sensortightly associated with exocytotic machinery (Module 4:Figure Ca2+-induced membrane fusion). In between thesetwo extremes, there are many variations in the way theCa2+ signal is presented to cells. Perhaps the most dra-matic are the Ca2+ waves that initiate at fixed localitiesand then process through the cytoplasm in a regenerativemanner through the process of CICR (Module 2: FigureCa2+-induced Ca2+ release).
These spatiotemporal aspects greatly enhance the ver-satility of Ca2+ signalling, thus providing the flexibility toregulate so many cellular processes.
Cyclic ADP-ribose (cADPR) signallingCyclic ADP-ribose (cADPR) is one of the messengers as-sociated with the NAD+ signalling pathways. cADPR hasattracted considerable attention as a putative messenger to
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �29
Module 2: Figure cADPR metabolism
The generation and metabolism of cyclic ADP-ribose (cADPR).The same enzyme ADP-ribosyl cyclase is responsible for both forming and degrading cADPR. The precursor NAD+ is converted into cADPR by thecyclase activity, whereas the hydrolase component inactivates cADPR by hydrolysing it to ADP-ribose (ADPR). Reproduced from Trends Cell Biol.,Vol. 4, Galione, A. and White, A., Ca2+ release induced by cyclic ADP-ribose, pp. 431–436. Copyright (1994), with permission from Elsevier; seeGalione and White (1994).
regulate the Ca2+ signalling pathway. cADPR generationand metabolism is unusual in that it is carried out by thesame enzyme that possesses both synthase and hydrolaseactivity. The cADPR control of Ca2+ release is somewhatcontroversial, as its precise mode of action is still unclear.A cADPR working hypothesis has been put forward toprovide a framework to understand some of the currentinformation on how this messenger appears to operate. Arelationship between cADPR and cell regulation has beenestablished in a number of different cell types.
cADPR working hypothesisThere is sufficient evidence to take seriously the possibilitythat cADPR functions to regulate Ca2+ signalling. Whatseems to be in question is exactly how it functions. Thisworking hypothesis has two main components:
• Firstly, the generation of cADPR is closely coupled tocellular metabolism, as described in cADPR generationand metabolism (Module 2: Figure cADPR/NAADPfunction). The idea is that cADPR may function as ametabolic messenger responsible for relaying informa-tion about the state of metabolism to various systems inthe cell, especially those that require a heavy expendit-ure of energy. An example would be Ca2+ signallingand the downstream elements regulated by Ca2+. Whenenergy is abundant, an increase in cADPR will set thestage for Ca2+ signalling to occur.
• Secondly, the cADPR control of Ca2+ releasemay occur indirectly through an activation of thesarco/endo-plasmic reticulum Ca2+-ATPase (SERCA)pump to increase the uptake of Ca2+ into the endoplas-mic reticulum/sarcoplasmic reticulum. This increasedloading of the internal store will serve to sensitize re-lease channels such as the RYRs, thus leading to an in-crease in Ca2+ signalling. This messenger should thusbe considered as a modulator rather than a mediator ofCa2+ signalling.
cADPR generation and metabolismThe generation and metabolism of cADPR are describedtogether because both processes are carried out by thesame enzyme: the ADP-ribosyl cyclase (Module 2: Fig-ure cADPR metabolism). The hydrolysis of cADPR pro-duces ADPR, which has been implicated as a messengerregulating melastatin-related transient receptor potential2 (TRPM2), which is a Ca2+ channel in the plasma mem-brane (Module 2: Figure cADPR/NAADP function). Thisis a highly versatile enzyme. In addition to synthesizingand metabolizing cADPR, it is also responsible for syn-thesizing NAADP. In mammals, this bifunctional enzymeappears to be the lymphocyte antigen CD38, which is ex-pressed widely and is located both in the plasma membraneand at internal sites. With regard to the former location, itsenzymatic region is located on the outside. This is unusualbecause its substrate and its site of action are on the inside
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �30
Module 2: Figure cADPR/NAADP function
cADPR
NAADP
NAAD
NAD
NADP
SH
Ca2+
2+
Ca
Ca
2+
2+
Ca2+
CaCa2+
CaCa2+
CaCa2+
Ca
+
+
+
+
+
+
+RYR
TCP1/2
ADPR
SERCA ADP ribosyl cyclase
Cellularmetabolism
ATPNADH
AGONISTS
Lysosome-related organelle
?
Phosphatase
HH
++
VOC TRPM2
Synthesis and mode of action of cADPR and NAADP.The enzyme ADP-ribosyl cyclase is a bifunctional enzyme that has a synthase (S) component that synthesizes cADPR and NAADP from theprecursors NAD+ and NADP respectively, but it also has a hydrolase (H) activity that converts cADPR into ADPR. This hydrolase is sensitive tometabolism because it is inhibited by either ATP or NADH. The cADPR may act by stimulating the sarco/endo-plasmic reticulum Ca2+-ATPase(SERCA) pump to increase the uptake of Ca2+ into the endoplasmic reticulum. NAADP acts on a channel to release Ca2+ from a lysosome-relatedorganelle.
of the cell. It has been suggested that the enzyme mightuse NAD+ derived from dying cells at sites of infectionsto generate cADPR, which is then transported into thecell.
The intracellular enzyme is more likely to be the onethat functions in most cells. Just how the cyclase is activ-ated is still unclear. One suggestion is that the formation ofcADPR and NAADP is sensitive to cellular metabolism(Module 2: Figure cADPR/NAADP function). In otherwords, cADPR and NAADP might be metabolic messen-gers that are capable of relaying information about thestate of cellular metabolism to the Ca2+ signalling path-ways. Such a notion is supported by the fact that cADPRmetabolism by the hydrolase is inhibited by either ATPor NADH. Another suggestion is that it might be ac-tivated by agonists acting through cell-surface receptors,but the coupling mechanism remains to be established(Module 2: Figure cADPR/NAADP function). This ab-sence of a coupling mechanism might be explained by thecADPR working hypothesis if the external agonist en-hanced cADPR formation indirectly by first increasingcellular metabolism. Such a mechanism could explain theability of β-adrenergic agents to increase cADPR levels inheart. Likewise, the glucose-dependent increase in cADPRin β-cells can be directly linked to the metabolism of gluc-ose, with the resulting increase in ATP acting to reduce
the hydrolase activity (Module 2: Figure cADPR/NAADPfunction).
In CD38−/ − mice, there is a decrease in the amountof oxytocin (OT) released from hypothalamic neurons.Such mice show defects in maternal nurturing and in socialbehaviour.
cADPR control of Ca2+ releaseOne of the major uncertainties about cADPR is its mode ofaction in controlling the release of Ca2+. There have beensuggestions that it is a Ca2+-mobilizing second messengerthat acts by stimulating the ryanodine receptors (RYRs) torelease Ca2+. However, direct evidence for this assertion isnot particularly convincing. Early single channel record-ings seemed to provide such evidence by showing thatcADPR could open RYRs in lipid membranes, but theseobservations were challenged on the basis that the cADPRwas acting through the ATP-binding site. When cADPR isinjected into cells, it usually fails to release Ca2+, but aftera period of time, it can begin to enhance the sensitivity ofthe RYRs. An example of such an effect is shown in heartcells, where there is a gradual increase in spark frequencyfollowing addition of cADPR (Module 2: Figure cADPRaction in heart cells). This observation on cardiac cellsforms the basis of the second part of the cADPR workinghypothesis, which argues that cADPR acts indirectly as a
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �31
Module 2: Figure cADPR action in heart cells
Effect of cADPR on spark frequency in heart cells.When permeabilized rat ventricular myocytes were treated with 5 µM cADPR, there was a gradual increase in spark frequency that recovered tothe resting level when it was washed off (wo). There was no change in spark amplitude. This increase in spark frequency was shown to dependupon sensitization of the RYRs due to an increase in the luminal load of Ca2+ resulting from an increase in the activity of the sarco/endo-plasmicreticulum Ca2+-ATPase (SERCA) pump. Reproduced from Lukyanenko, V., Gyorke, I., Wiesner, T.F. and Gyorke, S. (2001) Potentiation of Ca2+ releaseby cADP-ribose in the heart is mediated by enhanced SR Ca2+ uptake into the sarcoplasmic reticulum. Circ. Res. 89:614–622, with permission fromLippincott Williams & Wilkins (http://lww.com); see Lukyanenko et al. 2001.
modulator of Ca2+ release (Module 2: Figure cADPR/NAADP function) by stimulating the sarco/endo-plasmicreticulum Ca2+-ATPase (SERCA) pump to increase theload of Ca2+ within the lumen of the store. This increasein luminal Ca2+ then sensitizes the RYRs so that they eitherbegin to open spontaneously to give Ca2+ sparks (Module2: Figure cADPR action in heart cells) or begin to producelarger Ca2+ transients when activated by a pulse of trig-ger Ca2+ as occurs in neurons (Module 2: Figure cADPRaction in neurons).
cADPR and cell regulationA role for cADPR has been implicated in a number ofdifferent cell types:
• Insulin-secreting β-cells (Module 7: Figure β-cell sig-nalling)
• Hypothalamic neurons: in CD38−/ − mice, there is adecrease in the amount of oxytocin (OT) released fromhypothalamic neurons. Such mice show defects in ma-ternal nurturing and in social behaviour.
Nicotinic acid–adenine dinucleotidephosphate (NAADP) signallingNAADP is one of the messengers associated with theNAD signalling pathways. NAADP has attracted consid-erable attention as a Ca2+-mobilizing second messenger.NAADP generation and metabolism is unusual in that itis closely allied to that of cADPR in that the same en-zyme synthesizes both messengers. The NAADP controlof Ca2+ release depends upon this messenger acting on astore that is different from that controlled by the inositol1,4,5-trisphosphate receptors (InsP3Rs) or the ryanodinereceptors (RYRs). A relationship between NAADP andcell regulation seems to depend upon its interaction withthe other Ca2+-mobilizing messenger systems.
NAADP generation and metabolismNAADP (Module 2: Figure NAADP structure) is an-other member of the NAD signalling pathways. Its syn-thesis from NADP is closely linked to that of cADPR inthat they both share the same enzyme: the ADP-ribosylcyclase (Module 2: Figure cADPR/NAADP function).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �32
Module 2: Figure cADPR action in neurons
cADPR potentiates Ca2+ transients induced by action potentials in sympathetic neurons.When sympathetic neurons are stimulated for 10 ms at 20 Hz, there is a distinct transient made up of Ca2+ entering through voltage-operated channels(VOCs) together with release of Ca2+ from internal stores through a process of Ca2+-induced Ca2+ release (CICR). The latter component is quitevariable and depends upon the sensitivity of the ryanodine receptors (RYRs). When the neurons are perfused with cyclic ADP-ribose (cADPR), there isa large increase in the amplitude of the transient (compare responses a and b in panel A). Note how the increase developed slowly over the recordingperiod. There was no augmentation when the neurons were perfused with control solution (panel B). Reproduced from Neuron, Vol. 12, Hua, S.-Y.,Tokimasa, T., Takasawa, S., Furuya, S., Nohmi, M., Okamoto, H. and Kuba, K., Cyclic ADP-ribose modulates Ca2+ release channels for activation byphysiological Ca2+ entry in bullfrog sympathetic neurons, pp. 1073–1709. Copyright (1994), with permission from Elsevier; see Hua et al. 1994.
Module 2: Figure NAADP structure
The structure of NAADP.The space-filling model on the right illustrates the molecular organization of the different components of NAADP. Reproduced from Curr. Biol., Vol. 13,Lee, H.C., Ca2+ signalling: NAADP ascends as a new messenger, pp. R186–R188. Copyright (2003), with permission from Elsevier; see Lee 2003.
This enzyme uses NAD+ to make cADPR through a cyc-lization reaction, or it can use NADP as a substrate to pro-duce NAADP through a base-exchange reaction duringwhich the nicotinamide group is exchanged for nicotinicacid.
Unlike cADPR, which is hydrolysed by the same en-zyme, NAADP is degraded to NAAD by phosphatasessuch as alkaline phosphatase.
NAADP control of Ca2+ releaseNAADP functions as a Ca2+-mobilizing messenger torelease Ca2+ from an internal store (Module 2: Fig-ure cADPR/NAADP function). This NAADP-sensitivestore, which is distinct from the endoplasmic retic-ulum/sarcoplasmic reticulum store that is regulated by theinositol 1,4,5-trisphosphate receptors (InsP3Rs) and theryanodine receptors (RYRs), is still being defined. On the
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �33
basis of studies carried out in sea urchin eggs, this novelstore appears to reside in a lysosome-related organelle. Thereceptors for NAADP appear to be a family of two-porechannels (TCP), with TCP1 being located on the endo-somal membranes, whereas TCP2 is on lysosomal mem-branes.
NAADP and cell regulationThe amount of Ca2+ released by NAADP is relativelysmall and is unlikely to have a direct role in Ca2+ sig-nalling. However, there are suggestions that it may actindirectly to trigger the release of Ca2+ by the other re-lease channels [ryanodine receptors (RYRs) or the inositol1,4,5-trisphosphate receptors (InsP3Rs) (Module 2: FigurecADPR/NAADP function)].
NAADP has been implicated in the control of insulin re-lease by β-cells (Module 7: Figure β-cell signalling) and inthe control of secretion by pancreatic acinar cells (Module7: Figure control of pancreatic secretion).
ADP-ribosyl cyclaseADP-ribosyl cyclase is the enzyme responsible for syn-thesizing the Ca2+ mobilizing messengers cADPR andNAADP. During cADPR generation and metabolism ituses NAD+ to make cADPR through a cyclization reac-tion (Module 2: Figure cADPR/NAADP function). Thesame enzyme is also responsible for the hydrolysis ofcADPR to ADPR. This enzyme can also use NADP as asubstrate to produce NAADP through a base-exchange re-action during which the nicotinamide group is exchangedfor nicotinic acid.
In mammals, this bifunctional enzyme appears to be thelymphocyte antigen CD38, which is expressed widely andis located both in the plasma membrane and at internalsites.
Phosphoinositide signallingSignalling through the phosphoinositide lipids is com-plex because there are a number of signalling cassettesassociated with both the synthesis and hydrolysis of thephosphoinositides. Phosphoinositide metabolism can bedivided into two main parts. Firstly, there is inositol lipidmetabolism, which describes the way in which the parentmolecule PtdIns is metabolized to form a number of lipidintermediates, some of which are key elements in differ-ent signalling cassettes. Secondly, there is inositol phos-phate metabolism, which is a complex pathway respons-ible for metabolizing soluble inositol phosphates. Someof the inositol phosphates generated by this metabolismhave been implicated as messengers operating within themultipurpose inositol polyphosphate signalling pathway.
PtdIns4,5P2 is of particular interest because it isthe precursor used to generate the second messen-gers inositol 1,4,5-triphosphate (InsP3) and diacylglycerol(DAG) that function in the inositol 1,4,5-trisphosphate(InsP3)/Ca2+ signalling cassette and the diacylglycerol(DAG)/protein kinase C (PKC) signalling cassette re-spectively. PtdIns4,5P2 is also a precursor that is metabol-ized to PtdIns3,4P2 and PtdIns3,4,5P3, which function as
second messengers to activate protein kinases in the PtdIns3-kinase signalling pathway. In addition to being a pre-cursor for these various signalling pathways, PtdIns4,5P2
can also function as a messenger for the PtdIns4,5P2 sig-nalling cassette, where localized synthesis or hydrolysis ofthis lipid functions to regulate a number of cellular sys-tems such as the actin cytoskeleton, membrane trafficking,exocytosis, ion channels and exchangers.
Alterations in the activity of this phosphoinositidelipid signalling pathway have been implicated in variousdiseases such as manic-depressive illness, Lowe’s oculo-cerebrorenal (OCRL) syndrome and Cowden’s disease.
Phosphoinositide metabolismPhosphoinositide metabolism can be separated into twomain components (Module 2: Figure phosphoinositidemetabolism):
• Inositol lipid metabolism concerns the pathways re-sponsible for converting PtdIns into a variety ofphosphoinositide lipid signalling molecules.
• Inositol phosphate metabolism is responsible for creat-ing a large array of inositol phosphates, some of whichfunction in the multipurpose inositol polyphosphatesignalling pathway. This metabolic pathway also formsthe inositol that is used to resynthesize the lipid pre-cursor PtdIns.
These two pathways are connected by the processes oflipid hydrolysis and lipid synthesis. The hydrolysis occurswhen external signals activate phospholipase C (PLC) tohydrolyse PtdIns4,5P2 to form the second messengers in-ositol 1,4,5-trisphosphate (Ins1,4,5P3) and diacylglycerol(DAG). The Ins1,4,5P3 is one of the major inputs into theinositol phosphate metabolic pathway that produces theinositol required for lipid synthesis.
This complex phosphoinositide metabolic network par-ticipates in a number of highly versatile signalling cas-settes (Module 2: Figure phosphoinositide signalling sys-tems):
• Inositol 1,4,5-trisphosphate (InsP3)/Ca2+ signalling cas-sette
• Diacylglycerol (DAG)/protein kinase C (PKC) sig-nalling cassette
• PtdIns 3-kinase signalling• PtdIns4,5P2 signalling cassette• Multipurpose inositol polyphosphate signalling path-
way
Inositol lipid metabolismThe different phosphoinositide signalling pathways arederived from the parent molecule phosphatidylinositol(PtdIns) (Module 2: Figure PtdIns structure). Unlike theother phospholipids found in cellular membranes, PtdInsis unique in that the free hydroxy groups at the 3-, 4- and5-positions on the inositol ring can be phosphorylatedfurther to create a family of phosphoinositides (Module 2:Figure phosphoinositide metabolism). One of the moreimportant lipids involved in phosphoinositide signallingis PtdIns4,5P2, which is a nodal point for a number of
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �34
Module 2: Figure phosphoinositide metabolism
PtdIns PtdIns4P PtdIns3,4P PtdIns3,4,5P 3
PLC
PtdIns3P PtdIns3,5P 2
2
PtdIns5P PtdIns4,5P
Lipid resynthesis
Inositol synthase InositolInositol phosphate metabolism
Inositol lipid metabolism
Ins1,4,5PDAG
3
4
5
6
6
7
8
8
10
2
3G-6-P
1 11
2
9
Summary of phosphoinositide metabolism.The metabolism of phosphoinositides is separated into inositol lipid metabolism and inositol phosphate metabolism. In the former case, the parentmolecule phosphatidylinositol (PtdIns) (Module 2: Figure PtdIns structure) undergoes a series of phosphorylation reactions to create various poly-phosphoinositides. One of these is PtdIns4,5P2, which is of key importance as it can function both as a messenger and as a precursor for the formationof other phosphoinositide lipid signalling molecules such as PtdIns3,4,5P3. In addition, it can be hydrolysed to form the second messengers inositol1,4,5-trisphosphate (Ins1,4,5P3) and diacylglycerol (DAG). The Ins1,4,5P3 is the major input into a complex pathway of inositol phosphate metabolism(Module 2: Figure inositol phosphate metabolism). One of the outputs of this metabolism is inositol that is used for the resynthesis of PtdIns. Cellsalso have a 1-l-myo-inositol-1-phosphate synthase that can convert glucose 6-phosphate (G-6-P) into inositol.
Module 2: Figure PtdIns structure
Phosphatidylinositol (PtdIns)
O2 2
OH
OHOH OH
OH
C C
1
43 5
OO O
H HHC CC P
Molecular structure of PtdIns.The backbone of the molecule consists of a glycerol moiety. Two of the carbons carry fatty acids that insert into the hydrophobic domain of themembrane, whereas the remaining carbon is attached to a phosphate group that is linked to an inositol ring that projects out into the cytosol. Thehydroxy groups on the 3-, 4- and 5-positions can be phosphorylated further (see Module 2: Figure phosphoinositide metabolism).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �35
Module 2: Figure PI 3-K family
Helical
Accessorydomain
Helical PX
C2
C2
CaM-binding domain
C2 C2
Ras
Regulatory subunits
Catalytic subunits
Catalytic subunit
Catalytic subunit
Regulatory subunits
Regulatory subunit
Ras
Class IA
Class IB
Class II
Class III
Kinase
Kinase
Kinase
p110α, p110 p110β, δ
p110γ
PIK3C3 (hVPS34)
PP
SH2
SH2
SH2
SH2
BCRp85 p85α, β
p55 p50 pα, α, 55γ
p110 binding
p110 binding
SH3 P P
P
p101 p84, Homolgy I Homolgy II
Kinasep150Heat domains Wd40 domains
PIP32PIP
HelicalC2Ras
Catalytic subunits
p85 Kinase
PI3KC2αPI3KC2βPI3KC2γ
The regulatory and catalytic subunits of the PtdIns 3-kinase family.The family of PtdIns 3-kinase is divided into three classes. The kinase domain (shown in yellow) phosphorylates PtdIns4,5P2 (PIP2) to form the lipidmessenger PtdIns3,4,5P3 (PIP3). The domain structures of the regulatory and catalytic subunits are described in the text. Information for this figurewas taken from Hawkins et al. 2006.
signalling systems (Module 2: Figure phosphoinositide sig-nalling systems).
The metabolism of PtdIns is carried out by a collectionof lipid kinases and lipid phosphatases. The numbers belowrefer to the different reactions shown in Module 2: Figurephosphoinositide metabolism:
1. The Class III PtdIns 3-kinase (PtdIns 3-K) addsa phosphate to the 3-position of PtdIns to formPtdIns3P.
2. PtdIns 4-kinase (PtdIns 4-K) adds a phosphate tothe 4-position of PtdIns to form PtdIns4P. It canalso be formed by removal of the 3-phosphate fromPtdIns3,4P2 by phosphatase and tensin homologuedeleted on chromosome 10 (PTEN).
3. PtdInsP kinase III, which has not been fully charac-terized, adds a phosphate to the 5-position of PtdInsto form PtdIns5P.
4. The Type 1 PtdIns4P 5-kinase adds a phosphate tothe 5-position of PtdIns3P to form PtdIns3,5P2. Thelevel of PtdIns3,5P2 changes in cells following osmoticstress.
5. Types I and II PtdIns 3-kinase (PtdIns 3-K) adda phosphate to the 3-position of PtdIns4P to formPtdIns3,4P2. PtdIns3,4P2 has been suggested to func-tion as a messenger operating within the plasma mem-brane, where it serves to recruit and activate proteinkinases such as protein kinase B (PKB). PtdIns3,4P2
can also be formed by other enzymes (see Steps 6and 7).
6. PtdInsP kinase II adds a phosphate to the 4-positionof PtdIns3P to form PtdIns3,4P2. The same enzymecan add a phosphate to the 4-position of PtdIns5P toform PtdIns4,5P2.
7. Inositol polyphosphate 5-phosphatase such as Src ho-mology 2 (SH2) domain-containing inositol phos-phatase (SHIP) removes a phosphate from the 5-position of PtdIns 3,4,5P3 to form PtdIns3,4P2.
8. A PtdIns4P 5-kinase (PtdIns4P 5-K) adds a phosphateto the 5-position of PtdIns4P to form PtdIns4,5P2.The latter can also be formed by Steps 6 and 9.PtdIns4P5-K can also phosphorylate PtdIns3,4P2 toform PtdIns3,4,5P3.
9. Phosphatase and tensin homologue deleted on chro-mosome 10 (PTEN) removes a phosphate from the3-position of PtdIns3,4,5P3.
10. A Class I PtdIns 3-kinase adds a phosphate to the3-position of PtdIns4,5P2 to form PtdIns3,4,5P3.PtdIns3,4,5P3, which functions as one of the secondmessengers in the PtdIns 3-kinase signalling pathway,can also be formed by the addition of a phosphateto the 5-position of PtdIns3,4P2 by the PtdIns4P5-kinase (PtdIns4P 5-K) (i.e. the same enzyme usedfor Step 8).
11. The myotubularins remove the 3-phosphate fromPtdIns3P.
PtdIns 3-kinase (PtdIns 3-K)The PtdIns 3-kinases (PtdIns 3-Ks) are a family of en-zymes that have been classified into three classes (Mod-ule 2: Figure PI 3-K family).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �36
Module 2: Figure phosphoinositide signalling systems
Inositol phosphate metabolism
InsP DAG
The multipurposeinositol polyphosphatesignalling cassette
The diacylglycerol (DAG)/ protein kinase C signalling cassette
The InsP / Ca signalling cassette
2+
PtdIns PtdIns4P PtdIns3,4,5P3
3
3
PtdIns4,5P2
The PtdIns 3-kinasesignalling cassette
The PtdIns4,5Psignalling cassette
2
PtdIns metabolism spawns a variety of signalling cassettes.The metabolism of PtdIns creates a number of lipid and inositol phosphate derivatives that operate a variety of signalling cassettes. PtdIns4,5P2 is anodal point for a number of signalling cassettes. It is the precursor that is hydrolysed to generate the second messengers inositol 1,4,5-trisphosphate(InsP3) and diacylglycerol (DAG). It is phosphorylated further to form the lipid second messenger PtdIns3,4,5P3. The localized turnover of PtdIns4,5P2also has a signalling role within the PtdIns4,5P2 signalling cassette that regulates a variety of processes such as the cytoskeleton, control of theprocesses of membrane trafficking and exocytosis and the regulation of ion channels and exchangers. Finally, the InsP3 enters a complex pathwayof inositol phosphate metabolism that generates components of the multipurpose inositol polyphosphate signalling cassette.
Class I PtdIns 3-kinasesThe primary function of the Class I PtdIns 3-kinases is tophosphorylate PtdIns4,5P2 to form the lipid second mes-senger PtdIns3,4,5P3 (Module 2: Figure PtdIns 3-kinasesignalling). These Class I enzymes are divided into twogroups.
The Class IA enzymes are heterodimers that are formedfrom five regulatory subunits (p85α, p85β, p55α, p50α andp55γ), which are typical adaptor proteins. The p85 regu-latory subunit contains an Src homology 3 (SH3) domain,a breakpoint-cluster-region homology (BH) domain, twoproline-rich regions (P) on either side of the BH domainand two Src homology (SH2) domains that are separatedby a p110-binding domain, enabling them to interact with ap85-binding domain located on the three catalytic subunits(p110α, p110β and p110δ). The different binding domainson the regulatory subunits enhance the versatility of theClass IA enzyme by enabling the catalytic subunit to beactivated by interacting with a variety of signalling mo-lecules:
• The Src homology (SH2) domains of the regulatory sub-unit enable the enzyme to associate with phosphotyr-osine residues on the cytoplasmic domains of recept-ors such as the platelet-derived growth factor receptor(PDGFR) (Module 1: Figure PDGFR activation).
• The Src homology 3 (SH3) domain can bind to proline-rich regions of Shc, Cbl and dynamin.
• The proline-rich regions can bind to the SH3 domainsof Abl, Src, Lck, Lyn, Fyn and growth factor receptorbound protein 2 (Grb2).
The Class IB enzyme has a different activation mechan-ism in that it is stimulated by heterotrimeric G proteins(Module 2: Figure PtdIns 3-kinase signalling). It has ap110γ catalytic subunit and two regulatory subunits (p101and p84) (Module 2: Figure PI 3-K family). These regulat-ory subunits may mediate the translocation of the p110γ
catalytic subunit to the membrane by binding to the Gβγ
subunit.The Class I enzymes interact strongly with the onco-
gene Ras (Module 2: Figure PtdIns 3-kinase signalling) andthis can have important consequences for both upstreamand downstream events, depending on the cellular context.With regard to the former, the binding of Ras can enhanceenzyme activation and the generation of lipid messengers.On the other hand, the association with Ras might mediatesome of the downstream effects of the PtdIns 3-kinases.
Class I PtdIns 3-kinases are strongly inhibited by thefungal metabolite wortmannin (an irreversible inhibitor)and by LY294002 (a reversible inhibitor).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �37
Class II PtdIns 3-kinaseThe Class II PtdIns 3-kinase has a more restricted sub-strate range in that it phosphorylates just PtdIns andPtdIns4P. There are no regulatory subunits, and the threecatalytic subunits (PI3KC2α, PI3KC2β and PI3KC2γ)are characterized by having a C-terminal phox homology(PX) domain and a C2 domain (Module 2: Figure PI 3-Kfamily).
Class III PtdIns 3-kinaseThe Class III PtdIns 3-kinase has a restricted substraterange in that it phosphorylates just PtdIns. The prototypeof this class is vacuolar protein sorting 34 (Vps34) pro-tein found in Saccharomyces cerevisiae of which there isa human homologue (hVps34). The activity of hVps34 isregulated by Ca2+ and this contributes to the regulationof the target of rapamycin (TOR), which functions in cellgrowth control (see steps 4 and 5 in Module 9: Figure targetof rapamycin signalling). There is a Ca2+-dependent CaM-binding domain located between residues 318 and 334 inthe accessory domain (Module 2: Figure PI3-K family).
PtdIns 4-kinase (PtdIns 4-K)PtdIns 4-kinase catalyses phosphorylation of PtdIns onthe 4-position of the inositol ring to produce PtdIns4P(Step 2 in Module 2: Figure phosphoinositide metabol-ism). Enzyme activity is found in most cellular membranes(plasma membrane, endoplasmic reticulum, Golgi, nuclearmembrane and secretory vesicles). Cells have two classesof PtdIns 4-K: Type II and Type III. The latter has α-and β-isoforms and is sensitive to the drug wortmannin.Neither the PtdIns 4-KIIIα found in the endoplasmic re-ticulum nor the PtdIns 4-KIIIβ found in the cytosol andGolgi seem to be responsible for producing the PtdIns4Pin the plasma membrane. It is the Type III PtdIns 4-K iso-form that is activated by the neuronal Ca2+ sensor-1 pro-tein (NCS-1) to enhance exocytosis. The Type II PtdIns4-K also contains α- and β-isoforms that are insensitiveto wortmannin and may be responsible for generating thelipid substrates in the membrane that are used for sig-nalling. The enzymatic activity of PtdIns 4-kinase is greatlyenhanced in various cancer cells.
Originally, it was considered that PtdIns4P functionedsolely as a precursor for the formation of PtdIns4,5P2, butthere is increasing evidence for a role as part of a PtdIns4Psignalling cassette.
PtdIns phosphate kinasesThere is a family of PtdIns phosphate kinases that are di-vided into three subfamilies (Types I–III) that are localizedto different compartments and function to phosphorylatedifferent phosphoinositides.
PtdIns4P 5-kinase (PtdIns4P 5-K)This versatile family of Type I PtdIns phosphate kinases iscomposed of three isoforms (α, β and γ) that are locatedin different cellular regions such as the plasma membrane,focal adhesions, Golgi and nucleus. Their primary role is tophosphorylate PtdIns4P to PtdIns4,5P2 (Step 8 in Module2: Figure phosphoinositide metabolism).
PtdIns4P 5-kinase IαThis enzyme functions at multiple locations in the cell. It isthe major enzyme responsible for forming PtdIns4,5P2 atthe plasma membrane. It is also found within the nucleusat nuclear speckles that are sites of mRNA processing.
PtdIns4P 5-kinase IβThis isoform is found on membranes surrounding thenucleus.
Ins4P 5-kinase IγThe PtdIns4,5P2 function in focal adhesions depends upona spliced form of this enzyme, which is targeted to focaladhesions by interacting with talin (Module 6: Figure in-tegrin signalling). Talin contains a FERM domain, whichis used to bring PtdIns4P 5-kinase Iγ into the adhesioncomplex.
This Type I enzyme can be regulated by two separatemechanisms. Firstly, its activity is stimulated by phospha-tidic acid (PA) that is formed either by the phospholipaseD (PLD) signalling pathway where phosphatidylcholine(PC) is hydrolysed by phospholipase D (Module 2: Fig-ure PLD signalling) or by the phosphorylation of diacyl-glycerol (DAG) by diacylglycerol kinase (DAG) kinase(Module 2: Figure InsP3/DAG recycling). Secondly, en-zyme activity can be enhanced by small monomeric Gproteins such as Rho (Module 2: Figure Rho signalling).During its activation, PtdIns4P 5-kinase Iα translocates tothe plasma membrane through an activation mechanismthat is dependent upon Rac and Rho. This G protein reg-ulation results in the localized synthesis of PtdIns4,5P2,which then acts as a second messenger for the PtdIns4,5P2
signalling cassette. For example, it can influence a varietyof proteins to regulate processes such as actin remodelling,exocytosis, the PLD signalling pathway (Module 2: FigurePLD signalling) and the permeability of ion channels.
The PtdIns4P 5-Kα has an additional role in thatit can phosphorylate PtdIns3,4P2 to form the lipidsecond messenger PtdIns3,4,5P3 (Step 8 in Mod-ule 2: Figure phosphoinositide metabolism).
PtdInsP kinase IIThis enzyme, which is found in the cytosol, endoplas-mic reticulum and nucleus, but not in the plasma mem-brane, is a PtdInsP 4-kinase capable of phosphorylat-ing both PtdIns3P and PtdIns5P to produce PtdIns3,4P2
and PtdIns4,5P2 respectively (Steps 6 in Module 2: Figurephosphoinositide metabolism).
PtdInsP kinase IIIDespite being classified as a PtdInsP kinase, the main func-tion of PtdInsP kinase III is to phosphorylate PtdIns onthe 5-position to form PtdIns5P. One of the isoforms hasa FYVE domain to target it to endomembranes.
Phosphatase and tensin homologue deleted onchromosome 10 (PTEN)PTEN (phosphatase and tensin homologue deleted onchromosome 10) is a lipid phosphatase that is mutatedin many human cancers. Its gene is a tumour suppressor,
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �38
which functions in cell migration, proliferation and sur-vival. PTEN inactivates the 3-phosphorylated lipid secondmessengers that operate within the PtdIns 3-kinase sig-nalling cassette. It functions to remove the phosphate fromthe 3-position of PtdIns3,4P2 and PtdIns3,4,5P3 (Mod-ule 2: Figure phosphoinositide metabolism).
PTEN is a redox-sensitive enzyme, which may contrib-ute to a positive-feedback loop that enhances redox sig-nalling (Module 2: Figure plasma membrane ROS form-ation). It is inhibited by H2O2, which induces Cys-124in the active site to form a disulphide bond with Cys-71.This disulphide bond formation is specifically reversed bythioredoxin. A reversible inactivation of PTEN may thuscontribute to the accumulation of PtdIns3,4,5P3 (Mod-ule 2: Figure plasma membrane ROS formation), whichmay thus help to promote proliferation by switching offthe metabolism of the 3-phosphorylated lipid second mes-sengers. This reversible inactivation of PTEN contrib-utes to the accumulation of PtdIns3,4,5P3 by setting upa positive-feedback loop, since the formation of this lipidmessenger is responsible for stimulating the production ofH2O2 (Module 2: Figure plasma membrane ROS forma-tion).
There are additional enzymes that hydrolyse the 3-phosphorylated position. Transmembrane phosphatasewith tensin homology (TPTE) is localized to the plasmamembrane, but appears not to have phosphatase activity.TPTE and PTEN homologous inositol lipid phosphatase(TPIP) occurs as α and β isoforms. The TPIPα isoformhydrolyses PtdIns3,4,5P3, PtdIns3,5P2, PtdIns3,4P2 andPtdIns3P. It has N-terminal transmembrane domains thatappear to localize the protein to the ER.
PTEN is a potent tumour suppressor that is frequentlyinactivated in many different cancers, e.g. endometrial,prostate, mammary carcinomas, melanomas and thyroidtumours. When PTEN is inactivated in neurons, there is aprogressive increase in cell size and increased phosphoryla-tion of protein kinase B (PKB) that create cerebellar ab-normalities resembling those seen in human Lhermitte–Duclos disease (LDD).
Germline mutations in the PTEN gene have been im-plicated in the development of Cowden’s disease andBannayan–Zonana syndrome, where there is an increasedrisk of breast and thyroid cancers. An increase in the ex-pression of PTEN by the tumour suppressor p53 maycontribute to p53-induced apoptosis.
MyotubularinsThere is a large family of myotubularins that functionas lipid phosphatases that specifically dephosphorylatePtdIns3P. They may also act on PtdIns3,5P2. Mammaliancells express 14 myotubularins. The founding member isMTM1 and the remainder are MTMR2–MTMR13. Theseenzymes have a phosphatase domain, and a GRAM do-main, which associates with membranes and a C-terminalcoiled-coil domain that links these enzymes to other pro-teins. One of the isoforms, MTMR6, appears to have aspecific role in the PtdIns3P signalling cassette. Hydro-lysis of PtdIns3P may control the activity of the Ca2+-activated intermediate-conductance (IK) channel (KCa3.1),
and this may be of particular significance for the relation-ship between K+ channels and cell proliferation.
These enzymes are of interest because mutationsin MTM1 are responsible for X-linked recessivemyotubular myopathy and MTMR2 is mutated inCharcot-Marie-Tooth disease 4B.
Inositol phosphate metabolismThe Ca2+-mobilizing messenger function of inositol 1,4,5-trisphosphate (InsP3) is terminated through its metabol-ism by a complex inositol phosphate metabolic pathway(Module 2: Figure inositol phosphate metabolism) that hastwo main functions. Firstly, it produces free inositol thatis resynthesized to PtdIns, which can be reused for furthersignalling. Secondly, it generates an assortment of inositolphosphates, some of which contribute to the multipurposeinositol polyphosphate signalling pathway.
The Ins1,4,5P3 that enters the metabolic pathway ismetabolized via two pathways. It is dephosphorylated byType I inositol polyphosphate 5-phosphatase (step 1) toform Ins1,4P2 or it can be phosphorylated by InsP3 3-kinase to form Ins1,3,4,5P4 (step 4). Both Ins1,4P2 andIns1,3,4,5P4 are putative messengers in the multipurposeinositol polyphosphate signalling pathway. The Ins1,4P2
is sequentially dephosphorylated to free inositol. TheIns1,3,4,5P4 is dephosphorylated to Ins1,3,4P3, which oc-cupies an important position in the metabolic pathwayin that either it is dephosphorylated down to inositol,as part of an inositol recycling pathway, or it can bephosphorylated further to form additional inositol poly-phosphates. The inositol phosphates in the pink boxes inModule 2: Figure inositol phosphate metabolism have beenimplicated as intracellular messengers. InsP3 mobilizes in-ternal Ca2+ and the putative functions of the others aredescribed in the multipurpose inositol polyphosphate sig-nalling pathway.
Metabolism of the inositol phosphates is carriedout by a large number of inositol phosphate kinasesand phosphatases (Module 2: Figure inositol phosphatemetabolism).
Inositol polyphosphate 5-phosphataseStep 1 in Figure 2 inositol phosphate metabolism. Thislarge family of enzymes, which are products of multiplegenes and splice variants, removes the 5-phosphate groupfrom inositol lipids and, in some cases, can also act oninositol phosphates. All members of the family share acentral conserved catalytic domain:
• 5-Phosphatase I. Functions to hydrolyses Ins1,4,5P3
and Ins1,3,4,5P4. If this enzyme is deleted, thereis an increase in the level of InsP3, and thiswas associated with a transformed phenotype.This enzyme is inhibited following phosphoryla-tion by Ca2+/calmodulin-dependent protein kinase II(CaMKII). Type I, which hydrolyses the inositol phos-phates, is directed to the membrane by isoprenylationof the C-terminal region. If these membrane attachmentsites are removed, allowing the enzyme to disperse intothe cytosol, InsP3 metabolism appears to be severely
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �39
curtailed. It seems that much of the InsP3 is metabol-ized close to its site of action at the plasma membrane.
• 5-Phosphatase II. A 75 kDa protein that hydrolysesinositol phospholipids.
• OCRL. A gap-domain-containing enzyme that canhydrolyses both inositol phosphates (Ins1,4,5P3 andIns1,3,4,5P4) and the phosphoinositides PtdIns4,5P2
and PtdIns3,4,5P3. A deficiency of this enzyme is re-sponsible for Lowe’s oculocerebrorenal (OCRL) syn-drome.
• SHIP. The Src homology 2 (SH2) domain-containinginositol phosphatases (SHIP1 and SHIP2) hydrolyseIns1,3,4,5P4 and PtdIns3,4,5P3. The SHIPs, which comein different forms, are found in haematopoietic cells,where they play a role in inhibiting signalling. In Bcells, for example, the FcγRIIB receptor has an im-munoreceptor tyrosine-based inhibitory motif (ITIM)to which SHIPs attach causing a reduction in Ca2+ sig-nalling by hydrolysing PtdIns3,4,5P3 and thereby re-ducing the stimulatory effect of the Tec tyrosine kinasefamily on phospholipase Cγ (Module 2: Figure ROS ef-fects on Ca2+ signalling). SHIPs perform a similar func-tion in mast cells (Module 11: Figure mast cell inhibit-ory signalling). SHIP1 functions to localize PIP3 at thefront of the cell during neutrophil chemotaxis (Mod-ule 11: Figure neutrophil chemotaxis).
• Synaptojanin-1.• Synaptojanin-2.• Proline-rich inositol polyphosphate 5-phosphatase
(PIPP).• Skeletal muscle and kidney enriched inositol phos-
phatase (SKIP).
Inositol polyphosphate 1-phosphataseSteps 2 in Module 2: Figure inositol phosphate meta-bolism. This enzyme dephosphorylates both Ins1,3,4P3
and Ins1,4P2. This enzyme has been implicated in thesignalling pathways of compensatory hypertrophy and isup-regulated in human colorectal cancer.
Inositol monophosphataseSteps 3 in Module 2: Figure inositol phosphate metabol-ism. The free inositol that is formed by this enzyme isthen used to resynthesize PtdIns, which is returned tothe plasma membrane, where it can be reused to functionas the precursor for the phosphoinositide signalling lip-ids (Module 2: Figure phosphoinositide metabolism). Thisinositol monophosphatase is inhibited by lithium (Li+),which thus acts to reduce the supply of inositol (Module2: Figure InsP3/DAG recycling). This ability of Li+ tolower the level of inositol is the basis of an inositol de-pletion hypothesis to account for its action in controllingmanic-depressive illness.
Ins1,4,5P3 3-kinaseStep 4 in Module 2: Figure inositol phosphate metabolism.This kinase is one of the major pathways for metabolizingthe second messenger Ins1,4,5P3. There are three isoformsof this enzyme (InsP3 3-kinase A, B and C). Both type A
and type B are activated by Ca2+/calmodulin, with thetype B being the most sensitive. This enzyme is also sens-itive to other signalling pathways in that both forms areinhibited by PKC. On the other hand, protein kinase A(PKA) activates type A but inhibits type B.
Inositol polyphosphate 4-phosphataseSteps 5 in Module 2: Figure inositol phosphate metabol-ism. A multifunctional phosphatase capable of dephos-phorylating both Ins3,4P2 and Ins1,3,4P3, as well as theinositol lipid PtdIns3,4P2. This enzyme might be a regu-lator of cell proliferation, since it is absent in hyperprolif-erative megakaryocytes that lack the transcription factorGATA-1.
Inositol polyphosphate 3-phosphataseSteps 6 in Module 2: Figure inositol phosphate metabol-ism. A multifunctional phosphatase capable of dephos-phorylating both Ins1,3P2 and PtdIns3P.
Ins1,3,4P3 6-kinaseStep 7 in Module 2: Figure inositol phosphate metabolism.
Ins1,3,4,6P4 5-kinaseStep 8 in Module 2: Figure inositol phosphate metabolism.
Ins3,4,5,6P4 1-kinaseStep 9 in Module 2: Figure inositol phosphate metabolism.This enzyme is responsible for inactivating the Ins3,4,5,6P4
that regulates chloride channels. It is an unusual enzymein that it has alternative positional specificity (5/6-kinaseactivity against Ins1,3,4P3). The enzyme is the same as theIns1,3,4,5,6P5 1-phosphatase.
Ins1,3,4,5,6P5 1-phosphataseStep 10 in Module 2: Figure inositol phosphate metabol-ism. The enzyme is equivalent to the Ins 3,4,5,6P4 1-kinase.
Ins1,4,5,6P4 3-kinaseStep 11 in Module 2: Figure inositol phosphate meta-bolism.
Ins1,3,4,5,6P5/InsP6 3-phosphataseSteps 12 in Module 2: Figure inositol phosphate metabol-ism. This enzyme is also known as the multiple inositolphosphate phosphatase (MIPP). Its function is somewhatunclear because it is located within the lumen of the ER,apparently insulated from its two substrates.
Ins1,3,4,5,6P5 2-kinaseStep 13 in Module 2: Figure inositol phosphate metabol-ism. The production of InsP6 occurs primarily through thephosphorylation of Ins1,3,4,5,6P5 by a 2-kinase.
InsP6 kinaseSteps 14 in Module 2: Figure inositol phosphate meta-bolism. This kinase adds an additional phosphate to thatalready present on the 5-position to produce PP-InsP5
(InsP7). This kinase can run in reverse and is thus poten-tially capable of reforming ATP. The same enzyme may act
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �40
on Ins1,3,4,5,6P5 to produce PPInsP4. There are three iso-forms (InsP6K1–InsP6K3). InsP6K2 has been implicatedin the action of interferons on cell growth and apoptosis.
Diphosphoinositol phosphate phosphohydrolase(DIPP)Steps 15 in Module 2: Figure inositol phosphate metabol-ism. This enzyme dephosphorylates both PP-InsP5 (InsP7)and [PP]2-InsP4 (InsP8), during which it removes the β-phosphate from the diphosphate groups.
PP-InsP5 kinaseStep 16 in Module 2: Figure inositol phosphate metabol-ism. This kinase adds a further phosphate to PP-InsP5 toform [PP]2-InsP4, but the positional specificity of the lat-ter remains to be established. Like the InsP6 kinase, thisenzyme is capable of forming ATP by running in a reversemode.
Mammalian inositol phosphate multikinase (mIPMK)Steps 17 in Module 2: Figure inositol phosphate metabol-ism. This mammalian inositol phosphate multikinase (mI-PMK) is a multifunctional kinase capable of the followingphosphorylations:
• Ins4,5P2 → Ins1,4,5P3
• Ins1,4,5P3 → Ins1,3,4,5P4
• Ins1,3,4,5P4 → InsP5
• InsP5 → PP-InsP4
Inositol 1,4,5-trisphosphate (InsP3)/Ca2+signalling cassetteThis signalling cassette uses the second messenger inos-itol 1,4,5-trisphosphate (InsP3) to mobilize Ca2+ from in-ternal stores. When external stimuli engage receptors onthe cell surface, they activate the enzyme phospholipase C(PLC), which hydrolyses PtdIns4,5P2 to yield InsP3 anddiacylglycerol (DAG) (Module 2: Figure InsP3 and DAGformation). This is a bifurcating signalling pathway in thatInsP3 generates a Ca2+ signal, whereas DAG functions tostimulate protein kinase C (PKC). The InsP3 released fromthe membrane diffuses into the cytosol where it engagesthe InsP3 receptors (InsP3Rs) to release Ca2+ from theendoplasmic reticulum.
The operation of this InsP3/Ca2+ signalling cassette canbe divided into separate components:
• PtdIns conversion into the precursor lipid PtdIns4,5P2
• Hydrolysis of PtdIns4,5P2 to generate InsP3 and DAG• Metabolism of InsP3 and DAG and the resynthesis of
PtdIns• Inositol 1,4,5-trisphosphate (InsP3) and Ca2+ release
PtdIns conversion into the precursor lipid PtdIns4,5P2
The precursor PtdIns4,5P2, which is hydrolysed to giveInsP3, is part of the complex inositol lipid metabolic path-way (Module 2: Figure phosphoinositide metabolism). Thesynthesis of the lipid precursor PtdIns4,5P2 from PtdIns iscontrolled by two substrate cycles involving lipid kinasesand phosphatases (Module 2: Figure InsP3 and DAG form-ation). In the first step, the hydroxy group at the 4-position
of the inositol ring is phosphorylated to form PtdIns4P.In the next step, the PtdIns4P is phosphorylated furtherto yield the precursor PtdIns4,5P2. The next step is theagonist-dependent hydrolysis of PtdIns4,5P2 to generateInsP3 and diacylglycerol (DAG).
Hydrolysis of PtdIns4,5P2 to generate InsP3 anddiacylglycerol (DAG)External stimuli (e.g. hormones, neurotransmitters andgrowth factors) gain access to this signalling pathway byactivating cell-surface receptors. The latter fall into twomain classes, the G protein-coupled receptors (GPCRs)and the protein tyrosine kinase-linked receptors (PTKRs).During the transduction process, the precursor lipidPtdIns4,5P2 is hydrolysed by phospholipase C (PLC) toproduce both InsP3 and DAG. The family of PLCs can bedistinguished by the way they are coupled to cell-surfacereceptors. In general, the GPCRs use the PLCβ isoforms,whereas the receptor tyrosine kinases (RTKs) are coupledto the PLCγ isoforms (Module 2: Figure PLC structureand function).
DAG functions to activate the diacylglycerol(DAG)/protein kinase C (PKC) signalling cassette,whereas the InsP3 diffuses into the cytosol to activate theInsP3 receptors to release Ca2+ stored in the endoplasmicreticulum. The signalling function of this bifurcating sig-nalling pathway is curtailed by the metabolism of InsP3
and the resynthesis of PtdIns.
Metabolism of InsP3 and DAG and the resynthesis ofPtdInsThe metabolism of InsP3 and DAG and the resynthesisof PtdIns are the OFF reactions that terminate the ac-tions of these two messengers. There are two pathwaysfor diacylglycerol (DAG) metabolism. Similarly, there aretwo separate mechanisms of inositol 1,4,5-trisphosphate(InsP3) metabolism.
Diacylglycerol (DAG) metabolismThe second messenger DAG is metabolized via two separ-ate pathways. It can be phosphorylated by diacylglycerol(DAG) kinase to form phosphatidic acid (PA) or it is hy-drolysed by diacylglycerol (DAG) lipase. The PA is trans-ferred to the endoplasmic reticulum, where it interactswith CTP to form the CDP/DAG complex, which is aprecursor in the resynthesis of PtdIns (Module 2: FigureInsP3/DAG recycling).
Diacylglycerol (DAG) kinaseDiacylglycerol (DAG) kinase α (DAGKα), which is oneof a family of nine mammalian isotypes, has a numberof domains, including Ca2+-binding EF-hand motifs andan N-terminal recoverin homology domain that is relatedto the recoverin family of neuronal Ca2+ sensors. Thesetwo domains appear to function as a unit during Ca2+-induced activation of DAGKα. In response to an increasein Ca2+, DAGKα translocates to the membrane, whereit phosphorylates DAG to phosphatidic acid (PA) (Mod-ule 2: Figure InsP3/DAG recycling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �41
Module 2: Figure inositol phosphate metabolism
Ins1,4,5P3
Ins4P Ins1P Ins3P
Inositol
Inos
itol l
ipid
sig
nalli
ng
Inositol lipid synthesis
Ins1,3,4,5P4Ins1,3,4,6P4
Ins1,3,4P3
Ins3,4P2Ins4,5P2 Ins1,3P2Ins1,4P2
Ins3,4,5,6P 4Ins1,4,5,6P
Ins1,3,4,5,6P5
InsP6
ADP + Pi
ATP
ATP
5PPInsP (InsP )7
42[PP] InsP (InsP ) 8
4
44
PP-InsP
52
2
1
1
5
3 33
4
17
17
17
7
6
8 910 12
121314
11
14 15
1516
Cl-
+-
Inositol phosphate metabolism.The major input into this metabolic system is the Ins1,4,5P3 released from the membrane following receptor-activated inositol lipid signalling. One ofthe functions of this pathway is to form free inositol that is used for inositol lipid synthesis. The pathway also generates additional inositol phosphates(highlighted in pink) that have putative signalling functions. See the text for further details of the enzymes responsible for Steps 1–17.
Diacylglycerol (DAG) lipaseA diacylglycerol (DAG) lipase is responsible for removingone of the fatty acid tails from the sn-position of DAGto form monoacylglycerol. This hydrolysis of DAG mayrepresent the primary mechanism for removing the DAGthat is produced following the hydrolysis of inositol lipids.
Inositol 1,4,5-trisphosphate (InsP3) and Ca2+ signallingThe primary function of inositol 1,4,5-trisphosphate(InsP3) is to function as a second messenger to releaseCa2+ from the internal stores. The 1,4,5-trisphosphate re-ceptors (InsP3Rs) located on the endoplasmic reticulumrespond to InsP3 by releasing puffs of Ca2+ (Module 3:Figure InsP3R activation). These InsP3Rs are sensitive toboth InsP3 and Ca2+ and can thus function as co-incidentdetectors (Module 2: Figure Ca2+-induced Ca2+ release).
This InsP3/Ca2+ signalling system controls many differentcellular processes in a large number of different cell types(Module 2: Figure InsP3/Ca2+ signalling functions).
Inositol 1,4,5-trisphosphate (InsP3) metabolismThe inositol 1,4,5-trisphosphate (InsP3) that is formedduring signalling enters the pathways of inositol phos-phate metabolism from which it emerges as free inositol.It is dephosphorylated by Type I inositol polyphos-phate 5-phosphatase (Steps 1 in Module 2: Figure inositolphosphate metabolism) to form Ins1,4P2 or it can be phos-phorylated by InsP3 3-kinase to form Ins1,3,4,5P4 (Step4). These two products then enter a complex metabolicpathway that plays an important role in recycling the inos-itol headgroup. Cells have access to three separate sourcesof inositol: recycling the second messenger InsP3, de novo
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �42
Module 2: Figure InsP3 and DAG formation
PtdIns PtdIns4P PtdIns4,5P 2
InsP
OH
3
Diacylglycerol
Calcium signalling
Protein phosphorylation
P
P P
P P
P
P
P PPKC
PtdIns 4-kinase
PtdIns4P 5-kinase
PtdIns4,5P Pase
PtdIns4P Pase
2
PLCGq/11
Agonist
Plasma membrane
R
ecep
tor
Agonist-dependent formation of the second messengers inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG).The inositol lipids that function in signalling are embedded in the inner leaflet of the plasma membrane. The precursor lipid is phosphatidylinositol(PtdIns), which is successively phosphorylated, first on the 4-position to form PtdIns4P and then on the 5-position to form PtdIns4,5P2. Activatedcell-surface receptors are coupled through the G protein Gq/11 to phospholipase C (PLC) that hydrolyses PtdIns4,5P2 to generate inositol 1,4,5-trisphosphate (InsP3). InsP3 activates Ca2+ signalling, and diacylglycerol (DAG) stimulates protein kinase C (PKC) to initiate protein phosphorylation(Module 2: Figure PKC structure and activation). An animated version of this figure is available.
Module 2: Figure InsP3/Ca2+ signalling functions
Proliferation T cell Mesangial cellSmooth muscleBrown fat cell
Fluid secretion Intestinal cell Parietal cell Pancreas Salivary gland Sweat gland
ContractionSmooth muscleMesangial cellModulation ofventricular andatrial cell contraction
Aldosterone secretion Glomerulosa cell
Exocytosis b-CellL CellMast cell,MacrophageParathyroid glandPiuitary cellsAstrocytesRenin-producing granular cells
Differentiation Osteoclasts Brown fat cells
ChemotaxisNeutrophilsOsteoclast precursors (?)
Ion channel opening AstrocytesT Cells
FertilizationMetabolism Liver cells
Neuronal synapticplasticityPurkinje neuronsHippocampal neurons
Aggregation Blood platelets
InsP /Ca32+
InsP3/Ca2+ signalling functions.The mobilization of Ca2+ by inositol 1,4,5-trisphosphate (InsP3) functions in the control of many different cellular processes in a wide range of celltypes.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �43
Module 2: Figure InsP3/DAG recycling
PtdIns
PtdIns
PtdIns4P PtdIns4,5P2
PLC DAG DAGkinase
DAGlipase
PA
MAG
Na+
2
3
4 PA
CTP
InsPkinase
InsP
InsPPase
3
3
3
InsP InsP
InsP2 4
1
CDP.DAG
PtdInsPITP
PITP
PtdInssynthetase
CDP.DAGsynthetase
Lithium
Valproate
-
-
-
Endoplasmic reticulum
Inositol
Plasma inositolStimulus
G-6-PO4
de novo synthesis
Recycling
Dietary inositol
Inositol synthase
Inositolmonophosphatase
1
6
5
SMIT1
Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG) recycling.The cell has access to three sources of inositol. It can be synthesized de novo from glucose 6-phosphate (G-6-PO4) by an inositol synthase, itcan come into the cell from the plasma using a sodium-dependent myo-inositol cotransporter-1 (SMIT1) or it can be obtained by recycling InsP3.Agonist-dependent inositol lipid metabolism occurs through a series of steps as outlined in the text.
synthesis from glucose 6-phosphate or uptake of di-etary inositol circulating in the plasma (Module 2:Figure InsP3/DAG recycling). Drugs such as lithiumand valproate may control manic-depressive illness byreducing the supply of inositol by inhibiting the inositolmonophosphatase and inositol synthase respectively. Thesupply of free inositol is one of the essential precursorsfor the synthesis of PtdIns.
Synthesis of PtdInsThe two second messengers that are formed during phos-phoinositide signalling can be recycled back to the pre-cursor lipid PtdIns4,5P2 through a series of steps (Mod-ule 2: Figure InsP3/DAG recycling):
1. Conversion of PtdIns into the precursor lipidPtdIns4,5P2.
2. The agonist-dependent activation of phospholipase C(PLC), which hydrolyses PtdIns4,5P2 to generate thesecond messengers inositol 1,4,5-trisphosphate (InsP3)and diacylglycerol (DAG).
3. The InsP3 is recycled back to free inositol by a complexmetabolic pathway (Module 2: Figure inositol phos-phate metabolism). DAG is phosphorylated by DAGkinase to form phosphatidic acid (PA) or it is hy-drolysed by DAG lipase to form monoacylglycerol(MAG).
4. The PA is transferred from the plasma membrane to theER where it interacts with CTP to form CDP/DAG
by a CDP/DAG synthetase. The inositol, which canbe produced through three mechanisms (de novo syn-thesis, recycling or uptake of dietary inositol), is at-tached to CDP/DAG by the PtdIns synthetase [cytid-ine diphosphate (CDP)/diacylglycerol (DAG):myo-inositol 3-phosphatidyltransferase] located on the en-doplasmic reticulum (ER) to form PtdIns.
5. The PtdIns is then transported from the ER back to theplasma membrane by a PtdIns transfer protein (PITP).Cells express two PITPs, an α and a β isoform, pro-duced by separate genes. The latter appears to be thehousekeeping isoform that is essential for cell survival,whereas the α isoform has a more specialized function.When PtdIns is added back to the plasma membrane, itcan once again be converted through the two phos-phorylation reactions to maintain the supply of thePtdIns4,5P2 that is the precursor for the phosphoin-ositide signalling pathway.
6. Cells can also use plasma inositol, which is taken upby a sodium-dependent myo-inositol cotransporter-1(SMIT1).
Phospholipase C (PLC)Phospholipase C (PLC) hydrolyses the lipid precursorPtdIns4,5P2 to produce both InsP3 and DAG. It is madeup of five subclasses, which have variable isoforms PLCβ
(β1–β4), PLCδ (δ1–δ4), PLCγ (γ1 and γ2), PLCε andPLCζ (Module 2: Figure PLC structure and function).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �44
Module 2: Figure PLC structure and function
PLC 1 - 4)β(
PLCβGPCR PTKR
PHEF hands
X C2
C2
C2 RA
RARA
RA
Y
Y
Y
Y
X
X
EF hands
C2YX
XRasGEF
RasGEF
PH
PH
PH SH3SH2 SH2PLC (1, 2)γ
PLCγ
Translocation
PLC (1 - 4)δ
PLCδ
PLCε
PLCε
PLCζ
IP IPIP IP3 33 3
BL
NK
Ca
CaCa Ca Ca
2+
2+2+ 2+ 2+
PP P
P
PPH
PH
PH PHPHEF
EF
EF EFEFC2Gα
Gα
Gβγ
Gβγ
GqC2
XY
XY
XY XYXYSH2
SH2
SH2
SH2
Ras
+
+
+C2
The domain structure and activation mechanisms of PLC isoforms.The pleckstrin homology (PH) domain, four EF-hand domains, the catalytic X and Y domains, and the C2 domain are a common feature of most ofthe PLC isoforms. The two PLCγ isoforms differ from the others by lacking a C2 domain and by having additional domains such as the Src homology2 (SH2) and Src homology 3 (SH3) domains located between a split PH domain. The black triangles indicate the sites of tyrosine phosphorylationthat are important for the activation of PLCγ. PLCε also has additional domains related to its activation by Ras, such as the Ras association motifs(RA) and the Ras guanine nucleotide exchange factor (RasGEF). The bottom panel illustrates how the various PLC isoforms are activated by differentmechanisms. Not included in this figure is the action of PLCζ, which is unusual in that it is activated at fertilization following its injection into the oocyte(Module 8: Figure mammalian fertilization).
All forms of the enzyme have an absolute requirementfor Ca2+ that plays a critical role in the catalytic site. ThePtdIns4,5P2 is cleaved through two sequential reactions:first, the phosphodiester bond is cleaved to DAG and in-ositol 1,2-cyclic phosphate, the latter is then hydrolysed togive the acyclic InsP3 that is released into the cytoplasm.The domain structure of the different PLC isoforms re-veals a number of common structural features related tothe way the enzyme associates with the membrane andfunctions to hydrolyse PtdIns4,5P2. The catalytic domainis made up from the X and Y regions. They have at leasttwo potential lipid-binding domains: the pleckstrin homo-logy (PH) domain and the C2 domain (Module 6: Figuremodular lipid-binding domains). In the case of PLCδ1, theenzyme may first associate with the membrane through itsPH domain, which has a high affinity for PtdIns4,5P2. Fur-ther interactions may then occur through the C2 domain,which has an extensive interface with the catalytic domainand may thus enable the catalytic site to integrate itself intothe membrane to hydrolyse the lipid. The main differencebetween these PLC isoforms concerns the way in whichthey are activated.
Phospholipase Cβ (PLCβ)The PLCβ isoforms are mainly activated by G protein-coupled receptors (GPCRs). PLCβ1 and PLCβ3 are fairlyubiquitous, whereas PLCβ2 and PLCβ4 have more lim-ited tissue distributions.
PLCβ isoform distribution and function
PLCβ1 is highly concentrated in brain (pyramidal cells ofhippocampus, Purkinje cells and granule cells). The knock-out phenotype is characterized by seizures leading to sud-den death. These seizures are similar to those seen dur-ing epilepsy. Many of the defects are found in the centralnervous system (CNS). Activation of PLC by muscarinicreceptors is suppressed in the temporal lobe, cerebellumand hippocampus, and this could decrease the inhibitorytone, leading to the seizures.
PLCβ2 is mainly expressed in cells of the immune sys-tem. Knockout mice show some disruption of chemokinesignalling. For example, neutrophils fail to respond tothe chemoattractant fMet-Leu-Phe with the usual changesin PLC activation, Ca2+ release and superoxide radical(O2
−•) production. However, the response to lipopolysac-charide (LPS) is normal. Despite this absence of PLC activ-ation in leucocytes, chemotactic responses are enhanced,suggesting that this signalling pathway may normally ant-agonize the signalling pathways normally used to controlchemotaxis.
PLCβ3 is found mainly in brain, parotid, smoothmuscle and liver. Disruption of this gene is lethal, withthe mice dying by day 2.5. The embryos are highly dis-organized and have low cell numbers, suggesting a rolefor this isoenzyme in cell division. This isoform has twosites (Ser-26 and Ser-105) that are phosphorylated by cyclic
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �45
GMP-dependent protein kinase (PKG) resulting in a de-crease in enzyme activity (Module 7: Figure smooth musclecell cyclic GMP signalling).
PLCβ4 is found in cerebellum and granule cells. Knock-out mice appear to have defects in the cerebellum, result-ing in poor motor co-ordination due to a decrease in PLCstimulation through metabotropic glutamatergic and mus-carinic receptors. There also are defects in the processingof visual information.
The primary activation mechanism of the PLCβ iso-forms is through the Gq family of heterotrimeric G pro-teins (Gq, G11, G14, G15 and G16) (Module 2: Figure PLCstructure and function). While Gq and G11 are foundin most tissues, the other three are restricted to cells ofhaematopoetic origin. The function of these G proteinsis complicated because the PLCβ isoforms are sensitiveto both the α and βγ components of the heterotrimericcomplex. PLCβ1 and PLCβ4 are most sensitive to stim-ulation through α subunits, whereas βγ is more effectiveat activating PLCβ2 and PLCβ3. This sensitivity to βγ
subunits may explain the pertussis-toxin-sensitive stimu-lation of PLC by the Gi family of G proteins. For example,the adenosine A1, muscarinic M2, somatostatin and µ-, δ-and κ-opioid receptors can couple to PLC through Gi andGo. Of the five Gβ and 11 Gγ subunits, there appears tobe little specificity with regard to the activation of PLC.In general, Gβγ is less efficacious than Gα in stimulatingPLC.
The two G protein subunits interact with PLCβ at dif-ferent sites. The α subunit interacts with a region withinthe long C-terminal tail, whereas the βγ subunit interactswith a site within the Y region of the catalytic site (Module2: Figure PLC structure and function).
PLCβ also contains GAP (GTPase-activating protein)activity that stimulates the intrinsic GTPase activity of theα subunit. When an agonist binds to the GPCRs, it inducesa conformational change that is transmitted to the under-lying G protein, causing it to dissociate. At this time theGDP bound to the α subunit is exchanged for GTP. Theactive GTP/α subunit complex can then activate PLCβ
and this activation process is terminated when the intrinsicGTPase activity of the α subunit hydrolyses GTP backto GDP allowing the GDP/α subunit to once again bindto the βγ subunit to form the inactive complex. The in-trinsic GTPase activity of the α subunit is rather low, butis greatly enhanced by two mechanisms. There is a GAPactivity associated with the long C-terminal tail of PLCβ.Therefore PLCβ plays a direct role in terminating its ownactivity. In addition, there are regulators of G protein sig-nalling (RGS) proteins. There are about 20 of these RGSproteins, of which RGS2–RGS4 seem particularly effect-ive in interacting with Gαq. The GTPase activity of Gαq
is enhanced 25-fold by RGS4. There is considerable dis-crimination with regard to how individual RGS proteinsbind to specific receptor/Gαq complexes enabling themto exert an agonist-specific inhibitory mechanism. Thismechanism could explain the different types of Ca2+ sig-nals observed in hepatocytes following stimulation withdifferent agonists.
Modulation of PLCβ by other signalling pathwaysIn general, the phosphorylation of PLCβ, particularly iso-forms 2 and 3, by protein kinase A (PKA) results in inhib-ition of the enzyme.
Phospholipase Cγ (PLCγ)PLCγ functions predominantly in early development andin the signalling pathways that control cell proliferation.PLCγ1 knockout mice die at embryonic day 9, by whichtime the embryos appear normal, albeit somewhat smaller.
PLCγ is characterized by having a large insert betweenthe X and Y domains (Module 2: Figure PLC structure andfunction). This insert contains an additional pleckstrin ho-mology (PH) domain, which is itself split by another insertcontaining two Src homology 2 (SH2) domains and a singleSrc homology 3 (SH3) domain. The SH2 domains play acritical role in the activation of PLCγ because they providea docking module that enables the enzyme to translocate tothe membrane to dock to phosphorylated tyrosine residueson activated receptors or associated scaffolding proteins.This tyrosine phosphorylation occurs during activation ofprotein tyrosine kinase-linked receptors (PTKRs) or thenon-receptor protein tyrosine kinases (e.g. Src, Syk, Btk,Lck, Fyn). This sequence of events is well illustrated bythe activation of PLCγ1 during activation of the T cell re-ceptor (Module 9: Figure TCR signalling) or PLCγ2 fol-lowing stimulation of the B cell receptor (BCR) (Module 2:Figure ROS effects on Ca2+ signalling). This translocationto the cell surface has two important consequences for theactivation of the enzyme. Firstly, it brings the enzyme closeto its substrate in the membrane. Secondly, by interactingwith tyrosine kinases, PLCγ1 is itself phosphorylated ontyrosine residues located at positions 771, 783 and 1254(black triangles in Module 2: Figure PLC structure andfunction). Phosphorylation at Tyr-783 is particularly im-portant for switching on enzyme activity. The PLCγ2 iso-form is activated by phosphorylation of tyrosine residues753 and 759, mainly by the tyrosine kinase Btk (Module2: Figure ROS effects on Ca2+ signalling).
The enzymatic activity of PLCγ is also stimulated bythe PtdIns3,4,5P3 produced by the PtdIns 3-kinase sig-nalling pathway and thus represents a major point of in-teraction between these two signalling pathways. This in-teraction is particularly important during lymphocyte ac-tivation, where the Tec tyrosine kinase family are activ-ated by PtdIns3,4,5P3 to phosphorylate and activate bothPLCγ1 and PLCγ2. Both isoforms bind strongly to thishighly charged lipid through the N-terminal PH domain,and this interaction enhances enzymatic activity.
The Src homology 3 (SH3) domain, which binds toproline-rich sequences, may enable PLCγ to bind to othercomponents, such as the cytoskeleton and the proteindynamin.
Phospholipase Cδ (PLCδ)Less is known about the activation of PLCδ. Unlike PLCβ
and PLCγ, PLCδ appears not to be regulated by receptorsdirectly. Once PLCδ1 has associated with the membrane,it appears to be activated by an elevation in cytosolic Ca2+.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �46
This sensitivity to changes in intracellular Ca2+ is thoughtto depend on the C2 domain (Module 2: Figure PLC struc-ture and function).
Crystallographic studies of PLCδ1 have begun to revealhow PLC enzymes function to cleave their lipid substratesthat are embedded in the plasma membrane.
Phospholipase Cε (PLCε)Phospholipase Cε (PLCε) differs from the other isoformsby having two Ras-association domains (RA) at its C-ter-minus (Module 2: Figure PLC structure and function).It binds specifically to the GTP-bound forms of Ha-Rasand Rap1A. The enzyme is targeted to the membranethrough its ability to bind to these G proteins. It also hasa Cdc25 homology domain that is a guanine nucleotideexchange factor (GEF) motif for Rap1, which enables it totranslocate to the perinuclear region following epidermalgrowth factor (EGF) stimulation, where it can activate themitogen-activated protein kinase (MAPK) signalling path-way. There is evidence that cyclic AMP acting through theexchange protein activated by cAMP (EPAC) can stimu-late Rap1 to activate PLCε (Module 2: Figure cyclic AMPsignalling).
Phospholipase Cζ (PLCζ)Phospholipase Cζ (PLCζ) has a restricted expression inthat it is only found in mammalian sperm. PLCζ lacksthe N-terminal PH domain (Module 2: Figure PLC struc-ture and function), which means that there is no obviousmechanism for it to bind to phospholipids. PLCζ has asensitivity to Ca2+ that is 100-fold greater than that ofPLCδ1. Both the C2 and the tandem EF-hands are neces-sary for PLCζ to hydrolyse PtdIns4,5P2 to generate theInsP3 that plays a central role in sperm-induced oocyte ac-tivation during fertilization (Module 8: Figure mammalianfertilization).
Diacylglycerol (DAG)/protein kinase C (PKC)signalling cassetteThe diacylglycerol (DAG) that is formed whenPtdIns4,5P2 is hydrolysed by phospholipase C (PLC) re-mains within the plane of the membrane where it acts as alipid messenger to stimulate some members of the proteinkinase C (PKC) family (Module 2: Figure InsP3 and DAGformation). This PKC family contains a number of iso-forms with diverse structures and activation mechanisms.The PKC signalling function is equally diverse, and it hasproved difficult to pin down its precise function in theregulation of specific cellular processes.
Protein kinase C (PKC)The protein kinase C family has been divided into threesubgroups (Module 2: Figure PKC structure and activa-tion):
• Conventional PKCs (cPKCs: α, β1, β2 and γ) containC1 and C2 domains
• Novel PKCs (nPKCs: δ, ε, η and θ) contain a C1 and aC2-like domain
• Atypical PKCs (aPKCs: ζ, ι and λ) contain a truncatedC1 domain
The activation of these different PKC families is com-plex in that it depends on a number of processes suchas priming, translocation and association with scaffoldingproteins. The priming process is driven by a sequence ofmultisite phosphorylation events that have to occur beforethe enzyme can perform its signalling function. The newlysynthesized proteins are inactive and undergo a prim-ing process that begins when phosphoinositide-dependentprotein kinase 1 (PDK1), which is part of the PtdIns 3-k-inase signalling pathway, phosphorylates a site in the C4domain (Module 2: Figure PKC structure and activation).This is then followed by intramolecular autophosphoryla-tion reactions of two further sites in the C-terminal tail.The primed enzyme is now ready to perform its signallingfunction by responding to both diacylglycerol (DAG)and Ca2+. The cPKCs and most of the nPKCs are lipid-sensitive enzymes in that they are activated by DAG thatbinds to the C1 domain. This C1 domain is also respons-ible for binding the tumour-promoting phorbol esters. Inthe case of the cPKCs, Ca2+ also plays an important roleas a cofactor that binds to the C2 domain to increase itsaffinity for acidic phospholipids and is thus responsible forinducing the translocation of cPKCs to the plasma mem-brane.
The activation of the cPKC isoforms follows a setsequence of events that begins with the hydrolysis ofPtdIns4,5P2 to form DAG and inositol 1,4,5-trisphosphate(InsP3). The InsP3 releases Ca2+, which initiates the activa-tion process by binding to the C2 domain to induce a con-formational change that greatly enhances the membraneaffinity of cPKC and thus promotes its translocation tothe membrane where it is activated when it makes contactwith DAG. The pseudosubstrate (PS) domain swings awayfrom the active site, which is now free to phosphorylate itssubstrates.
The nPKCs, which are also activated by DAG, havea much slower activation process because the Ca2+-dependent facilitation of membrane translocation isabsent.
Protein kinase Cζ (PKCζ)This is one of the atypical PKC isoforms (aPKC),which fails to respond to diacylglycerol (DAG) or Ca2+,but is sensitive to low levels of ceramide (Module 2:Figure sphingomyelin signalling). PKCζ is also activ-ated following phosphorylation by kinases from othersignalling pathways. For example, PKCζ is activatedby phosphoinositide-dependent kinase 1/2 (PDK1/2)(Module 2: Figure PtdIns 3-kinase signalling). In somecells, it is responsible for activating nuclear factor κB (NF-κB). PKCζ can also phosphorylate the glucose transporterin skeletal muscle (Module 7: Figure skeletal muscle E-Ccoupling).
Protein kinase Cθ (PKCθ)Protein kinase Cθ (PKCθ) is one of the novel PKCs(nPKCs) that plays an important role in lymphocyte ac-tivation where it is concentrated in the central region oftheimmunological synapse (Module 9: Figure immunolo-gical synapse structure).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �47
Module 2: Figure PKC structure and activation
C1A
C1A
C1
C1
C1
C1
C1B
C1B
C2
C2
C2
C2
C2-like
C3 C4
C4
C4C3
C3
PS
PS
PS
PS
P
P
P
P
P
P
P
P
Conventional (cPKC)
(α, βΙ, , γ)βΙΙ
Novel (nPKC)
(δ, ε, η)θ,
Atypical (aPKC)
(ζ, ι/λ)
Plasma membrane
TranslocationInsP3
AGONIST
Ca
Ca
2+
2+
R
DAG
DAG ATP PDK1
cPKCSubstrate Substrate
PLC
2 PtdIns4,5P
C4C4
C4
SubstrateP
P
P
PP
P
P
P P
P
Structure of PKC isoforms and activation mechanism of the conventional protein kinase C (cPKC) isoforms.The nine genes that code for protein kinase C (PKC) fall into three families. Conventional PKCs (cPKCs) have a domain structure that has an N-terminalpseudosubstrate (PS) domain followed by regulatory domains (C1 and C2), which are responsible for binding the diacylglycerol (DAG) and Ca2+
responsible for membrane targeting, an ATP-binding C3 domain and the C4 catalytic domain. The C1 region is present as a tandem repeat (C1Aand C1B). The PS domain has an autoinhibitory function in that it associates with the catalytic site on C4, thus preventing it from phosphorylating itssubstrates. Novel PKCs (nPKCs) have a similar domain structure, except that they have a C2-like domain that does not bind Ca2+. Atypical (aPKCs)also lack a C2 domain, and the C1 domain is small and fails to bind DAG. The illustration at the bottom indicates how cPKC is activated, as describedin the text.
PKC signalling functionsThe different PKC isoforms function to regulate a widerange of cellular processes:
• PKCε functions to control the N-type Ca2+ channel(Module 3: Figure CaV2 channel family).
• PKC is one of the kinases responsible for phosphorylat-ing the transcription factor p53 (Module 4: Figure p53domains).
• PKC functions to regulate the calmodulin (CaM)-binding properties of proteins such as neuromodulinand neurogranin.
• Protein kinase C functions in the modulation of CaV1.2L-type channels from heart muscle (Module 3: FigureCaV1.2 L-type channel).
Phosphoinositide lipid signalling moleculesSome of the phosphoinositides derived from the metabol-ism of PtdIns (Module 2: Figure phosphoinositide meta-bolism) have important signalling functions that are oftenlocated in specific regions of the cell. One of the mostversatile systems is the PtdIns4,5P2 signalling cassette.Another important system is PtdIns 3-kinase signalling,where the formation of PtdIns3,4,5P3 functions as a mes-senger to activate a great variety of cellular processes. Thereis increasing evidence that some of the other lipids may alsoplay signalling roles as described in the PtdIns3P signalling
cassette, the PtdIns4P signalling cassette and the PtdIns5Psignalling cassette.
PtdIns4,5P2 signalling cassetteThe phosphoinositide PtdIns4,5P2 is widely distributedthroughout the cell. Not only is it found in the plasmamembrane, but also it occurs in the Golgi, endosomes,endoplasmic reticulum (ER) and within dense structureswithin the nucleus. This lipid has two main roles. Firstly, itis hydrolysed to form InsP3 and diacylglycerol (DAG) or itcan be phosphorylated further to form the lipid messengerPtdIns3,4,5P3. Secondly, PtdIns4,5P2 can also function as alipid messenger (Module 2: Figure PtdIns4,5P2 signalling).Such a signalling role is supported by the observation thatcell stimulation can result in highly localized increases inPtdIns4,5P2. The localized formation of this lipid messen-ger can be activated by the monomeric G proteins and inparticular the family of Rho proteins. For example, Racand Rho can function to target PtdIns4P 5-kinase I to theplasma membrane. In addition, Rho-GTP can activate thiskinase directly to form PtdIns4,5P2 (Module 2: Figure Rhosignalling).
There is increasing evidence that a variety of cellularprocesses are activated in response to a change in the levelof this lipid:
• PtdIns4,5P2 regulation of actin remodelling
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �48
Module 2: Figure PtdIns4,5P2 signalling
PtdIns PtdIns4P
InsP
Ion channels (Agonist-induced hydrolysis of PtdIns4,5P activates TRPV1 but inhibits potassium channels)
Exocytosis(Primes vesicles)
Endocytosis (Recruits AP-2 toform clathrin-coated pits)
Phagocytosis
Cytoskeleton
PLD
Rac Rho Arf PA
DAG Ca
Ca
2+
2+
PA SPHK S1P
PtdIns4,5P
PtdIns3,4,5P
2
2
3
3
+
Multiple signalling functions of PtdIns4,5P2.PtdIns4,5P2 has multiple signalling functions in the plasma membrane. It is the lipid precursor used for the generation of second messengers such asInsP3, diacylglycerol (DAG) and PtdIns3,4,5P3. It functions in the regulation of the phospholipase D (PLD) signalling pathway. In addition, PtdIns4,5P2can function as a messenger to control a whole variety of cellular systems, including the cytoskeleton, phagocytosis, exocytosis, endocytosis and ionchannels. All of these processes are sensitive to changes in the membrane level of this lipid.
• PtdIns4,5P2 regulation of membrane trafficking and en-docytosis
• PtdIns4,5P2 regulation of exocytosis• PtdIns4,5P2 activation of phospholipase D• PtdIns4,5P2 regulation of ion channels and exchangers• PtdIns4,5P2 function in focal adhesions• PtdIns4,5P2 regulation of phagocytosis
PtdIns4,5P2 regulation of actin remodellingThere are a number of cellular processes, such as chemo-taxis, locomotion, phagocytosis, shape change and cy-tokinesis, where there are rapid changes in the cytoskel-eton in response to external stimuli such as growth factorsand cytokines. Phosphoinositides play a significant role intransmitting information from cell-surface receptors to thebattery of proteins that are responsible for actin remodel-ling. Its main actions are to uncap the barbed end of actinfilaments and to facilitate actin nucleation and polymeriz-ation by controlling a variety of actin regulatory proteins.A critical component of the signalling network is the Rhofamily of monomeric G proteins (Rho, Cdc42 and Rac)that relay information from various signalling pathwayssuch as the PtdIns 3-kinase signalling pathway to the ef-fector molecules that are directly responsible for remodel-ling the actin network. One of the signalling molecules isPtdIns4,5P2.
The signalling cascade begins when external signals act-ing through guanine nucleotide exchange factors (GEFs)
stimulate members of the Rho family of G proteins bycatalysing the exchange of GDP for GTP. The active Gproteins can then have a number of functions. For example,one function of Rac-GTP is to activate actin polymeriz-ation (Module 2: Figure Rac signalling). Similarly, Cdc42also stimulates actin remodelling (Module 2: Figure Cdc42signalling). The consequence of this remodelling varies de-pending on which G protein is operating. For example,the Cdc42 and Rac polymerization of actin results in theformation of filopodia and ruffles respectively (Module 4:Figure actin remodelling).
The formation of PtdIns4,5P2 contributes to actinremodelling by acting on a number of the mol-ecules that control actin polymerization such as theWiskott-Aldrich syndrome protein (WASP) and the re-lated Wiskott-Aldrich syndrome protein (WASP) verpro-lin homologous (WAVE) proteins. WASP and WAVE relayinformation from the upstream signalling elements to thedownstream cytoskeletal regulators. Perhaps the most im-portant example of the latter is the actin-related protein2/3 (Arp2/3) complex (containing seven strongly associ-ated subunits of which two are the actin-related proteinsArp2 and Arp3) that is responsible for catalysing the poly-merization of actin. PtdIns4,5P2 appears to have two ac-tions. It binds to and activates WASP and to Scar andthus contributes to the relay of information to Arp2/3. Inaddition, it can alter the activity of various proteins thatmodify the structure of actin. For example, it is an indirectactivator of Cdc42 and it may loosen the gelsolin caps on
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �49
the end of the barbed ends, opening up new sites for actinpolymerization. The PtdIns4,5P2 may also induce a con-formational change in vinculin that enables it to bind totalin during the formation of the focal adhesion complex(Module 6: Figure vinculin function).
PtdIns4,5P2 function in focal adhesionsThe Iγ isoform of PtdIns4P 5-kinase (PtdIns4P 5-K) ishighly localized at the focal adhesion complex, where itis tightly bound to talin to regulate the local formation ofPtdIns4,5P2 (Module 6: Figure integrin signalling).
PtdIns4,5P2 regulation of exocytosisThe synthesis of PtdIns4,5P2 has been implicated in theATP-dependent processes of priming vesicles as part ofthe exocytotic mechanism. The priming process seems todepend upon a number of steps that begin with the PtdInstransfer protein carrying PtdIns to the vesicle membranewhere it is phosphorylated to PtdIns4P by PtdIns 4-kinase(PtdIns 4-K) and then to PtdIns4,5P2 by the PtdIns4P5-kinase (PtdIns4P 5-K). It is still not clear exactly how thisPtdIns4,5P2 functions to prime the vesicle for exocytosis.
The ability of the neuronal Ca2+ sensor-1 (NCS-1) tofacilitate exocytosis may depend on its ability to activatethe PtdIns 4-kinase (PtdIns 4-K).
PtdIns4,5P2 regulation of membrane trafficking andendocytosisPtdIns4,5P2 plays a role in endocytosis by targetingclathrin-associated proteins to endocytic vesicles, leadingto the formation of clathrin-coated pits.
The PtdIns4P 5-kinase (PtdIns4P 5-K) Iβ is recruited tothe plasma membrane, where it phosphorylates PtdIns4Pto PtdIns4,5P2, which recruits the adaptor protein AP-2to form clathrin-coated pits.
PtdIns4,5P2 regulation of phagocytosisWhen macrophages engulf foreign particles, PtdIns4P 5-k-inase (PtdIns4P 5-K) Iα is recruited to the plasma mem-brane at the phagosome cup, where it induces a local pulseof PtdIns4,5P2, which is then rapidly degraded by phos-pholipase C (PLC) to leave behind diacylglycerol (DAG).This local formation of PtdIns4,5P2 seems essential for theearly process of phagocytosis.
The localized transient increase in PtdIns4,5P2 mayfunction in the initial recruitment of actin to the phagocyte.
PtdIns4,5P2 activation of phospholipase DPhospholipase D (PLD), which catalyses the hydrolysisof phosphatidylcholine (PC) to generate phosphatidic acid(PA), exists as two isoforms, PLD1 and PLD2. The PLD1is primarily located on vesicles associated with the en-dosomal/lysosomal pathway, whereas PLD2 is mainlyfound on the plasma membrane. An interaction betweenPtdIns4P 5-kinase α (PtdIns4P 5-Kα) and the PLDs en-sures that there is a local generation of PtdIns4,5P2 thatplays an important role in regulating PLD activity (Module2: Figure PLD signalling).
PtdIns4,5P2 regulation of ion channels and exchangersA number of ion channels and exchangers appear to beregulated by PtdIns4,5P2. In some cases, the hydrolysis ofPtdIns4,5P2 results in channel opening as occurs for thetransient receptor potential (TRP) vanilloid 1 (TRPV1)channel, whereas the TRPC6 channel, TRP melastatin 4(TRPM4), TRP melastatin 7 (TRPM7) and TRP melast-atin 8 (TRPM8) channels are closed (Module 2: FigurePtdIns4,5P2 regulation of TRP channels). In the case ofthe inward rectifying K+ channels, lipid hydrolysis resultsin channel opening (Module 2: Figure PtdIns4,5P2 regula-tion of K+ channels). Modulation of the TRPV1 channel isparticularly important in hyperalgesia (Module 10: Figurenociception).
Much of the information on this regulatory functionof PtdIns4,5P2 has emerged from studies on K+ chan-nels. Stimulation of phospholipase C (PLC) by G protein-coupled receptors (GPCRs) causes a reduction in the levelof PtdIns4,5P2, which results in a reduction in the activityof some of the voltage-dependent K+ (Kv) channels:
• The human ether-a-go-go-related (HERG) K+ channel(Kv11.1 channel)
• The Kv7.1 channel, which is coded for by the KCNQ1gene, is regulated by PtdIns4,5P2. This inositol lipidkeeps the channel open, but when PtdIns4,5P2 is hy-drolysed by angiotensin II, the channel closes. Suchlipid regulation is seen in adrenal zona glomerulosa cells(Module 7: Figure glomerulosa cell signalling) and alsoin the heart. Mutation of KCNQ1 causes long QT) syn-drome (LQT).
• The heteromultimer KCNQ2/3 is responsible for theM channel in neurons and has been linked to a form ofepilepsy known as benign familial neonatal convulsions.
• KCNQ4, which codes for Kv7.4 (Module 3: Tablevoltage-dependent K+ channels), is the delayed rectifierthat controls K+ efflux from sensory hair cells of the in-ner ear. KCNQ4 mutations cause autosomal dominantnonsyndromic deafness type 2 (DFNA2).
PtdIns 3-kinase signallingThe PtdIns 3-kinase signalling cassette, which generatesthe lipid second messenger phosphatidylinositol 3,4,5-tetrakis phosphate (PtdIns3,4,5P3), has multiple functionsin regulating a wide range of cellular processes such as thecontrol of glycogen metabolism, lipid synthesis, proteinsynthesis, gene transcription and cell growth, inflamma-tion, cytoskeletal rearrangement and apoptosis. With re-gard to the latter, the PtdIns 3-kinase signalling cassette hasa special function in cell survival through its contributionto the hormonal modulation of apoptosis and by enhan-cing the activity of the target of rapamycin (TOR). PtdIns3-kinase signalling is one of the major pathways used bythe insulin receptor to regulate energy uptake and storage.
This lipid signalling pathway has a special relation-ship to Ca2+ signalling in that it can regulate the way inwhich external signals can maintain information flowingthrough the inositol 1,4,5-trisphosphate (InsP3)/Ca2+ sig-nalling cassette.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �50
Module 2: Figure PtdIns4,5P2 regulation of K+ channels
InsP3
AGONIST
K+
R
Inositol
Phosphatidic acid
Inositol phosphatemetabolism
DAG
PLC
Channel open membranehyperpolarized
Channel closed membrane depolarized
PtdIns4P 5-Kinase
DAGKinase
PtdInssynthase
PtdIns 4-Kinase
2 PtdIns4,5P PtdIns4P PtdIns
PtdIns
Regulation of K+ channels by agonist-dependent hydrolysis of PtdIns4,5P2.A number of different K+ channels are activated when they bind to PtdIns4,5P2. Upon stimulation by agonists that are coupled to phospholipase C(PLC), this lipid is hydrolysed to InsP3 and diacylglycerol (DAG), and this removal causes the channel to shut. The channel will open again whenanother PtdIns4,5P2 molecule associates with the lipid-binding site. The supply of PtdIns4,5P2 is maintained by the resynthesis of PtdIns and itsrephosphorylation by PtdIns 4-kinase and then by PtdIns4P 5-kinase (Module 2: Figure InsP3/DAG recycling).
Module 2: Figure PtdIns4,5P2 regulation of TRP channels
InsP3
AGONISTTRPC6TRPM4TRPM7TRPM8TRPV1
Cations
R
Inositol
Phosphatidic acid
Inositol phosphatemetabolism
DAG
PLC
TRPM4 TRPM7 TRPM8 closes
TRPV1 opens
PtdIns4P 5-Kinase
DAGKinase
PtdInssynthase
PtdIns 4-Kinase
2 PtdIns4,5P PtdIns4P PtdIns
PtdIns
Regulation of TRP channels by agonist-dependent hydrolysis of PtdIns4,5P2.A number of transient receptor potential (TRP) channels are opened (TRPM4, TRPM7 and TRPM8) or closed (TRPV1) when they are bound toPtdIns4,5P2. Upon stimulation by agonists that are coupled to phospholipase C (PLC), this lipid is hydrolysed, and its removal causes the channel toopen (TRPV1) or closed (TRPM4, TRPM7 and TRPM8). The channel returns to the resting state when PtdIns4,5P2 re-associates with the lipid-bindingsite. The supply of PtdIns4,5P2 is maintained by the resynthesis of PtdIns and its sequential phosphorylation by PtdIns 4-kinase and PtdIns4P 5-kinase.The operation of this signalling system is particularly important in nociception (Module 10: Figure nociception).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �51
Module 2: Figure PtdIns 3-kinase signalling
PtdIns3,4,5P3
Btk
Ras Ras
Itk
PLCγ
DAG
Ca2+
PDK1/2
PKB
BAD FOXO
RacRho
Cdc42
GPCRPTKRs
IP3
GSK3S6K TOR
Cytoskeletalrearrangement
Superoxide formation
Genetranscription
Cellsurvival
Glycogenmetabolism
Proteinsynthesis
PtdIns- 4,5P2
PtdIns- 4,5P2
α,β
δ,ζPKCs
PKCs
Class IA PI 3-K
Class IB PI 3-K
p85 p110 p110γ
PP
PP
PP
Growth factor
α,β,δ
β/γp101/ p84
The PtdIns 3-kinase signalling pathway.The precursor lipid PtdIns4,5P2 is phosphorylated on the 3-position by Class I PtdIns 3-kinases (PtdIns 3-Ks) to generate the lipid second messengerPtdIns3,4,5P3. The Class 1A enzyme has regulatory subunits such as p85 that attaches the catalytic p110 subunits to the phosphorylated tyrosineresidues on the cytoplasmic domains of activated growth factor receptors. The Class 1B enzymes (p110γ), which are activated by G protein-coupledreceptors (GPCRs), translocate to the membrane by binding to the Gβγ subunit. When brought into the vicinity of the membrane, these PtdIns 3-kinasesform the 3-phosphorylated lipid messenger PtdIns3,4,5P3, to regulate a large number of processes. It binds to other signalling components suchas Btk and PLCγ (shown in pink). It stimulates phosphoinositide-dependent kinase 1/2 (PDK1/2) and protein kinase B (PKB), which activate a largenumber of downstream targets (yellow). It also activates monomeric G proteins (Rac, Rho and Cdc42) to stimulate both cytoskeletal rearrangementand superoxide radical (O2
−•) formation (shown in green).
Operation of the PtdIns 3-kinase signalling cassette canbe considered in two parts:
• Generation and metabolism of the 3-phosphorylatedlipid messengers
• Mode of action of the 3-phosphorylated lipid messen-gers
One of the important functions of this pathway is to reg-ulate cell proliferation. Since a number of human cancersare found to have mutations in certain components of thispathway, there is considerable interest in the relationshipbetween PtdIns 3-kinase signalling and cancer.
Generation and metabolism of the 3-phosphorylatedlipid messengersCells contain a number of phosphoinositides carrying aphosphate on the 3-position (Module 2: Figure phosphoin-ositide metabolism). One of these is PtdIns3,4,5P3, whichis the main second messenger operating in the PtdIns3-kinase signalling pathway. This highly phosphorylatedlipid is generated in cells following activation of eitherG protein-coupled or tyrosine kinase-coupled receptors(Module 2: Figure PtdIns 3-kinase signalling).
These 3-phosphorylated lipid messengers are metabol-ized either by the type II inositol polyphosphate 5-phos-
phatases or by phosphatase and tensin homologue deletedon chromosome 10 (PTEN).
Mode of action of the 3-phosphorylated lipidmessengersThe lipid messengers PtdIns3,4P2 and PtdIns3,4,5P3 actwithin the plane of the plasma membrane by binding to agreat variety of target proteins (Module 2: Figure PtdIns3-kinase signalling). Most of these downstream targets aresoluble proteins that translocate to the membrane by bind-ing to the lipid messengers through various lipid-bindingdomains [e.g. pleckstrin homology (PH), Phox homology(PX), C2 domains and basic amino acid regions] (Mod-ule 6: Figure modular lipid-binding domains). Once drawnon to the membrane, these target proteins are activated andfunction as effectors to control a large number of cellularprocesses.
• Contribution to liver cell signalling mechanisms to con-trol glycogen metabolism (Module 7: Figure liver cellsignalling).
• Functions in the insulin control of skeletal muscle gly-cogen synthesis (Module 7: Figure skeletal muscle E-Ccoupling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �52
• Functions in white fat cells to control lipid metabolism(Module 7: Figure lipolysis and lipogenesis).
• Functions in the control of cell proliferation (Module 9:Figure growth factor signalling).
• PtdIns 3-kinase signalling in cardiac hypertrophy is anexample where this signalling system carries out mul-tiple roles in the same cell (i.e. inhibits apoptosis, ac-tivates protein synthesis and facilitates a programmeof foetal gene transcription (Module 12: Figure hyper-trophy signalling mechanisms).
• Modulation of InsP3-induced Ca2+ release by phos-phorylation of the InsP3 receptor, and this could providea possible mechanism for the action of insulin in livercells (Module 7: Figure liver cell signalling).
• PtdIns3,4,5P3 activates the monomeric G proteins Rac(Module 2: Figure Rac signalling), Rho (Module 2: Fig-ure Rho signalling) and Cdc42 (Module 2: Figure Cdc42signalling).
• Formation of osteoclast podosomes (Module 7: Figureosteoclast podosomes).
• Contributes to the amplification of the early polaritysignalling during neutrophil chemotaxis (Module 11:Figure neutrophil chemotactic signalling).
• Functions as a regulator of autophagy (Module 11: Fig-ure autophagy).
Phosphoinositide-dependent kinase 1 (PDK1)One of the main functions of phosphoinositide-dependentkinase 1 (PDK1) is to phosphorylate protein kinase B(PKB), which translocates to the membrane, where it bindsto PtdIns3,4,5P3. This binding alters the conformation ofPKB so that critical sites become available to PDK1. LikePKB, PDK1 has pleckstrin homology (PH) domains thatalso bind to the 3-phosphorylated lipid messengers thatserve to activate their kinase activity. PDK1 is also re-sponsible for phosphorylating and activating other sig-nalling molecules such as ribosomal S6 protein kinase 1(S6K1) and atypical protein kinase Cζ (PKCζ).
Protein kinase B (PKB)Protein kinase B (PKB), which is also known as Akt, isa serine/threonine protein kinase. PKB has three mem-bers (PKBα, PKBβ and PKBγ) that are activated througha two-stage process. Firstly, it translocates to the mem-brane by binding to either PtdIns3,4P2 or PtdIns3,4,5P3
through pleckstrin homology (PH) domains. The latterappears to be particularly important when cells are stud-ied in vivo. The next stage depends upon its interactionwith phosphoinositide-dependent kinase 1 (PDK1), whichthen completes the activation process by phosphorylat-ing PKB on Thr-308. In addition, a DNA-dependentprotein kinase (previously called PDK2) phosphorylatesSer-473. The activated PKB then functions to stimulatea variety of molecular targets, including glycogen syn-thase kinase-3 (GSK-3) (which mediates the effect ofinsulin on glycogen metabolism) and the pro-apoptoticfactor Bad. PKB also plays a role in redox signalling inapoptosis.
PKB is a key player in cell growth control, where itfunctions by phosphorylating tuberous sclerosis 1 and 2
(TSC1/2), which integrate a number of inputs that controlthe activity of the target of rapamycin (TOR) (Module9: Figure target of rapamycin signalling). PKB can alsotranslocate into the nucleus, where it phosphorylates theForkhead box O (FOXO) transcription factors (Module 4:Figure FOXO control mechanisms) that play an importantrole in regulating the cell cycle.
Ribosomal S6 protein kinase 1 (S6K1)This kinase plays an important role in regulating proteinsynthesis by phosphorylating the S6 ribosomal protein(a component of the 40S ribosomal subunit), which thenenhances the translation of those mRNA transcripts thatcontain a polypyrimidine tract at the 5
′transcriptional start
site. The control of S6K1 is complex in that it depends upona priming step that is followed by series of phosphoryla-tion events at multiple sites that are sensitive to a number ofkinases. The priming step depends on Ca2+, which appearsto act by opening up the enzyme so that it becomes sensit-ive to phosphorylation by different kinases. One of these isphosphoinositide-dependent kinase 1 (PDK1), which car-ries out the wortmannin-sensitive phosphorylation. In ad-dition there are rapamycin-sensitive sites phosphorylatedby the target of rapamycin (TOR) (Module 9: Figure targetof rapamycin signalling).
Glycogen synthase kinase-3 (GSK-3)One of the primary functions of insulin is to act throughglycogen synthase kinase-3 (GSK-3) to stimulate the con-version of glucose into glycogen by increasing the activityof glycogen synthase (Module 7: Figure skeletal muscleE-C coupling). GSK-3 is a proline-directed protein kinasethat usually requires a priming kinase to add a phosphateto its substrate before it can carry out further phosphoryla-tions. Casein kinase I (CKI) often acts as the priming kinaseas it does for β-catenin in the Wnt signalling pathway(Module 2: Figure Wnt canonical pathway).
GSK-3 also has a number of other signalling func-tions:
• It phosphorylates nuclear factor of activated T cells(NFAT) and thus contributes to the NFAT shuttle(Module 4: Figure NFAT activation).
• It contributes to the Wnt signalling pathway (Mod-ule 2: Figure Wnt canonical pathway).
• It functions in dorsoventral specification during devel-opment (Module 8: Figure dorsoventral specification).
• It is one of the kinases that phosphorylates the tran-scription factor p53 (Module 4: Figure p53 domains).
• It contributes to Myc degradation.• It phosphorylates the neuron-specific microtubule-
associated protein tau, which forms tangles in neuronsduring the onset of Alzheimer’s disease (Module 12:Figure amyloid plaques and tangles).
• It plays a significant role in cardiac gene transcription,where it has an important role in the NFAT shuttle(Module 12: Figure hypertrophy signalling mechan-isms).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �53
Module 2: Figure insulin receptor
SSSS SS
SS SSSS
PP
1
23
64
PPP
PP P
P
P
P
PPP
P
PDK1/2
PtdIns4,5P PtdIns3,4,5P
Insulin
L1
L2
α
β
IRS
IRS
972
115811621163
13281334
PKB
Glucose entry
Tyrosine kinase domain
Cysteine-rich domain
Fibronectin III repeat
Glycogen synthesis
Lipidsynthesis
Proteinsynthesis
PI 3-K
PI 3-K
2 3
p85
p85
5
Activation of the PtdIns 3-kinase signalling pathway by the insulin receptor.The insulin receptor is a homodimer that is connected together by disulphide bonds. Each monomer consists of an α-chain, which is extracellularand has the insulin-binding region. The β-chain has a single membrane-spanning region with a large intracellular region that contains the tyrosinekinase domain that is critical for the process of signal transduction that proceeds through a sequence of events as described in the text.
Tec tyrosine kinase familyAll members of this tyrosine kinase family are cytoso-lic, but they translocate rapidly to the membrane throughpleckstrin homology (PH) domains that are particularlysensitive to PtdIns3,4,5P3. Once bound to the mem-brane, these enzymes are also phosphorylated by Srcfamily tyrosine kinases such as Lyn and Src. Activa-tion thus requires both translocation to the membraneand phosphorylation on tyrosine residues. One of thefunctions of these Tec tyrosine kinases is to enhanceCa2+ signalling by maintaining the activity of phosphol-ipase Cγ (PLCγ). This enhancement of PLCγ activityby the Tec kinases represents a major point of interac-tion between the inositol 1,4,5-trisphosphate (InsP3)/Ca2+
signalling cassette and the PtdIns 3-kinase signallingcassette.
Bruton’s tyrosine kinase (Btk)Bruton’s tyrosine kinase (Btk) is one of the non-receptorprotein tyrosine kinases (Module 1: Figure non-re-ceptor tyrosine kinases) that plays an important rolein B cell maturation. One of the main functions ofBtk is to activate PLCγ2 in B cells (Module 2: Fig-ure ROS effects on Ca2+ signalling). It has a similarmode of action in the mast cell FcεRI signalling path-way (Step 7 in Module 11: Figure FcεRI mast cellsignalling).
Inactivation of Btk results in Bruton’s type X-linkedagammaglobulinaemia.
Inducible T cell kinase (Itk)As for Bruton’s tyrosine kinase (Btk) in B cells, inducibleT cell kinase (Itk) has a similar function in controllingthe maturation of T cells by activating phospholipase Cγ1(PLCγ1) (Module 9: Figure TCR signalling).
Insulin receptorInsulin has a major role to play in regulating a variety ofcellular processes, with particular emphasis on the regula-tion of energy uptake and storage. The action of insulin iscarried out by the insulin receptor, which is a disulphide-linked homodimer that belongs to the large family of re-ceptors that have tyrosine kinase domains (Module 1: Fig-ure tyrosine kinase-linked receptors). An important com-ponent of the signal transduction mechanism used by theinsulin receptor is the insulin receptor substrate (IRS),which is a classical scaffolding protein (Module 6: FigureIRS domain structure).
The function of the insulin receptor depends upon theactivation of the PtdIns 3-kinase signalling pathway thatoccurs through the sequence of events shown in Mod-ule 2: Figure insulin receptor:
1. Insulin binds to the extracellular domain to induce aconformational change in the receptor resulting in theactivation of the intracellular tyrosine kinase domains.
2. Once activated, the tyrosine kinase domains undergoautophosphorylation whereby they phosphorylate upto six tyrosine residues on the opposite β chain.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �54
3. The insulin receptor substrate (IRS), which is a scaf-folding domain (Module 6: Figure IRS domain struc-ture), attaches itself to juxtamembrane phosphotyr-osine residue 972. Once it is drawn into the vicinityof the receptor, the tyrosine kinase domain also phos-phorylates IRS on multiple residues.
4. The phosphotyrosine residues on IRS then function asdocking sites for Class IA PtdIns 3-kinase (PI 3-K).
5. The PI 3-K then phosphorylates PtdIns4,5P2 to formthe lipid second messenger PtdIns3,4,5P3.
6. The PtdIns3,4,5P3 then activates the PtdIns 3-kinasesignalling pathway (Module 2: Figure PtdIns 3-kinasesignalling) that controls a number of cellular processes:
7. It stimulates lipogenesis in white fat cells (Module 7:Figure lipolysis and lipogenesis).
8. It stimulates glucose uptake and glycogen synthesis inskeletal muscle cells (Module 7: Figure skeletal muscleE-C coupling).
9. It stimulates glycogen synthesis in liver cells (Mod-ule 7: Figure liver cell signalling).
The onset of diabetes (i.e. Type 2 diabetes) begins whencells become resistant to the action of insulin in energyuptake and storage.
PtdIns3P signalling cassettePtdIns3P, which is formed by the phosphorylation ofPtdIns by the type III PtdIns 3-kinase, has a specific roleto play in controlling a number of cellular processes:
• One of the main functions of PtdIns3P is to regulateintracellular vesicle trafficking. Most of the PtdIns3Pis found on internal membranes and particularly thosethat function in vesicle trafficking. This function is par-ticularly evident in phagosome maturation (Module 4:Figure phagosome maturation).
• Formation of PtdIns3P also has a role to play in activat-ing the isolation membrane responsible for initiating theevents of autophagy (Module 11: Figure autophagy).
• PtdIns3P plays a role in regulating the activity of theintermediate conductance (IK) channel, particularly aspart of the relationship between K+ channels and cellproliferation.
PtdIns3P levels can be reduced through the action ofthe myotubularin family of 3-phosphatases (MTM1 andMTMR1–MTMR8). These enzymes are of interest be-cause mutations in MTM1 are responsible for X-linkedrecessive myotubular myopathy and MTMR2 is mutatedin Charcot-Marie-Tooth disease 4B.
PtdIns4P signalling cassetteFor a long time, PtdIns4P was considered to be only a pre-cursor of PtdIns4,5P2, but now there are indications thatit might have a signalling role to regulate trafficking at theGolgi, where it is formed by at least two PtdIns 4-kinases(PtdIns 4-Ks). The small GTPase ADP-ribosylation factorArf-1 recruits the type III PtdIns 4-kinase to the Golgi. Inaddition, the Golgi also has a PtdIns 4-kinase α that pro-duces the PtdIns4P that has a specific role to associate with
activating protein 1 (AP-1) to regulate the trafficking ofclathrin-coated vesicles through the trans-Golgi network.
PtdIns5P signalling cassettePtdIns5P has been implicated in several signalling events.It may modulate the PtdIns 3-kinase signalling pathwayby interfering with the metabolism of the lipid messengerPtdIns3,4,5P3. It may also have a signalling role within thenucleus to control the response to DNA damage. PtdIns5Plocated on chromatin may provide an anchor to bind in-hibitor of growth family, member 2 (ING2), which thenresults in acetylation of the p53 tumour suppressor.
Multipurpose inositol polyphosphate signallingpathwayThe process of inositol phosphate metabolism generatesa large number of inositol phosphates (Module 2: Figureinositol phosphate metabolism). Many of these are meta-bolic intermediates, but some have been implicated in avariety of control functions.
Ins1,4P2
There is some evidence to suggest that Ins1,4P2 may func-tion within the nucleus to activate DNA polymerase. Thisinositol phosphate has also been implicated in Ca2+ sig-nalling and cardiac hypertrophy.
Ins1,3,4P3
This inositol phosphate has an important signalling func-tion as a negative regulator of the Ins1,3,4,5,6P5 1-phos-phatase (Module 2: Figure inositol phosphate metabolism)that controls the level of Ins3,4,5,6P4, which is an inhibitorof Ca2+-sensitive Cl− channels.
Ins1,3,4,5P4
This inositol phosphate, which is formed by phosphorylat-ing Ins1,4,5P3, has been implicated in the control of Ca2+
entry into cells. However, its mode of action is unknown.Some clues concerning its action may come from the iden-tification of GAP1IP4BP, which is one of the GTPase-activating protein (GAP) family (Module 2: Table mono-meric G protein toolkit). GAP1IP4BP normally associateswith the plasma membrane. When Ins1,3,4,5P4 binds tothe membrane-anchoring domain of GAP1IP4BP, it causesthis GAP to come off the membrane. A closely relatedGAP1m is located on the ER.
Ins3,4,5,6P4
Ins3,4,5,6P4 functions as an inhibitor of the Ca2+-sensitiveCl− channels (CLCAs) in epithelial cells, which regulatesalt and fluid secretion, cell volume homoeostasis and elec-trical excitability in neurons and smooth muscle cells. Itappears to act by preventing Ca2+/calmodulin-dependentprotein kinase II (CaMKII) from activating the channel.
Ins3,4,5,6P4 is formed by an Ins1,3,4,5,6P5 1-phos-phatase, which is activated by Ins1,3,4P3 (Module 2: Figureinositol phosphate metabolism).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �55
InsP6
There are a number of suggestions concerning the possiblemessenger role of InsP6. Cells contain high levels of InsP6
(approximately 15–100 µM), and much of this is prob-ably not in solution, but is probably attached to the phos-pholipids in membranes through electrostatic interactionswith bivalent cations. The level of InsP6 does not changemuch during acute stimulation, but its level can vary overthe long term, as occurs during the cell cycle and duringcellular differentiation. The various proposed messengerfunctions outlined below may thus depend upon highlylocalized fluctuations in specific cellular compartments.
Trafficking of vesiclesInsP6 may inhibit clathrin cage assembly by binding to ad-aptor protein (AP)-2 and -3. Such a role in membrane traf-ficking is also consistent with the observation that InsP6
binds to synaptotagmin by competing with the inositollipid-binding site. It therefore seems that InsP6 may func-tion as a negative regulator of endocytic vesicle traffic.
Such a possibility may explain the observation thatGRAB, which is a guanine nucleotide exchange factor thatacts on Rab3A, interacts with InsP6 kinase which convertsInsP6 into InsP7.
EndocytosisIn insulin-secreting β-cells, InsP6 appears to promote dy-namin I-mediated endocytosis through a mechanism thatseems to depend upon protein kinase C (PKC), which mayact to inhibit the phosphoinositide phosphatase synapto-janin, thereby raising the level of PtdIns4,5P2 that has beenimplicated in vesicle dynamics.
Regulation of Ca2+ channelsInsP6 is present in many brain regions and appears to be el-evated following neural activity. It may act by stimulatingadenylyl cyclase to produce cyclic AMP, which then in-creases the activity of L-type Ca2+ channels. Alternatively,it may activate L-type Ca2+ channels through an inhibitionof protein phosphatases. In vascular smooth muscle cells,InsP6 appears to act through a protein kinase C (PKC)-dependent pathway.
Regulation of gene transcriptionStudies on yeast have revealed that ARG82, which is anIns1,4,5P3 6-kinase that phosphorylates Ins1,4,5P3 withinthe nucleus to form Ins1,4,5,6P4, functions as a transcrip-tional regulator.
Regulation of mRNA export from the nucleusStudies on yeast indicate that InsP6 formed by an inositolpolyphosphate kinase located on the nuclear pores mayfacilitate the export of mRNA from the nucleus.
PP-InsP4
This diphosphorylated inositol phosphate has been con-sidered as an orphan signal, as its function is unknown. Itslevel declines in response to either cyclic AMP or cyclicGMP.
PP-InsP5
This diphosphoinositide may function as an energy storeas does [PP]2-InsP4.
[PP]2-InsP4
This diphosphoinositide may function as a high-energystore in that it can donate a high-energy phosphate toADP to form ATP.
Nitric oxide (NO)/cyclic GMP signalling pathwayNitric oxide (NO) is a highly diffusible messenger, whichpasses rapidly through cell membranes. It can act as asecond messenger within its cell of origin or it can dif-fuse across membranes to act on neighbouring cells as aparacrine signalling agent. NO synthesis is carried outby nitric oxide synthase (NOS), which comes in threedifferent forms: neuronal nitric oxide synthase (nNOS),inducible nitric oxide synthase (iNOS) and endothelialnitric oxide synthase (eNOS). These different isoformsshare a similar NO synthetic reaction mechanism, whichuses L-arginine as a substrate and O2 and NADPH as co-substrates to form NO. NOS regulation is very differentfor the three isoforms and is partly dependent on the waythey are located in different parts of the cell. The actionof NO is complex in that it can transmit information inmarkedly different ways. One of its actions is mediatedthrough the cyclic GMP signalling pathway, where it stim-ulates soluble guanylyl cyclase to produce the cyclic GMPthat can modify the properties of ion channels, proteinphosphatases or cyclic nucleotide phosphodiesterase. NOcan also act through the reactive nitrogen species (RNS)signalling pathways, whereby the NO alters the activityof a variety of protein targets through a nitrosylation re-action. This diverse NO/cyclic GMP signalling pathwayoperates to control the following cellular processes:
• NO/cyclic GMP and smooth muscle relaxation• NO/cyclic GMP and synaptic plasticity• NO/cyclic GMP and cardiac hypertrophy
NO synthesisNitric oxide (NO) synthesis is carried out by NO syn-thase (NOS), of which there are three isoforms namedeither after the tissues where they were first discovered,i.e. neuronal nitric oxide synthase (nNOS) and endothelialnitric oxide synthase (eNOS) or by the way in which theyare controlled, i.e. inducible nitric oxide synthase (iNOS)(Module 2: Figure NO and cyclic GMP signalling). Theexpression of these enzymes is not as restricted as theirnames imply, but are widely expressed and can coexist inmany cell types. Even though these isoforms are regulateddifferently and have different cellular locations, they allseem to use the same NO synthetic reaction mechanism.
The excessive production of NO can have patholo-gical consequences and has been linked to various diseasestates such as Huntington’s disease, Alzheimer’s diseaseand hypertension.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �56
Module 2: Figure NO and cyclic GMP signalling
NO
NO
sGC
pGC
Proteintargets
ONO-NO GS-NO Mn - NO2+
Cy-SH Cy-SNO
cGMP
CNGC
cGK IcGK II
O2-
eNOSiNOSCa2+
Ca2+
GSH Mn 2+
PDE5 GMP
S-Nitrosylation
ROSSignalling
nNOS GC
GC
GCK
C-type natriuretic factor (CNP)Guanylin
Atrial natriuretic factor (ANP) Brain natriuretic factor (BNP)
Nitric oxide (NO) and cyclic GMP (cGMP) signalling pathways.The nitric oxide (NO) signal can either diffuse in from other cells as a paracrine signal or it can be generated within the cell by different NO synthases(NOSs). The NO has two main actions. It can stimulate soluble guanylyl cyclase (sGC) to form the messenger cyclic GMP (cGMP), which can actthrough cyclic nucleotide-gated channels (CNGCs) to promote Ca2+ entry or it can activate cyclic GMP-dependent protein kinase (cGK). The othermain action is through reactive oxygen species (ROS) signalling mechanisms. Cyclic GMP is also formed by a plasma membrane guanylyl cyclase(pGC), which is part of the single membrane-spanning receptor activated by a variety of peptides such as atrial natriuretic peptide (ANP), brain typenatriuretic factor (BNP), C-type natriuretic factor (CNP) and guanylin.
NO synthetic reaction mechanismThe different nitric oxide synthase (NOS) enzymes func-tion as homodimers, which are arranged in a head-to-headorientation with the N-terminal oxidase domain of onemonomer lined up alongside the C-terminal reductase do-main of its neighbour. The substrates for the enzymaticreaction mechanism are L-arginine, oxygen and NADPH,which combine to form citrulline with the liberation ofNO (Module 2: Figure NO synthase mechanism). NOSregulation is complicated because each isoform appears tobe regulated by different mechanisms.
Endothelial nitric oxide synthase (eNOS)As its name implies, endothelial nitric oxide synthase(eNOS) was first described in endothelial cells, where itgenerates NO in response either to agonists such as acet-ylcholine and bradykinin that elevate Ca2+ or to bloodflow-induced shear stress. It is now evident that eNOS isexpressed in many different cell types (lung epithelial cells,blood platelets, cardiac myocytes and hippocampal neur-ons). It has a complex regulation, which is very dependenton its attachment to caveolin, one of the proteins in cave-olae (Module 6: Figure caveolae molecular organization),where it contributes to their signalling function. One ofthe key regulators of eNOS is Ca2+, which acts throughcalmodulin (CaM) to stimulate the enzyme to release NO(Module 2: Figure eNOS activation).
The NO released from endothelial cells diffuses out toregulate smooth muscle cell contraction and hence controlsblood pressure, smooth muscle cell proliferation, aggrega-tion of blood platelets and leucocyte adhesion. Studies onNO/cyclic GMP and cardiac hypertrophy have revealedthat expression of eNOS in endothelial cells can inhibithypertrophy in neighbouring cardiac cells. Given its cent-ral role in regulating so many cellular processes, alterationsin the endothelial production of NO have been implicatedin many disease states, such as hypertension, diabetes andhypercholesterolaemia.
Inducible nitric oxide synthase (iNOS)Inducible nitric oxide synthase (iNOS), which is alsoknown as immunocyte NOS, was first described in mac-rophages, where its expression is up-regulated by inflam-matory mediators. Although macrophages are the maincells that express iNOS, it is also found in other cell types(cardiac cells, vascular smooth muscle cells and glial cells).Unlike the other isoforms, the activation of iNOS doesnot require an elevation of Ca2+. However, iNOS doesbind calmodulin (CaM), which is essential for its activa-tion. Since the enzyme is constitutively active, its primaryregulation depends upon its induction by inflammatory re-ceptors such as those that respond to interferon-γ (IFN-γ)or lipopolysaccharide (LPS). As large amounts of enzymeare produced, this is a high-output pathway capable of
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �57
Module 2 : Figure NO synthase mechanism
Reductase domain
Reductase domain
Oxidase domain
Oxidase domain
NADPHFADFMN
Haem
Arginine
NOCitrulline
S
O2
1177
CAM
CAM
Ca2+
BH4
Nitric oxide synthase (NOS) reaction mechanism.The two nitric oxide synthase (NOS) monomers are lined up alongside each other so that the reductase domain of one functions together withthe oxidase domain of its neighbour. The enzyme dimer also functions as a scaffold to organize the other components of the reaction mechanismsuch as the bound cofactors [flavin–adenine dinucleotide (FAD), flavin adenine mononucleotide (FMN), haem and tetrahydrobiopterin (BH4)] andthe tightly bound prosthetic group calmodulin (CaM). The formation of NO is driven by an NADPH-dependent electron flux that passes from thereductase towards the oxidase domain. The attached haem is the terminal electron acceptor, which binds the oxygen that is inserted into arginine toform the hydroxyarginine that decays to release NO. One of the important regulators of NOS is calmodulin, which is constitutively active in inducibleNOS (iNOS), but requires an elevation of Ca2+ for both neuronal NOS (nNOS) and endothelial NOS (eNOS). One consequence of increasing theconcentration of Ca2+ in cells is therefore to increase the formation of NO.
delivering NO for prolonged periods as part of the cellsdefence against invading micro-organisms. As such, it doesnot strictly function as a messenger. However, the largeproduction of NO at the sites of inflammation will spillover to affect neighbouring cells. The large up-regulationof iNOS may account for the fall in blood pressure duringendotoxic shock.
Neuronal nitric oxide synthase (nNOS)Neuronal nitric oxide synthase (nNOS) was first describedin neurons, but has since been found in other cell types suchas skeletal muscle, which has an alternatively spliced vari-ant that has a 34-amino-acid insert between exons 16 and17. In both neurons and muscle, nNOS is closely associatedwith the plasma membrane where it binds to various pro-teins through its PDZ domains. In neurons, nNOS bindsto the postsynaptic density (PSD) proteins such as PSD-93 and PSD-95 (Module 10: Figure postsynaptic density),whereas in skeletal muscle it interacts with α1-syntrophin(a binding partner of dystrophin).
Cyclic GMP signalling pathwayThe cyclic GMP signalling pathway (Module 2: FigureNO and cGMP signalling) is governed by the secondmessenger cyclic GMP, which is synthesized by guanylyl
cyclase (GC). The latter comes in two different forms:there are the soluble GC (sGC) and membrane-boundparticulate guanylyl cyclases (pGCs). Many of the sig-nalling functions of cyclic GMP are carried out by cyclicGMP-dependent protein kinase (cGK). In addition, cyc-lic GMP can act directly to open cyclic nucleotide-g-ated channels. Cyclic GMP hydrolysis is carried out by acyclic GMP-specific phosphodiesterase (PDE5). Throughthese different signalling pathways, cyclic GMP func-tions to regulate a diverse collection of cellular processes.There are those where cyclic GMP mediates the actionof NO:
• NO/cyclic GMP and smooth muscle relaxation• NO/cyclic GMP and synaptic plasticity• NO/cyclic GMP and cardiac hypertrophy
In those cases where cyclic GMP is formed by the pGC,cyclic GMP acts independently of NO to regulate cellularprocesses such as phototransduction (Module 10: Figurephototransduction overview). Certain strains of Escheri-chia coli, which secrete the STa toxin, increase intestinal se-cretion and cause diarrhoea by activating the cyclic GMPsignalling pathway by stimulating the particulate guanylylcyclase C (pGC-C) receptor that is normally activated byguanylin (Module 7: Figure intestinal secretion).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �58
Module 2: Figure eNOS activation
NO
Caveolin
Caveolin
NADPH NADPArginine Citrulline
Ca2+
CAM
CAM
O2
FAD
FAD
FMN
FMN
Haem
Heme
BH4
BH4
eNOS
eNOS
Inactive
Active
Palmitic acid
Myristic acid
Calcium-induced NOS activation
Ca2+-dependent activation of endothelial nitric oxide synthase (eNOS).An important feature of eNOS is its location on the membrane of caveolae (Module 6: Figure caveolae molecular organization). Its membranelocalization is facilitated by an N-terminal myristic acid and by two palmitic acid residues attached to two cysteine residues (Cys-15 and Cys-26),whereas its association with the caveolae depends upon its attachment to caveolin, which is responsible for keeping the enzyme inactive underresting conditions. There appears to be a competition between caveolin and calmodulin (CaM) for the caveolin-binding site (amino acids 350–358)on eNOS. In the absence of Ca2+, caveolin dominates, but when Ca2+ increases and binds to CaM, the latter relieves the inhibitory effect of caveolin,and the enzyme is activated to generate NO from arginine using oxygen and NADPH as co-substrates.
NO/cyclic GMP and synaptic plasticityThe enzymes responsible for NO formation and its actionare richly expressed in the nervous system. Although theprecise function of NO is still debated, there are indica-tions that it might function in synaptic plasticity by con-tributing to cerebellar cell long-term potentiation (LTP) atthe parallel fibre/Purkinje cell synapse.
Guanylyl cyclase (GC)Guanylyl cyclase (GC) is the enzyme that synthesizes cyc-lic GMP from ATP (Module 2: Figure NO and cyclicGMP signalling). It comes in two main forms, soluble GC(sGC), which is activated by NO, and membrane-boundparticulate guanylyl cyclases (pGCs). The latter belongs to
a group of single membrane-spanning receptors that usean enzyme to transduce information. In this case, it is theguanylyl cyclase region of the cytoplasmic domain thatfunctions both as a transducer and an amplifier to generatethe second messenger cyclic GMP.
Cyclic GMP hydrolysisThe enzyme that reverses the second messenger action ofcyclic GMP by hydrolysing it to GMP is the cyclic GMP-specific phosphodiesterase (PDE5), which can be activatedby cGKI, thus setting up a negative-feedback loop. Excess-ive signalling through the cyclic GMP signalling pathwaywill thus be curtailed through this ability of cyclic GMPto enhance its own hydrolysis. PDE5 is of considerableinterest as it is the target of Viagra.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �59
Cyclic GMP-dependent protein kinase (cGK)There are two cyclic GMP-dependent protein kinases(cGKs): cGKI and cGKII. The cGKI comes in two al-ternatively spliced forms, cGKIα and cGKIβ. These cGKsare serine/threonine protein kinases that exist as homodi-mers that are held together by leucine zippers in their N-terminal domains, which fold over to inhibit the catalyticdomain. The binding of cyclic GMP to a regulatory siteinduces a conformational change that relieves this inhibi-tion, thus enabling the catalytic domain to phosphorylateits substrates.
cGK targetsThese cGKs are targeted to specific sites in the cell. TheN-terminal domain is responsible for targeting cGKIα andcGKIβ to specific cellular regions as is particularly import-ant for smooth muscle relaxation mediated by nitric oxide(NO) and cyclic GMP (Module 7: Figure smooth musclecell cGMP signalling).
The cGKII has an N-terminal Gly-2 myristic acidresidue that serves to target it to the plasma membrane.
cGK protein substratesThe cGMP-sensitive target proteins regulated by cGKsinclude inositol 1,4,5-trisphosphate receptor-associatedcGKI substrate (IRAG), large conductance (BK) chan-nels, cGMP-specific phosphodiesterase (PDE5), cerebellarG substrate, vesicle-associated membrane protein (VASP)and telokin.
Inositol 1,4,5-trisphosphate receptor-associated cGKIsubstrate (IRAG)Inositol 1,4,5-trisphosphate receptor (InsP3R)-associatedcGKI substrate (IRAG) is located on the endoplasmicreticulum (ER), where it appears to associate with theInsP3R. When it is phosphorylated by cGKIβ, IRAGacts to inhibit channel opening (Module 7: Figure smoothmuscle cell cGMP signalling).
Reactive nitrogen species (RNS) signallingOne of the ways by which NO functions in cells is throughan S-nitrosylation reaction. This covalent modificationresults from the addition of NO to reactive cysteineresidues on specific target proteins. NO does not reactdirectly with these proteins, but is first of all convertedinto reactive nitrogen species (RNS) are then respons-ible for carrying out the nitrosylation reaction. Thesenitrosylation-sensitive signalling pathways carry out manyof the functions of NO, and there is growing evidence fornitrosylation dysfunction in disease.
Reactive nitrogen species (RNS)The reactive nitrogen species (RNS) that carry out thenitrosylation reaction are formed when NO interactswith various acceptors such as superoxide radical (O2
−•),cysteine (Cys), glutathione (GSH) or transition metal ions(Mn+, e.g. Fe3+ or Cu2+) (Module 2: Figure NO and cGMPsignalling):
NO + O2− → ONO-NO
NO + Mn+ → Mnn+-NO
NO + GSH → GS-NONO + Cys → Cys-NO
Nitrosylation reactionThe S-nitrosylation reaction depends upon the transferof NO from one of the reactive nitrogen species (RNS)to a peptidyl cysteine thiol group of target proteins (RS)(Module 2: Figure NO and cGMP signalling):
RS + ONO-NO → RS-NORS + Mn+-NO → RS-NORS + GS-NO → RS-NORS + Cys-NO → RS-NO
Specificity is determined by the fact that the target pro-teins (RS) have hyperreactive cysteine groups where thethiol moiety exists as a thiolate anion due to the presenceof positively charged amino acids in the immediate vicinityof the protein chain.
Redox signallingCells have evolved a sophisticated mechanism of intracel-lular signalling based on localized changes in the oxidationstate of specific proteins. The internal environment of cellsis normally highly reduced. Certain forms of stress are as-sociated with an increase in the oxidative state, and thiscan induce apoptosis. It is also well known that certainphagocytic cells, such as neutrophils, can rapidly generatesuperoxide radical (O2
−•) and hydrogen peroxide (H2O2)that are used to kill other cells during inflammatory re-sponses. In addition to these pathological effects, there isincreasing evidence that the redox system has been adaptedto perform a variety of signalling functions and can mod-ulate the activity of other signalling pathways. As such,they can control many cellular processes, including cellproliferation, apoptosis and cellular senescence. Some ofthese effects are exerted through a two-way interactionwith Ca2+ signalling. For example, redox signalling canhelp to promote the tyrosine phosphorylation events thatgenerate many signalling cascades and it can modulate theactivity of the ryanodine receptors (RYRs) and inositol1,4,5-trisphosphate receptors (InsP3Rs) that release Ca2+.Conversely, Ca2+ can stimulate redox signalling, particu-larly within the mitochondrion, indicating that there aredynamic interactions operating between these signallingpathways.
There are two main types of redox signalling. The firsttype is reactive oxygen species (ROS) signalling, whichdepends on the formation of ROS. The second type isreactive nitrogen species (RNS) signalling, which is carriedout by RNS and is linked to the nitric oxide (NO)/cyclicGMP signalling pathway.
Reactive oxygen species (ROS) signallingROS signalling has all the hallmarks of a classical signallingmechanism. The second messengers are the reactive oxy-gen species (ROS) formed in response to many agonists.Reactive oxygen species (ROS) formation depends on thestimulation of a NADPH oxidase that removes an electronfrom NADPH and adds it to oxygen to create superoxide
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �60
Module 2: Figure summary of redox signalling
H2
O O H2O2
_2
S
S
OHOH
SH
SH
Reduction
Oxidation
SOD Catalase
Cellular response
Messenger formation
Messengermetabolism
Messenger action
Recovery
SIGNAL REDOXSIGNALLING PATHWAY
Oxidation-sensitive processes
R
Summary of the main features of redox signalling.The sequential processes that constitute redox signalling begin with an external signal activating a receptor (R) that then generates reactive oxygenspecies, such as superoxide radical (O2
−•), which is then converted into hydrogen peroxide (H2O2) by superoxide dismutase (SOD). O2−• and H2O2
diffuse into the cell where they exert their messenger action by stimulating the oxidation of specific proteins, sometimes forming an internal disulphidebond. Recovery is carried out by various enzyme systems that return the target protein to its reduced state. The oxidized protein acts to stimulate avariety of cellular processes.
radical (O2−•), which is one of the ROS found in cells.
Superoxide dismutase (SOD) rapidly converts the O2−•
into hydrogen peroxide (H2O2), which is one of the mainmessenger molecules used by the redox signalling pathway(Module 2: Figure summary of redox signalling).
Rapid reactive oxygen species (ROS) metabolism en-sures that H2O2, like other intracellular messengers, has ashort half-life. This metabolism of H2O2 is carried out bya range of enzymes, including catalase, glutathione peroxi-dase (GPx) and peroxiredoxin (Prx). Since the inside of thecell is a highly reducing environment, the last two enzymescan draw upon a large reservoir of reducing equivalents inorder to metabolize the ROS. The cell has a large redoxbuffer capacity in the form of glutathione, which func-tions to maintain the redox balance in the cell. The factthat H2O2 is metabolized so rapidly means that its site ofaction is highly localized close to its site of production.
The reactive oxygen species (ROS) messenger action ofH2O2 depends upon its ability to react with the cysteineresidues of a certain group of target proteins. The latter aremarked out by virtue of having hyperreactive thiol groupsthat are rapidly oxidized to form a disulphide bond. Therecovery from this oxidized state back to a fully reducedthiol group is carried out by the glutaredoxin and/or thethioredoxin systems.
Finally, the oxidized target proteins activate a number ofoxidation-sensitive processes that bring about a number of
cellular responses such as gene activation, modulation ofion channels and the activity of other signalling pathways[mitogen-activated protein kinase (MAPK) cascade andCa2+ signalling]. It is therefore not surprising to find arole for redox signalling in proliferation and cancer.
Reactive oxygen species (ROS)Reactive oxygen species (ROS) is a collective termthat refers to those oxygen species [superoxide (O2
−•),hydrogen peroxide (H2O2) and the hydroxyl radical(OH•)] that are more reactive than ground-state oxygen[Module 2: Figure reactive oxygen species (ROS)]. Theseare sometimes considered synonymous with free radicals,but not all ROS are free radicals. The latter are definedas atoms or molecules that contain one or more unpairedelectrons. With regard to signalling, it is the O2
−• andH2O2 that appear to be the most important messengers.Also, there are indications that these two messengers mayperform different functions.
There are two important sources of reactive oxygen spe-cies (ROS): one is at the plasma membrane and the otheris within the mitochondria (Module 2: Figure sites of ROSformation).
Superoxide (O2−•)
The superoxide radical (O2−•) that is formed by the
one-electron reduction of O2 is short-lived (half-life of
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �61
10−6 s) in that it rapidly dismutates into hydrogen peroxide(H2O2). This conversion can occur spontaneously, but thereaction is greatly accelerated by the enzyme superoxidedismutase (SOD).
Hydrogen peroxide (H2O2)Much attention is focused on hydrogen peroxide (H2O2)because it appears to be the primary messenger moleculefunctioning in the redox signalling pathway. Since it has nounpaired electrons, H2O2 is not a free radical and thus isnot a particularly powerful oxidizing agent. This meansthat it can function as a messenger by diffusing awayfrom its site of action to interact with more distant tar-gets. However, its sphere of influence is restricted by itsshort half-life, which is determined by the rapid reactiveoxygen species (ROS) metabolism of H2O2.
Hydroxyl radical (OH•)While H2O2 is relatively benign, it can be converted intothe highly toxic hydroxyl radical (OH•) through a reduc-tion process catalysed by transition metals (Fe3+ or Cu2+).OH• has a half life of 10−9 s indicative of its very highreactivity in that it reacts immediately and indiscrimin-ately with the first molecule it finds. Much of the oxidativedamage cause by ROS is mediated by OH•.
Superoxide dismutase (SOD)A family of metalloproteinases that converts superox-ide radical (O2
−•) into hydrogen peroxide (H2O2) (Mod-ule 2: Figure plasma membrane ROS formation):
2O2−• + 2H+ → H2O2 + O2
There are four families of SOD enzymes:
• Copper/zinc-containing superoxide dismutases(CuZnSODs)
• Manganese-containing superoxide dismutases (MnS-ODs)
• Copper-containing superoxide dismutases (CuSODs)• Iron-containing superoxide dismutases (FeSODs)
Expression of MnSOD is regulated by the FOXO3atranscription factor (Module 4: Figure FOXO controlmechanisms).
Mutation of SOD is the cause of amyotrophic lateralsclerosis (ALS), which is a debilitating and progressiveneurological disease.
Reactive oxygen species (ROS) formationReactive oxygen species (ROS) are formed at two mainsites: there is plasma membrane reactive oxygen species(ROS) formation and mitochondrial reactive oxygen spe-cies (ROS) formation (Module 2: Figure sites of ROSformation). This production of ROS appears to be highlylocalized suggesting the existence of reactive oxygen spe-cies (ROS) microdomains.
Plasma membrane reactive oxygen species (ROS)formationA large number of receptors responding to external sig-nals stimulate the formation of reactive oxygen species
(ROS), including cytokine receptors (tumour necrosisfactor α, interleukin-1 and interferon-γ), growth factorreceptors [platelet-derived growth factor (PDGF), epi-dermal growth factor (EGF) and basic fibroblast growthfactor (bFGF) receptors] and G protein-coupled receptors(GPCRs) (5-hydroxytryptamine, bradykinin, angiotensinII, thrombin and endothelin receptors). Just how all thesereceptors are coupled to the formation of ROS is still some-what uncertain, but there is increasing evidence that themechanism might resemble that found in phagocytic cells.
In phagocytes, external signals acting through cell-surface receptors stimulate the PtdIns 3-kinase signallingpathway to form the lipid messenger PtdIns3,4,5P3
(Module 2: Figure PtdIns 3-kinase signalling), which thenacts through Rac to stimulate NADPH oxidase. This mul-ticomponent enzyme uses NADPH as an electron donorto carry out the first step of ROS formation i.e. the re-duction of oxygen to superoxide radical (O2
−•) (Module2: Figure plasma membrane ROS formation). In additionto this activation through Rac, it seems that the enzymecan also be regulated through diacylglycerol (DAG) andCa2+ acting through protein kinase C (PKC), which phos-phorylates p47phox, one of the cytoplasmic components ofNADPH oxidase, to bring about the assembly of the mul-ticomponent complex. Non-phagocytic cells that generateROS for signalling use a similar, but genetically distinct,NADPH oxidase called Nox1, which plays a significantrole in redox signalling in proliferation and cancer.
In phagocytes, the O2−• is released to the outside of the
cell, where it is able to attack invading micro-organisms,whereas, in non-phagocytic cells, it functions as a mes-senger to activate a signalling cascade on the inside ofthe cell (Module 2: Figure plasma membrane ROS form-ation). Superoxide dismutase (SOD) rapidly converts theO2
−• into hydrogen peroxide (H2O2), which appears tobe the primary messenger molecule of this redox signallingpathway.
NADPH oxidaseThe NADPH oxidase (NOX/DUOX) family (Module 2:Table redox signalling components) consists of a numberof enzymes with different cellular locations.
NOX2, also known as gp91phox, has been described bestfor phagocytes, where it is made up of a number of sub-units. The catalytic component is cytochrome b558, whichis a heterodimer formed from gp91phox and p22phox. Inaddition to this heterodimer, which is located in the mem-brane, there are cytoplasmic components (e.g. p47phox,p67phox, Rap1A and Rac) that play a role in regulatingenzyme activity. The DUOX (dual oxidase) enzymes aresensitive to Ca2+ and play an important role in interactingwith the Ca2+ signalling pathway (Module 2: Figure ROSeffects on Ca2+ signalling).
Mitochondrial reactive oxygen species (ROS)formationMost of the electrons that enter the electron transportchain are transferred to oxygen in an orderly manner, butthere is always a 1–2% leakage during which an electron istransferred directly to oxygen to form superoxide (O2
−•)and this is the source of mitochondrial reactive oxygen
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �62
Module 2: Table redox signalling componentsThe major components of the redox signalling pathway
Component CommentNADPH oxidases (NOXs)
NOX1 Inducible enzyme found in colonand smooth muscle
NOX2 (gp91phox) Major NOX in phagocytesNOX3 Foetal kidneyNOX4 WidespreadNOX5 Brain, spleen and spermDUOX1 (dual oxidase 1) A Ca2+-sensitive isoformDUOX2 (dual oxidase 2) A Ca2+-sensitive isoform
NOX/DUOX regulatory factorsp47phox
p67phox
p40phox
p22phox
NOXO 1NOXA 1Rac1/Rac2
ROS metabolismSuperoxide dismutase (SOD)Catalase Localized in peroxisomesGlutathione peroxidase (GPx) Localized in cytosol and
mitochondriaPeroxiredoxin (Prx)
Prx I–IV (2-Cys) I and II in cytosol; III inmitochondria; IV in endoplasmicreticulum
Prx V (atypical 2-Cys)Prx VI (1 Cys)
Thiol-containingproteins/peptidesGlutathione (GSH)Glutaredoxin (Grx)Thioredoxin (Trx)
Trx-1Trx-2 Mitochondria-specific
ReductasesGlutathione reductaseGlutaredoxin reductaseThioredoxin reductase (TrxR)Sulphiredoxin (Srx) Catalyses reduction of
hyperperoxidized proteins
species (ROS) (Module 5: Figure mitochondrial Ca2+ sig-nalling). This orderly electron transfer to oxygen occursduring energy metabolism, where oxygen is reduced towater by accepting four electrons (e−) from cytochrome coxidase:
O2 + 4e− + 4H+ → 2H2O
Mitochondrial energy metabolism is inherently danger-ous, because the electron transport chain is somewhatleaky in that some of the molecular oxygen is divertedinto the formation of the superoxide radical (O2
−•), whichis then converted into hydrogen peroxide (H2O2) and thehydroxyl radical (OH•). These mitochondrial ROS mayplay an important role in apoptosis by acting synergist-ically with Ca2+ to stimulate the formation of the mito-chondrial permeability transition pore (MTP) (Module 5:Figure mitochondrial Ca2+ signalling). The formation ofmitochondrial ROS appears to be highly localized in thatsmall superoxide flashes have been recorded in single mi-tochondria. This is another example of reactive oxygenspecies (ROS) microdomains.
The generation of ROS by mitochondria might be aregenerative process in that a local release of ROS in car-diac myocytes causes a rapid mitochondrial depolarization
due to formation of a MTP and a concomitant increase inintrinsic ROS production, i.e. a process of ROS-inducedROS release (RIRR). This RIRR often occurs synchron-ously and reversibly among long chains of adjacent mito-chondria, suggesting a co-operative mechanism.
Redox balanceThe redox balance of the cell is maintained by energy meta-bolism, primarily the pentose phosphate shunt that feedsreducing equivalents into the cell in the form of NADPH.The latter is then used to maintain redox buffers such asglutathione (GSH) (Module 2: Figure GSH/GSSG couple)and thioredoxin (Trx) in their reduced forms. The concen-tration of GSH in the cytoplasm lies within the 1–10 mMrange, with over 99% existing as the reduced GSH form.The 2GSH/GSSG redox couple is thus a measure of theredox balance in the cell. A similar balance exists for theoxidized and reduced forms of thioredoxin (Trx). The re-duced GSH and Trx are used for a number of reductiveprocesses such as the metabolism of hydrogen peroxideby glutathione peroxidase (GPx) (Module 2: Figure H2O2
metabolism) or as a source of reducing equivalents for theglutaredoxin system (Module 2: Figure recovery of proteinoxidation). The GSSG (the oxidized form of glutathione)is converted back into GSH by glutathione reductase.
Similar redox control enzymes and buffers are located inboth the mitochondria and within the lumen of the endo-plasmic reticulum (ER). Like the cytoplasm, the mitochon-drial matrix maintains a reducing environment and usessimilar enzymatic mechanisms to control the ROS eman-ating from the electron transport chain. The ER, however,is somewhat different in that the GSH/GSSG ratio is closeto 1 and this more oxidizing environment is necessary forthe formation of the disulphide bonds that are an integralcomponent of the extracellular proteins that are processedand packaged within the ER.
Since the cytoplasm and the ER lumen have differentredox potentials, there is a redox potential gradient acrossthe ER membrane and this might be used to modulateCa2+signalling by altering the activity of the ion channelsthat release Ca2+.
Reactive oxygen species (ROS) metabolismLike all other intracellular signalling molecules, ROS aremetabolized rapidly. Superoxide dismutase (SOD) rapidlyconverts superoxide radical (O2
−•) into hydrogen perox-ide (H2O2), which is then metabolized by a number ofenzyme systems including catalase, glutathione peroxidase(GPx) and peroxiredoxin (Prx) (Module 2: Figure H2O2
metabolism).With so many enzyme systems co-operating to meta-
bolize H2O2, it is likely that this messenger will have ahighly restricted sphere of influence localized to its site ofproduction either at the plasma membrane or within themitochondrion.
CatalaseCatalase is a haem-containing protein that decomposeshydrogen peroxide (H2O2) to water and oxygen (Mod-ule 2: Figure H2O2 metabolism).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �63
Module 2: Figure reactive oxygen species (ROS)
Oxygen
Superoxide
Hydrogenperoxide
Hydroxylradical
REACTIVE OXYGEN SPECIES (ROS)
2O
2O
O OH H
O H
O O
O O
e-
e-
e-
.
.-
OH
H O2 2
Unpaired electronsPaired electrons
Formation and metabolism of the reactive oxygen species (ROS).To understand the properties of reactive oxygen species (ROS) and how they are formed, it is best to begin with oxygen. Oxygen is a strong oxidizingagent in that it has two unpaired electrons (e−; red dots), which have parallel spins (i.e. they spin in the same direction as indicated by the two blackarrows) and occupy separate π -antibonding orbitals. Given that they have these unpaired electrons, oxygen qualifies as a free radical. However,oxygen is relatively inert because, in order to react with another molecule, it has to accept a pair of electrons with antiparallel spins to fit into the emptyspaces in the π orbitals. Because of this restriction, oxygen accepts electrons (e−) one at a time and this leads to the formation of the different ROS.
Most of the catalase in cells is found in peroxisomes,thus restricting the role of the enzyme in dealing with theH2O2 generated during the redox signalling mechanism atthe plasma membrane.
Glutathione peroxidase (GPx)The glutathione peroxidase (GPx) family uses the reducingpower of glutathione to convert H2O2 into water (Mod-ule 2: Figure H2O2 metabolism).
Peroxiredoxin (Prx)The peroxiredoxins (Prxs) are a family of small antioxidantproteins that function to metabolize hydrogen peroxide(H2O2) to water, thus curtailing its messenger action. Theoperation of the catalytic cycle goes through the followingsteps (Module 2: Figure peroxiredoxin catalytic cycles):
1. H2O2 is generated near the plasma membrane whenthe PtdIns3,4,5P3 (PIP3) formed by receptor activationstimulates NADPH oxidase.
2. H2O2 interacts with the reduced cysteine residues(Cys-SH) in the N-terminal regions of the thioredoxin(Trx) dimers to form two oxidized sulphenic residues(Cys-SOH).
3. The Cys-SOH can then interact with the conservedCys-SH on the C-terminal regions of the neighbouringdimer to form two intermolecular disulphides.
4. The Prx disulphide is converted back into the reducedform by Trx, which is regenerated by thioredoxin re-ductase (Module 2: Figure recovery of protein oxida-tion).
5. The sulphenic residues formed by Reaction 2 can un-dergo hyperperoxidation by interacting with furthermolecules of H2O2 to form the sulphinic acid residues.
6. This hyperperoxidation reaction can be reversed by areaction that requires ATP catalysed by the enzymesulphiredoxin (Srx).
7. The phosphorylated intermediate is reduced back tothe reduced form of Prx by the Trx system.
Mammals express five Prxs which contain catalyticcysteine residues and use Trx as an electron donor (Mod-ule 2: Figure peroxiredoxin catalytic cycles).
The reducing equivalents derived from thioredoxin(Trx) are used to regenerate Prx-(SH)2:
Prx-S2 + Trx-(SH)2 → Prx-(SH)2 + Trx-S2
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �64
Module 2: Figure sites of ROS formation
O
O
2
2H
NADH
NAD+
Agonist
NADPH oxidasecomplex on the plasma membrane
Leakage of electronsfrom mitochondrialelectron transportchain
ROS
ROS
The two main sites of reactive oxygen species (ROS) formation.There are two main sites of ROS formation in cells. One is at the level of the plasma membrane, where an NADPH oxidase complex is activated bycell signalling pathways. The other is at the mitochondria, where ROS are produced as a result of electron leakage from the electron transport chain.As indicated, this production of ROS appears as microdomains indicating that ROS signalling might be highly localized in cells.
Module 2: Figure plasma membrane ROS formation
PI 3-K PIPPIP
PIP
2
2
3 Rac Cyt b NADPH
NADP+
OO
OH
22
22
PTEN
Superoxidedismutase
NADPH oxidase
CytokinesGrowthfactors Hormones
Gene transcription
Apoptosis
Channelmodulation
SH
SH
S
S
Reduction
Oxidation
Tyrosine phosphatase
-
Receptor-dependent reactive oxygen species (ROS) formation at the plasma membrane.The formation of reactive oxygen species (ROS) occurs in response to many external signals such as cytokines, growth factors and hormones. Inmany cases, these signals activate receptors coupled to PtdIns 3-kinase (PI 3-K) which produces PtdIns3,4,5P3 (PIP3) that then acts through Racto stimulate NADPH oxidase at the plasma membrane. An electron is removed from NADPH and transferred to oxygen to form superoxide radical(O2
−•). This O2−• is then transformed by the addition of further electrons by superoxide dismutase (SOD) to form hydrogen peroxide (H2O2). There is
a positive-feedback loop that will amplify the redox signalling pathway because one of the actions of H2O2 is to inhibit the enzyme phosphatase andtensin homologue deleted on chromosome 10 (PTEN) that hydrolyses PIP3.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �65
Module 2: Figure GSH/GSSG couple
Glu Cys Gly
Glu Cys Gly
Glu Cys Gly
SH
SS
GSH
GSSG
Oxidation
Reduction
GSH
GSSG
Grx-S
Grx-(SH)2
2S
SH2H O2
H O2S
S
*
The GSH/GSSG redox couple.GSH is a tripeptide consisting of glutamic acid, cysteine and glycine. In its oxidized state, two molecules of GSH are joined together through adisulphide bond to form GSSG. This is the most abundant redox couple in the cell. The state of this couple can be determined by measuring thehalf-cell reduction potential (E-hc). Under normal reducing conditions, this potential is high, i.e. −240 mV, and this seems to be associated with cellproliferation. Differentiation seems to occur at lower potentials (−200 mV), whereas still lower potentials of −170 mV favour apoptosis. At these lowerpotentials, where there is an alteration in the redox balance, the build-up of GSSG within the cell can reverse the operation of the glutaredoxin systemthat functions normally in the recovery of oxidation-sensitive processes. GSSG interacts with reduced glutaredoxin [Grx-(SH)2] to form oxidized Grx-S2,and this disulphide bond can be transferred to oxidize target proteins.
When Prx1 in mice is knocked out, animals develophaemolytic anaemia and malignant cancers.
Reactive oxygen species (ROS) messenger actionThe primary action of hydrogen peroxide (H2O2) is toreversibly oxidize a variety of target proteins with a highdegree of specificity (Steps 1–7 in Module 2: Figure revers-ible and irreversible ROS oxidations):
1. The primary action of H2O2 is to oxidize the hyperre-active cysteine to form a sulphenic acid group (-SOH),which can be metabolized further along a number ofpathways.
2. The sulphenic acid residue can interact with nitrogen ona neighbouring serine residue to form an intramolecularcyclic sulphenyl amide as occurs during the oxidation ofprotein tyrosine phosphatases (Module 2: Figure ROSformation and action).
3. The sulphenic acid residue can be converted into anintramolecular disulphide bond with the elimination ofwater.
4. The sulphenic acid residue can interact with glutathione(GSH) to form an intermolecular disulphide bond.
5. The sulphenic acid residue can undergo hyperperoxid-ation by interacting with another molecule of H2O2 toform a sulphinic acid intermediate (Cys-SO2H).
6. The sulphinic acid intermediate undergoes further hy-perperoxidation to form the sulphonic acid intermedi-ate (Cys-SO3H).
7. The sulphinic acid group (Cys-SO2H) can be reducedby a reaction that requires ATP and is catalysed by theenzyme sulphiredoxin (Srx).
The various oxidated intermediates can be convertedback into the initial reduced state by either the thioredoxin(Trx) or the glutaredoxin (Grx) system (Module 2: Figurerecovery of protein oxidation).
These oxidation processes can occur through a num-ber of mechanisms. What is remarkable about this processis its specificity. Only a subset of proteins are modified,and within these there is a high degree of specificity inthat only certain thiols are modified. How is it that anoxidizing agent such as H2O2 is able to seek out and se-lectively modify specific target proteins? The answer liesin the fact that proteins vary considerably in their sensit-ivity to mild oxidizing agents, such as H2O2. Most of thecysteine residues in proteins have a high acidic constant
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �66
Module 2: Figure H2O2 metabolism
2GSH GSSG
GSHperoxidase
++
Prx(SH)Peroxiredoxin
PrxS++2
CatalaseH
H
H
2
2
2
O
O
O
2
2
2
O2H
H
H
O
O
O
2
2
2
2
2
2 2
+2
Hydrogen peroxide (H2O2) metabolism by different enzyme systems.Hydrogen peroxide (H2O2) can be metabolized by three main mechanisms. The enzyme catalase, which is restricted to peroxisomes, converts H2O2into water and oxygen. GSH peroxidase uses the reducing power of glutathione (GSH) to convert H2O2 into water with the formation of GSSG. Theperoxiredoxin (Prx) family is a major player in the metabolism of H2O2 through a series of catalytic cycles (Module 2: Figure peroxiredoxin catalyticcycles).
Module 2: Figure peroxiredoxin catalytic cycles
PI 3-K
TrxTrx
Trx
Trx
Srx
ATPH O
2
H O2
H O2
NOX/DUOXPIP
-SH
-SH-SH-SH
-SH
-SH
-SH-SH
-SH
-SH
S
P
P
1
2
3
4
5
6
7
S
S
S -SH
-SH-SH
-SH-SH
-SH
S
S
S
SS
S
S
O
O
O
O
OO
S
-SH
-SH-SOH
-SOH
3
OH 22
CytokinesGrowthfactors Hormones
Prx
Prx
Prx
Prx
Prx
Prx
Metabolism of H2O2 by peroxiredoxin.Peroxiredoxin (Prx) plays a major role in restricting the microdomain of hydrogen peroxide (H2O2) that forms beneath the plasma membrane by rapidlyremoving this messenger through a series of catalytic reactions as described in the text. Information adapted from Curr. Opin. Cell Biol., Vol. 17, Rhee,S.G., Kang, S.W., Jeong, W., Chang, T.-S., Yang, K.-S. and Woo, H.A., Intracellular messenger function of hydrogen peroxide and its regulation byperoxyredoxins, pp. 183–189. Copyright (2005), with permission from Elsevier; see Rhee et al. 2005.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �67
Module 2: Figure reversible and irreversible ROS oxidations
S
O
PO4
SO H
SH
SH
ATP
SH
SH
H
H
H
H
SSG
3
H O2
H O2
H O2
GSH
Sulphonic acid
SN
N
N
N
N
S
S
SOH
SH
HSulphenic acid
N
SO H
SH
H
2
Sulphinic acid
N
S
SH
H Proteinsubstrate
N *
Srx
Grx and Trx systems
H O2 2
H O2
H O2 2
H O2
H O2 2
H O2
12
3
45
6
7
8
Reactive oxygen species (ROS) messenger action through the reversible oxidation of target proteins.The target protein (blue) undergoes a variety of oxidative covalent modifications, some of which induce the conformational changes responsible formediating a variety of oxidation-sensitive processes. The target proteins that are modified by reactive oxygen species (ROS) have a hyperreactivecysteine thiolate anion (asterisk) that is highly sensitive to oxidation by H2O2 to initiate a cascade of reactions as described in the text.
(i.e. pKa values of approximately 8.5), which means thatthey are resistant to attack by H2O2. However, some of thecysteine residues, particularly those located next to posit-ively charged amino acids, have pKa values between 4 and5 and thus exist as a thiolate anion (Cys-S−), which is veryvulnerable to oxidation, and these have been referred to ashyperreactive cysteine residues (the S− group marked withan asterisk in Module 2: Figure reversible and irreversibleROS oxidations).
The sulphenic acid intermediate is somewhat unstableand can be converted into a number of intermediates byeither eliminating water or causing it to interact with GSH(Reactions 2, 3 and 4 in Module 2: Figure reversible and ir-reversible ROS oxidations). In addition, sulphenic acid canbe oxidized further by H2O2 to sulphinic acid and sulph-onic acid. The formation of sulphinic acid is reversiblethrough a reaction that requires ATP and sulphiredoxin(Srx). However, the final step to form sulphonic acid isirreversible and can result in serious damage and has beenimplicated in the ageing process. One idea is that this ir-reversible change may accumulate with time and the pro-gressive damage may result in ageing.
There is a suggestion that the hyperperoxidation reac-tions that lead to the irreversible oxidation may be avoidedby an internal reaction whereby the sulphenic acid is rap-idly converted into a sulphenylamide species by interactingwith the main-chain nitrogen atom of an adjacent serineresidue (Step 2 in Module 2: Figure reversible and irrevers-ible ROS oxidations). The formation of this sulphenyl-
amide intermediate protects against further oxidation, andthe enzyme can be reactivated by converting the intermedi-ate back into a thiol group via a mixed disulphide reactioninvolving GSH.
Two of the main signalling molecules whose activitiesare reduced by oxidation are the protein tyrosine phos-phatases and phosphatase and tensin homologue deletedon chromosome 10 (PTEN) (Module 2: Figure ROS form-ation and action). An analysis of tyrosine phosphatasestructure and function reveals the presence of a hyperre-active cysteine residue in the catalytic domain that is sens-itive to oxidants resulting in inactivation of the enzyme.Other examples of proteins that have such hypersensitiveresidues include the cell cycle regulatory enzyme Cdc25C,the Ca2+-release channels ryanodine receptors (RYRs) andinositol 1,4,5-trisphosphate receptors (InsP3Rs). The spe-cificity with which this redox signalling system exerts itseffects therefore depends on the fact that H2O2 will onlymodify hyperreactive cysteine residues that exist in thesetarget proteins of the different oxidation-sensitive pro-cesses.
Recovery of oxidation-sensitive processesLike other signalling pathways, there are mechanisms inplace for the recovery of oxidation-sensitive processesbased on the thioredoxin (Trx) and glutaredoxin (Grx)systems (Module 2: Figure recovery of protein oxidation).Although the two systems have much in common, thereare some differences, not least of which are their substratespecificity and the kinds of disulphide bonds that they
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �68
Module 2: Figure ROS formation and action
PI 3-K
PTP
PTP
PIP PIP
-SH
SHH
-SH
S
S S
N
N
PIP23 3
OH 22 PTEN
PTEN
Cytokines
Growthfactors
Hormones
Proteinkinase
Signalling protein
Signalling protein
Inactivated proteintyrosine phosphatase (PTP)
Trx
Trx
Trx
Trx
-SH
-SH
-SH
-SH
S
S
S
S
P
NOX/DUOX
1 2
Inactivated PTEN
Formation of H2O2 and its action to regulate phosphotyrosine phosphatase (PTP) and phosphatase and tensin homologue deleted on chromo-some 10 (PTEN).The hydrogen peroxide (H2O2) formed by the NADPH oxidase (NOX)/DUOX (dual oxidase) complex creates a microdomain in which it can inactivateenzymes involved in various signalling pathways: 1. The phosphotyrosine phosphatase (PTP) that dephosphorylates various signalling proteins isinactivated following its oxidation by H2O2; 2. The inositol lipid phosphatase and tensin homologue deleted on chromosome 10 (PTEN), whichdephosphorylates the lipid second messenger PtdIns3,4,5P3, is inactivated following its oxidation by H2O2. These two enzymes can be reactivatedfollowing their reduction by the thioredoxin (Trx) system (Module 2: Figure recovery of protein oxidation).
can reduce. For example, Trx is more effective at reducingprotein tyrosine phosphatase 1B than is Grx.
Trx and Grx exist in a reduced or oxidized state, andit is the former that enables them to reduce their sub-strates. In doing so, they become oxidized and have to beconverted back into a reduced state by the Trx and Grxsystems respectively. In the case of Trx, this is carried outby thioredoxin reductase. The Grx system is somewhatmore complicated in that it depends upon glutathione thatis regenerated by a glutathione reductase.
The level of Trx is markedly elevated during rheumatoidarthritis and this may influence the rate of secretion ofmatrix metalloproteinases (MMPs).
Thioredoxin reductaseThioredoxin (Trx) reductase together with thioredoxin isan important oxidoreductase system that has a signific-ant role in regulating the redox state (Module 2: Figurerecovery of protein oxidation).
Trx reductase is unusual in that it contains seleno-cysteine (SeCys) located in the C-terminal active site,which has a highly conserved -Gly-Cys-SeCys-Gly- se-quence. The N-terminal region contains one mol of flavin–adenine dinucleotide (FAD). The enzyme operates bytransferring electrons from NADPH to FAD and thenon to the active site in the C-terminus.
The level of Trx reductase is greatly increased in varioustumour cells, where it may play an important role in inhib-iting apoptosis. Reduced Trx-(SH)2 is known to bind tothe apoptosis signal-regulating kinase 1 (ASK1), but whenit is oxidized to Trx-S2, the ASK1 is released, and proceedsto induce apoptosis. An enhanced level of Trx reductasemay prevent this process by ensuring the Trx remainsreduced.
The ability of this thioredoxin system to reverseredox signalling is inhibited by Ca2+, which acts toconvert a large proportion of the reduced Trx-(SH)2
into its oxidized form. Such an action would enhancethe growth-promoting activity of the redox signallingsystem.
Thioredoxin-2 (Trx-2) is a mitochondria-specific mem-ber of the Trx family. It functions to regulate the mitochon-drial membrane potential and contributes to the inhibitionof apoptosis.
Glutathione reductaseGlutathione reductase is responsible for converting oxid-ized GSSG back into the reduced GSH (Module 2: Figurerecovery of protein oxidation):
GSSG + NADPH + H+ → 2GSH + NADP+
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �69
Module 2: Figure recovery of protein oxidation
GSH
GSSG Grx-S
Grx-(SH)
SH
SH
SH
SH
S
S
S
S
NADPH
NADP
Pentose cycle
Glucose
Trx-(SH)22
Trx-S2
TrxR-(SH)
TrxR-S2
1
1
M
M
1
1
M
M
-20
- 20
M
M
100
100
NADPH
NADP
2
2
THIOREDOXIN SYSTEM
GrxR-S2
GrxR-(SH)2
GLUTAREDOXIN SYSTEM
1-10mM
+
+
Recovery of protein oxidation by the thioredoxin and glutaredoxin systems.These two systems operate through protein disulphide oxidoreductases, which function to reduce the disulphide bond on the oxidized protein (red)back to the reduced thiol groups (blue). Thioredoxin (Trx) system: the active site on Trx is a -Cys-32-Gly-Pro-Cys-35- motif and it is these two cysteineresidues in the reduced Trx-(SH)2 form that are responsible for reducing disulphide bonds. Upon transferring the two protons to the substrate protein,the Trx becomes oxidized to Trx-S2. Before it can operate again, the Trx-S2 must be converted back into Trx-(SH)2 by thioredoxin reductase (TrxR),which extracts reducing equivalents from the NADPH formed from the pentose cycle. Glutaredoxin (Grx) system: Grx has a -Cys-Pro-Tyr-Cys- motif,which is the active site for the oxidoreduction reaction. Glutaredoxin can act both as a dithiol–disulphide oxidoreductase and as a GSH–disulphideoxidoreductase. The latter action enables Grx to reverse mixed protein disulphides (protein-SSG not shown on the figure) formed when proteinsinteract with GSH. In order to continue its reducing function, the Grx-S2 or Grx-SSG must be reduced back to Grx-(SH)2 by its interaction with GSH,which is maintained in a reduced form by glutathione reductase.
Oxidation-sensitive processesThe redox signalling pathway acts to regulate a vari-ety of oxidation-sensitive processes (Module 2: Figureplasma membrane ROS formation). The cellular pro-teins that are sensitive to oxidation are those that containhyperreactive cysteine residues. One of the main func-tions of redox signalling is to modulate the activity ofother signalling systems that contain protein compon-ents with such hyperreactive cysteine residues, e.g. Ca2+-releasing channels [inositol 1,4,5-trisphosphate receptors(InsP3Rs) and ryanodine receptors (RYRs)], transcriptionfactors, protein tyrosine phosphatases and by activatingthe mitochondrial permeability transition pore (MTP) toinduce apoptosis. As so many key signalling functions arebeing influenced, it is not surprising to find that there is arole for redox signalling in many cellular processes:
• redox signalling in proliferation and cancer• redox signalling in apoptosis• Redox signalling and DNA damage• redox signalling in vascular homoeostasis• redox signalling and gene transcription• redox signalling and modulation of Ca2+ signalling
Redox signalling in proliferation and cancerOne of the main actions of redox signalling is to con-trol growth. During the action of many growth factorsthere is an increase in the production of hydrogen peroxide(H2O2) (Module 2: Figure plasma membrane ROS form-ation), which facilitates growth factor signalling by in-hibiting tyrosine phosphatases and the tumour suppressorphosphatase and tensin homologue deleted on chromo-some 10 (PTEN). The latter inhibits the hydrolysis ofPtdIns3,4,5P3 (PIP3), which functions in cell migration,proliferation and survival. H2O2 inactivates PTEN by in-ducing a disulphide bond to form between Cys-124 in theactive site and Cys-71. This disulphide is specifically re-versed by thioredoxin. A reversible inactivation of PTENmay thus contribute to the accumulation of PIP3, whichwill thus set up a positive-feedback loop, since the forma-tion of PIP3 is responsible for stimulating the productionof H2O2.
Many cancer cells are known to use ROS to control theirproliferation. There are five human homologues of Nox1.When Nox1 is overexpressed in fibroblasts, there is anincrease in proliferation and tumour formation. The addi-tion of antioxidants can reduce the growth of cancer cells.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �70
Module 2: Figure thimerosal-induced Ca2+ signalling
Thimerosal-induced Ca2+ oscillations in oocytes.A. The normal Ca2+ oscillation induced by sperm fusion in a mouse oocyte. B–D. Examples of Ca2+ oscillations induced by addition of the oxidizingagent thimerosal at three different concentrations: B, 100 µM; C, 10 µM; D, 1 µM. Reproduced from Cheek, T.R., McGuinness, O.M., Vincent, C.,Moreton, R.B., Berridge, M.J. and Johnson, M.H. (1993) Fertilisation and thimerosal stimulate similar calcium spiking patterns in mouse oocytes butby separate mechanisms. Development 119, 179–189, with permission from The Company of Biologists; see Cheek et al. 1993.
Many cancer cells, like a number of stem cells, have en-hanced ROS defences in the form of elevated levels of GSHand thioredoxin, which make them particularly resistant toapoptosis. Thioredoxin may also play a role in increasingthe expression of the hypoxia-inducible factor 1α (HIF-1α), resulting in an increase in vascular endothelial growthfactor (VEGF) and tumour angiogenesis.
Redox signalling in apoptosisIn addition to playing a role in cell proliferation, thereis a darker side to reactive oxygen species (ROS) in thatthey can also activate apoptosis. One of the actions ofredox signalling is that it contributes to Ca2+-inducedapoptosis at the level of the mitochondria. The uptakeof Ca2+ and the resulting increase in ROS formationact synergistically to open the mitochondrial permeabil-ity transition pore (MTP) (Module 5: Figure mitochon-drial Ca2+ signalling). ROS can also increase apoptosisby stimulating the acidic sphingomyelinases (SMases)that produce ceramide (Module 2: Figure sphingomyelinsignalling).
Cells have different ways of suppressing this ROS-induced apoptosis. One mechanism is carried out byPtdIns 3-kinase signalling during which protein kinaseB (PKB) plays a prominent role through the Forkheadbox O 3a (FOXO3a) transcription factor, which actsby increasing the amount of manganese superoxide dis-mutase (MnSOD) to provide greater protection againstROS (Module 4: Figure FOXO control mechanisms).
An overactive redox signalling system may contributeto the increase in neuronal cell death that characterizesAlzheimer’s disease (Module 12: Figure astrocyte-inducedneuronal death) and Down’s syndrome.
Redox signalling and DNA damageOne of the major pathological consequences of excessROS formation is DNA damage. When this damage oc-curs during the G1 phase of the cell cycle, specific repairmechanisms function to repair the damage and they alsoinduce the process of G1 checkpoint signalling to DNAdouble-strand breaks (DSBs) (Module 9: Figure G1 check-point signalling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �71
Redox signalling in vascular homoeostasisHydrogen peroxide (H2O2) may function as anendothelium-derived hyperpolarizing factor (EDHF) thatdiffuses across to relax neighbouring smooth muscle cells.This action may be particularly important in regulating thetone of cerebral arteries.
Redox signalling and gene transcriptionThe redox signalling pathway has been implicated in thecontrol of gene transcription, particularly with regard tothe activation of nuclear factor κB (NF-κB). Whether thisactivation is due to a direct modulation of the transcriptionfactor by messengers such as H2O2 or indirectly throughactivation of other signalling pathways remains to be de-termined.
A large number of other transcription factors [activatingprotein 1 (AP-1), specificity protein 1 (SP1), c-Myb, p53and Egr-1] are redox-sensitive. Many of these have a highlyconserved cysteine residue located within their DNA-binding domains that has to be reduced in order for thefactor to bind DNA (Module 4: Figure SRF and AP-1). Intheory, therefore, such factors would be inhibited by ox-idation. There is a nuclear redox factor 1 (Ref-1) that func-tions to control transcription by reducing this cysteine.
HDAC oxidation is an important mechanism used tocontrol chromatin remodelling and gene transcription.
Redox factor 1 (Ref-1)Ref-1 plays a role in the nucleus to promote gene tran-scription (Module 4: Figure SRF and AP-1) and to protectcells against oxidative stress. There is a possibility that itmight also have a function within the cytoplasm to reducethe Rac-1-regulated production of reactive oxygen species(ROS).
Redox signalling and modulation of Ca2+ signallingThere are reciprocal interactions operating between theredox and Ca2+ signalling pathways. There are Ca2+ sig-nalling effects on redox signalling and there are redox sig-nalling effects on Ca2+ signalling.
Ca2+ signalling effects on redox signallingOne of the actions of Ca2+ is to enhance redox signallingby interfering with the recovery of the oxidation-sensitiveprocesses. Ca2+ acts by turning down the thioredoxin sys-tem by inhibiting the thioredoxin reductase that normallyswitches off redox signalling.
Redox signalling effects on Ca2+ signallingThere are numerous examples of redox signalling acting toenhance Ca2+ signalling. For example, the two Ca2+ releasechannels ryanodine receptors (RYRs) and the InsP3 recept-ors (InsP3Rs) can be activated by oxidation of key cysteineresidues. In the case of the latter, the oxidizing agent thi-merosal faithfully reproduces the Ca2+ transients normallyinduced by sperm fusion during fertilization (Module 2:Figure thimerosal-induced Ca2+ signalling).
Another example is that hydrogen peroxide (H2O2) canmarkedly enhance Ca2+ signalling by inhibiting the Srchomology 2 (SH2) domain-containing protein tyrosinephosphatase-1 (SHP-1), which normally acts to keep in
check the protein tyrosine phosphorylation cascade thatoccurs during B cell receptor (BCR) activation (Module 2:Figure ROS effects on Ca2+ signalling). This is a reciprocalinteraction because it is the increase in Ca2+ that activatesthe formation of H2O2 through a Ca2+-dependent activa-tion of dual oxidase (DUOX). This is a good example ofthe cross-talk that can exist between signalling pathways.
Mitogen-activated protein kinase(MAPK) signalling
OverviewThe multifunctional mitogen-activated protein kinase(MAPK) signalling system consists of separate pathwaysthat function to control a number of different cellular pro-cesses such as gene transcription, metabolism, motility, cellproliferation, apoptosis, synaptic plasticity and long-termmemory. These different downstream effectors are activ-ated by the final MAPK components associated with thethree main signalling pathways:
• Extracellular-signal-regulated kinase (ERK) pathway• c-Jun N-terminal kinase (JNK) pathway• p38 pathway
These different pathways are assembled by combiningcomponents from an extensive mitogen-activated proteinkinase (MAPK) signalling toolkit.
The mitogen-activated protein kinase (MAPK) sig-nalling properties such as their spatio-temporal controlmechanisms help to explain how they operate to regulateso many cellular processes.
The activity of the MAPK signalling pathway is reversedby the mitogen-activated protein kinase (MAPK) phos-phatases.
Mitogen-activated protein kinase (MAPK)signalling toolkitThere is an extensive mitogen-activated protein kinase(MAPK) signalling toolkit, which can be divided intodifferent functional components such as the transducers,the MAPK kinase kinases (MAPKKKs), MAPK kinases(MAPKKs), MAPKs, MAPK scaffolding proteins andMAPK target proteins (Module 2: Table MAPK signallingtoolkit). Specific components from this toolkit are thenassembled into the different signalling pathways (Mod-ule 2: Figure MAPK signalling).
Extracellular-signal-regulated kinase (ERK)pathwayThe extracellular-signal-regulated kinase (ERK) pathwayis one of the major signalling cassettes of the mitogen-activated protein kinase (MAPK) signalling pathway(Module 2: Figure MAPK signalling). It performs a num-ber of important signalling functions, including the controlof cell proliferation and the synaptic plasticity responsiblefor learning and memory. The main MAPK/ERK kinasekinase (MEKK) components are the Raf family membersRaf-1, A-Raf and B-Raf that phosphorylate two serineresidues on the MAPK/ERK kinase (MEK) components
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �72
Module 2: Figure ROS effects on Ca2+ signalling
H O2 2
O2
IP3BL
NK
DUOX
Antigen
BCR
Ca2+
SHP1SHP1
SHP1
BtkSykLy
n
P PP PP
PP
PP
P
P
P
+
_ __
2 PtdIns4,5P
PLCγ2
Reciprocal interaction between the ROS and Ca2+ signalling pathways during B cell receptor (BCR) activation.Cross-linking the B cell receptor (BCR) with antigen sets up a protein phosphorylation cascade that begins with Lyn phosphorylating both the receptorand Syk. The latter then binds to the phosphorylated receptor, where it phosphorylates the adaptor protein B cell linker (BLNK) and the tyrosine kinaseBruton’s tyrosine kinase (Btk). The latter phosphorylates BLNK and phospholipase Cγ2 (PLCγ2), which associates with BLNK and begins to hydrolysePtdIns4,5P2 to form inositol 1,4,5-trisphosphate (IP3). This phosphorylation cascade is kept in check by the tyrosine phosphatase Src homology 2(SH2) domain-containing protein tyrosine phosphatase-1 (SHP-1). One of the functions of the Ca2+ released by IP3 is to set up a positive-feedbackloop based on the activation of the Ca2+-sensitive enzyme dual oxidase (DUOX) that generates hydrogen peroxide (H2O2). The latter feeds back toinhibit SHP-1, which enables the signalling cascade to work more effectively in generating Ca2+ signals. This formation of H2O2 is highly localizedas a microdomain in the immediate vicinity of the BCR (Module 2: Figure ROS microdomains). This figure is based on information taken fromSingh et al. 2005.
MEK1/2. The latter are dual-specificity protein kinasesthat phosphorylate the tyrosine and threonine residues ofthe characteristic MAPK components ERK1/2 that are re-sponsible for stimulating the downstream effectors, manyof which are transcription factors (Module 2: Figure ERKsignalling). There is thus a linear transfer of informationthrough a phospho-relay system based on a sequentialseries of phosphorylation events.
An important feature of this ERK pathway, which can beactivated by both protein tyrosine kinase-linked receptors(PTKRs) and by G protein-coupled receptors (GPCRs), isits spatial organization (Module 2: Figure ERK signalling).
In the case of the PTKRs, growth factors such asplatelet-derived growth factor (PDGF) usually cause re-ceptor dimerization, which allows the cytosolic tyrosinekinase domains to come together and to phosphorylateeach other (Module 1: Figure PDGFR activation). Thesephosphorylated residues then function as docking motifsto pull in signalling components such as Shc, growth factorreceptor-bound protein 2 (Grb2) and Son-of-sevenless(SoS) that then activate the small GTP-binding protein Ras.Activated Ras then interacts with the protein kinase Raf,which initiates the phosphorylation cascade of the ERKpathway. The three components of this pathway (Raf-1,MEK1/2 and ERK1/2) are held in place at the cell sur-
face by the scaffolding protein kinase suppressor of Ras1 (KSR1). Up to this point, all the signal transductionprocesses have occurred at the cell surface, and the nextinformation transfer step depends upon the diffusion ofthe activated enzyme from the cell surface to the nucleus.Once it is phosphorylated, the activated phospho-ERK1/2leaves the plasma membrane to diffuse into the cytoplasmand then into the nucleus, where it phosphorylates and ac-tivates a number of transcription factors (Module 2: FigureERK signalling). Some of these genes code for MAPK sig-nalling components, such as MAPK phosphatase 1 (MKP-1), which sets up a negative-feedback loop. In addition,phospho-ERK1/2 can also act together with Ca2+ to stim-ulate cytoplasmic phospholipase A2 (cPLA2). The Ca2+
induces the cPLA2 to associate with cytosolic membranes,whereas the ERK1/2 phosphorylates Ser-505 and Ser-727to stimulate the enzymatic release of arachidonic acid fromphospholipid precursors.
There are a number of putative mechanisms for linkingthe activation of G protein-coupled receptors (GPCRs) tothe ERK pathway. The release of βγ subunits may activ-ate Src, which can then feed into the processes that ac-tivate Ras. The arrestins can also function as scaffolds toassemble components of the ERK pathway such as Raf1and ERK1/2. Alternatively, activation of phosphoinositide
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �73
Module 2: Table MAPK signalling toolkitMitogen-activated protein kinase (MAPK) signalling toolkitComponent CommentTransducers
GCK Germinal centre kinaseGLK GCK-like kinaseHPK1 Haematopoietic progenitor kinase 1MST1 Mammalian Ste20-like protein kinase
MAPKKKsASK1 Apoptosis signal-regulating kinase 1ASK2 Apoptosis signal-regulating kinase 2DLK Dual leucine-zipper-bearing kinaseMEKK1 MAPK/ERK kinase kinase 1MEKK2 MAPK/ERK kinase kinase 2MEKK3 MAPK/ERK kinase kinase 3MEKK4 MAPK/ERK kinase kinase 4MLK1 Mixed lineage kinase 1MLK2 Mixed lineage kinase 2MLK3 Mixed lineage kinase 3MosRaf-1B-RafPAK p21-activated kinaseTAK1 TGFβ-activated protein kinase 1Tpl2 Tumour progression locus 2
MAPKKsMEK1 MAPK/ERK kinase 1MEK2 MAPK/ERK kinase 2MKK3 MAPK kinase 3MKK4 MAPK kinase 4MEK5 MAPK/ERK kinase 5MKK6 MAPK kinase 6MKK7 MAPK kinase 7
MAPKsERK1 Extracellular-signal-regulated kinase 1ERK2 Extracellular-signal-regulated kinase 2ERK3-relatedERK3 Extracellular-signal-regulated kinase 3ERK5 Extracellular-signal-regulated kinase 5ERK7 Extracellular-signal-regulated kinase 7ERK8 Extracellular-signal-regulated kinase 8JNK1 c-Jun N-terminal kinase 1JNK2 c-Jun N-terminal kinase 2JNK3 c-Jun N-terminal kinase 3p38α
p38β
p38γ
p38δ
MAPK scaffolding proteinsβ-Arrestin-2JIP1 JNK-interacting protein 1JIP2 JNK-interacting protein 2JIP3 JNK-interacting protein 3KSR1 Kinase suppressor of Ras 1KSR2 Kinase suppressor of Ras 2MP1 MEK partner 1MKPX Phosphatase that may also function as
a scaffoldSKRP1 Stress-activated protein kinase (SAPK)
pathway-regulating phosphatase 1MAPK target proteins
ATF2 Activating transcription factor 2Cytoplasmic PLA2 Cytoplasmic phospholipase A2
ETSElk-1c-JunJun-BJun-DMAPKAP-kinase MAPK-activated protein kinaseMEF2 Myocyte enhancer factor 2MSK1 Mitogen- and stress-activated protein
kinase 1MSK2 Mitogen- and stress-activated protein
kinase 2SAP-1 Stomach cancer-associated protein
tyrosine phosphatase-1MAPK phosphatases
MKP-1 MAPK phosphatase-1 (Module 2:Figure ERK signalling)
Module 2 Table continuedComponent Comment
MKP-2 MAPK phosphatase-2MKP-3 MAPK phosphatase-3, acts
specifically to dephosphorylateERK2
MKP-4 MAPK phosphatase-4MKP-5 MAPK phosphatase-5VHR VH1-relatedhVH3/B23hVH5/M3/6PAC1Pyst2STEP Striatal-enriched protein tyrosine
phosphatase; dephosphorylatesERK in neurons (Module 10:Figure medium spiny neuronsignalling).
hydrolysis by Gq to produce inositol 1,4,5-trisphosphate(InsP3) and diacylglycerol (DAG) can access the ERKpathway via two mechanisms. The InsP3/Ca2+ can actthrough proline-rich tyrosine kinase 2 (Pyk2), whereasDAG and Ca2+ act through protein kinase C (PKC).
One of the major functions of the ERK pathway isto activate a range of different transcription factors suchas cyclic AMP response element-binding protein (CREB)(Module 4: Figure CREB activation) and Elk-1 (Module4: Figure ETS activation).
This ERK pathway contributes to the control of a largenumber of cellular processes:
• Regulation of cell proliferation such as T cell activation(Module 9: Figure TCR signalling)
• Cardiac hypertrophy (Module 12: Figure hypertrophysignalling mechanisms)
• Synaptic plasticity such as long-term potentiation (LTP)in hippocampal neurons
• Proliferation of endothelial cells during angiogenesis(Module 9: Figure VEGF-induced proliferation)
• Phosphorylation of the transcription factor p53(Module 4: Figure p53 domains)
• Remodelling the ERK signalling pathway may contrib-ute to the development of polycystic kidney disease(Module 12: Figure polycystins and polycystic kidneydisease)
• Activation of phospholipase A2 (PLA2) in mast cells(Module 11: Figure mast cell signalling)
c-Jun N-terminal kinase (JNK) pathwayThe c-Jun N-terminal kinase (JNK) pathway is one of themajor signalling cassettes of the mitogen-activated pro-tein kinase (MAPK) signalling pathway. It functions in thecontrol of a number of cellular processes, including prolif-eration, embryonic development and apoptosis. The path-way takes its name from the c-Jun N-terminal kinases 1–3(JNK1–JNK3), which are the MAPKs that interact withthe final effectors (Module 2: Figure MAPK signalling).They contain the dual phosphorylation motif Thr-Pro-Tyr, which is phosphorylated following activation of theupstream phosphorylation cascade.
The JNK pathway is activated by a bewildering numberof mechanisms. This complexity is evident by the fact that
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �74
Module 2: Figure ROS microdomains
PLC
PLC
IP
IP
3
3
BL
NK
BL
NK
DUOX
BCR
2 2H O microdomain
Ca
Ca
2+
2+
DUOX
Antigen
PROLIFERATION
Resting
Stimulated
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
SHP1
Btk
Btk
Syk
Syk
Lyn
Lyn
Localized ROS signalling in a microdomain surrounding the B cell receptor.Under resting conditions, the protein phosphorylation cascade that operates between Lyn, Syk, Bruton’s tyrosine kinase (Btk) and phospholipase C(PLC) (Module 2: Figure ROS effects on Ca2+ signalling) is inhibited by the tyrosine phosphatase Src homology 2 (SH2) domain-containing proteintyrosine phosphatase-1 (SHP-1), which is present at very high levels. When the receptor is stimulated by antigen, there is a Ca2+-dependent activationof dual oxidase (DUOX) that creates a microdomain of H2O2 (yellow oblong) to inhibit the SHP-1 enzymes in the immediate vicinity of the B cellreceptor (BCR). This figure is based on information taken from Singh et al. 2005.
there are 13 MAPK kinase kinases (MAPKKKs) respons-ible for feeding information into the JNK pathway. Oneway of trying to cope with this complexity is to exam-ine specific examples such as the activation of JNK by theinterleukin-1 receptor (Module 2: Figure JNK signalling).
The JNK pathway can also be activated through G pro-tein-coupled receptors (GPCRs) using G proteins such asG12/13. Just how G proteins feed into the cascade is unclear,but it seems that they activate the GTP-binding proteinssuch as Rac and Cdc42. Alternatively, the arrestins thatassociate with G protein-coupled receptors (GPCRs) dur-ing the process of receptor desensitization may functionas a scaffold to bring together components of the JNKpathway such as MKP7 and JNK3.
The MAPK signalling system operates a negative-feedback loop in that some of the genes activated by theJNK pathway code for signalling components such as thescaffold protein JNK-interacting protein 1 (JIP1), whichwill bind JNK and thus limits its action.
This JNK pathway contributes to the control of a largenumber of cellular processes:
• Phosphorylation of the transcription factor p53(Module 4: Figure p53 domains).
• The JNK pathway has been implicated in themitogen-activated protein kinase (MAPK) signalling incardiac hypertrophy (Module 12: Figure hypertrophysignalling mechanisms).
p38 pathwayThe p38 pathway is one of the major signalling cassettes ofthe mitogen-activated protein kinase (MAPK) signallingpathway. It functions in the control of apoptosis and therelease of cytokines by macrophages and neutrophils. Thepathway takes its name from the family of p38 kinases,which are the MAPKs that interact with the final effectors(Module 2: Figure MAPK signalling).
The p38 pathway can be activated either by differentreceptor mechanisms or by various environmental stressessuch as osmotic, redox or radiation stress. For example, oneof the targets of the p38 pathway that is activated by UVirradiation is one of the Cdc25 enzymes that control cellcycle progression. Phosphorylation of Ser-323 on Cdc25Bresults in the binding of 14–3-3 protein, which then pre-vents this enzyme from initiating entry into mitosis.
The Toll receptor signalling pathway provides an ex-ample of how the p38 pathway is activated by a Toll re-ceptor for lipopolysaccharide (LPS) (Module 2: Figure Tollreceptor signalling).
Mitogen-activated protein kinase (MAPK)signalling propertiesThe different mitogen-activated protein kinase (MAPK)signalling pathways have a number of important propertiesthat greatly enhance their signalling efficiency.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �75
Module 2: Figure MAPK signalling
Raf-1, A-Raf, B-Raf
MEK1/2
c-Jun, Jun-B, Jun-D, ATF2, Elk1
MEK4/7 MEK3
ATF2, Ets, MEF2,SAP1, MAPKAP2
JNK1-3ERK1/2
ERK pathway JNK pathway p38 pathway
p38 αβγδ
MAPKKK MEKK1-4, MLK1-3, TAK1, ASK1/3, TPL-2
TAK1
MAPKK
Effectors Ets, Elk1, SAP1 cPLA2
StimulusGrowth factors Osmotic stress
redox stress, radiation
Osmotic stress, radiation
FasL, cytokines
Cellularresponses
Proliferation Proliferation, apoptosis,embryonic development
Apoptosis, cytokinerelease, cell cycle arrest
MAPK
TransducersSOSGrb2Ras
Rac
Rac
Cdc42
Cdc42 Rho
Organization of the main mitogen-activated protein kinase (MAPK) signalling pathways.The generic pathway on the left summarizes the sequential organization of the mitogen-activated protein kinase (MAPK) signalling system. Externalstimuli act through a variety of transducers to stimulate the first element of the signalling pathway, which is one of the 14 different MAPK kinasekinases (MAPKKKs). This MAPKKK then phosphorylates the next element, which is one of the seven MAPK kinases (MAPKKs). This MAPKK thenphosphorylates one of the 12 MAPKs, the names of which define the different signalling pathways. The three main pathways are the ERK pathway(Module 2: Figure ERK signalling), the JNK pathway (Module 2: Figure JNK signalling) and the p38 pathway. The components that make up thesedifferent signalling pathways are summarized in the MAPK signalling toolkit (Module 2: Table MAPK signalling toolkit).
Module 2: Figure ERK signalling
Growth factors
SOSGrb2 Ras
KSR1
Raf-1
MEK1/2
ERK1/2
SHC
DAG
PKC
Pyk2
PLC
ERK1/2
Arachidonic acid ERK1/2
GPCR PTKR
Ca
MKP-1
Src
cPLA
cPLA
PL
P
P
P
P
P
P
P
P
PGqγβ
2
2
2+
Proliferation
MKP1
ETS SAP1 ELK1Negative feedback loop
+
++
+
The organization and topology of the extracellular-signal-regulated kinase (ERK) pathway.The extracellular-signal-regulated kinase (ERK) pathway, which can be made up from different components (Module 2: Figure MAPK signalling), isrepresented here by Raf-1, mitogen-activated protein kinase (MAPK)/ERK kinase 1/2 (MEK1/2) and ERK1/2, which can be activated by either proteintyrosine kinase-linked receptors (PTKRs) or by G protein-coupled receptors (GPCRs). (See the text for further details.)
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �76
Module 2: Figure JNK signalling
JIP1
MEKK1
MKK7
JNK
TAK1RAC/
Cdc42
JNK
Cytochrome c
Caspases
JNK
GPCR
PI3K-γ
Interleukin-1
?
TRAF6
P
P
P
P
P
P
Gα12/13 γβ
Proliferation, embryonic development
C-Jun ATF2 ELK1
JIP1
Negative feedback loop
APOPTOSIS
The organization and topology of the c-Jun N-terminal kinase (JNK) pathway.The c-Jun N-terminal kinase (JNK) pathway can be activated in many ways, including via different receptor mechanisms and by various environmentalstresses such as osmotic, redox and radiation stress. These different inputs are transduced by separate mechanisms that all feed into the JNKsignalling cascade. With regard to receptor activation, the JNK pathway can be activated by various cytokines such as interleukin-1 as illustratedhere. The interleukin-1 receptor (IL-1R) is composed of two receptor-binding domains that interact with interleukin-1 and a non-binding accessoryprotein. Once activated by interleukin-1, the IL-1R recruits the adaptor protein tumour-necrosis-factor-receptor-associated factor 6 (TRAF6), which thenrecruits the mitogen-activated protein kinase kinase kinase (MAPKKK) called transforming growth factor β-activated kinase 1 (TAK1) responsible forinitiating the phosphorylation cascade by phosphorylating MAPK/extracellular-signal-regulated kinase (ERK) kinase kinase 1 (MEKK1). The MEKK1then phosphorylates the dual-specificity protein kinase MAPK kinase 7 (MKK7) responsible for phosphorylating the tyrosine and threonine residues onJNK. This activation cascade occurs in the vicinity of the plasma membrane, where it is organized by the scaffolding protein JNK-interacting protein 1(JIP1). Once JNK is phosphorylated, it leaves the multimolecular activation complex and then diffuses into the nucleus, where it activates transcriptionfactors responsible for controlling processes such as proliferation, apoptosis and embryonic development.
Temporal aspects of mitogen-activated protein kinase(MAPK) signallingThe outcome of these pathways is very dependent on tem-poral aspects, particularly signal duration. For example,prolonged stimulation is necessary to induce cell prolifer-ation.
FidelityAnother important property is the fidelity of these sig-nalling pathways. The three major signalling pathwaysshare many common features: they often share signallingcomponents, they are often activated by similar inputs [es-pecially in the case of the c-Jun N-terminal kinase (JNK)and p38 pathways], and they can also regulate similar cel-lular processes. A question therefore arises as to how thefidelity of these signalling pathways is achieved in orderto reduce cross-talk and to ensure that they carry outtheir particular functions. It seems that much of this fi-delity is achieved by using molecular scaffolds to holdtogether all the components of each signalling pathway ina multimolecular complex. In this way, information canbe passed from one component to the next without inter-ference from other signalling pathways (Module 6: Figure
signalling hierarchies). Examples of such scaffolds are thescaffolding proteins kinase suppressor of Ras 1 (KSR1)(Module 2: Figure ERK signalling) and JNK-interactingprotein 1 (JIP1) (Module 2: Figure JNK signalling).
Other determinants of fidelity are the docking sitesthat enable the different mitogen-activated protein kinases(MAPKs) to bind to their specific downstream effectors.
Phenotypic remodelling of the mitogen-activatedprotein kinase (MAPK) signalling pathwayThe mitogen-activated protein kinase (MAPK) signallingpathway operates autoregulatory loops in that the dif-ferent signalling pathways can regulate the expressionof their own signalling components. For example, theextracellular-signal-regulated kinase (ERK) pathway canregulate the expression of MAPK phosphatase 1 (MKP1)(Module 2: Figure ERK signalling), whereas the c-Jun N-terminal kinase (JNK) pathway induces the expression ofJNK-interacting protein 1 (JIP1) (Module 2: Figure JNKsignalling). This induction of MKP1 and JIP1 effectivelyset up negative-feedback loops that limit the activity of theMAPK signalling pathway, and is an example of signalsomestability.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �77
Phenotypic remodelling of the MAPK signalsome mayresult in the abnormal cell proliferation observed inpolycystic kidney disease (Module 12: Figure polycystinsand polycystic kidney disease).
Nuclear factor κB (NF-κB) signallingpathwayThe transcription factor nuclear factor κB (NF-κB) isactivated by a large number of external stimuli such asthe tumour necrosis factors (TNFs), interleukin-1 (IL-1)and the pathogen-associated molecular patterns (PAMPs),which are responsible for controlling processes such asinflammation, cell proliferation and apoptosis. NF-κB be-longs to the group of transcription factors that lie latent inthe cytoplasm and then translocate into the nucleus uponactivation (mechanism 2 in Module 4: Figure transcriptionfactor activation). This diversity of downstream effectorprocesses indicates that there must be separate signallingpathways, and this is evident from the nuclear factor κB(NF-κB) signalling toolkit, which contains multiple iso-forms both of the transcription factors (the NF-κB/Relfamily) and of the activation components. This complex-ity is carried through to the nuclear factor κB (NF-κB)signalling pathway, where there are many variations onthe basic theme of NF-κB activation (i.e. Mechanism 2 inModule 4: Figure transcription factor activation).
There are two remarkable aspects of the NF-κB sig-nalling pathway. Firstly, it can control a very large numberof genes that are often activated as large cohorts in spe-cific cells by different stimuli. Secondly, it is used by anumber of different signalling systems with subtle vari-ations in the mechanism and the components that are usedto convey information into the nucleus. The tumour nec-rosis factor α (TNFα) signalling pathway and the Tollreceptor signalling pathway will be described to illustratethe main features of the NF-κB signalling pathway. Thereceptor activator of nuclear factor κB (NF-κB) ligand(RANKL), which is a transmembrane protein that belongsto the TNF family of cytokines, activates the RANKL re-ceptor (RANK) that uses the NF-κB signalling pathwayto control osteoclastogenesis (Module 8: Figure osteoclastdifferentiation).
Nuclear factor κB (NF-κB) signalling toolkitThe NF-κB signalling toolkit is composed of four mainclasses of signalling components (Module 2: Table NF-κBsignalling toolkit).
Nuclear factor κB (NF-κB)/Rel familyThe nuclear factor κB (NF-κB)/Rel family consists offive members (Module 2: Table NF-κB signalling toolkit).There is some confusion concerning the nomenclature ofthis family. The notations shown at the beginning of thetable (p50, p52, p65, RelB and c-Rel) will be used here.All members of the family share an N-terminal Rel ho-mology domain (RHD), which binds to DNA (Module2: Figure NF-κB, IκB and IKK structure). The RHD isalso used to associate NF-κB with the inhibitor of nuclearfactor κB (NF-κB) (IκB) proteins. The NF-κB/Rel family
Module 2: Table NF-κB signalling toolkitThe nuclear factor κB (NF-κB) signalling toolkitNuclear factor κB (NF-κB)transcription system
Comments
NF-κB/Rel family oftranscription factorsp50 (NF-κB1) p50 has a p105 precursor proteinp52 (NF-κB2) p52 has a p100 precursor proteinp65 (NF-κB3, also knownas RelA)RelBc-Rel
Inhibitor of NF-κB (IκB) familyIκBα Bind to p65 and c-RelIκBβ
IκBγ
IκBε
IκBζ Binds preferentially to p50Bcl3 B-cell lymphoma 3 binds to
p50/p50 and p53/p53homodimers.
Inhibitor of NF-κB (IκB)kinases (IKKs)IKKα
IKKβ
NEMO (IKKγ) A regulatory subunit responsiblefor the interaction withupstream kinases on thereceptor complex (Module 2:Figure NF-κB activation)
Associated proteinsCdc37Hsp90
of transcription factors normally function as heterodimers,and the p50/p65 complex was the first to be discovered.Some of the family members can form homodimers, suchas p50/p50 and p52/p52, and these act as repressors ofNF-κB-sensitive genes.
Inhibitor of nuclear factor κB (NF-κB) (IκB)The inhibitor of nuclear factor κB (NF-κB) (IκB) familyare characterized by an ankyrin repeat domain (Module2: Figure NF-κB, IκB and IKK structure), which func-tions in its interaction with NF-κB to form the inactivecomplex that resides in the cytoplasm. The IκB inhibitstranscription by masking the nuclear localization signal(NLS) on NF-κB, which thus prevent it from entering thenucleus. The NF-κB/IκB complex remains inactive withinthe cytoplasm until the IκB is removed following activa-tion of the nuclear factor κB (NF-κB) signalling pathway.Much of the specificity within the NF-κB signalling path-way depends on these IκB isoforms being able to bind todifferent dimers of the NF-κB/Rel family.
Inhibitor of nuclear factor κB (IκB) kinases (IKKs)The inhibitor of nuclear factor κB (IκB) kinases (IKKs)function as heterodimers and are responsible for phos-phorylating IκB to mark it for subsequent degradationby the proteasome. The kinase domain is located in theN-terminus, whereas the C-terminus has leucine zipperand helix–loop–helix domains (Module 2: Figure NF-κB,IκB and IKK structure). These two protein-associationdomains enable the enzyme to associate with a largemultisubunit complex in the cytoplasm. Another import-ant component of this complex is nuclear factor κB (NF-κB) essential modulator (NEMO) (IKKγ), which is a
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �78
Module 2: Figure NF-κB, IκB and IKK structure
HLH
HLH
ZIP
ZIP
ZIP
CCD CCD
RHD
RHD
RHD
Ser kinase
Ser kinase
RHD
RHD
NF- B/Rel familyκ
I B familyκ
p65
p52
c-Rel
RelB
Ankyrin repeat domain
Zn finger
Bcl3
I Bκ α
IKKα
I Bκ β
IKKβ
NEMO
I Bκ γ
I Bκ ε
p50
IKK family
The structure of components of the nuclear factor κB (NF-κB) signalling toolkit.The nuclear factor κB (NF-κB)/Rel family all share a Rel homology region (RHD). The mauve box in the N-terminal region of RelB is a putative leucinezipper region. The p50 and p52 isoforms also have ankyrin repeat domains that resemble those found in the inhibitor of NF-κB (IκB) family. The IκBkinase (IKK) family has both kinase (IKKα and IKKβ) and regulatory components [NF-κB essential modulator (NEMO)]. ZIP, leucine zipper; HLH,helix–loop–helix domain; CCD, coiled-coil domain. Redrawn from Handbook of Cell Signalling, Vol. 3 (R.A. Bradshaw and E.A. Dennis, eds), Westwick,J.K., Schwamborn, K. and Mercurio, F., NFκB: a key integrator of cell signalling, pp. 107–114. Copyright (2003), with permission from Elsevier; seeWestwick et al. 2003.
regulatory/structural subunit responsible for the inter-action with upstream kinases on the receptor as part ofthe tumour necrosis factor α (TNFα) signalling pathway(Module 2: Figure NF-κB activation).
The tumour suppressor CYLD binds to NEMO andinhibits the subsequent phosphorylation of IκB as part ofthe TNFα signalling pathway.
Tumour necrosis factor α (TNFα) signallingpathwayThe basic operation of the nuclear factor κB (NF-κB)/Relfamily of transcription factors is that they are activatedin the cytoplasm and then translocate into the nucleus toactivate transcription (mechanism 2 in Module 4: Figuretranscription factor activation). There are a number of vari-ations in the way that this translocation process is initiated,depending on the nature of the incoming signals and thereceptors that are being activated. In addition to the activ-ation of NF-κB, these non-enzyme-containing receptors
can activate other signalling pathways (Module 1: Figurecytokines). In this section, attention will focus on the NF-κB signalling pathway, which is used by the tumour nec-rosis factor α (TNFα). It is considered to be one of the‘classical’ mechanisms as illustrated by the following se-quence of events (Module 2: Figure NF-κB activation):
1. Occupation of the TNF receptor (TNF-R) by TNFinduces receptor oligomerization to form a complexthat attracts the adaptors TRADD and the TNFR-associated factor 2 (TRAF2), which is an adaptorthat belongs to the TNF-receptor-associated factor(TRAF) family.
2. TRAF2 is a RING domain E3 ubiquitin ligase that as-sociates with the heterotrimeric ubiquitin-conjugating(E2) complex that contains Ubc13 and Uev1A. Thisis a K63 ubiquitinating complex that adds ubiquitinchains to TRAF2 and this then helps to recruit thereceptor-interacting protein 1 (RIP1), which is also
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �79
ubiquitinated. These ubiquitin chains provide an im-portant scaffolding role in the assembly of additionalelements of the signal transduction pathway.
3. The developing receptor complex attracts additionalcomponents such as the transforming growth factor β
activated kinase-1 (TAK1), the TAK1-binding (TAB)proteins 2 and 3 (TAB2/3) and the multisubunit cyto-plasmic complex containing the inhibitor of NF-κB(IκB) kinase (IKK) α/IKKβ dimer and the regulatoryNF-κB essential modifier (NEMO) subunit, which isa central player in this translocation sequence.
4. Once this complex is complete, the TAK1 phos-phorylates and activates IKKβ.
5. The IKKβ then phosphorylates IκBα on two sites(Ser-32 and Ser-36).
6. The phosphorylated IκBα is then susceptible to ubi-quitination by the Skp1/cullin/F-box (SCF) ubiquitinligase.
7. The polyubiquitinated IκBα is then sent to the protea-some, where it is degraded, resulting in the liberationof the NF-κB heterodimer p50/p65.
8. The NF-κB is imported into the nucleus, where itbinds to the κB promoter elements to activate expres-sion of many different genes.
9. Transcription ceases when NF-κB is exported fromthe nucleus. One of the genes activated by NF-κB isIκBα, which thus sets up a negative-feedback loop,because it binds to the NF-κB that is exported fromthe nucleus to reconstitute the inactive cytoplasmiccomplex.
10. The ubiquitin signalling pathway contributes tothe recovery of this signalling cascade after TNFα
is withdrawn by removing the ubiquitin scaffoldsthat hold together the transducing complex. Thedeubiquitinating enzymes A20 and CYLD are par-ticularly active in removing the ubiquitin chains. Thereversible ubiquitination of signal transducing com-ponents is thus an essential part of the processing ofinformation by this TNF signalling pathway.
Tumour necrosis factor (TNF)-receptor-associatedfactor (TRAF) familyThe tumour necrosis factor (TNF)-receptor-associatedfactor (TRAF) family has seven members, which func-tion in the signalling pathways of various TNF receptorsuperfamily and Toll/interleukin-1 receptor mechanisms.TRAF2 functions as an adaptor for the tumour necrosisfactor receptor (Module 2: Figure NF-κB activation). Inthe case of osteoclasts, it is TRAF6 that couples thereceptor activator of nuclear factor κB (NF-κB) ligand(RANKL) receptor (RANK) to downstream signallingpathways (Module 8: Figure osteoclast differentiation).
Toll receptor signalling pathwayThe Toll receptor signalling pathway plays a centralrole in inflammatory responses (Module 11: Figure in-flammation). This pathway is activated by two types ofstimuli: cytokines, represented by interleukin-1 (IL-1),and stimuli derived from pathogens that are knownas pathogen-associated molecular patterns (PAMPs)
(Module 11: Figure formation and action of PAMPs).These PAMPs act on Toll-like receptors (TLRs) and theIL-1 receptor (IL-1R) and these form a TLR/IL-1R fam-ily that act through a similar Toll receptor signalling path-way (Module 1: Figure cytokines). The primary functionof this signalling pathway is to stimulate the transcrip-tional processes that result in the expression of a widerange of inflammatory cytokines and immunoregulators(Module 2: Figure Toll receptor signalling). As such, theToll receptor signalling pathway has to transmit informa-tion from the TLRs and the IL-1R on the cell surface to thetranscriptional factors such as nuclear factor κB (NF-κB)and activating protein 1 (AP-1) (c-Jun/Fos) that act withinthe nucleus. This information transfer system will be il-lustrated by reference to the response of the receptorTLR4 to lipopolysaccharide (LPS) (Module 11: Figureformation and action of PAMPs). As for many other sig-nalling pathways, information is transferred through bothprotein–protein interactions and protein phosphorylationreactions. The ubiquitin signalling system also has an im-portant role in orchestrating this Toll receptor signallingpathway as shown in the following sequence (Module 2:Figure Toll receptor signalling):
1. The TLR4 is a transmembrane protein that hasleucine-rich repeats in its ectodomain, while thecytoplasmic domain has a Toll/interleukin 1 (IL-1) receptor (TIR) domain. The lipopolysaccharide(LPS) that initiates this signalling pathway binds firstto an extracellular LPS-binding protein (LBP) andto CD14, which is a glycosylphosphatidylinositol-anchored membrane glycoprotein, and this complexcarries the LPS to the Toll-like receptor 4 (TLR4).
2. The TIR domain on the TLR4 receptor forms ho-mophilic interaction with the TIR domain of theadaptor proteins TIR adaptor protein (TIRAP) andMyD88. The latter is specific for certain TLRs, suchas TLR4, but not others. The other end of these ad-aptors have a death domain that draws in the IL-1 receptor-associated kinases 1 and 4 (IRAK-1 andIRAK-4), which undergoes an autophosporylation re-action that enables them to bind to another adaptorcalled TRAF6, which belongs to the tumour necrosisfactor (TNF)-receptor-associated factor (TRAF) fam-ily that has a critical role to play in the next series ofreactions.
3. The IRAK-1 and TRAF6 dissociate from the receptorand move in to the cytoplasm.
4. The TRAF 6 is a RING domain E3 ubiquitin lig-ase that associates with the heterotrimeric ubiquitin-conjugating (E2) complex that contains Ubc13 andUev1A. This is a K63 ubiquitinating complex thatresults in the autoubiquitination of TRAF6. Theubiquitinated TRAF6 then binds the transforminggrowth factor β activated kinase-1 (TAK1) and theTAK1-binding (TAB) proteins 2 and 3 (TAB2/3). Themultisubunit cytoplasmic complex containing the in-hibitor of NF-κB (IκB) kinase (IKK) α/IKKβ dimerand the regulatory NF-κB essential modifier (NEMO)subunit are also drawn into the complex. The
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �80
Module 2: Figure NF-κB activation
345
6
7
8
9
10
Import
ExportNF- Bκ
Bκ
TAB2/3TAB2/3
p50
p50
p50
p50p50p65
p65
p65
p65p65
P P
P
P P
RIP1 RIP1
TRA
F2
TRA
F2
TRA
F2
Proteasome IKKβ
IKKβ
IKKα
IKKα
NEMO
CYLD
A20
TAK1TAK1
NEMO
I BIL-6CYLDA20
κ α
I Bκ α
I Bκ α
I Bκ αI Bκ α
TNFR
TRADD
Ubiquitin UbiquitinUbc13/Uev1a
Ubc13/Uev1a
TNFα TNFαTNFα
DD
DDDD
DD
DDDD
DD
2
1
The ‘classical’nuclear factor κB (NF-κB) signalling pathway activated by the tumour necrosis factor receptor (TNFR).The p50 and p65 isoforms of the nuclear factor κB (NF-κB)/Rel family form the NF-κB dimer that is activated in the tumour necrosis factor α (TNFα)signalling pathway. The activated TNF receptor (TNFR) recruits a signalling complex to the membrane (Steps 2–4), which contains the inhibitor ofNF-κB (IκB) kinase (IKK) α/IKKβ dimer that is responsible for phosphorylating the IκBα subunit that retains p50/p65 in the cytoplasm (Step 5). Whenthe IκBα is phosphorylated, it is ubiquitinated and degraded by the proteasome (Steps 6 and 7). The p50/p65 homodimer is imported into the nucleus(Step 8), where it activates a large number of genes. One of these genes codes for IκBα, which sets up a negative-feedback loop by exportingp50/p65 from the nucleus (Step 9). Adapted from Trends Biochem. Sci., Vol. 30, Viatour, P., Merville, M.-P., Bours, V. and Chariot, A., Phosphorylationof NF-κB and IκB proteins: implications in cancer and inflammation, pp. 43–52. Copyright (2004), with permission from Elsevier; see Viatour et al.2005.
Module 2: Figure Toll receptor signalling
P
P
P
P
P
TRAF6
TRAF6
NF- Bκ
p38JNK
JUN Fos
LPSCd1
4
LBP
IRAK-1
IRAK-1
IRAK-4
MyD88
Inflammatory cytokines
TNF- IL-6; IL-1α; β
κBCRE
Immunoregulators
TGF- prostaglandinsPDE4a
β;
TLR4
TIRAP
5
P
P
P
P
CYLD
A20
Ubiquitin
Ubc13/Uev1a
2
3
4
9
10
TAB2/3
p50 p50
p50
p65 p65
p65
PIKKβIKKα
TAK1NEMO
I Bκ α
I Bκ α
I Bκ α7
8
Import
Proteasome
1
6
The Toll receptor signalling pathway.The lipopolysaccharide (LPS) that initiates this signalling pathway binds first to an extracellular LPS-binding protein (LBP) and to CD14, which is aglycosylphosphatidylinositol-anchored membrane glycoprotein, and this complex carries the LPS to the Toll-like receptor 4 (TLR4). The activated TLR4then recruits a signalling complex to relay information to both the p38 and nuclear factor κB (NF-κB) signalling pathways to induce the transcriptionof a number of inflammatory cytokines and immunoregulators as described in the text.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �81
resulting activation of TAK1 is then responsible forrelaying information out to other components of thesignalling pathways.
5. The TAK1 activates both the c-Jun N-terminal kinase(JNK) pathway and the p38 pathway (Module 2: Fig-ure MAPK signalling). The JNK and p38 functionto phosphorylate transcription factors such as AP-1, which binds to the cyclic AMP response element(CRE) site.
6. The other action of TAK1 is to phosphorylate theIKKβ component of the inhibitor of NF-κB (IκB)kinase (IKK) α/IKKβ dimer.
7. The phosphorylated IKKβ then acts to phosphorylateIκBα to liberate the NF-κB.
8. The phosphorylated IκBα is then susceptible to ubi-quitination by the Skp1/cullin/F-box (SCF) ubiquitinligase. The polyubiquitinated IκBα is then sent to theproteasome, where it is degraded.
9. The NF-κB enters the nucleus, binds to the κB siteand activates the genes that code for the inflammatorycytokines and immunoregulators that contribute toinflammatory responses (Module 11: Figure inflam-mation).
10. An important aspect of the recovery of this signallingcascade after LPS is withdrawn, is the removal ofthe ubiquitin scaffolds that hold together the trans-ducing complex. The deubiquitinating enzymes A20and CYLD are particularly active in removing the ubi-quitin chains. This ubiquitin signalling system thusplays an essential role in the processing of informa-tion by this Toll receptor signalling pathway.
This signalling pathway is particularly evident on res-ident macrophages (Module 11: Figure macrophage sig-nalling) and mast cells (Module 11: Figure mast cell sig-nalling) that respond to invading pathogens.
Virus recognition and antiviral responsesThe recognition and initiation of antiviral responses, whichis a part of the innate immune system, depend upon the ac-tivation of a number of signalling pathways such as the Tollreceptor signalling pathway. When viruses enter cells theyare broken down into fragments such as double-strandedRNA (dsRNA), single-stranded RNA (ssRNA), 5′ tri-phosphate ssRNA or CpG DNA, which are located eitherin the cytoplasm or within the endosomal compartment(Module 2: Figure viral recognition). These fragments areexamples of the pathogen-associated molecular patterns(PAMPs) that are responsible for triggering a variety of in-flammatory responses (Module 11: Figure formation andaction of PAMPs). The following sequence of reactionsdescribes how three main groups of receptors detect theseviral PAMPs to initiate a number of antiviral signallingpathways (Module 2: Figure viral recognition):
Toll-like receptors (TLRs)
1. Those viral PAMPs, which are directed into theendosome, interact with Toll-like receptors (TLR3,TLR7/8 and TLR9) that are located in the endosomalmembrane (see green box on the left of Module 2:Figure viral recognition).
2. The TLR9, which responds to CpG DNA, and theTLR7/8 that bind ssRNA both recruit the adaptorprotein MyD88. The latter then acts through TRAF6,which is one of the tumour necrosis factor (TNF)-receptor-associated factor (TRAF) family, to stimulatethe IKKαβ. The latter then activates the transcrip-tion factor NF-κB through the Toll receptor signallingpathway described in more detail earlier (Module 2:Figure Toll receptor signalling).
3. The NF-κB is imported into the nucleus where it func-tions to induce the transcription of both the inflam-matory cytokines and the genes for type I interferon(IFN).
4. When activated by dsRNA, the TLR3 receptor in-teracts with the TIR-domain-containing adaptor pro-tein inducing IFN-β (TRIF). The latter then activ-ates the two IKK-related proteins TBK1 [TRAF-family member-associated NF-κB activator (TANK)-binding protein] and IKKε, which is also known asinducible IKK (iIKK).
5. The activated TBK1/IKKε complex then phos-phorylates the interferon-regulatory factors (IFRs)IRF3 and IRF7. When phosphorylated, these twoIRFs form homodimers that are then imported intothe nucleus where they contribute to the transcriptionof the type I interferons (IFN-α and IFN-β).
Retinoic acid-inducible gene I (RIG-I)-like receptors(RLRs)
6. Those viral PAMPs that are directed into the cyto-plasm interact with cytoplasmic receptors such as theRIG-I-like receptors (RLRs) (see yellow box in themiddle of Module 2: Figure viral recognition).
7. The two main RLRs are retinoic acid-inducible gene I(RIG-I) itself and melanoma-associated gene 5(MDA5). These two receptors have a characteristicDExD/H box helicase domain that is responsiblefor binding double-stranded RNA (dsRNA). RIG-Ican also respond to 5’ triphosphate ssRNA. Thesetwo receptors also have a pair of N-terminal caspaserecruitment domains (CARD), which are the func-tional transducing components that signal to down-stream elements. The LGP2 (laboratory of geneticsand physiology-2), which also has a helicase domain,can also bind dsRNA, but it lacks the CARD domainsand thus fails to transduce cellular signals but may actas a negative regulator of MDA5 and RIG-I.
8. The CARD domains on MDA5 and RIG-I interactwith the IFN-β promoter stimulator-1 (IPS-1), whichalso contains an N-terminal CARD domain and isattached to the outer mitochondrial membrane. IPS1is also known as MAVS (mitochondrial antiviral sig-nalling), VISA (virus-inducing signalling adaptor) orCARDIF (CARD adaptor inducing IFN-β). The IPS1protein then stimulates the TBK1/IKKε complex thatthen phosphorylates the interferon-regulatory factors(IFRs) IRF3 and IRF7 as described above (see step 5).
Nucleotide oligomerization domain (NOD) protein-likereceptors (NLRs)
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �82
Module 2: Figure viral recognition
TRAF6
NF- Bκ
MyD88MyD88
Inflammatory cytokines
TNF- IL-6 IL-1
α, β
κB κB
Type I IFN genes
TLR3
Endosome
TRIF
TLRsRLRs
NLRs
IPS1
ASC
dsRNAdsRNA
Viruses
CpGDNA
ssRNA
5’Tr
ipho
spha
te s
sRN
A
dsRNA
TLR7/8
TLR9MDA5
RIG-I
NALP3
5P P P P
PP
2
3
4
9
10
p50
p50
IRF3
IRF3
IRF7
IRF7
p50 p50
p65
p65
p65 p65
IKKβIKKε
IKKα
TBK1I Bκ α
I Bκ α
7
8
Import
1 6
Pro-IL-1βIL-1β
Mitochondrion
IFN /α β
P
PP
Caspase-1
LGP2
Viral recognition and antiviral responses.When viruses enter cells they are degraded into short fragments of double-stranded RNA (dsRNA), single-stranded RNA (ssRNA), 5’ triphosphatessRNA or CpG DNA. There are three main groups of sensors that detect these fragments: Toll-like receptors (TLRs, see green box), retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs, see yellow box) and nucleotide oligomerization domain (NOD)-like receptors (NLRs, see pink box).These different receptors then activate different signalling systems to induce transcription of the inflammatory cytokines and interferon as describedin the text. The information used to construct this figure was taken from McCartney and Colonna (2009) and Takeuchi and Akira (2009).
9. The double-stranded RNA (dsRNA) in the cytoplasmcan also interact with the nucleotide oligomerizationdomain (NOD) protein-like receptors (NLRs) (seepink box on the right of Module 2: Figure viral re-cognition). One of the NLRs is the Nacht domain-,Leucine-rich repeat-, and PYD-containing protein 3(NALP3), which is also known as cryopyrin that re-sponds to dsRNA to activate caspase-1. The interac-tion between NALP3 and caspase-1 is facilitated by anadaptor called apoptosis-associated speck-like proteincontaining a CARD (ASC). The interaction betweenthese three proteins takes place in a macromolecularcomplex known as the inflammasome.
10. The activated caspase-1 contributes to the develop-ment of an inflammatory response by cleaving pro-IL-1β to form the inflammatory cytokine interleukin1β (IL-1β).
Phospholipase D (PLD) signallingpathwayThe phospholipase D (PLD) signalling pathway functionsby generating phosphatidic acid (PA), which acts to reg-ulate a wide range of cellular processes. Phospholipase D(PLD) activation depends upon a number of mechanisms,and these may vary depending on where the signallingmechanism is located within the cell. The primary mes-senger produced by this signalling pathway is PA, andphosphatidic acid (PA) action is carried out through anumber of downstream effectors. Phosphatidic acid (PA)
metabolism occurs through different pathways that gen-erate further signalling molecules such as diacylglycerol(DAG) and lysophosphatidic acid (LPA).
Phospholipase D (PLD) activationMammals have two phospholipase D (PLD) genes (PLD1and PLD2), both of which have splice variants. Most atten-tion has focused on PLD1 as a signal transducer because ithas a low basal activity that increases markedly in responseto external stimuli. On the other hand, PLD2 has a highbasal activity and its role in signalling is uncertain. Whilemost of the PLD2 is located on the plasma membrane,PLD1 is found predominantly on intracellular membranes(e.g. Golgi, endoplasmic reticulum and endosomes), buthas also been located at the plasma membrane, particu-larly at caveolae. The structure of the enzyme containsmotifs responsible for its membrane location and catalyticactivity (Module 2: Figure PLD isoforms). The activityof PLD1 increases following stimulation of both proteintyrosine kinase-linked receptors (PTKRs) and G protein-coupled receptors (GPCRs) (Module 2: Figure PLD sig-nalling). One of the problems with trying to understandthe signalling function of this pathway is to determinejust how these different receptors act to stimulate PLD1.The fact that the activation of PLD1 seems to depend onthe prior activation of other signalling pathways such asthe diacylglycerol (DAG)/protein kinase C (PKC) sig-nalling cassette or the PtdIns 3-kinase signalling cassette(Module 2: Figure PLD signalling) suggests that the PLDsignalling pathway is not an autonomous signalling system,
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �83
Module 2: Figure PLD isoforms
1072
1034
PHPX
PX PH
I
I
III
IIIIII
IV
IVIV
II
IIII
Palmitate (Cys 240)
Palmitate (Cys 240)
Palmitate (Cys 241)
Palmitate (Cys 241)
PLD1
PLD2
HKD
HKD
HKD
HKD
The domain structure of phospholipase D (PLD).The two isoforms of phospholipase D (PLD) have similar domain structures. Palmitoylation of Cys-240 and Cys-241 within the pleckstrin homology(PH) domain play a critical role in localizing the enzyme to membranes. There are four conserved sequences (I–IV). Domains II and IV containHXKXXXXD, the so-called HKD motif, which come together to form the catalytic domain. The C-terminal region has a phox homology (PX) domainand a PH domain responsible for the binding of PtdIns4,5P2 during activation of the enzyme (Module 2: Figure PLD signalling).
Module 2: Figure PLD signalling
PIP
PIP2
PIP2
PIP2
PIP3
DAG
PC
PA
PKCα
RhoArf
PLD1
PI3KPLCγ PLCβ
LPA
LPA
Sphingosine kinase
Protein phosphatase 1
mTOR
Membrane budding and fusion
Membrane traffickingCytoskeletal organization
InflammationPhagocytosisProliferation
Secretion (Mast cells)Respiratory burst
PAhydrolase
PLA2
RalA
Ras
Ral-GDS
+
+
ChemokinesChemotactic factors
Growth factors Antigen
Plasma membrane OREndosomal membranes
PtdIns4P-5K
The phospholipase D (PLD) signalling pathway.This signalling pathway depends upon the enzyme phospholipase D1 (PLD1), which associates primarily with intracellular membranes (e.g. Golgi andendosomes), but is also found in the plasma membrane. The activation of PLD1 is linked to many receptors through a number of mechanisms that mayvary depending on the location and which of its many functions PLD is carrying out. One activator is protein kinase Cα (PKCα), through a mechanismthat is independent of its catalytic activity. Other major regulators are various small G proteins. Arf and Rho may be activated by the lipid messengerPtdIns3,4,5-trisphosphate (PIP3) formed by the PtdIns 3-kinase signalling cassette, whereas Ral A may be activated via Ras and the GTP/GDPexchange factor Ral-GDS. In addition, PLD1 has an absolute requirement for PtdIns4,5P2 (PIP2), which may be part of a positive-feedback loopbecause the PtdIns4P 5-kinase (PtdIns4P-5K) is activated by phosphatidic acid (PA). The action of PA is terminated either by a PA phosphohydrolase,which removes phosphate to leave behind diacylglycerol (DAG), or by a phospholipase A2 (PLA2) to produce lysophosphatidic acid (LPA).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �84
but should be considered more as a downstream effectorof these other signalling pathways.
Phosphatidic acid (PA) actionThe primary messenger of the phospholipase D (PLD) sig-nalling pathway is the lipid phosphatidic acid (PA), whichhas a number of actions within the cell (Module 2: FigurePLD signalling). The signalling function of PA is mainlydirected towards the regulation of various enzymes such asstimulation of the target of rapamycin (TOR) and sphin-gosine kinase or inhibition of protein phosphatase 1. Italso plays an important role in regulating phagocytosis(Module 4: Figure phagosome maturation).
In addition to such signalling functions, a local accumu-lation of PA may also alter the physical properties of themembrane by creating curvatures to facilitate the forma-tion of vesicles for intracellular trafficking.
Phosphatidic acid (PA) metabolismTwo separate enzymes carry out the metabolism of phos-phatidic acid (PA). It can be dephosphorylated to diacyl-glycerol (DAG) by a PA phosphohydrolase or it can bepartially deacylated by a phospholipase A2 (PLA2) to formlysophosphatidic acid (LPA) (Module 2: Figure PLD sig-nalling). The LPA released from the cell is a potent agoniston receptors of the endothelial differentiation gene (EDG)family (Module 2: Figure sphingomyelin signalling).
Sphingomyelin signalling pathwayThe sphingomyelin signalling pathway that has been im-plicated in the control of a whole host of cellular pro-cesses through the generation and function of ceramideand sphingosine 1-phosphate (S1P), which are the mainmessengers operating in this signalling pathway. The ac-tion of sphingosine 1-phosphate (S1P) is complicated be-cause, in addition to acting internally, S1P is released fromcells to function as an external ligand acting on cell-surfacereceptors. This sphingomyelinase signalling pathway alsoproduces ceramide, another messenger that plays a signi-ficant part in processes such as cell proliferation, apoptosisand the response of the cell to stress and injury. One of thedifficulties in understanding this pathway is its pleiotropiceffect on cells with responses that are often diametricallyopposed, such as proliferation and apoptosis. A clue tothis complexity lies in the fact that the pathway can spawna number of messengers, and the action of these is verydependent on the current state of the cell, especially withregard to what other signalling pathways are active.
Generation and function of ceramide andsphingosine 1-phosphate (S1P)The generation of ceramide and sphingosine 1-phosphate(S1P), which are the two main bioactive ‘messenger’ lip-ids of the shingomyelin signalling pathway, begins withthe hydrolysis of sphingomyelin (SM) by various sphin-gomyelinases (SMases) to form ceramide (Module 2: Fig-ure sphingolipid metabolism). A ceramidase (CDase) thencleaves off one of the fatty acid chains to form sphingosine(Sph), which is then phosphorylated by sphingosine kinase
(SPHK). As illustrated by the figures in Module 2: Fig-ure sphingolipid metabolism, the relative cellular levels ofthese sphingolipids vary enormously. The precursor SM ispresent at the highest level and this then declines duringthe different metabolic transformations.
An important aspect of this sphingomyelin signallingpathway is the cellular location and function of these dif-ferent metabolic steps (Module 2: Figure sphingomyelinsignalling):
1. Sphingomyelin synthesis is carried out first in the en-doplasmic reticulum (ER), where ceramide is formed,and is completed by the Golgi. The first step is carriedout by serine palmitoyl transferase (SPT) that com-bines serine and palmitate to form dihydrosphingosine(dhSph), which is then converted into dihydroceram-ide (dhCer) by dihydroceramide synthase (CerS). Fi-nally, a desaturase converts dhCer into ceramide (Cer).
2. The ceramide formed at the ER is then transportedto the Golgi by a ceramide transfer protein (CERT).A sphingomyelin synthase (SMS) located in the Golgiconverts ceramide into sphingomyelin (SM), which istransferred to the membrane through a vesicle trans-port mechanism.
3. The hydrolysis of SM in the plasma membrane is activ-ated by receptors sensitive to stimuli such as tumournecrosis factor α (TNFα), interleukin 1, CD28 andFas. The coupling between receptors and the neutralsphingomyelinases (SMases) has been worked out insome detail for the TNFα receptor. A factor associatedwith neutral SMase (FAN) functions to couple the en-zyme to the neutral SMase activation domain (NSD) ofthe TNFα receptor. These components of the sphin-gomyelinase signalling pathway are often highly con-centrated in lipid rafts and caveolae, which representthe site where some of the sphingomyelinases functionto hydrolyse sphingomyelin.
4. The activation of neutral SMase at the plasma mem-brane provides one of the major sources of ceramide(Cer), which can then be metabolized to other sig-nalling sphingolipids (Module 2: Figure sphingolipidmetabolism). A ceramidase (CDase) hydrolyses Cer tosphingosine (Sph), which can then be phosphorylatedby sphingosine kinase (SPHK) to form sphingosine 1-phosphate (S1P). Since the latter is soluble, it leaves themembrane and is free to diffuse both into and out ofthe cell (see later). The SPHK is activated by a numberof signalling pathways using messengers such as Ca2+,DAG, cyclic AMP and ERK1/2 that are produced bysignalling pathways activated by growth factors andhormones. Ceramide can also be phosphorylated bya ceramide kinase to form ceramide 1-phosphate, forwhich there is no apparent function.
5. A variety of stress stimuli, such as ionizing radiation,UV irradiation and reactive oxygen species (ROS) actby stimulating the acidic sphingomyelinases (SMases)localized in lysosomes to hydrolyse sphingomyelin(SM) to form ceramide (Cer) (Module 2: Figure sphin-gomyelin signalling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �85
Module 2: Figure sphingolipid metabolism
Sphingomyelin (SM)
Ceramide (Cer)
Sphingosine (Sph)
C C
C C
C
C CC C C C
C C
C
C C
CH
CH
CH3
3
3
O OOH
Sphingosine-1-PO (S1P)
4
HO HO
OH
OH OH
OH
H H
H H
O
O O
OO
O
OP P
N N N N
N
CDase SPHK SMase
30,000 3,000 100 1
Sphingolipid metabolism.The sphingomyelin signalling pathway depends on the conversion of sphingomyelin (SM) into a series of bioactive lipids capable of activatingvarious signalling mechanisms as described in Module 2: Figure sphingomyelin signalling. The numbers represent the relative cellular levels of thesesphingolipids. SMase, sphingomyelinase; CDase, ceramidase; SPHK, sphingosine kinase. Information for this Figure was taken from Hannun andObeid (2008).
Module 2: Figure sphingomyelin signalling
EDGreceptors
Growth factors Hormones
ABCC1
CAPPsCAPKJNK
Cathepsin D
PKCζ
TNFα
6
7
8
9
1
SM
SM
Cer
Lysosome
ER
Cer
dhSph
dhCer
Cer
ROS
Sph
AcidicSMase
Neutral SMaseN
NP
N
NP
OHN
OH
N
NO
HN
N
OHN
S1PNP
S1P LPANP
CDase
Ca 2+
DAG
Gq
Phospho-ethanolamine
SPHK
S1P lyase SPPs
FAN
Golgi
SPT
CerS
Serine + palmitate
DesaturaseIonizingradiation
InsP3
Ras
PKB
ERK1/2
PKC
CaMKII
PI 3-Kβ/γ
Apoptosis
Cell cyclearrest
Senescence
Survival
Proliferation
Anti-inflammatory responses
ERK1/2DAGCa2+
2
34
5
The sphingomyelin signalling pathway.A number of stimuli can activate the neutral or acidic sphingomyelinases (SMase) to hydrolyse sphingomyelin (SM) to ceramide (Cer), which is thenconverted into sphingosine (Sph) by ceramidase (CDase). The sphingosine is converted into sphingosine 1-phosphate (S1P) by a sphingosine kinase(SPHK), which is sensitive to other signalling pathways using messengers such as Ca2+, diacylglycerol (DAG), cyclic AMP (cAMP) and extracellular-signal-regulated kinase 1/2 (ERK1/2). Ceramide can activate a number of targets, and some of these can activate apoptosis. On the other hand, S1Pcan promote survival and proliferation by passing out of the cell, where it functions as an external ligand to activate endothelial differentiation gene(EDG) receptors.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �86
6. S1P is metabolized via two separate pathways: it canbe converted back into sphingosine by a sphingosine1-phosphate (S1P) phosphatase (SPP) or it can becleaved by an S1P lyase to give palmitaldehyde andphosphoethanolamine. The lyase has a 20-amino-acidtransmembrane domain that positions the enzymes incellular membranes.
The sphingomyelinase signalling pathway is thus sens-itive to external signals at a number of discrete steps. Theproduction of ceramide is accelerated by receptors sens-itive to cytokines such as TNFα or to various stress sig-nals such as ionizing radiation and reactive oxygen species(ROS). The formation of S1P can also be enhanced bygrowth factor and hormone receptors that elevate secondmessengers such as Ca2+, DAG, ERK1/2 or cyclic AMPthat act by stimulating SPHK. In the absence of these sig-nals, the main messenger will be ceramide, but this willswitch over to S1P if the SPHK is activated. This inter-relationship between ceramide and S1P has led to a cer-amide/S1P rheostat model, where a balance between thesetwo messengers is thought to determine cell fate. In gen-eral, ceramide seems to tip the balance in favour of cellcycle arrest, senescence and apoptosis, whereas S1P pro-motes survival and proliferation (Module 2: Figure sphin-gomyelin signalling). The different outcomes controlledby these two messengers may depend on their ability toactivate separate signalling cascades.
7. The ceramide formed at the plasma membraneor in the lysosomes acts on a number of tar-gets such as ceramide-activated protein kinase(CAPK), protein kinase Cζ (PKCζ), cathepsin Dand ceramide-activated protein phosphatases (CAPP),such as PP1 and PP2A. Most of these ceramide-sensitive targets act to promote cell cycle arrest andapoptosis either directly or indirectly. For example,the cathepsin D converts Bid into tBid to promote ap-optosis (see Step 3 in Module 11: Figure apoptosis).On the other hand, the CAPPs such as PP2A can de-phosphorylate various components that are used bythe EDG receptors to drive cell survival and prolifer-ation. For example the dephosphorylation of proteinkinase B (PKB) not only prevents it from inhibitingapoptosis but it will also reduce its ability to enhanceproliferation.
8. One of the actions of the soluble sphingosine 1-phos-phate (S1P) is to diffuse out of the cell passing throughthe ABCC1 transporter, which is a family mem-ber of the ATP-binding cassette (ABC) transporters(Module 3: Table ABC transporters). The extracellu-lar S1P then activates EDG receptors, which belong tothe family of G protein-coupled receptors (GPCRs)(Module 1: Table G protein-coupled receptors) thatare capable of relaying information down a number ofsignalling pathways.
9. These signalling pathways control cellular pro-cesses, such as cell survival, proliferation and anti-inflammatory responses, which are opposite to thosecontrolled by ceramide.
This sphingomyelin signalling pathway has been implic-ated in a number of cellular processes:
• S1P has been proposed to release the Ca2+ necessary toactivate the formation of PtdIns3P on endosomes duringphagosome maturation (Module 4: Figure phagosomematuration).
• S1P appears to contribute to the CD28 co-stimulatorypathway during T cell activation (Module 9: Figure Tcell signalling map).
• S1P contributes to the communication between en-dothelial cells and pericytes during angiogenesis (seeStep 6 in Module 9: Figure angiogenesis signalling).
Sphingosine 1-phosphate (S1P)The key enzyme in the formation of sphingosine 1-phosphate (S1P) is sphingosine kinase (SPHK). The mes-senger function of S1P is complicated by the fact that itmay have both intra- and extra-cellular actions.
S1P can exert its extracellular action either in anautocrine mode (activate the same cell from which it isreleased) (see Step 8 in Module 2: Figure sphingomyelinsignalling) or a paracrine mode (diffuse away to activateneighbouring cells). This extracellular action is mediatedby endothelial differentiation gene (EDG) receptors, so-called because they were first described in human um-bilical vein endothelial cells induced to differentiate byphorbol esters. There is a family of these EDG receptorsthat are all G protein-coupled receptors (GPCRs) capableof activating most of the conventional signalling pathways(Module 1: Table G protein-coupled receptors). The EDG1, 3, 5, 6 and 8 receptors mediate the action of S1P, whereasthe EDG 2, 4 and 7 receptors appear to respond to lyso-phosphatidic acid (LPA), a related lysolipid. The EDG re-ceptors that function to induce a mobilization of internalCa2+ act by stimulating the formation of inositol 1,4,5-trisphosphate (InsP3). One function of the EDG receptorsis to regulate cell motility and directional migration. Anexample of the latter is preosteoclast chemotaxis where S1Pdirects preosteoclasts form the bone marrow back into theblood (Module 8: Figure preosteoclast homeostasis).
The intracellular action of S1P is not as well defined,mainly because it is often difficult to separate an intra-cellular action from that induced by its extracellular ac-tion. An intracellular action of S1P has been proposed forphagosome maturation, where it releases the Ca2+ neces-sary to activate the formation of PtdIns3P on endosomes(Module 4: Figure phagosome maturation). One of theproblems with trying to establish such an intracellular ac-tion for S1P is that it could also mobilize internal Ca2+
by acting through the EDG receptors to generate InsP3.However, there are indications that S1P can act directly torelease Ca2+ from an internal store. Unlike InsP3, how-ever, release occurs without the appearance of elementaryevents. One of the difficulties with this hypothesis is thatthe channel on the endoplasmic reticulum (ER) that isopened by S1P remains to be identified. An earlier pro-posal that S1P might act through Scamper has not beensubstantiated. Another suggestion is that S1P functions asthe Ca2+ influx factor (CIF) that is proposed to link store
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �87
depletion to Ca2+ entry. Another proposed action of S1P isto activate the extracellular-signal-regulated kinase (ERK)cascade to promote proliferation. Overexpression of theSPHK can induce tumour formation. Therefore the pre-cise role of S1P and its relationship to ER signalling requirefurther clarification.
CeramideThis is an enigmatic messenger in that it has been linkedto both proliferation and apoptosis, depending very muchon the background activity of other messenger systems.In keeping with its different functions, ceramide can ac-tivate a number of signalling components such as theceramide-activated protein kinase (CAPK), protein kinaseCζ (PKCζ) and ceramide-activated protein phosphatases(CAPP), such as PP1 and PP2A. Some of these ceramidetargets enable it to interact with other signalling systemssuch as the c-Jun N-terminal kinase (JNK) and mitochon-drial systems. With regard to the latter, ceramide may ac-tivate mitochondrial reactive oxygen species (ROS) form-ation through ceramide-activated protein kinase (CAPK).It is important to stress that apoptosis can occur withoutthe need for ceramide. However, there is overwhelm-ing evidence that the formation of ceramide can stronglytip the balance in favour of apoptosis. It contributes tothis activation of the cell death programme by activat-ing the intrinsic pathway at the level of the mitochon-dria (Module 11: Figure apoptosis). One way of inter-acting with the mitochondria depends on its activationof the JNK pathway, which is known to induce apop-tosis (Module 2: Figure JNK signalling). Ceramide canalso activate cathepsin D, which converts Bid into tBid,which is one of the pro-apoptotic members of the Bcl-2superfamily.
Sphingomyelinases (SMases)Sphingomyelinases (SMases) exist in different isoformsthat can be distinguished by their pH optima: acidic, neut-ral or alkaline. The last is found in bile and plays a role in di-gestion. Sphingomyelin signalling seems to depend on theacidic and neutral isoforms (Module 2: Figure sphingomy-elin signalling). The neutral SMase functions to generateceramide in response to the activation of receptors sensit-ive to stimuli such as tumour necrosis factor α (TNFα) andinterleukin 1 (see Step 3 in Module 2: Figure sphingomy-elin signalling). The acidic form was originally identifiedin lysosomes where it generates ceramide in response tostress stimuli (see Step 5). There also are indications thatthe acidic SMase may act on sphingomyelin located in theouter leaflet. The acidic SMase is a component of lipid raftsand caveolae, which are microdomains of the plasma mem-brane containing high levels of the precursor sphingomy-elin. It is this acidic isoform that is defective in patientswith Niemann-Pick disease.
Ceramide-activated protein kinase (CAPK)Ceramide-activated protein kinase (CAPK) is amembrane-bound proline-directed protein kinasethat acts by phosphorylating Raf-1, thereby enablingceramide to plug into the mitogen-activated protein
kinase (MAPK) cascade and could account for those caseswhere ceramide promotes both inflammation and proli-feration.
Sphingosine kinase (SPHK)Sphingosine kinase (SPHK) appears as two isoforms(SPHK1 and SPHK2), which have different tissue distri-butions. SPHK1 is found at high levels in the lung andspleen, whereas SPHK2 is expressed mainly in liver andheart. Both have five highly conserved domains, with theATP-binding site and catalytic site located in the C2 do-main.
The enzyme is both membrane-bound and free in thecytosol, and there is some uncertainty as to how it isactivated by cell-surface receptors. It seems likely thatit responds to various downstream signals emanatingfrom these receptors such as Ca2+/calmodulin, diacylgly-cerol (DAG)/protein kinase C (PKC), cyclic AMP/proteinkinase A (PKA) or extracellular-signal-regulated kinase1/2 (ERK1/2) (see Step 4 in Module 2: Figure sphin-gomyelin signalling). Another activator is oxidized low-density lipoprotein (LDL), which may play a role incell proliferation. Consistent with the activation by vari-ous messenger pathways is the fact that SPHK con-tains consensus sequences for Ca2+/calmodulin bindingand phosphorylation sites for PKA, casein kinase II andPKC.
Sphingosine 1-phosphate (S1P) phosphatase (SPP)Mammals have two sphingosine 1-phosphate (S1P) phos-phatases (SPPs). Both SPP1 and SPP2 have eight to tentransmembrane domains that locate the enzymes withinthe endoplasmic reticulum (ER). SPP-1 is located mainlyin the placenta and kidneys, whereas SPP2 is found in thebrain, heart, colon, kidney, small intestine and lung. Theseenzymes convert S1P back into ceramide and can diminishsurvival and promote apoptosis (see Step 6 in Module 2:Figure sphingomyelin signalling).
Ceramide-activated protein phosphatase (CAPP)Ceramide-activated protein phosphatase (CAPP) is amember of the protein phosphatase 2A (PP2A) family ofserine/threonine phosphatases that consist of three sub-units: A and B are regulatory, whereas C is the catalyticsubunit. In the case of CAPP, ceramide acts to stimulatephosphatase activity by binding to one of the regulatoryB subunits. The ability of ceramide to inhibit growth isprobably mediated through PP2A.
ScamperA sphingolipid Ca2+-release-mediating protein of the en-doplasmic reticulum (ER) (Scamper) was originally pro-posed to be the Ca2+ channel on the ER that respondsto signals from the sphingomyelin signalling pathway,but more recent studies have shown that it has a singlemembrane-spanning segment that seems to remodel theactin cytoskeleton.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �88
Module 2: Figure JAK and STAT structure
200 400 600 800 1000 12000
JH1JH2
Pseudokinase domain
Kinase domain
JH5JH6JH7 JH4 JH3
JAKs
STATs
2 & 6
1,3,45A & 5B
Y
Y
SH2SH2
SH2SH2
SH3SH3
SH3SH3
DNA binding domain
STATdimerization domain
TADTAD
S
Receptorbinding region
Domain structure of the JAKs and STATs.There are four mammalian Janus kinases (JAKs): JAK1, JAK2, JAK3 and Tyk2. They all have a similar domain structure, which has seven JAKhomology (JH) domains. JH1 is the kinase domain, whereas JH2 is a pseudokinase domain. Some of the other JH regions seem to contribute tothe binding of JAKs to various cell-surface receptors, where they function to activate the signal transducers and activators of transcription (STATs).There are seven mammalian STAT genes. STATs 2 and 6 have 850 amino acids, whereas the others are somewhat shorter. The tyrosine residuenear residue 700 is phosphorylated during the activation domain and functions as a binding site for the Src homology 2 (SH2) sites on other STATsduring the dimerization process (Module 2: Figure JAK/STAT function). The DNA-binding domain is located between the SH3 (Src homology 3) andSH2 domains. The C-terminus has a transcriptional activation domain (TAD), which, on the shorter-length STATs, contains a serine residue that canmodulate transcriptional activity when phosphorylated.
Janus kinase (JAK)/signal transducerand activator of transcription (STAT)signalling pathwayThe Janus kinase (JAK)/signal transducer and activatorof transcription (STAT) signalling pathway provides amechanism for rapidly activating gene transcription inresponse to a large number (>35) of external ligands.This signalling pathway is mainly activated by cytokinessuch as interferon, but is also used by receptor tyr-osine kinases [epidermal growth factor receptor (EGFR),platelet-derived growth factor receptor (PDGFR)], non-receptor tyrosine kinases and G protein-coupled receptors(GPCRs). The Janus kinase (JAK)/signal transducer andactivator of transcription (STAT) structure reveals the ma-jor features of these two components and how they arelinked during the signal transducer and activator of tran-scription (STAT) activation cascade. Cell-surface recept-ors act by phosphorylating the STATs, which are latenttranscription factors. Once phosphorylated, these STATsleave the membrane and then dimerize before migratinginto the nucleus where they bind to specific DNA-bindingelements to activate transcription. There is considerableevidence for a Janus kinase (JAK)/signal transducer andactivator of transcription (STAT) function in growth anddevelopment.
Janus kinase (JAK)/signal transducer andactivator of transcription (STAT) structureThe two major components of the signalling pathwayare the Janus kinases (JAKs) and their substrates the sig-nal transducers and activators of transcription (STATs)(Module 2: Figure JAK and STAT structure). Of the fourmammalian JAKs, three are expressed fairly ubiquitously,whereas JAK3 is restricted to natural killer (NK) cellsand thymocytes, with some expression in vascular smoothmuscle cells and endothelium. The main structural com-ponent of the JAKs is the kinase domain that functions tophosphorylate the STATs at a key tyrosine in the region ofresidue 700 during the signal transducer and activator oftranscription (STAT) activation cascade. The STATs havea number of functional domains whose three-dimensionalstructure reveals how the STAT dimers are formed andhow they bind to DNA (Module 2: Figure STAT1/DNAcomplex).
Signal transducer and activator of transcription(STAT) activation cascadeThe signal transducers and activators of transcription(STATs) are a family of latent cytoplasmic receptors thatare activated by a phosphorylation cascade that can beinduced by many different receptors (e.g. cytokine re-ceptors, tyrosine kinase-linked receptors, non-receptor
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �89
Module 2: Figure STAT1/DNA complex
Organization of a STAT1/DNA complex.The ribbon diagram shown in (A) illustrates how the two DNA-binding domains of the two subunits attach the dimer to the DNA helix. The linker domain(orange) attaches to the SH2 (Src homology 2) domains (light blue) that hold the molecule together through the intermolecular SH2–phosphotyrosineinteraction. The molecular surface representation shown in (B) has the same orientation as in (A). The local electrostatic potential over the cell surfaceis represented by the colouring, with blue representing positive and red negative potentials. (Reproduced from Cell, Vol. 93, Chen, X., Vinkemeier,U., Zhao, Y., Jeruzalmi, D., Darnell, Jr, J.E. and Kuriyan, J., Crystal structure of a tyrosine phosphorylated STAT-1 dimer bound to DNA, pp. 827–839.Copyright (1998), with permission from Elsevier; see Chen et al. 1998.
tyrosine kinases and G protein-linked receptors). The ver-satility of this signalling mechanism is greatly enhanced bythe way the Janus kinases (JAKs) and STATs can be mixedand matched to generate an enormous number of com-binations as illustrated by the different cytokine receptors(Module 2: Figure JAK/STAT heterogeneity).
A typical activation cascade for cytokine receptors,which illustrates the essential role of the JAKs in a se-quence of tyrosine phosphorylations that culminates in thephosphorylation and activation of the STATs, is outlinedin the following sequence (Module 2: Figure JAK/STATfunction):
1. Agonists induce dimerization by binding to the extra-cellular domains of the receptor subunits.
2. The JAKs, which are associated with the cytoplasmicdomain of these receptors, phosphorylate each other.
3. The activated JAKs then phosphorylate tyrosineresidues on the receptors to provide docking sites forthe Src homology 2 (SH2) domains on the STATs.
4. Once the STATs have docked, they are then phos-phorylated by the JAKs on the tyrosine residue loc-ated in the C-terminal region (Module 2: Figure JAKand STAT structure).
5. The phosphorylated STATs, which are dimerizedthrough an intermolecular SH2–phosphotyrosine in-teraction, leave the receptor.
6. These dimers are imported into the nucleus usingimportin-α (Imp-α) and attach to promoter regionsthrough their DNA-binding domains (Module 2: Fig-ure STAT1/DNA complex).
7. STATs activate the transcription of a number of tar-get genes. One group of these genes code for thesuppressor of cytokine signalling proteins (SOCS),which thus function as part of a negative-feedbackloop to limit the action of the signalling pathway(Module 2: Figure JAK/STAT function).
8. STAT activity is terminated by a nuclear protein tyr-osine phosphatase (N-PTP) that removes the tyrosylphosphate groups. The inactive STAT is then exported
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �90
Module 2: Figure JAK/STAT heterogeneity
TYPE I CYTOKINES
TYPE II CYTOKINES
IL-2, IL-7,IL-9, IL-15
IL-4
IL-13
IL-3, IL-5, G-CSF, GM-CSF
IL-6, IL-11, OSM CNTF, LIF, CT-1
IL-12
Growth hormone
Prolactin
Erythropoietin
Thrombopoietin
IFNα, IFNβ
IFNγ
IL-10
JAK1JAK3
STAT5a, STAT5b STAT3
STAT5a, STAT5b STAT3
STAT5a, STAT5b
STAT5a, STAT5b
STAT5a, STAT5b
STAT5a, STAT5b
JAK1JAK3 STAT6
STAT3
STAT4
STAT6JAK1,JAK2 Tyk2
JAK1,JAK2 Tyk2
JAK2
JAK2
JAK2
JAK2
JAK2
JAK2, Tyk2
JAK1, Tyk2
JAK1, Tyk2
JAK1, Tyk2
STAT1, STAT2
STAT1
STAT3
Heterogeneity of the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) combinations used by different cytokine receptors.The cytokines acting on cell-surface receptors (blue and mauve boxes) activate different combinations of Janus kinases (JAKs) and signal transducersand activators of transcription (STATs). CNTF, ciliary neurotrophic factor; GCSF, granulocyte colony-stimulating factor; IFN, interferon; IL, interleukin;LIF, leukaemia inhibitory factor; OSM, oncostatin M. The organization of some of the type I cytokine receptors is shown in Module 1: Figure type Icytokine receptors.
from the nucleus by chromosome region maintenance1 (CRM1), thus completing the cycle.
For some of the other receptors, these tyrosine phos-phorylations are carried out by kinases other than theJAKs. After the STATs are phosphorylated, they leavethe receptor and then dimerize before translocating intothe nucleus. This activation cascade can occur quickly,with the activated STATs appearing in the nucleus withinminutes. Transcriptional activity can be modulated byphosphorylation of the serine residue in the transcrip-tional activation domain (TAD) of STAT 1, 3, 4, 5A and 5B(Module 2: Figure JAK and STAT structure). In the case ofSTAT 1 and STAT4, phosphorylation enhances transcrip-tional activity, whereas the binding of STAT5a to DNA isgreatly prolonged.
Suppressor of cytokine signalling proteins(SOCS)The suppressor of cytokine signalling proteins (SOCSs)are induced during the activation of the Janus kinase
(JAK)/signal transducer and activator of transcription(STAT) signalling pathway (Module 2: Figure JAK/STATfunction). The SOCS act by inhibiting the Janus kinases(JAKs) and thus operate a negative-feedback loop to limitthe action of cytokines.
There is a marked increase in expression of the SOCS-3isoform in hypothalamic neurons following the action ofleptin during the control of food intake and body weight(Module 7: control of food intake).
Janus kinase (JAK)/signal transducer andactivator of transcription (STAT) function ingrowth and developmentThe Janus kinase (JAK)/signal transducer and activator oftranscription (STAT) signalling pathway has a primary rolein the regulation of growth and development, particularlyof haematopoietic cells. For example, it functions in theinterleukin-2 signalling pathway responsible for drivingthe final signalling steps in lymphocyte activation (Mod-ule 9: Figure T cell signalling map).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �91
Module 2: Figure JAK/STAT function
JAK
STAT
Agonist
PP P P P
P P P
P P P
P P P1 2
3
4
5
6
7
8
P
P
CRM1
SH2 domainDNA-binding domain
Importin
ImportinCRM1
ImportExport
Target genesSOCS
P
P
N-PTP
SOCS
_
The Janus kinase (JAK)/signal transducer and activator of transcription (STAT) activation cascade.A schematic summary of the main sequence of events responsible for the activation of signal transducers and activators of transcription (STATs).Agonist activation of cell-surface receptors induces dimerization of receptor subunits, resulting in the stimulation of resident Janus kinases (JAKs). TheJAKs then carry out a sequential series of three phosphorylation reactions to activate the STATs, which then enter the nucleus to induce transcription.The binding between STATs and DNA is shown in Module 2: Figure STAT1/DNA complex.
A mutation in the γc component of cytokine recept-ors results in X-linked severe combined immune defi-ciency (X-SCID). Since JAK3 associates with γc, it seemedlikely that X-SCID may result from an alteration in theJAK/STAT signalling pathway. This was confirmed whenSCID was found in patients carrying mutations in JAK3.SCID resulted from a dramatic reduction in the number ofT cells, highlighting the important role for this signallingpathway in lymphocyte development.
JAKs have also been implicated in certain forms of ma-lignant transformation such as Sezary’s syndrome, v-Abl-transformed cells and in some leukaemias.
Smad signalling pathwayThis signalling pathway takes its name from the Smads,which are a collection of intracellular signalling moleculesthat act collectively to transfer information from cell-surface receptors into the nucleus. As such, some of theSmads function as transcription factors, whereas oth-ers either facilitate or inhibit this transcriptional activ-ity. These Smads mediate the action of the transforminggrowth factor β (TGF-β) superfamily, which are cytokinesthat regulate many cellular functions such as proliferation,apoptosis, extracellular matrix formation and angiogen-esis. In addition, they play a critical role in controllingevents during early development and cell differentiation.There is an extensive Smad signalling toolkit, and the manycomponents can be mixed and matched to assemble a largevariety of Smad signalling pathways. The domain struc-ture of the Smad family illustrates their multifunctional
roles in the Smad signalling mechanism that functions totransfer information from activated receptors on the cellsurface to gene targets in the nucleus. This signalling mech-anism has two main components. Firstly, there is a processof transforming growth factor β (TGF-β) receptor ac-tivation, which is responsible for activating the Smads.Secondly, there is the Smad activation of transcription.
There are a number of mechanisms for the modulation ofSmad signalling. Transforming growth factor β (TGF-β)inhibition of cell proliferation is one of the major functionsof the Smad signalling pathway. It plays an important rolein the differentiation of intestinal cells. Alterations in thesignalling pathways controlled by the TGF-β superfamilyhave been associated with many cancers of epithelial andlymphoid origins, which fail to respond to the normal anti-proliferative effects of TGF-β. The TGF-β receptor is oneof the tumour suppressors that are switched off in manycancers and particularly in colorectal cancer (CRC).
An increase in the expression of TGF-β1 may play acritical role in the transition from stable hypertrophy tocongestive heart failure (CHF).
Smad signalling toolkitThe Smad signalling pathway is made up from a largenumber of components (Module 2: Table Smad signallingtoolkit). Specific cell types express different combinationsof these components so that there is considerable variab-ility in the nature of Smad signalling pathways. However,the overall organization is fairly similar and is best ex-emplified by the action of transforming growth factor β
(TGF-β) itself. The different components of the toolkit
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �92
Module 2: Table Smad signalling toolkitComponents of the Smad signalling toolkit.
Component Comment
Ligand trapsLAP Latency-associated polypeptide
that binds TGF-βDecorin Binds TGF-βα2-Macroglobulin Binds TGF-βNoggin Binds to BMPs (Module 8: Figure
epidermal stem cell)Follistatin Binds to activins and BMPs. It
inhibits activin action ongonadotrophs (Module 10:Figure gonadotroph regulation)
Chordin/SOG Binds to BMPsDAN/Cerberus Binds to BMPs
LigandsTGF-β1 Transforming growth factor β
(Module 1: Figure enzyme-linkedreceptors)
TGF-β2TGF-β3Activin One of its functions is to increase
the transcription of FSH ingonadotrophs (Module 10:Figure gonadotroph regulation)
Myostatin Growth and differentiation factor 8(GDF8) (see Module 8: Figuresatellite cell activation)
NodalBMP Bone morphogenetic factor
functions in the control ofepidermal stem cells (Module 8:Figure epidermal stem cell),proliferation of SNO cells(Module 8: Figure HSCregulation) and differentiation ofwhite fat cells (Module 8: Figurewhite fat cell differentiation)
Inhibin Inhibits activin action ingonadotrophs (Module 10:Figure gonadotroph regulation)
Accessory receptorsBetaglycan Facilitates the binding of TGF-β to
the Type II receptors. Mediatesthe action of inhibins ingonadotrophs (Module 10:Figure gonadotroph regulation)
CriptoEndoglin
Signalling receptorsType I receptors
ALK1 Activin receptor-like 1ALK2 Activin receptor-like 2ALK3 Activin receptor-like 3ALK4 Activin receptor-like 4ALK5 Activin receptor-like 5ALK6 Activin receptor-like 6ALK7 Activin receptor-like 7
Type II receptorsActRIIActRIIBBMPRIITβRII
Receptor-regulated Smads(R-Smads)Smad1Smad2Smad3Smad5Smad8
Co-mediator Smad(Co-Smad)Smad4 A germ-line mutation in Smad4 has
been linked to juvenile polyposissyndromes (JPSs).
Inhibitory Smads (I-Smad)Smad6Smad7
Module 2 Table continued
Component Comment
Smad regulatory factorsSARA Smad anchor for receptor activationSmurf1 Smad ubiquitin-regulatory factor 1.
Smirf1 also regulates thedegradation of Rho duringneutrophil chemotaxis (Module11: Figure neutrophilchemotactic signalling)
Smurf2 Smad ubiquitin-regulatory factor 2
have been grouped together on the basis of their functionin the signalling pathway (Module 2: Table Smad signallingtoolkit):
Ligand trapsThese are a large group of proteins that control accessof the ligands to their cell-surface receptors. These ligandtraps can have an important inhibitory action since theycan prevent the ligands from reaching their cell-surfacereceptors. For example, follistatin inhibits the action ofactivin on gonadotrophs (Module 10: Figure gonadotrophregulation).
LigandsThe Smad signalling pathway is activated by a number ofclosely related ligands. The flagship ligand is transforminggrowth factor β (TGF-β), but there are a number of otherligands that also act through the Smads.
Accessory receptorsThese are cell-surface receptors that function as co-receptors in that they promote the binding of the ligandsto their signalling receptors.
Signalling receptorsThere are two groups of signalling receptors (Type Iand II). These receptors contain a single membrane-spanning region that separates the extracellular ligand-binding domain from the cytosolic region that containsa serine/threonine kinase domain. The Type I receptoralso contains a glycine/serine-rich (GS) domain, which isphosphorylated by the Type II receptor during the signaltransduction process. Ligands act to complex these recept-ors so that the Type II can then activate the Type I, whichin turn activates the Smads.
SmadsThe Smads are the intracellular transducers of the Smadsignalling pathway. There are three Smad types. Receptor-regulated Smads (R-Smads), which are activated by thesignalling receptors, carry information into the nucleus. Asingle co-mediator Smad (Co-Smad) acts together with theR-Smads. Inhibitory Smads (I-Smads) set up a negative-feedback loop to limit the activity of the R-Smads. Thesedifferent activities are reflected in the domain structure ofthe Smad family.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �93
Module 2: Figure Smad domain structure
MH1
MH1
MH2
MH2
MH2
--SSXS-
NLS
NLS NES
Smurf1/2 binding
Smurf1/2 binding
Type I receptorphosphorylation sites
MAPK phosphorylation sites
CaMKII phosphorylation sitesPKC phosphorylation sites
PY
PY
PP (Smad1,2,3,5,8)
R-Smads
Co-Smad (Smad4)
I-Smads (Smad6,7)
The domain structure of the three Smad family members.The receptor-regulated Smads (R-Smads) have a MAD homology domain 1 (MH1) in the N-terminal region and an MH2 in the C-terminal region.These two domains play a critical role in carrying out the protein–protein and protein–DNA interactions. A nuclear localization signal (NLS) is locatedwithin MH1 and functions in the transport of the R-Smads into the nucleus. The SSXS motif at the C-terminus contains the two serine residues thatare phosphorylated by the Type I receptors during the process of signal transduction (Module 2: Figure TGF-βR activation).
Smad regulatory factorsThere are a number of proteins that contribute to the Smadsignalling pathway.
Domain structure of the Smad familyThe main structural feature of the Smads are the two MADhomology domains (MH1 and MH2) that function bothin the protein–protein and protein–DNA interactions thatoccur during the process of signal transduction (Mod-ule 2: Figure Smad domain structure). The regulatorySmads also have a C-terminal SSXS motif, which is crit-ical for the transduction process because two of the serineresidues are phosphorylated by the type I receptors as partof the Smad signalling mechanism. There also are numer-ous other sites that are phosphorylated by various kinasesfrom other signalling pathways. The linker region betweenthe MH1 and MH2 domains contains a phosphotyrosine(PY) motif, which is an interaction site for the binding ofthe Smad ubiquitin-regulatory factors 1 and 2 (Smurf1/2),which is an ubiquitin ligase that controls the selective pro-teolysis of the Smads. The single co-mediator Smad (Co-Smad, i.e. Smad4) resembles the R-Smads in some aspects.It also has MH1 and MH2 domains, but here the latter issplit. In addition to the NLS, it also has a nuclear exportsignal (NES). Smad4 lacks the C-terminal phosphoryla-tion motif, but it does contain a number of phosphoryla-tion sites. The two inhibitory Smads (I-Smads) lack anMH1 domain, but they have the Smurf1/2-binding PYmotif.
Smad signalling mechanismThe Smad signalling mechanism can be divided into twoparts. Firstly, there is the process of transforming growthfactor β (TGF-β) receptor activation, which concernsthe way in which ligands such as TGF-β interact withthe signalling receptors to trigger Smad activation (Mod-ule 2: Figure TGF-βR activation). The critical aspects ofthis activation process are the phosphorylation reactionsthat occur within the receptor complex. The Type II re-ceptors are constitutively active and phosphorylate theType I receptors. These activated Type I receptors thenact to phosphorylate the Smads.
The second part is the Smad activation of transcrip-tion, during which the phosphorylated receptor-regulatedSmads (R-Smads), together with their partner Smad4,translocate into the nucleus to induce gene transcription(Module 2: Figure Smad signalling).
Transforming growth factor β (TGF-β) receptoractivationActivation of the transforming growth factor β (TGF-β)receptor depends upon a series of reactions as illustrated bythe following steps shown in Module 2: Figure TGF-βRactivation.
1. The ligand, in this case TGF-β, is often held in a latentligand complex by being bound to one of the ligand trapproteins such as latency-associated polypeptide (LAP)or decorin. When it dissociates from the ligand trap, it
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �94
Module 2: Figure TGF-βR activation
P P
P MH1
MH2-SXSS--
MH1Glycine-serine-rich domain (GS)
Ser/Thr kinase domain
Betaglycan
1
2
3
4
4
5
TGF-β
SARA
SA
RA
SA
RA
TGF-β
Ligand trap (e.g. LAP, decorin)
Smad2Smad3
Type IIreceptors Type I
receptors
MH2 -SXSS--
(See Module 2: Figure Smad signalling)
P PP
MH1
MH2-SXSS--
GENE TRANSCRIPTION
Activation of the transforming growth factor β (TGF-β) receptor superfamily.The transforming growth factor β (TGF-β) receptor superfamily uses the Smad signalling pathway to transmit information into the cell. The mainfeatures of this pathway can be illustrated by considering the action of TGF-β, which activates the TGF-β receptor complex through a sequentialseries of reactions as described in the text.
can be taken up by one of the accessory receptors suchas betaglycan.
2. The TGF-β then associates with the two TGF-β re-ceptor components to assemble an agonist/receptorcomplex. In the absence of ligand, the Type I and IIreceptor components exist as homodimers which arethen brought together by TGF-β.
3. When the two receptor types have been complexedby TGF-β, the serine/threonine kinase domain on theType II receptors phosphorylates the serine residueson the glycine/serine-rich (GS) region of the Type Ireceptors.
4. These phosphorylated GS regions on the Type I re-ceptors provide a docking site for the MAD homologydomain 1 (MH1) domain of Smad2 or Smad3. This re-cruitment of the Smads to the membrane is facilitatedby Smad anchor for receptor activation (SARA). Onceattached to the receptor, the SSXS motif is brought intocontact with the serine/threonine kinase domain, andtwo of the serine residues are phosphorylated.
5. Once Smad2/3 have been phosphorylated, their affinityfor both the receptor complex and for SARA is reduced,and the two proteins pass into the cytoplasm. The ac-tivated Smads then translocate into the nucleus wherethey activate transcription (Module 2: Figure Smad sig-nalling).
Smad activation of transcriptionThe receptor-regulated Smads (R-Smads) function as tran-scription factors responsible for activating a large num-ber of target genes. Once they have been activated by thecell-surface receptors, the activated Smads translocate intothe nucleus (Module 2: Figure Smad signalling). Some ofthe ligands, such as transforming growth factor β (TGF-β), activin and Nodal activate receptors that are coupledto Smad2 and Smad3, whereas bone morphogenetic pro-
tein (BMP) acts through Smad1, Smad5 or Smad8. Oncethese Smads have been phosphorylated on their C-terminalSSXS motif (Module 2: Figure TGF-βR activation) theyleave the receptor, where they combine with Smad4 to forma dimer that then translocates into the nucleus (Module 2:Figure Smad signalling). Once the dimer enters the nuc-leus, it recognizes and binds to a specific DNA motif.
The transcriptional activity of the Smads is facilitatedby associating with other site-specific transcription factors(TFs). Their activity is also modulated by coactivators andrepressors. For example, Smad4 binds directly to the co-activator p300, which has an important role in activat-ing transcription. The pathway on the right, which car-ries out the action of receptors that bind ligands such asbone morphogenetic protein (BMP), use Smad1, 5 or 8 totransmit information into the nucleus. As for the pathwaydescribed above, Smad4 is again used as a partner for thetranslocation process, and the transcriptional processes arealso similar.
The Smads stimulate transcription of the collagen type Iduring the fibrogenesis induced in activated hepatic stellatecells in the liver (Module 7: Figure hepatic stellate cell).
Modulation of Smad signallingThere are a number of ways that the Smad signalling path-way can be modulated. One mechanism is by an increasein the expression of inhibitory Smads (I-Smads), whichtarget the cell-surface receptors for degradation. This pro-cess is mediated by the Smad ubiquitin-regulatory factors(Smurfs), which are a family of ubiquitin E3 ligases thatbind to the PY motif on the linker region of I-Smads(Module 2: Figure Smad domain structure). The E3 ligasefunctions in the ubiquitin-proteasome system (Module 1:Figure ubiquitin-proteasome system).
The mitogen-activated protein kinase (MAPK) sig-nalling pathway can also modulate Smad signalling by
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �95
Module 2: Figure Smad signalling
P
P
P P
P
P
Smad4
Coactivator/repressor
Coactivator/repressor
BMPs/GDFs
Smad2Smad3
Smad1Smad5Smad8
TGF /activintarget genes
β
TGF /activin/Nodalβ
SBE SBE
BMPtarget genesTF TF
The Smad signalling pathway links cell-surface transforming growth factor β (TGF-β) receptors to gene transcription.There are two main types of Smad signalling. In the one shown on the left, receptors activated by ligands such as transforming growth factor β (TGF-β),activin and Nodal phosphorylate either Smad2 or Smad3. The phosphorylated Smads then heterodimerize with the co-mediator Smad (Co-Smad)Smad4 to form a dimer that then translocates into the nucleus. Once in the nucleus, the Smad dimer then binds to a specific Smad-binding element(SBE), which has a GTCT motif that is recognized by the MAD homology domain 1 (MH1).
phosphorylating sites located in the linker regions ofSmad1 and 2 (Module 2: Figure Smad domain structure).This is an example of signalling cross-talk in that it providesa mechanism for the tyrosine kinase-linked receptors toantagonize the action of the transforming growth factor β
(TGF-β) superfamily.
Transforming growth factor β (TGF-β) inhibitionof cell proliferationOne of the major functions of transforming growth factorβ (TGF-β) is to inhibit cell proliferation by altering ex-pression of some of the key regulators of the cell cyclesignalling pathway. One of its actions is to increase thetranscription of the p15INK4B gene that codes for p15,which is one of the cyclin-dependent kinase (CDK) in-hibitors that inhibits cyclin D/cyclin-dependent kinase 4(CDK4) complex that is one of the early events of thecell cycle signalling cascade (Module 9: Figure cell cyclesignalling mechanisms). Another important action is torepress c-Myc, which is one of the major transcriptionalactivators of genes that function in cell proliferation.
TGF-β plays an important role in regulating the pro-liferation of mesangial cells (Module 7: Figure mesangialcells). It also regulates the proliferation of stem cells suchas the skeletal muscle satellite cells (Module 8: Figure Satel-lite cell activation) and the epidermal stem cells (Module8: Figure epidermal stem cell).
Smad2 and 4 are tumour suppressors. A germline muta-tion in Smad4 has been linked to juvenile polyposis syn-dromes (JPSs).
Wnt signalling pathwaysThe Wnt signalling pathways play a critical role in thecontrol of cell proliferation, cell fate specification and dif-ferentiation. These different pathways are activated by ex-tracellular lipoprotein signalling molecules called Wnts re-sponsible for transmitting information between cells overrelatively short distances. There are three main pathways:the canonical and two non-canonical pathways (Module 2:Figure Wnt signalling pathways).
The primary function of the canonical Wnt/β-cateninpathway is to activate gene transcription to control pro-cesses during both development and in the adult organism.The functions of the non-canonical Wnt pathways are stillsomewhat of an enigma, since their precise functions havenot been clearly identified, but there are clear indicationsthat they may function in planar cell polarity (PCP). TheWnt/planar cell polarity (PCP) pathway, which has beencharacterized in Drosophila, appears to act through variousGTP-binding proteins. A closely related Wnt/Ca2+ sig-nalling pathway, which has been described in vertebrates,has a number of similar signalling components but also hasadditional features such as the hydrolysis of PtdIns4,5P2 toactivate signalling through InsP3/Ca2+ and diacylglycerol(DAG)/protein kinase C (PKC).
WntsThe term Wnt results from the fusion of the names oftwo orthologous genes, the Drosophila segment polaritygene Wingless (Wg) and a mouse proto-oncogene Int-1.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �96
Module 2: Figure Wnt signalling pathways
LEF-1 TCF
β-catenin
p300
LRP5/6
Wntgenes
Wnt
Wnt
Wnt
Fz Fz
CR
D
CR
D
CR
D
PtdIns4,5P2PLC
DAG
Rho
ROK
Wnt/planar cell polarity pathwayCanonical Wnt/catenin pathway Wnt/Ca signalling pathway
JNK
?
?
Rac
PKC
InsP3
DshDsh
DAAM1
Dsh
Ca2+
2+
β-catenin
Myosin IIP
Wnt5A,11
NFATCaMKII
Fz2, Fz6
Summary of the signalling mechanisms used by the canonical and non-canonical Wnt signalling pathways.All three signalling pathways are activated by frizzled (Fz) receptors that depend upon the dishevelled protein (Dsh) as part of the transductionmechanism to transfer information into the cell. Most is known about the canonical Wnt/β-catenin pathway (Module 2: Figure Wnt canonical pathway)with less information on the two non-canonical pathways. In the Wnt/planar cell polarity (PCP) pathway, Dsh transfers information to the small GTP-binding proteins Rho and Rac. The Wnt/Ca2+ signalling pathway is connected to the dishevelled-associated activator of morphogenesis 1 (DAAM1),which relays information to the Rho pathway. In addition, Dsh also relays information to phospholipase C (PLC) to produce InsP3 and diacylglycerol(DAG).
The human genome has 19 Wnt genes, many of whichappear to have distinct functions. They contain numer-ous cysteine residues, one of which is palmitoylated, thusmaking the Wnts somewhat insoluble, which may help toexplain why they are short-range ligands. A large num-ber of Wnt-binding proteins, such as secreted frizzled-re-lated protein (SFRP) and Wnt inhibitory factor (WIF) alsorestrict their sphere of influence by acting as Wntbuffers.
Canonical Wnt/β-catenin pathwayThe defining feature of the canonical Wnt/β-catenin path-way is the transcription factor β-catenin, which is respons-ible for regulating the transcription of Wnt target genes(Module 2: Figure Wnt canonical pathway). However, thisis not the sole function of β-catenin, which also functionsas a scaffolding protein in cell adhesion by providing a linkbetween cadherin and the actin cytoskeleton. Here we con-sider how β-catenin functions in Wnt signalling to regulategene transcription. There are a number of transcriptionfactor activation mechanisms, and β-catenin belongs tothose that depend on cell-surface receptors that generatecytosolic signals to activate latent transcription factors inthe cytoplasm, which are then imported into the nucleus(Mechanism 2 in Module 4: Figure transcription factoractivation). In the case of β-catenin, its cytosolic level iskept low because it is constantly being degraded by theproteasome as shown in the sequence of events shown in
the left-hand panel in Module 2: Figure Wnt canonicalpathway:
1. Formation of β-catenin by protein synthesis occurscontinuously.
2. β-Catenin is drawn into a β-catenin degradation com-plex, which consists of the scaffolding protein axin,which binds the adenomatous polyposis coli (APC)tumour suppressor, the protein phosphatase PP2A,casein kinase Iα (CKIα), glycogen synthase kinase-3β
(GSK-3β) and β-catenin. Within this multiprotein de-gradation complex, the GSK-3β phosphorylates β-catenin and thus targets it for destruction by the pro-teasome. Before β-catenin can be phosphorylated byGSK-3β it must first be primed by phosphorylatingSer-45 by casein kinase Iα (CKIα). PP2A, which con-sists of a scaffolding A subunit and a regulatory B sub-unit (Module 5: Figure PP2A holoenzyme), may inac-tivate the complex by dephosporylating GSK-3β.
3. The phosphorylated β-catenin is recognized by the F-box/β-TrCP/ubiquitin ligase complex, which targets itfor destruction by the proteasome.
4. In the absence of β-catenin, the monomeric high-mobility group (HMG) DNA-binding proteins lymph-ocyte enhancer factor-1 (LEF-1) and T cell factor(TCF) inhibit the transcription of Wnt genes. An addi-tional component of this repressor complex is histonedeacetylase (HDAC), which prevents chromatin re-modelling through histone deacetylation. Wnt acts to
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �97
inhibit the destruction of β-catenin, which then accu-mulates in the cytoplasm and can enter the nucleus topromote transcription as shown in the right-hand panelin Module 2: Figure Wnt canonical pathway.
5. Wnt initiates the signalling process by binding togethertwo cell-surface proteins: it binds to the cysteine-richdomain (CRD) of the frizzled (Fz) receptor and italso draws in the Fz co-receptor LRP5 and LRP6(LRP5/6), which are members of the low-density lipo-protein receptor-related protein family (LRP).
6. The LRP5/6 co-receptor is then phosphorylated bymembrane-bound casein kinase Iγ (CKIγ) isoform,which adds phosphates to multiple sites that havePPPSP motifs. Some of these motifs are also phos-phorylated by a membrane-associated GSK-3β. Thesephosphorylated motifs then provide binding sites forthe attachment of the scaffolding protein axin. Anotherkey event is the binding of Dishevelled (Dsh), whichis another scaffolding protein containing various signaltransduction domains (e.g. DIX, PDZ and DEP). Dshbecomes hyperphosphorylated by an unknown mech-anism and this contributes to its role in inhibiting thedegradation complex. Axin and Dsh also bind to eachother through their DIX domains. As the multiproteincomplex associates with the membrane, the organiza-tion of the subunits is altered so that the activity ofGSK-3β is inhibited, thus reducing the degradation ofβ-catenin. Loss-of-function mutations of LRP5 havebeen linked to osteoporosis pseudoglioma (OPPG).
7. When GSK-3β is inhibited, the newly synthesized β-catenin is stabilized and accumulates within the cyto-plasm, from where it can enter the nucleus to activatetranscription.
8. β-Catenin binds to LEF-1 and TCF to reduce theirrepressor activity to initiate the transcription of the Wntgenes. β-Catenin replaces HDAC with p300, whichfacilitates transcription by acetylating histones to re-model chromatin.
9. Activation of transcription of the Wnt target genes res-ults in the activation of the developmental and prolifer-ative effects that characterize the operation of the Wntsignalling pathway.
In summary, the Wnt signalling pathway acts by switch-ing off the GSK-3β-dependent degradation pathway, thusenabling β-catenin to accumulate in the cytosol and toenter the nucleus to activate transcription of the Wnt tar-get genes.
Inhibitors of Wnt signallingThere are various extracellular molecules that can inhibitWnt signalling. Some of these, such as the Wnt inhibit-ory factor 1 (WIF-1) and secreted frizzled-related protein(sFRP), bind to Wnt and thus prevent it from activatingthe Fz receptor. There is another group, such as Dickkopf(Dkk) and sclerostin (SOST) that interfere with the activ-ity of the lipoprotein receptor-related protein (LRP) co-receptor LRP5/6. These inhibitors play an important rolein preventing the proliferation of stem cells until they arerequired for tissue repair as occurs for the epidermal stem
cells used for the hair follicle cycle (Module 8: Figure epi-dermal stem cell).
Wnt inhibitory factor 1 (WIF-1)The Wnt inhibitory factor 1 (WIF-1) is a soluble extracel-lular factor that binds to Wnt and prevents it from inter-acting with the frizzled receptor (Module 2: Figure Wntcanonical pathway).
Secreted frizzled-related protein (sFRP)The secreted frizzled-related protein (sFRP) is a solubleinhibitor that binds to Wnt and prevents it from interactingwith the frizzled receptor (Module 2: Figure Wnt canonicalpathway).
Dickkopf (Dkk)Dickkopf (Dkk) binds to Kremen and the lipoproteinreceptor-related protein (LRP) co-receptor LRP5/6 andthis causes the complex to internalize and will thus inac-tivate Wnt signalling.
Sclerostin (SOST)Sclerostin (SOST) inhibits Wnt signalling by binding tothe lipoprotein receptor-related protein (LRP) co-receptorLRP5/6 (Module 2: Figure Wnt canonical pathway). SOSTis produced by the osteoclasts to inhibit the proliferationand differentiation of the osteoblasts (Module 8: Figurebone cell differentiation).
Van Buchem disease is caused by a mutation in SOSTthat results in excessive activation of the Wnt signallingpathway.
Function of canonical Wnt signallingThe canonical Wnt pathway has an important role to playin regulating processes such as development and prolifer-ation:
• It functions in dorsoventral specification, as has beenshown for amphibia and zebrafish (Module 8: Figuredorsoventral specification).
• It functions in the control of stem cell proliferation(Module 8: Figure stem cell function). For example, Wntsignalling stimulates the proliferation of epidermal stemcells during the hair follicle cycle (Module 8: Figure epi-dermal stem cell).
• It controls the processes of osteoblastogenesis, whichis responsible for the development of the bone-formingosteoblasts (Module 8: Figure bone cell differentiation).
• It controls the differentiation of intestinal cells and al-terations in its signalling components are a major causeof colorectal cancer (CRC) (Module 12: Figure coloncancer).
• The canonical Wnt pathway inhibits the differentiationof white fat cells (Module 8: Figure white fat cell dif-ferentiation) and the differentiation of brown fat cells(Module 8: Figure brown fat cell differentiation).
The adenomatous polyposis coli (APC) protein, whichcontributes to the cytoplasmic complex that degrades β-catenin in the cytoplasm, is a potent tumour suppressorthat is frequently mutated in cancer cells and partic-ularly in those that develop within the intestine such
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �98
Module 2: Figure Wnt canonical pathway
Ac Ac Ac AcAc
HDAC
LEF-1
LEF-1 TCFTCF
Deacetylation Acetylation
4
1
2
3
5
6
7
8 9
β-catenin
β-catenin
β-ca
ten
inβ-catenin
β-cateninβ-catenin
β-catenin
β-catenin
-catenin protein synthesis anddegradation cycle
β
p300
APC
APCPP
P
PP P
Axin
Axi
nDsh
Dsh
LRP5/6
LRP5/6LRP5/6
Kremen
Proteinsynthesis
Proteinsynthesis
Amino acids
Amino acids
Proteasome
GSK3β
GS
K3β
CKIα
CK
Iα
Wntgenes
Wnt
Wnt
WntCRDFz Fz
CR
D
β-catenin accumulates and translocates into the nucleus β-catenin
degradation complex
PP
PP2A
B
AB A
PP
2A
CKIγCKIγ PP
Dkk1SOST
WIF1 WIF1
SFRP SFRP
The canonical Wnt/β-catenin signalling pathway.The primary function of this signalling pathway is to regulate the activity of β-catenin, which controls transcription of the Wnt genes. The left-handpanel illustrates the resting condition where the cytosolic level of β-catenin is kept low by its continuous degradation. In response to the arrival of Wnt(as shown on the right), this degradation is inhibited and the level of β-catenin rises enabling it to induce the transcription of the Wnt genes.
as colorectal cancer (CRC). Germline mutations ofAPC are responsible for familial adenomatous polyposis(FAP). Mutation in the LRP5 gene, which binds to theWnt antagonist Dickkopf1 (DKK1), causes osteoporosispseudoglioma (OPPG) syndrome.
Wnt/planar cell polarity (PCP) pathwayThe Wnt/planar cell polarity (PCP) pathway has beencharacterized in insects, where it functions to establishplanar cell polarity (PCP) during development (Mod-ule 2: Figure Wnt signalling pathways). Just how thispathway functions is still unclear, but there are indica-tions that it relays information through the Rac signallingmechanisms and the Rho signalling mechanism, both ofwhich function in the remodelling of actin. The Rho path-way acts through the Rho kinase (ROK) to activate con-traction of the actin/myosin system (Module 2: FigureRho signalling). These effects on actin remodelling andcontraction may play an important role in planar cellpolarity.
In vertebrates, there are similar PCP processes such asneural tube closure, the orientation of hair cell stereociliarybundles in the ear, mammalian hair follicle orientation andconvergent extension (CE), during which there are large-scale movements of cells that occur during gastrulation.Components of the insect planar cell polarity pathwayhave also been described in vertebrates, where it includessignalling through Ca2+ and has thus been referred to asthe Wnt/Ca2+ signalling pathway.
Wnt/Ca2+ signalling pathwayThe Wnt/Ca2+ signalling system (Module 2: Figure Wntsignalling pathways) has been implicated in a number ofplanar cell polarity (PCP) processes in vertebrates. Thereare a number of similarities between this signalling path-way and the PCP pathway in Drosophila. They both canactivate Rho and contraction of the actin/myosin system.The main difference is that the vertebrate system has an ad-ditional signalling component in that it can activate Ca2+
signalling. The activation of this Wnt/Ca2+ signalling path-way depends upon the frizzled receptor plugging into theinositol 1,4,5-trisphosphate (InsP3)/Ca2+ signalling cas-sette (Module 2: Figure InsP3 and DAG formation). Thereare reports that the Ca2+ signalling events associated withthis pathway may act to inhibit the canonical Wnt/cateninpathway during dorsal/ventral axis specification (Module8: Figure dorsoventral specification).
Mutations of the Fz4 receptor, which activates thisWnt/Ca2+ pathway, have been linked to familial exudativevitreoretinopathy (FEVR).
Casein kinase I (CKI)The casein kinase I (CKI) family has seven members(CKIα, α2, δ, ε, γ1, γ2 and γ3), which can have verydifferent functions in cells.
Casein kinase Iα and α2CKIα functions as a priming kinase for glycogen syn-thase kinase-3β (GSK-3β) during the operation of the
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �99
Wnt signalling pathway (Module 2: Figure Wnt canonicalpathway).
Casein kinase IδCasein kinase Iδ (CKIδ), which is 97% homologous withcasein kinase Iε (CKIε), contributes to the operation ofthe circadian clock by phosphorylating PER1 and PER2to control the nuclear entry and stability of these clock pro-teins (Module 6: Figure circadian clock molecular mech-anism).
Casein kinase IεCKIε participates in circadian rhythmicity by phos-phorylating the PER and CRY proteins to regulate theirnuclear entry and stability during the operation of thePER regulatory loop (Module 6: Figure circadian clockmolecular mechanism). Mutation of CKIε, which resultsin a decrease in the ability of this kinase to phosphorylatethe PER proteins of the circadian clock, is responsible forfamilial advanced sleep phase syndrome (FASPS).
Casein kinase Iγ1, γ2 and γ3CKIγ is an unusual member of the family in that it hasa palmitoylation site at its C-terminus and this fatty acidanchor attaches it to the plasma membrane. This membranelocation is critical for one of its main functions, which isto phosphorylate the frizzled lipoprotein receptor-relatedprotein (LRP) co-receptor LRP5/6 during activation of theWnt signalling pathway (Module 2: Figure Wnt canonicalpathway).
Casein kinase II (CK2)Casein kinase II (CK2) is a serine/threonine protein kinase.It functions as a heterotetramer consisting of two 44 kDacatalytic α-subunits and two regulatory β-subunits. It hasa unique ability to use GTP as well as ATP as a phosphatedonor. CK2 is also known as phosvitin kinase, glycogensynthase kinase 5, troponin T kinase and casein kinase G,which reflects the fact that CK2 can phosphorylate manydifferent substrates and thus contributes to many controlmechanisms. This multifunctional kinase has been implic-ated in many cellular processes and seems to be particu-larly active in controlling cell proliferation and has alsobeen implicated in cell transformation and tumorigenesis.Many of its actions depend on its ability to phosphorylatetranscription factors such as Myc, p53, Rb and activatingprotein 1 (AP-1). Although CK2 is constitutively active,it can also be activated by certain growth factors (insulin,IGF-I and EGF).
Hedgehog signalling pathwayThe Hedgehog signalling pathway in mammals closely re-sembles that originally discovered and characterized inDrosophila. The mammalian Hedgehog signalling toolkithas many of the components found in insects. In compar-ison with the latter, however, much less is known aboutthe mechanism of Hedgehog activation of transcription inmammals. There are multiple Hedgehog signalling func-tions that operate during both development and adult life.
Hedgehog signalling toolkitThe Hedgehog signalling pathway has a number of com-ponents (Module 2: Table Hedgehog signalling toolkit)whose names resemble those given to similar componentsin Drosophila, where this signalling system was origin-ally identified and characterized. While many of the mam-malian components have similar functions to those foundin insects, it is clear that there are important differences.Since there are mammalian components that are not foundin insects, the mechanism of Hedgehog activation of tran-scription has to be worked out separately for vertebrates.
Hedgehog activation of transcriptionHedgehog mediates its effects by activating gene tran-scription. Hedgehog uses mechanism 2 of the differenttranscription factor activation mechanisms found in cells(Module 4: Figure transcription factor activation). Thereare three Hedgehog transcription factors (GLI 1–3), whichare held in an inactive state within the cytoplasm in rest-ing cells (Module 2: Figure Hedgehog signalling path-way). This inactive state is maintained by the Hedge-hog receptor patched (PTC), which inhibits the seven-membrane-spanning protein smoothened (SMO) that actsas the Hedgehog transducer. In the absence of a signal fromSMO, the GLI transcription factors are maintained in a lat-ent state by interacting with a large number of cytoplasmicfactors (Module 2: Table Hedgehog signalling toolkit). Theprecise function of all these factors is still being workedout. Hedgehog arriving at the cell surface induces a train ofevents that activate these transcription factors so that theytranslocate into the nucleus to induce gene transcription(Module 2: Figure Hedgehog signalling pathway). First ofall, Hedgehog binds to its receptor PTC and this removesthe inhibitory effect of PTC on SMO. The latter is thenable to activate GLI by removing it from the inhibitoryconstraints of the cytoplasmic factors so that it is now freeto translocate into the nucleus to activate transcription.Some of the genes that are activated are components of theHedgehog signalling pathway and thus set up both posit-ive and negative feedback loops. Many of the other genescontribute to Hedgehog signalling functions.
Hedgehog signalling functionsThe Hedgehog signalling pathway functions both dur-ing development and during adult life. Its function dur-ing development is wide-ranging in that it can controlcell proliferation, cell determination and pattern forma-tion of the developing embryo and the final processes ofcell differentiation. Hedgehog signalling seems to play animportant role in controlling the development of organssuch as the skin, brain, digestive tract, pancreas and pro-state. Mutations of components of this signalling pathwayresult in congenital malformations such as holoprosen-cephaly, which is a cranial defect that occurs when themidline structures of the brain and face fail to separate.The most dramatic phenotype of such malformation iscyclopia where only one eye is formed.
The function of Hedgehog continues in the adult or-ganism, where it plays a major role in the formation andmaintenance of the stem cell population. Since Hedgehog
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �100
Module 2: Table Hedgehog signalling toolkitThe Hedgehog signalling toolkit.Component CommentHedgehog (Hh) ligands
Shh Sonic HedgehogIhh Indian HedgehogDhh Desert Hedgehog
Hedgehog-binding proteinsHIP Hh-interacting proteinMegalin A member of the low-density lipoprotein (LDL)-receptor-related family
Hedgehog receptorsPTC1 Patched 1PTC2 Patched 2
TransducerSMO Smoothened; a seven-membrane-spanning protein
Cytoplasmic regulatorsKIF7 Functions to repress Sh responsesFU The fused serine/threonine protein kinaseSUFU Suppressor of fused; a negative regulator of GLI transcription factorsMIM Missing in metastasis; an actin-binding protein that potentiates transcriptionIguana A zinc-finger protein that promotes localization of GLI1IFT88 Ciliary protein that regulates GLI functionIFT172 Ciliary protein that regulates GLI functionFKBP8 Antagonizes Shh actionSIL Cytoplasmic protein that acts downstream of PTCRab23 This regulator of vesicular traffic is a negative regulator of Hh signalling
Transcription factorsThe multifunctional GLI transcription factors function toeither activate or repress transcription
GLI1 A zinc-finger transcriptional activatorGLI2 A zinc-finger transcriptional activator/repressorGLI3 A zinc-finger transcriptional repressor
The function of some of these toolkit components are illustrated in Module 2: Figure Hedgehog signalling pathway.
Module 2: Figure Hedgehog signalling pathway
SMO SMO PTCPTC
Wnt, BMP
PTC, HIP
GLI1, MIM
KIF7 KIF7
KIF7
GLI1
GLI1
GLI1
GLI1
GLI1
Iguana MIM
SU SU
SU
HIP
SUFUSUFU
X
Positive feedback
Negative feedback
++
Hh
Hh
Hh
Activation of gene transcription by the Hedgehog signalling pathway.Under resting conditions, the patched (PTC) receptor for Hedgehog (Hh) inhibits the activity of the seven-membrane-spanning receptor smoothened(SMO). In this inhibited state, SMO is not able to act on the complex of cytoplasmic factors that collectively regulate the transcription factor GLI1.When Hh is present, it binds to PTC, and this removes the inhibitory effect of the latter on SMO, which is now capable of activating transcriptionby releasing GLI from its associated cytoplasmic factors. GLI1 then translocates into the nucleus, where it activates a variety of genes that fall intothree main groups. One group consists of components of the signalling pathway, such as GLI1 and missing in metastasis (MIM), which set up apositive-feedback loop. The second group consists of genes coding for PTC and Hh-interacting protein (HIP), which thus set up a negative-feedbackloop. The HIP protein acts on the outside to bind Hh and thus reduce its activity. The last group of genes code for proteins not involved in Hedgehogsignalling, but some do function in other signalling pathways such as Wnt (Module 2: Figure Wnt canonical pathway) and the bone morphogeneticproteins (BMPs) (Module 2: Figure Smad signalling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �101
Module 2 : Figure Notch signalling
Acetylation
1 2
3
4
9
10
5
6
7
8
P
P
ADAM
PEST
S2
S36 Ankyrin repeats
CSLX
Gene activationGene repression
CSL
CDK-8SEL-10
Notch target genes
Endosome
γ-secretase
NICD
NICD
HES1HES2HES7HEY1HEY2HEYLESRNRARP
DeltaJagged
Notch
SHARP
Proteasome
MAMp300
UbUbUbUb
HDACDeacetylation
SMRT
SKIP
Notch signalling pathway.One of the stimuli for the Notch signalling pathway is Jagged, which is located on the surface of the communicating cell. The receptor for Jagged isNotch, which is located on the surface of the receiving cell. When Jagged interacts with Notch (Step 1), it triggers a series of steps that result in theproteolytic release of the Notch intracellular domain (NICD), which then enters the nucleus to activate transcription of Notch target genes. See the textfor details of this sequence of events.
signalling plays such a major role in regulating cell prolif-eration, it is not surprising that alterations in this signallingpathway have been detected in many cancers such as skincancer and some brain cancers.
For example, patched (PTC) is a tumour suppressor thatis inactivated in basal cell carcinomas (BCCs), medullo-blastomas, gliomas in the brain and prostate cancers.
Mutations of the PTC gene are responsible for Gorlin’ssyndrome, which is also known as nevoid basal cell car-cinoma syndrome (NBCC). The Hedgehog signallingpathway may also contribute to the growth of tumoursby enhancing the activity of stromal cells that provide thetumour cell microenvironment that supports cancer cellsurvival and growth. Cancer cells release hedgehog thatthen uses a paracrine mechanism to stimulate neighbouringstromal cells such as the blood vessels, fibroblasts, immunecells and epithelial cells. These stromal cells assist cancercell growth by providing both an extracellular matrix andessential growth factors such as insulin-like growth factor(IGF) and Wnt.
Notch signalling pathwayThe Notch signalling pathway is a highly conserved sig-nalling system that functions in both development andadulthood. Many of its functions relate to cell-fate de-termination and this is particularly the case in its controlof binary cell-fate decisions in stem cells. During the assy-metrical divisions of stem cells, one cell retains its stem cell
fate (self-renewal) while the daughter cell (progenitor cell)adopts a different state that will drive it towards prolifera-tion and differentiation into a specific cell type (Module 8:Figure stem cell function). As these progenitor cells adopttheir new cell fate, they use the Notch signalling pathwayto feed information back to suppress their neighbour fromadopting a similar cell fate. This is a short-range informa-tion transfer mechanism that depends upon direct contactbetween the cells, which is a hallmark of this signallingpathway. For example, the stimulus Jagged is an integralmembrane protein located on the surface of communic-ating cells, whereas the Notch receptor that responds toJagged is embedded in the surface of the receiving cell(Module 2: Figure Notch signalling).
The main feature of the transduction mechanism is de-ceptively simple. When Jagged binds to Notch, the Notchintracellular domain (NICD) is released into the cytoplasmand then diffuses into the nucleus where it induces thetranscription of multiple Notch target genes. Despite itssimplicity, this signalling system has an extensive numberof interacting components (Module 2: Table Notch sig-nalling components). Some of these function during thesignal transduction process itself, whereas others play arole in the modulation of Notch signalling, which regu-lates the expression of the stimuli (Delta and Jagged) andthe Notch receptor at the cell surface.
The operation of the Notch signalling pathway dependsupon the following steps (Module 2: Figure Notch sig-nalling):
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �102
Module 2: Table Notch signalling componentsNotch signalling toolkit.Components CommentsNotch stimuli These Notch stimuli contain an N-terminal DSL (Delta, Serrate and LAG-2)
domain that interacts with the Notch receptor (Module 2: Figure Notchsignalling)
Delta 1Delta 3Delta 4 Responsible for lateral inhibition during angiogenesis (Module 9: Figure
angiogenesis signalling)Delta-like-4 (DLL4)Jagged 1Jagged 2
ReceptorsNotch 1Notch 2Notch 3Notch 4
Notch modulatorsNumb A major inhibitor of Notch. It facilitates the endocytic removal of Notch from the
plasma membrane (Module 2: Figure Notch modulation)
Crumbs May function to inhibit γ-secretase activity thereby reducing the activationof Notch
ProteasesADAM-10 Also known as Kuzbanian or MADM. Functions in neural and cardiac
developmentADAM-17 Also known as tumour necrosis factor-α (TACE)Furin A protein convertase that cleaves site 1 (S1) during Notch receptor maturationγ-secretase An enzyme complex containing presenilin, nicastrin, PEN2 and APH1
GlycosyltransferasesFringe family A family of glycosyltransferase in the Golgi that glycosylates Notch to alter its
binding affinities to Delta and JaggedLunatic fringeRadical fringeManic fringe
Transcriptional regulatorsCSL
CSL [CBF-1, Su(H), Lag-1] represses Notch target genes (Module 2: FigureNotch signalling)
MAM MastermindSKIP Ski-interacting protein interacts with CSL and the ankyrin repeats of NICD
Notch target genesHES1 Hairy and enhancer of split 1 (HES1) represses transcription of tissue specific
genes responsible for differentiationHES2HES7HEY1HEY2HEYLESR
Ubiquitin ligasesItch A Hect domain E3 ligase that functions in Notch trafficking (Module 2: Figure
Notch modulation)Deltex A Ring finger E3 ligase that functions in Notch trafficking (Module 2: Figure
Notch modulation)Mind bomb (Mib) Ring finger E3 ubiquitin ligases that initiate the trafficking of Delta and Jagged
(Module 2: Figure Notch modulation)Neutralized (Neur) Ring finger E3 ubiquitin ligases that initiate the trafficking of Delta and Jagged
(Module 2: Figure Notch modulation)SEL10 A nuclear ubiquitin ligase that interacts with the phosphorylated form of NICD
(see Step 8 in Module 2: Figure Notch signalling)
1. Delta or Jagged located on one cell interacts withthe Notch receptor on a neighbouring cell. The N-terminal DSL (Delta, Serrate and LAG-2) domain(yellow bar) on Delta/Jagged binds to the EGF-repeats 11 and 12 (red bar) on Notch. By itself, thisinteraction with Notch seems to have little effect andonly initiates the signal transduction sequence as a res-ult of Delta/Jagged physically pulling on Notch.
2. The endocytosis of Delta/Jagged seems to be a crit-ical step for receptor activation. As Delta/Jagged are
drawn into the endosome, they pull on Notch, whichundergoes a conformational change to expose site-2(S2).
3. S2 is cleaved by ADAM proteases such as ADAM-10and ADAM-17 (Module 1: Figure ADAMs proteases).The external domain is shed leaving behind a shorttransmembrane region and the intracellular domain,which then becomes a substrate for γ-secretase.
4. γ-Secretase is an enzyme complex made up ofpresenilin, nicastrin, PEN2 and APH1 that cleaves
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �103
the S3 site to release the Notch intracellular domain(NICD). The activity of γ-secretase may be inhibitedby Crumbs, which is an apical polarity protein.
5. NICD released from the membrane diffuses into thenucleus and binds to CSL (CBF-1, Suppressor of Hair-less, Lag-1).
6. CSL normally acts to repress Notch target genes.Repression is enhanced by CSL providing a frame-work to recruit co-repressors such as SMRT, SHARP,SKIP and CIR. This repression complex also recruitshistone deacetylase (HDAC) that deacetylates chro-matin further shutting down transcriptional activity.When NICD binds to CSL, the repression complex isdisassembled and this paves the way for the assemblyof an activation complex. An important part of thecomplex is the co-activator Mastermind (MAM) andthe histone acetyl transferase p300. The latter is re-sponsible for protein acetylation of histones to openup the chromatin to facilitate gene transcription.
7. Gene activation results in an increase in the transcrip-tion of Notch target genes. Some of these such as thehairy and enhancer of split (HES) family encode tran-scriptional repressors responsible for suppressing theexpression of tissue specific proteins, which accountsfor the ability of the Notch signalling pathway to in-hibit differentiation.
8. The inactivation of target gene transcription beginswhen NICD is phosphorylated in the N-terminalPEST domain by kinases such as cyclin-dependentkinase-8 (CDK-8).
9. The phosphorylated NICD becomes a substrate fornuclear ubiquitin ligases such as SEL-10 and is thenexported to the cytoplasm where it is degraded by theproteasome.
10. Once NICD is removed from the activation com-plex and degraded, the different co-activators fallaway and are replaced by the repressors that resultin gene repression. Gene repression also depends onthe recruitment of histone deacetylase (HDAC) thatdeacetylates chromatin further shutting down tran-scriptional activity.
Notch signalling controls a number of developmentalevents and continues to have a role during adulthood par-ticular in maintaining the stem cell population:
• Proliferation of satellite cells in skeletal muscle is en-hanced by Notch signalling (Module 8: Figure Satellitecell activation).
• The self renewal of haematopoietic stem cells (HSCs)is facilitated by Notch signalling that acts by inhibitingdifferentiation (Module 8: Figure HSC regulation).
• Notch signalling is responsible for the process of lat-eral inhibition that inhibits tip cell formation duringangiogenesis (see Step 2 in Module 9: Figure angiogen-esis signalling).
Alterations in Notch signalling have been implicated in anumber of cancers. For example, overexpression of Notchhas been identified in both ovarian and medullablastomas.An up-regulation of Notch and a decrease in the inhib-
itor Numb has been observed in breast cancer. Mutationsin Notch have been linked to T-cell acute lymphoblasticleukaemia.
Modulation of Notch signallingThe primary mechanism for modulating Notch signallingis to adjust the mechanisms responsible for regulating bothligand (Delta and Jagged) and receptor (Notch) maturation(Module 2: Figure Notch modulation). In both cases, theligands and the receptor on the cell surface are taken upinto the endosomes and are then either recycled back to thecell surface or are degraded. The balance between recyclingand degradation thus determines their level of membraneexpression and thus sets the sensitivity of the Notch sig-nalling pathway. The ubiquitin-proteasome system playsa critical role in regulating the trafficking of these twosignalling components.
The trafficking of Delta and Jagged follows the fol-lowing series of events (Module 2: Figure Notch mod-ulation):
1. The cytoplasmic domain of Delta and Jagged are ubi-quitinylated by Ring finger E3 ubiquitin ligases such asMind bomb (Mib) and Neutralized (Neur).
2. Endocytosis of these ubiquitylated proteins is facil-itated by the protein epsin that binds to ubiquitin-containing cargo proteins. In addition, epsin bindsto both clathrin and to phosphatidylinositol 4,5-bisphosphate (PtdIns4,5P2), which functions in thePtdIns4,5P2 regulation of membrane trafficking and en-docytosis.
3. The endosome then has two paths: it can move downthe lysosomal path resulting in degradation of Delta andJagged. Alternatively, it can recycle back to the plasmamembrane where the ligands are then able to activateNotch.
Different players function in the regulation of Notch traf-ficking as shown in the following steps (Module 2: FigureNotch modulation):
4. Entry of Notch into the endosome is dependent on theprotein Numb, which is a major inhibitor of Notchsignalling. The Notch receptor interacts with Numb,which also binds to α-adaptin. The latter is a componentof the adaptor protein-2 (AP2) complex responsible forendocytosis. A Numb-associated kinase (NAK) is alsoa component of this complex. The Notch receptors inthe endosome have two fates.
5. Some of the Notch receptors are ubiquitinated byubiquitin ligases such as Itch (a Hect domain E3 lig-ase) and Deltex (a Ring finger E3 ligase).
6. Ubiquitinylated notch receptors are transferred to themultivesicular body (MVB) en route to degradation bythe lysosome.
7. Some of the Notch receptors are recycled back to theplasma membrane.
8. These Notch receptors on the cell surface are then ableto interact with ligands such as Delta and Jagged toinitiate the Notch signalling pathway (Module 2: FigureNotch signalling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �104
Module 2: Figure Notch modulation
1
23
4
56
7
8
DeltaJagged
Notch
NumbNAK
Neur
MVB
Endosome
α-adaptin
Ub
Ub
Ub
Ub
Ub Ub
UbUb
Ub
Ub
Deltex
Itch
Recycling
Recycling
Degredation
Degredation
Mib
Epsin
2 PtdIns4,5P
Ligand maturation
Receptor maturation
Modulation of Notch signalling.Trafficking through the endosomal compartment plays an important role in regulating the maturation/activity of the Notch ligands (shown at the top)and the Notch receptor (shown at the bottom). See text for further details. Mib, Mind bomb; MVB, multivesicular body; Neur, Neutralized; NAK,Numb-associated kinase; Ub, ubiquitin.
DeltaDelta is a transmembrane protein that functions as oneof the stimuli that activate the Notch receptor (Module2: Table Notch signalling components). The extracellu-lar domain is made up of EGF-like repeats and there isan N-terminal DSL (Delta, Serrate and LAG-2) domainthat interacts with the Notch receptor (Module 2: Fig-ure Notch signalling). In order for Delta to function as astimulus, it has to undergo a maturation process, whichdepends upon its passage through the endosome (Module2: Figure Notch modulation). This trafficking through theendosomal compartment determines the amount of Deltain the membrane and this is an important aspect of themodulation of Notch signalling.
The expression of Delta-1 is increased on the surfaceof myofibres during muscle damage and this activates theNotch signalling pathway to stimulate the proliferation ofsatellite cells (Module 8: Figure Satellite cell activation).
JaggedJagged is a transmembrane protein that functions as a oneof the stimuli that activate the Notch receptor (Module2: Table Notch signalling components). The extracellulardomain is made up of a cysteine-rich domain, EGF-likerepeats and there is an N-terminal DSL (Delta, Serrate andLAG-2) domain that interacts with the Notch receptor.In order for Jagged to function as a stimulus, it has toundergo a maturation process, which depends upon itspassage through the endosome (Module 2: Figure Notchmodulation). This trafficking through the endosomal com-partment determines the amount of Delta in the membrane
and this is an important aspect of the modulation of Notchsignalling.
NotchNotch is a transmembrane protein that functions as thereceptor for the Notch signalling pathway by respond-ing to stimuli such as Delta and Jagged (Module 2: FigureNotch signalling). The large extracellular domain of Notchis made up of a variable number (29–36) of EGF-like re-peats and a Lin21/Notch repeat region. EGF-repeats 11and 12 (red bar) on Notch provides the binding site that in-teracts with the N-terminal DSL (Delta, Serrate and LAG-2) domain on Delta and Jagged. The cytoplasmic domainhas a series of six ankyrin repeats and a terminal PESTdomain.
Notch undergoes a number of post-translational modi-fications as it moves through the endoplasmic reticulum(ER) and the Golgi. While in the ER, certain sites on theEGF repeats are fucosylated by the chaperone O-fut. Afterthe addition of these fucose groups, further extension ofthe carbohydrate chains is carried out by the Fringe family(Module 2: Table Notch signalling components). The de-gree of glycosylation can markedly influence the affinityof the Notch receptor for its ligands. The last modification,which occurs in the Golgi, is the cleavage of the moleculeat site 1 (S1) by the protease; the heterodimeric receptor isthen inserted into the membrane.
Trafficking of Notch through the endosomal compart-ment determines the amount of receptor in the membraneand this is an important aspect of the modulation of Notchsignalling (Module 2: Figure Notch modulation).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �105
Endoplasmic reticulum (ER) stresssignallingThe endoplasmic reticulum (ER) has sophisticated stresssignalling pathways that enable it to adapt to a wholehost of stress factors mainly concerned with the way inwhich proteins are synthesized and packaged. Mainten-ance of a constant level of Ca2+ within the lumen of theER is essential for the post-translational processing, fold-ing and export of proteins. This protein processing is car-ried out by a number of Ca2+-sensitive chaperones such as78 kDa glucose-regulatory protein (GRP78) [also knownas immunoglobulin heavy-chain-binding protein (BiP)],GRP94 (endoplasmin) and calnexin. Any decline in the lu-minal level of Ca2+ results in the accumulation of misfol-ded proteins and the activation of the ER stress signallingpathways (Module 2: Figure ER stress signalling):
1. Oligomerization and autophosphorylation of PKR(protein kinase R)-like ER kinase (PERK) sets off aphosphorylation cascade that culminates in the phos-phorylation and inactivation of the translation euka-ryotic initiation factor eIF-2α, resulting in protein syn-thesis being switched off.
2. Oligomerization and autophosphorylation of IRE1 ini-tiates one of the transcriptional signalling pathways re-sponsible for the up-regulation of the various chaper-ones.
3. Another of the transcriptional pathways dependsupon the activation of the ER membrane-boundactivating transcription factor 6 (ATF6), which isreleased from the ER to enter the nucleus, whereit interacts with the ER stress-response elementof the C/EBP (CCAAT/enhancer-binding protein)-homologous protein 10 (CHOP) gene.
4. The various chaperones are then expressed within theER, where they participate in protein folding.
5. One of the genes activated during the stress responseis CHOP, which acts as a transcription factor and cancontribute to apoptosis.
6. Caspase 12, which is associated with the ER membrane,is also activated and contributes to ER stress-inducedapoptosis.
7. An excessive accumulation of proteins within the ERresults in the activation of the transcription factornuclear factor κB (NF-κB), which acts to increase theproduction of interferons and cytokines, so contribut-ing to an inflammatory response.
These stress pathways are then responsible for switchingoff ongoing protein synthesis, for up-regulating the pro-duction of new chaperones, for inducing apoptosis and foractivating inflammatory responses. The degree to whichthese different responses are activated depends upon thenature of the stress. The fact that the ER can up-regulatechaperone levels results in the phenomenon of tolerance,whereby treatment of cells with low levels of stress stimulican make cells much more tolerant to subsequent stressfulstimuli.
Unfolded protein response (UPR)An accumulation of misfolded proteins induces an unfol-ded protein response (UPR), which switches off ongoingprotein synthesis and also activates various transcriptionalcascades that result in the up-regulation of many of thekey chaperones in an attempt to improve the defectiveprotein packaging machinery (Module 2: Figure ER stresssignalling). Activating transcription factor 6 (ATF6) is oneof the transcription factors activated by the UPR.
Endoplasmic reticulum (ER) overload response(EOR)An excessive build-up of proteins, as occurs during viral in-fections, switches on an endoplasmic reticulum (ER) over-load response (EOR) that acts through the nuclear factorκB (NF-κB) signalling cascade to stimulate the release ofinterferons and cytokines as part of an inflammatory re-sponse.
Activation of apoptosisEndoplasmic reticulum (ER) stress signalling pathwayscan also contribute to apoptosis (Module 11: Figure ap-optosis). For example, one of the UPR pathways dependsupon the release of the transcription factor activatingtranscription factor 6 (ATF6), which acts to switch onC/EBP (CCAAT/enhancer-binding protein)-homologousprotein 10 (CHOP). Another mechanism depends uponthe ER directly activating a subset of caspases duringER stress. A critical component is caspase 12, which isassociated with the ER membrane and is released byproteolytic cleavage following ER stress. Several mech-anisms have been proposed for this activation process.One suggestion is that the stress sensor molecule IRE1recruits tumour-necrosis-factor-receptor-associated factor(TRAF) which then binds to caspase 12, making it sens-itive to the Ca2+-responsive cysteine protease m-calpain.Another suggestion is that the hydrolysis of caspase 12 ismediated by caspase 7, which is recruited to the membraneduring ER stress. An interesting aspect of this mechanismis that glucose-regulated protein 78 kDa (GRP78) appearsto inhibit this activation process by forming a complexwith caspase 7 and caspase 12. Once caspase 12 is releasedinto the cytosol, it activates a specific cascade involvingcaspase 9 and caspase 3 in a cytochrome c-independentmanner.
Metabolic messengersThere are a number of cellular metabolites that function asmetabolic messengers to integrate the activities of cellularmetabolism and cell signalling (Module 2: Figure meta-bolic messengers). In this context, a metabolic messengeris defined fairly widely: it includes components that areeither a part of, or are derived from, cellular metabolism.Metabolism is regulated at many different levels. The mostdirect control depends upon feedback processes where cer-tain substrates or products function as positive or negativeregulators of the enzymes that synthesize or metabolizethem. They are not signalling mechanisms in the strictsense, but are based on relatively straightforward mass
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �106
Module 2: Figure ER stress signalling
Dysfunctionalchaperones
STRESS
Normal chaperones
Protein synthesis OFF
Protein synthesis ON
Interleukins Cytokines
e1F-2
e1F-2
PDI: GRP78: GRP94; Calreticulin etc
CHOP
ExportExportOligomerizationOligomerization Misfolded
protein
Excessprotein
Endoplasmic reticulum
ATF-6NF-kB
Transcription factors
Protein synthesisProtein synthesis
Protein foldingProtein folding
APOPTOSISINFLAMMATORY RESPONSES
1
2 3
4
5
6
IRE1
PERK
Caspase-12 7
P
PP
PP
Endoplasmic reticulum (ER) stress signalling pathways.An accumulation of misfolded proteins or an excessive accumulation of normal proteins activate a number of signalling pathways. Chaperones withinthe endoplasmic reticulum (ER) lumen are responsible for folding newly synthesized proteins into their tertiary structures prior to their export to theGolgi. A variety of stress factors, including a decline in the luminal level of Ca2+, results in dysfunctional chaperones and an accumulation of misfoldedproteins that can activate a number of signalling pathways.
Module 2: Figure metabolic messengers
Metabolicmessengers
cADPR
NAADP
NADNADP
SH
ADPR
ADP ribosyl cyclase
ATP
HCO 3
AMP
NADHNAD
Genetranscription
K
K
+Ca2+
Ca2+
Ca2+
Ca2+
P2XP2YTRPM2
+
++
+
+
ATP
ATP
AMP signalling pathway
Cyclic AMP signalling pathway
Calcium signalling pathway
PLC
InsPDAG 3
α γβ
Interaction between metabolic messengers and cell signalling pathways.A number of metabolic intermediates can interact with various cell signalling pathways. ATP plays a significant role through its ability to closeATP-sensitive K+ channels as occurs in insulin-secreting β-cells (Module 7: Figure β-cell signalling). ATP that is released from the cell functionsto activate ATP-sensitive P2X channels (Module 3: Figure P2X receptor structure). ATP may also play an important role in regulating the activity ofADP-ribosyl cyclase that produces the Ca2+-mobilizing messengers cyclic ADP-ribose (cADPR) and nicotinic acid–adenine dinucleotide phosphate(NAADP) (Module 2: Figure cADPR/NAADP function). AMP is an important activator of the AMP signalling pathway (Module 2: Figure AMPK control ofmetabolism). The breakdown product of cADPR is ADP-ribose (ADPR), which is an activator of the transient receptor potential melastatin 2 (TRPM2)channel. Bicarbonate (HCO3
−) is an activator of soluble adenylyl cyclase and thus contributes to the cyclic AMP signalling pathway (Module 2: Figurecyclic AMP signalling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �107
Module 2: Figure metabolic signalling
CELLULARRESPONSES
ROS/RNS
NAD / NADH+
NADP / NADPH+
1
2
3
4
5
6
7
89AMP/ATP
NAADPcADPR
HCO3
_
Metabolic messengers
Energy
ME
TAB
OLI
SM
SIG
NA
LLIN
G
PAT
HW
AYS
Stimuli Substrates Oxygen
Function of metabolic hormones in integrating the operation of cell metabolism and cell signalling pathways.The signalling cascade begins with the arrival of an agonist (1) that recruits specific signalling pathways to activate a cellular response (2). Metabolismuses oxygen and substrates (3) to provide the energy (4) that not only powers the cellular responses (5), but also drives signalling pathways (6). Thesignalling pathways can also regulate the supply of energy by direct effects on metabolism (7). Superimposed on these more direct interactions, thereare a variety of metabolic messengers that form a complex signalling network (8 and 9) that integrate the activity of both the signalling and metaboliccascades.
action reactions that ensure that metabolism proceeds in anorderly and regulated manner. However, some of the meta-bolic components can be considered to be metabolic mes-sengers because they activate or modulate clearly definedsignalling pathways. There are a number of such metabolicmessengers:
• Adenosine triphosphate (ATP)• Adenosine monophosphate (AMP)• Fatty acids• Bicarbonate (HCO3
−)• NAD+ signalling pathways
The AMP signalling pathway is of major importancewith regard to metabolic signalling systems. Componentsof various redox signalling systems, such as the nitric ox-ide signalling pathway, are also related to such metabolicsignalling pathways.
Signalling through metabolites is a highly integrated sys-tem because some of the signalling pathways activated bythe metabolic messengers can feed back to regulate meta-bolism (Module 2: Figure metabolic signalling). An ex-ample of how a change in metabolism can affect a signallingpathway has been described in cardiac myocytes, where theaddition of pyruvate to increase metabolism has a markedeffect on Ca2+ signalling (Module 2: Figure pyruvate andCa2+ signalling).
AMP signalling pathway
Adenosine monophosphate (AMP)Cells have an AMP signalling pathway that is activated byan increase in the AMP/ATP ratio, which results in theactivation of an AMP-activated protein kinase (AMPK).AMP thus functions as a second messenger, since it is re-sponsible for activating the signalling pathway. The AMPKthat responds to AMP has been referred to as the “fuelgauge” in that it responds to a decrease in the level ofATP. This signalling cascade is sensitive to many stim-uli, such as cell stress, oxidative damage, hypoxia andglucose deprivation. Once activated, AMPK induces anup-regulation of ATP-generating systems while simultan-eously down-regulating processes that consume energy.One of its important actions is to reduce protein synthesiswhen energy levels are low by regulating the activity ofthe target of rapamycin (TOR) (Module 9: Figure target ofrapamycin signalling). Another of its actions is to regulatethe transcription of genes that function in the metabol-ism of glucose, fatty acids and cholesterol. The AMPKsignalling pathway may also play a role in stimulating mi-tochondrial biosynthesis.
AMPK has also been implicated in cell growth control,where it functions to regulate the protein kinase TORthat controls protein synthesis (Module 9: Figure target ofrapamycin signalling).
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �108
Module 2: Figure pyruvate and Ca2+ signalling
The effect of pyruvate on Ca2+ signalling in cardiac myocytes.The traces represent the Ca2+ spikes that are generated when isolated cardiac myocytes are electrically stimulated at 0.5 Hz. Under control conditions(0′), there are constant amplitude transients resulting from the release from the internal store. The size of this store is indicated by the amplitude ofthe much larger transient when caffeine (10 mM) is added. The next series of transients were taken 1, 2 and 8 min after addition of 10 mM pyruvate,which caused an increase in both the resting level and the transient amplitudes. This increase in the amplitude of the Ca2+ transients may haveresulted from an increase in the content of the internal store, as indicated by the very large caffeine-induced transient apparent at the end of the 8-minsequence. The effect of pyruvate was reversible because the signalling system was back to its control values 4 min after washing out the pyruvate(W). A possible interpretation of this experiment is that pyruvate enhanced metabolism and the increase in ATP concentration may have enhanced theformation of cyclic ADP-ribose (cADPR) to stimulate the pump that transfers Ca2+ into the internal store (Module 2: Figure cADPR/NAADP function).Reproduced from Zima, A.V., Kockskamper, J., Meijia-Alvarez, R. and Blatter, L.A. (2003) Pyruvate modulates cardiac sarcoplasmic reticulum Ca2+
release in rats via mitochondria-dependent and -independent mechanisms. J. Physiol. 550:765–783, with permission from Blackwell Publishing; seeZima et al. 2003.
AMP-activated protein kinase (AMPK)AMP-activated protein kinase (AMPK) plays a critical rolein regulating the usage of fuels such as glucose and fattyacids. However, it can have additional functions such asthe regulation of insulin secretion by pancreatic β-cells,and may also play a role in controlling satiety centres inthe hypothalamus. AMPK is a trimeric protein (Module2: Figure AMPK structure) made up of multiple isoformsof a catalytic α subunit (α1 and α2) associated with β-subunits (β1 and β2) and γ-subunits (γ1–γ3). Cells expressdifferent combinations of these different isoforms. Theactivity of AMPK is regulated by both the intracellularlevel of AMP and by phosphorylation through an AMPKkinase known as LKB1.
AMPK functions as a pleiotropic regulator of cell meta-bolism (Module 2: Figure AMPK control of metabolism).It stimulates the translocation of the glucose transporter(GLUT4) to the plasma membrane, where it facilitates theentry of glucose in skeletal muscle and heart cells. AMPKcan influence the balance between glycolysis and gluco-neogenesis by stimulating the formation of fructose 2,6-bisphosphate (F-2,6-P2), which is a potent regulator ofglycolysis through its ability to activate 6-phosphofructo-1-kinase (PFK-1) and to inhibit fructose-1,6-bisphosphate1-phosphatase. The formation of F-2,6-P2 is regulated bytwo separate signalling pathways. AMPK promotes glyco-lysis by stimulating the phosphofructokinase (PFK-2) ofthe bifunctional enzyme, which has both kinase and phos-phatase activities. Cyclic AMP acting through the fructose-2,6-bisphosphate 2-phosphatase component lowers thelevel of F-2,6-P2, which reduces glycolysis and promotesgluconeogenesis.
AMPK can inhibit the activity of various transcrip-tion factors such as the sterol regulatory element-bindingprotein 1c (SREBp1c) and hepatocyte nuclear factor 4α
(HNF4α), which regulate lipogenic and glycolytic genesrespectively. AMPK phosphorylates TORC2 to preventit from acting as a cofactor to activate genes respons-ible for gluconeogenesis. Glycogen can inhibit the activityof AMPK by interacting with the glycogen-binding do-main (GBD) of the β subunit (Module 2: Figure AMPKstructure). Finally, AMPK plays an important role in cellgrowth control by reducing protein synthesis when energylevels are low by acting through TOR (Module 9: Figuretarget of rapamycin signalling).
In summary, AMPK switches the cell away from energy-requiring processes towards energy conservation. It doesthis by exerting rapid effects on processes such as glucoseentry and glycolysis, as well as longer-term effects, by reg-ulating the transcription of both glycolytic and lipogenichormones (Module 2: Figure AMPK control of metabol-ism). Many of its actions are carried out in muscle cells andliver. With regard to the latter, it can phosphorylate trans-ducer of regulated cyclic AMP response element-bindingprotein 2 (TORC2), which is then prevented from act-ing as a cofactor for the cyclic AMP response element-binding protein (CREB) transcription factor, which canactivate genes responsible for gluconeogenesis (Module 7:Figure liver cell signalling). However, AMPK can also reg-ulate the activity of other cells such as insulin-secreting β1cells (Module 7: Figure β-cell signalling) and may func-tion in O2-sensing by the glomus cells in the carotidbody (Module 10: Figure carotid body chemorecep-tion). The fact that AMPK plays such a central role in
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �109
regulating energy metabolism has important implicationsfor diabetes.
Glycogen storage disease in humans is caused by a muta-tion of the AMPK γ-subunit.
LKB1LKB1 is a serine/threonine protein kinase that functions toactivate AMP-activated protein kinase (AMPK) by phos-phorylating Thr-172 on the α-subunit (Module 2: FigureAMPK structure). The activation of AMPK is very de-pendent upon LKB1. Inactivation of the latter is respons-ible for Peutz-Jeghers syndrome. As such, LKB1 is con-sidered to be one of the tumour suppressors.
Adenosine triphosphate (ATP)ATP is an important metabolic messenger in that it hasa number of different actions both as an internal andan external signal (Module 2: Figure metabolic messen-gers). With regard to the former, one of its main func-tions is to regulate the activity of the ATP-sensitive K+
(KATP) channel, which is particularly important in reg-ulating the release of insulin (Module 7: Figure β-cellsignalling). It also plays a role in regulating the activ-ity of the ADP-ribosyl cyclase that generates the Ca2+-mobilizing messengers cyclic ADP-ribose (cADPR) andnicotinic acid–dinucleotide phosphate (NAADP) (Module2: Figure cADPR/NAADP function). ATP is also releasedfrom the cell and can activate P2X receptors that activateCa2+ entry or P2Y receptors that are coupled to phos-pholipase C (PLC) to generate inositol 1,4,5-trisphosphate(InsP3) and diacylglycerol (DAG) (Module 2: Figure meta-bolic messengers).
Bicarbonate (HCO3−)
The CO2 produced during cellular metabolism is rap-idly converted into bicarbonate (HCO3
−), which acts as amessenger to report the current state of metabolism. TheHCO3
− activates soluble adenylyl cyclase (Module 2: Fig-ure cyclic AMP signalling) to generate cyclic AMP that hasmany signalling functions, including an effect on cellularmetabolism through its activation of glycogen metabolism.
Fatty acidsFatty acids can be considered as metabolic messen-gers because they contribute to a number of meta-bolic control mechanisms. For example, they con-trol gluconeogenesis in liver cells by activating theperoxisome-proliferator-activated receptor α (PPARα)(Module 7: Figure liver cell signalling). Free fatty acidsare also responsible for activating the uncoupling pro-tein 1 (UCP1) that provides the proton leak duringnoradrenaline-induced heat production by brown fat cellmitochondria (Module 7: Figure brown fat cell).
The positive-feedback loop that free fatty acids (FFAs)exert on the process of lipogenesis (i.e. Step 7 in Module7: Figure metabolic energy network) may exacerbate theonset of obesity by enhancing fat storage.
NAD signalling pathwaysAs the name implies, the NAD signalling pathways en-compass signalling systems that depend on nicotinamide–adenine dinucleotide (NAD+) and nicotinamide–adeninedinucleotide phosphate (NADP+). These two meta-bolic cofactors exist in the cell as NAD+/NADH andNADP+/NADPH redox couples and have a number ofimportant functions both as messengers and as precursorsof other metabolic messengers.
NAD and NADP as metabolic messengers• NAD+ functions to regulate a number of cellular pro-
cesses including energy metabolism, gene transcription,DNA repair and perhaps ageing as well.
• NADH activates the C-terminal binding protein(CtBP), which is a transcriptional corepressor that func-tions during growth and development.
• NADH and NADPH regulate the transcription factorsClock/BMAL1 and NPAS2/BMAL1 that control geneexpression during the operation of the circadian clock.
• NADPH appears to regulate the ADP-ribosyl cy-clase that is responsible for producing both cyclicADP-ribose (cADPR) and nicotinic acid–adenine di-nucleotide phosphate (NAADP) (Module 2: FigurecADPR/NAADP function).
NAD and NADP as precursors of metabolicmessengers• NAD+ is a precursor of cyclic ADP-ribose (cADPR)
signalling pathway that functions in Ca2+ signalling.• Metabolism of cADPR generates ADPR, which regu-
lates transient receptor potential melastatin 2 (TRPM2),a member of the transient receptor potential (TRP) ionchannel family that controls the entry of external Ca2+
(Module 2: Figure cADPR/NAADP function).• NADP+ is a precursor of nicotinic acid–adenine dinuc-
leotide phosphate (NAADP) signalling pathway thatfunctions in Ca2+ signalling.
• NADPH is the substrate used to generate reactive oxy-gen species (ROS), which function as second messen-gers to regulate a number of cellular proteins (Module2: Figure plasma membrane ROS formation).
Sterol sensing and cholesterol biosynthesisThe level of cholesterol in cell membranes is regulated bya signalling system that can sense the level of sterols andthen relay information to the nucleus to adjust the tran-scription of the genes responsible for cholesterol biosyn-thesis. The signalling system is based on membrane-boundtranscription factors: the sterol regulatory element-bind-ing proteins (SREBPs), which are integral membrane pro-teins located in the endoplasmic reticulum (ER). They havetwo membrane-spanning domains with the free ends pro-jecting into the cytosol. The N-terminal region is the lat-ent transcriptional regulator, which is cleaved by a sterol-regulated system of proteases (Module 2: Figure sterolsensing). Once released into the cytosol, these transcrip-tion factors dimerize through a basic loop–helix–leucinezipper and enter the nucleus, where they bind to sterol
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �110
Module 2: Figure AMPK structure
α
α
α
αβ
βγ
γ
γ
γ
BD
BDGBD
GBD
C
AMP
AMP
C
P
P
Enzyme activation byAMP and protein phosphorylation by an AMPK kinase (LKB1)
Protein-Protein
LKB1+
+
I
I
Structure and function of AMP-activated protein kinase (AMPK).AMP-activated protein kinase (AMPK) is an αβγ heterotrimer. The β-subunit, which contains a C-terminal αγ-binding domain (αγBD), functions asa scaffold to organize the other two subunits. The α-subunit has an N-terminal catalytic domain (C), which is kept quiescent at rest by binding toan autoinhibitory domain (I). This α-subunit is activated by both AMP and by an AMPK kinase (AMPKK). The γ-subunit binds AMP and undergoesa conformation change that is transmitted to the α-subunit, causing the enzyme to open up so that its catalytic site can begin to phosphorylate itssubstrate proteins. Enzyme activity is also regulated by an AMPKK known as LKB1, which phosphorylates Thr-172. The β-subunit also contains aglycogen-binding domain (GBD), which enables AMPK to associate with glycogen that serves to inhibit the enzyme.
Module 2: Figure AMPK control of metabolism
G-6-P
G-6-P
F-6-P
F-1,6-P
F-2,6-P
Pyruvate
PFK-1
2
2 F-1,6-P2ase
Exocytosis
PFK-2
F-2,6-P ase2
Gluconeogenesis
Glycolysis Proteinsynthesis
+++
+
+
+
++
+
+
__
AMPK
_
_
_
SREBp1c HNF4α
Lipogenic and glycolytic genes
Glycogen
AMPLKB1
TORC
TORC
Glucose
Glut4 Cyclic AMP
PKA
Glucagon
TSC1/2
TOR
Nucleus
P
The pleiotropic action of AMP-activated protein kinase (AMPK) on cell metabolism.An increase in the level of AMP during metabolic stress activates AMP-activated protein kinase (AMPK), which then has a number of actions, asoutlined in the text.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �111
Module 2: Figure sterol sensing
Sterol signalling at the endoplasmic reticulum (ER) membrane.Sterols bind to the sterol regulatory element-binding proteins (SREBP) cleavage-activating protein (SCAP) resulting in the activation of site-1 protease(S1P), which then cleaves the SREBPs at a point on the luminal loop. A site-2 protease (S2P) then hydrolyses the N-terminal bHLH (basic helix–loop–helix) fragment to release the cytosolic portion, which then diffuses into the nucleus to initiate gene transcription. Reproduced from Brown, M.S.and Goldstein, J.L. (1999) A proteolytic pathway that controls the cholesterol content of membranes, cells, and blood. Proc. Natl. Acad. Sci. U.S.A.96:11041–11048. Copyright (1999) National Academy of Sciences, U.S.A.; see Brown and Goldstein 1999).
regulatory elements on the genes that code for the en-zymes of lipid biosynthesis (cholesterol, unsaturated fattyacids and triacylglycerols) and lipid-uptake mechanisms.The sterol sensor is a SREBP cleavage-activating protein(SCAP). When sterols build up in the membranes, theybind to SCAP to inhibit the proteolytic cleavage of theSREBPs. The ER thus plays a central role both in sensingthe level of sterols and then providing the output signalsthat make the necessary adjustments to lipid synthesis tomaintain a constant level of membrane cholesterol.
References
Monomeric G ProteinsCullen, P.J. and Lockyer, P.J. (2002) Integration of calcium and Ras
signalling. Nat. Rev. Mol. Cell Biol. 3:339–348.Manser, E. (2003) Cytoskeletal regulation: small G-protein–kinase
interactions. Handbook of Cell Signaling Vol. 1 (Bradshaw, R.A. andDennis, E.A., eds), pp. 499–503, Academic Press, San Diego.
Mitlin, N., Rossman, K.L. and Der, C.J. (2005) Signalling interplay in Rassuperfamily function. Curr. Biol. 15:R563–R574.
Rossman, K.L., Der, C.J. and Sondek, J. (2005) GEF means go: turningon Rho GTPases with guanine nucleotide-exchange factors. Nat. Rev.Mol. Cell. Biol. 6:167–180.
Somlyo, A.P. and Somlyo, A.V. (2003) Ca2+ sensitivity of smooth muscleand nonmuscle myosin II: modulated by G proteins, kinases, andmyosin phosphatase. Physiol. Rev. 83:1325–1358.
Tybulewicz, V.L.J. (2005) Vav-family proteins in T-cell signalling.Curr. Opin. Immunol. 17:267–274.
Welch, H.C.E., Coadwell, W.J., Stephens, L.R. and Hawkins, P.T. (2003)Phosphoinositide 3-kinase-dependent activation of Rac. FEBS Lett.546:93–97.
Cyclic AMP signalling pathwayCooper, D.M. (2003) Regulation and organization of adenylyl cyclases
and cAMP. Biochem. J. 375:517–529.Whorton, M.R. and Sunahara, R.K. (2003) Adenylyl cyclases. Handbook
of Cell Signaling Vol. 2 (Bradshaw, R.A. and Dennis, E.A., eds),pp. 419–426, Academic Press, San Diego.
G proteinsRal-GTPase: approaching their 15 minutes of fame (2003) Trends Cell
Biol. 13:419–425.
Ca2+ signallingBerridge, M.J., Bootman, M.D. and Roderick, H.L. (2003) Calcium
signalling: dynamics, homeostasis and remodelling. Nat. Rev. Mol.Cell Biol. 4:517–529.
Maravall, M., Mainen, Z.F., Sabatini, B.L. and Svoboda, K. (2000)Estimating intracellular calcium concentration and buffering withoutwavelength ratioing. Biophys. J. 78:2655–2667.
Cyclic ADP-ribose (cADPR)Galione, A. and White, A. (1994) Ca2+ release induced by cyclic
ADP-ribose. Trends Cell Biol. 4:431–436.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �112
Hua, S.-Y., Tokimasa, T., Takasawa, S., Furuya, S., Nohmi, M.,Okamoto, H. and Kuba, K. (1994) Cyclic ADP-ribose modulates Ca2+release channels for activation by physiological Ca2+ entry in bullfrogsympathetic neurons. Neuron 12:1073–1709.
Lee, H.C. (1997) Mechanisms of calcium signalling by cyclic ADP-riboseand NAADP. Physiol. Rev. 77:1133–1164.
Lukyanenko, V., Gyorke, I., Wiesner, T.F. and Gyorke, S. (2001)Potentiation of Ca2+ release by cADP-ribose in the heart is mediatedby enhanced SR Ca2+ uptake into the sarcoplasmic reticulum.Circ. Res. 89:614–622.
Nicotinic acid–adenine dinucleotide phosphate(NAADP) signallingLee, H.C. (2003) Ca2+ signalling: NAADP ascends as a new messenger.
Curr. Biol. 13:R186–R188.Patel, S., Churchill, G.C. and Galione, A. (2001) Coordination of Ca2+
signalling by NAADP. Trends Biochem. Sci. 26:482–489.
Phospholipase C (PLC)Delmas, P., Crest, M. and Brown, D.A. (2004) Functional organization of
PLC signalling microdomains in neurons. Trends Neurosci. 27:41–47.Katan, M. (1998) Families of phosphoinositide-specific phospholipase
C: structure and function. Biochim. Biophys. Acta 1436:5–17.Kelley, G.G., Reks, S.E., Ondrako, J.M. and Smrcka, A.V. (2001)
Phospholipase Cε: a novel Ras effector. EMBO J. 20:743–754.Rebecchi, M.J. and Pentyala, S.N. (2000) Structure, function, and
control of phosphoinositide-specific phospholipase C. Physiol. Rev.80:1291–1335.
Rhee, S.G. and Bae, Y.S. (1997) Regulation of phosphoinositide-specificphospholipase C isozymes. J. Biol. Chem. 272:15045–15048.
Singer, W.D., Brown, H.A. and Sternweis, P.C. (1997) Regulation ofeukaryotic phosphatidylinositol-specific phospholipase C andphospholipase D. Annu. Rev. Biochem. 66:475–509.
Nuclear phosphoinositide cycleIrvine, R.F. (2003) Nuclear lipid signalling. Nat. Rev. Mol. Cell Biol.
4:349–361.Martelli, A.M., Manzoli, L., Faenza, I., Bortul, R., Billi, A.M. and Cocco, L.
(2002) Nuclear inositol lipid signalling and its potential involvement inmalignant transformation. Biochim. Biophys. Acta 1603:11–17.
Phosphoinositide lipid signalling moleculesCzech, M.P. (2000) PIP2 and PIP3: complex roles at the cell surface. Cell
100:603–606.Yin, H.Y. and Janmey, P.A. (2003) Phosphoinositide regulation of the
actin cytoskeleton. Annu. Rev. Physiol. 65:761–789.Cockcroft, S. and De Matteis, M.A. (2001) Inositol lipids as spatial
regulators of membrane traffic. J. Membr. Biol. 180:187–194.Suh, B.-C. and Hille, B. (2005) Regulation of ion channels by
phosphatidylinositol 4,5-bisphosphate. Curr. Opin. Neurobiol.15:370–378.
McLaughlin, S., Wang, J., Gambhir, A. and Murray, D. (2002) PIP2 andproteins: interactions, organization, and information flow. Annu. Rev.Biophys. Biomol. Struct. 31:151–175.
Pendaries, C., Tronchere, H., Racaud-Sultan, C., Gaits-Iaconvani, F.,Coronas, S., Manenti, S., Gratacap, M.-P., Plantavid, M. and Payrastre,B. (2005) Emerging roles of phosphatidylinositol monophosphates incellular signalling and trafficking. Adv. Enzyme Regul. 45:201–214.
Takenawa, T. and Itoh, T. (2001) Phosphoinositides, key regulatorymolecules for regulation of actin cytoskeleton and membrane trafficfrom the plasma membrane. Biochim. Biophys. Acta 1533:190–206.
PtdIns 3-kinase signallingHawkins, P.T., Anderson, K.E., Davidson, K. and Stephens, L.R. (2006)
Signalling through Class I PI3Ks in mammalian cells. Biochem Soc.Trans. 34:647–662.
Leslie, N.R. and Downes, C.P. (2004) PTEN function: how normal cellscontrol it and tumour cells lose it. Biochem. J. 382:1–11.
Woodgett, J.R. (2005) Recent advances in the protein kinase Bsignalling pathway. Curr. Opin. Cell Biol. 17:150–157.
Wymann, M.P. and Marone, R. (2005) Phosphoinositide 3-kinase indisease: timing, location, and scaffolding. Curr. Opin. Cell Biol.17:141–149.
Multipurpose inositol polyphosphate signallingpathwayShears, S.B. (2004) How versatile are inositol phosphate kinases?
Biochem. J. 377:265–280.
Nitric oxide (NO)/cyclic GMP signalling pathwayBogdan, C. (2001) Nitric oxide and the regulation of gene expression.
Trends Cell Biol. 11:66–75.Moncada, S., Palmer, R.M.J. and Higgs, E.A. (1991) Nitric oxide:
physiology, pathophysiology, and pharmacology. Pharmacol. Rev.43:109–142.
Pacher, P., Beckman, J.S. and Liaudet, L. (2007) Nitric oxide andperoxynitrite in health and disease. Physiol. Rev. 87:315–424.
Shaul, P.W. (2002) Regulation of endothelial nitric oxide synthase:location, location, location. Annu. Rev. Physiol. 64:749–774.
Stamler, J.S. (1994) Redox signalling: nitrosylation and related targetinteractions of nitric oxide. Cell 78:931–936.
Cyclic GMP (cGMP) signalling pathwayFriebe, A. and Koesling, D. (2003) Regulation of nitric oxide-sensitive
guanylyl cyclase. Circ. Res. 93:96–105.Hofmann, F. (2005) The biology of cyclic GMP-dependent protein
kinases. J. Biol. Chem. 280:1–4.Hofmann, F., Ammendola, A. and Schlossmann, J. (2000) Rising behind
NO: cGMP-dependent protein kinase. J. Cell Sci. 113:1671–1676.Lucas, K.A., Pitari, G.M., Kazerounian, S., Ruiz-Stewart, I., Park, J.,
Schulz, S., Chepenik, K.P. and Waldman, S.A. (2000) Guanylylcyclases and signalling by cyclic GMP. Pharmacol. Rev. 52:375–413.
Reactive nitrogen species (RNS) signallingFoster, M.W., McMahon, T.J. and Stamler, J.S. (2003) S-nitrosylation in
health and disease. Trends Mol. Med. 9:160–168.Marshall, H.E., Hess, D.T. and Stamler, J.S. (2004) S-nitrosylation:
physiological regulation of NF-κB. Proc. Natl. Acad. Sci. U.S.A.101:8841–8842.
Redox signallingBokoch, G.M. and Knaus, U.G. (2003) NADPH oxidases: not just for
leukocytes anymore!. Trends Biochem. Sci. 28:502–508.Cheek, T.R., McGuinness, O.M., Vincent, C., Moreton, R.B., Berridge,
M.J. and Johnson, M.H. (1993) Fertilisation and thimerosal stimulatesimilar calcium spiking patterns in mouse oocytes but by separatemechanisms. Development 119:179–189.
Chiarugi, P. and Cirri, P. (2003) Redox regulation of tyrosinephosphatases during receptor tyrosine kinase signal transduction.Trends Biochem. Sci. 28:509–514.
Droge, W. (2002) Free radicals in the physiological control of cellfunction. Physiol. Rev. 82:47–95.
Lambeth, J.D. (2004) NOX enzymes and the biology of reactive oxygen.Nat. Rev. Immunol. 4:181–189.
Mikkelsen, R.B. and Wardman, P. (2003) Biological chemistry of reactiveoxygen and nitrogen and radiation-induced signal transductionmechanisms. Oncogene 22:5734–5754.
Rhee, S.G., Kang, S.W., Jeong, W., Chang, T.-S., Yang, K.-S. and Woo,H.A. (2005) Intracellular messenger function of hydrogen peroxide andits regulation by peroxyredoxins. Curr. Opin. Cell Biol. 17:183–189.
Singh, D.K., Kumar, D., Siddiqui, Z., Basu, S.K., Kumar, V. and Rao,K.V.S. (2005) The strength of receptor signaling is centrally controlledthrough a cooperative loop between Ca2+ and an oxidant signal. Cell121:281–293.
Mitogen-activated protein kinase (MAPK)signallingDavis, R.J. (2000) Signal transduction by the JNK group of MAP kinases.
Cell 103:239–252.Johnson, G.L. and Lapadat, R. (2002) Mitogen-activated protein kinase
pathways mediated by ERK, JNK, and p38 protein kinases. Science298:1911–1912.
Morrison, D.K. and Davis, R.J. (2003) Regulation of MAP kinasesignalling modules by scaffold proteins in mammals. Annu. Rev. CellDev. Biol. 19:91–118.
Pouyssegur, J. and Lenormand, P. (2003) Fidelity and spatio-temporalcontrol of MAP kinase (ERKs) signalling. Eur. J. Biochem.270:3291–3299.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge � Module 2 � Cell Signalling Pathways 2 �113
Wellbrock, C., Karasarides, M. and Marais, R. (2004) The Raf proteinstake centre stage. Nat. Rev. Mol. Cell Biol. 5:875–885.
Nuclear factor κB (NF-κB) signalling pathwayAdhikari, A., Xu, M. and Chen, Z.J. (2007) Ubiquitin-mediated activation
of TAK1 and IKK. Oncogene 26:3214–3226.Akira, S. (2003) Toll-like receptor signalling. J. Biol. Chem.
278:38105–38108.Akira, S. (2004) Toll-like receptor signalling. Nat. Rev. Immunol.
4:499–511.Hayden, M.S. and Gosh, S. (2008) Shared principles in NF-κB
signalling. Cell 132:344–362.Nakanishi, C. and Toi, M. (2005) Nuclear factor-κB inhibitors and
sensitizers to anticancer drugs. Nat. Rev. Cancer 5:297–309.Perkins, N.D. (2007) Integrating cell-signalling pathways with NF-κB and
IKK function. Nat. Rev. Mol. Cell Biol. 8:49–62.Viatour, P., Merville, M.-P., Bours, V. and Chariot, A. (2004)
Phosphorylation of NF-κB and IκB proteins: implications in cancerand inflammation. Trends Biochem. Sci. 30:43–52.
Westwick, J.K., Schwamborn, K. and Mercurio, F. (2003) NFκB: a keyintegrator of cell signalling. Handbook of Cell Signaling, Vol. 3(Bradshaw, R.A. and Dennis, E.A., eds), pp. 107–114, AcademicPress, San Diego.
Virus recognition and antiviral responsesMcCartney, S.A. and Colonna, M. (2009) Viral sensors: diversity in
pathogen recognition. Immunol. Rev. 227:87–94.Takeuchi, O. and Akira, S. (2009) Innnate immunity to virus infection.
Immunol. Rev. 227:75–86.
Phospholipase D (PLD) signalling pathwayLiscovitch, M., Czarny, M., Fiucci, G. and Tang, X. (2000) Phospholipase
D: molecular and cell biology of a novel gene family. Biochem. J.345:401–415.
Singer, W.D., Brown, H.A. and Sternweis, P.C. (1997) Regulation ofeukaryotic phosphatidylinositol-specific phospholipase C andphospholipase D. Annu. Rev. Biochem. 66:475–509.
Sphingomyelin signalling pathwayFuterman, A.H. and Hannun, Y.A. (2004) The complex life of simple
sphingolipids. EMBO Rep. 5:777–782.Hla, T., Lee, M.-J., Ancellin, N., Paik, J.H. and Kluk, M.J. (2001)
Lysophospholipids: receptor revelations. Science 294:1875–1878.
Hannun, Y.A. and Obeid, L.M. (2008) Principles of bioactive lipidsignalling: lessons from sphingolipids. Nat. Rev. Mol. Cell Biol.9:139–150.
Le Stunff, H., Peterson, C., Liu, H., Milstien, S. and Spiegel, S. (2002)Sphingosine-1-phosphate and lipid phosphohydrolases.Biochim. Biophys. Acta 1582:8–17.
Spiegel, S., English, D. and Milstien, S. (2002) Sphingosine-1-phosphatesignaling: providing cells with a sense of direction. Trends Cell Biol.12:236–242.
Janus kinase (JAK)/signal transducer andactivator of transcription (STAT) signallingpathwayChen, X., Vinkemeier, U., Zhao, Y., Jeruzalmi, D., Darnell, J.E. Jr and
Kuriyan, J. (1998) Crystal structure of a tyrosine phosphorylatedSTAT-1 dimer bound to DNA. Cell 93:827–839.
Smad signalling pathwayDerynck, R. and Zhang, Y.E. (2003) Smad-dependent and
Smad-independent pathways in TGF-β family signalling. Nature425:577–584.
Shi, Y. and Massague, J. (2003) Mechanisms of TGF-β signalling fromcell membrane to the nucleus. Cell 113:685–700.
ten Dijke, P. and Hill, C.S. (2004) New insights into TGF-β-Smadsignalling. Trends Biochem. Sci. 29:265–273.
Wnt signalling pathwayGiles, R.H., van Es, J.H. and Clevers, H. (2003) Caught up in a Wnt
storm: Wnt signalling and cancer. Biochim. Biophys. Acta 1653:1–24.Logan, C.Y. and Nusse, R. (2004) The Wnt signalling pathway in
development and disease. Annu. Rev. Cell Dev. Biol. 20:781–810.Moon, R.T., Kohn, A.D., De Ferrari, G.V. and Kaykas, A. (2004) Wnt and
β-catenin signalling: diseases and therapies. Nat. Rev. Genet.5:691–701.
Nelson, W.J. and Nusse, R. (2004) Convergence of Wnt, β-catenin, andcadherin pathways. Science 303:1483–1487.
Reya, T. and Clevers, H. (2005) Wnt signalling in stem cells and cancer.Nature 434:843–850.
Strutt, D. (2003) Frizzled signalling and cell polarization in Drosophilaand vertebrates. Development 130:4501–4513.
Veeman, M.T., Axelrod, J.D. and Moon, R.T. (2003) A second canon:functions and mechanisms of β-catenin-independent Wnt signalling.Dev. Cell 5:367–377.
Hedgehog signalling pathwayDaya-Grosjean, L. and Couve-Privat, S. (2005) Sonic hedgehog
signalling in basal cell carcinomas. Cancer Lett. 225:181–192.Hooper, J.E. and Scott, M.P. (2005) Communicating with hedgehogs.
Nat. Rev. Mol. Cell Biol. 6:306–317.Ingham, P.W. and Kim, H.R. (2005) Hedgehog signalling and the
specification of muscle cell identity in the zebrafish embryo.Exp. Cell Res. 306:336–342.
Lum, L. and Beachy, P.A. (2004) The hedgehog response network:sensors, switches, and routers. Science 304:1755–1759.
Stecca, B., Mas, C. and Altaba, A.R. (2005) Interference with HH-GLIsignalling inhibits prostate cancer. Trends Mol. Med. 11:199–203.
Notch signalling pathwayBray, S.J. (2006) Notch signalling: a simple pathway becomes complex.
Nat. Rev. Mol. Cell Biol. 7:678–689.
Endoplasmic reticulum (ER) stress signallingRutkowski, D.T. and Kaufman, R.J. (2004) A trip to the ER: coping with
stress. Trends Cell Biol. 14:20–28.
Metabolic messengersZima, A.V., Kockskamper, J., Meijia-Alvarez, R. and Blatter, L.A. (2003)
Pyruvate modulates cardiac sarcoplasmic reticulum Ca2+ release inrats via mitochondria-dependent and -independent mechanisms.J. Physiol. 550:765–783.
Adenosine monophosphate (AMP)Hardie, D.G. (2005) New roles for the LKB1→AMPK pathway.
Curr. Opin. Cell Biol. 17:167–173.Kahn, B.B., Alquier, T., Carling, D. and Hardie, D.G. (2005)
AMP-activated protein kinase: ancient energy gauge provides clues tomodern understanding of metabolism. Cell Metab. 1:15–25.
Rutter, G.A., da Silva Xavier, G. and Leclerc, I. (2003) Roles of5’-AMP-activated protein kinase (AMPK) in mammalian glucosehomoeostasis. Biochem. J. 375:1–16.
Sterol sensing and the regulation of cholesterolbiosynthesisBrown, M. S. and Goldstein, J.L. (1999) A proteolytic pathway that
controls the cholesterol content of membranes, cells, and blood.Proc. Natl. Acad. Sci. U.S.A. 96:11041–11048.
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge 2 �114
Index of cited figures and tables
Figures Page no.
Module 1: Figure cytokines 1.26Module 1: Figure Eph receptor signalling 1.24Module 1: Figure homologous desensitization 1.55Module 1: Figure non-receptor tyrosine kinases 1.39Module 1: Figure PDGFR activation 1.23Module 1: Figure signal transmission mechanisms 1.37Module 1: Figure stimuli for cyclic AMP signalling 1.5Module 1: Figure stimuli for enzyme-linked receptors 1.20Module 1: Figure type I cytokine receptors 1.27Module 1: Figure tyrosine kinase-linked receptors 1.21Module 1: Figure ubiquitin–proteasome system 1.49Module 2: Figure adenylyl cyclase structure 2.4Module 2: Figure AMPK control of metabolism 2.110Module 2: Figure AMPK structure 2.110Module 2: Figure basic Ca2+ signalling mechanism 2.17Module 2: Figure Ca2+ modules 2.24Module 2: Figure Ca2+ signalling dynamics 2.18Module 2: Figure Ca2+ signalling toolkit 2.23Module 2: Figure Ca2+ transient mechanisms 2.26Module 2: Figure Ca2+-induced Ca2+ release 2.25Module 2: Figure cADPR action in heart cells 2.31Module 2: Figure cADPR action in neurons 2.32Module 2: Figure cADPR metabolism 2.29Module 2: Figure cADPR/NAADP function 2.30Module 2: Figure Cdc42 signalling 2.15Module 2: Figure cell signalling pathways 2.2Module 2: Figure cell-specific Ca2+ signalsomes 2.22Module 2: Figure cyclic AMP signalling 2.5Module 2: Figure eNOS activation 2.58Module 2: Figure ER stress signalling 2.106Module 2: Figure ERK signalling 2.75Module 2: Figure G protein binary switching 2.8Module 2: Figure GSH/GSSG couple 2.65Module 2: Figure H2O2 metabolism 2.66Module 2: Figure Hedgehog signalling pathway 2.100Module 2: Figure heterotrimeric G protein signalling 2.10Module 2: Figure inositol phosphate metabolism 2.41Module 2: Figure InsP3 and DAG formation 2.42Module 2: Figure InsP3/Ca2+ signalling functions 2.42Module 2: Figure InsP3/DAG recycling 2.43Module 2: Figure insulin receptor 2.53Module 2: Figure JAK and STAT structure 2.88Module 2: Figure JAK/STAT function 2.91Module 2: Figure JAK/STAT heterogeneity 2.90Module 2: Figure JNK signalling 2.76Module 2: Figure MAPK signalling 2.75Module 2: Figure metabolic messengers 2.106Module 2: Figure metabolic signalling 2.107Module 2: Figure NAADP structure 2.32Module 2: Figure NF-κB activation 2.80Module 2: Figure NF-κB, IκB and IKK structure 2.78Module 2: Figure NO and cyclic GMP signalling 2.56Module 2: Figure NO synthase mechanism 2.57Module 2: Figure Notch modulation 2.104Module 2: Figure Notch signalling 2.101
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge 2 �115
Figures Page no.
Module 2: Figure peroxiredoxin catalytic cycles 2.66Module 2: Figure phosphoinositide metabolism 2.34Module 2: Figure phosphoinositide signalling systems 2.36Module 2: Figure PI3-K family 2.35Module 2: Figure PKC structure and activation 2.47Module 2: Figure plasma membrane ROS formation 2.64Module 2: Figure PLC structure and function 2.44Module 2: Figure PLD isoforms 2.83Module 2: Figure PLD signalling 2.83Module 2: Figure protein kinase A (PKA) 2.6Module 2: Figure PtdIns 3-kinase signalling 2.51Module 2: Figure PtdIns structure 2.34Module 2: Figure PtdIns4,5P2 regulation of K+ channels 2.50Module 2: Figure PtdIns4,5P2 regulation of TRP channels 2.50Module 2: Figure PtdIns4,5P2 signalling 2.48Module 2: Figure pyruvate and Ca2+ signalling 2.108Module 2: Figure Rac signalling 2.12Module 2: Figure Ras signalling 2.11Module 2: Figure reactive oxygen species (ROS) 2.63Module 2: Figure recovery of protein oxidation 2.69Module 2: Figure reversible and irreversible ROS oxidations 2.67Module 2: Figure Rho signalling 2.13Module 2: Figure Rho-regulated kinases 2.16Module 2: Figure ROS effects on Ca2+ signalling 2.72Module 2: Figure ROS formation and action 2.68Module 2: Figure ROS microdomains 2.74Module 2: Figure sites of ROS formation 2.64Module 2: Figure Smad domain structure 2.93Module 2: Figure Smad signalling 2.95Module 2: Figure sphingolipid metabolism 2.85Module 2: Figure sphingomyelin signalling 2.85Module 2: Figure STAT1/DNA complex 2.89Module 2: Figure sterol sensing 2.111Module 2: Figure summary of redox signalling 2.60Module 2: Figure temporal aspects 2.28Module 2: Figure TGF-βR activation 2.94Module 2: Figure thimerosal-induced Ca2+ signalling 2.70Module 2: Figure Toll receptor signalling 2.80Module 2: Figure viral recognition 2.82Module 2: Figure Wnt canonical pathway 2.98Module 2: Figure Wnt signalling pathways 2.96Module 2: Figure phosphoinositide metabolism 2.34Module 2: Figure Rho-regulated kinases 2.16Module 3: Figure AMPA receptor phosphorylation 3.17Module 3: Figure Ca2+ entry mechanisms 3.12Module 3: Figure CaV1.1 L-type channel 3.6Module 3: Figure CaV1.2 L-type channel 3.7Module 3: Figure CaV2 channel family 3.9Module 3: Figure CFTR channel 3.52Module 3: Figure cyclic nucleotide-gated channels 3.20Module 3: Figure InsP3R activation 3.63Module 3: Figure InsP3R structure 3.62Module 3: Figure P2X receptor structure 3.19Module 3: Figure polycystin domain structure 3.33Module 3: Figure ryanodine receptor structure 3.59Module 3: Figure STIM-induced Ca2+ entry 3.26Module 3: Figure TRP channel phylogeny 3.26Module 4: Figure actin remodelling 4.54
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge 2 �116
Figures Page no.
Module 4: Figure annexin structure 4.5Module 4: Figure Ca2+-induced membrane fusion 4.18Module 4: Figure calcineurin 4.14Module 4: Figure CREB activation 4.41Module 4: Figure ETS activation 4.43Module 4: Figure FOXO control mechanisms 4.42Module 4: Figure NFAT activation 4.39Module 4: Figure p53 domains 4.29Module 4: Figure phagosome maturation 4.20Module 4: Figure S100 phylogenetic tree 4.7Module 4: Figure SRF and AP-1 4.46Module 4: Figure transcription factor activation 4.23Module 5: Figure Ca2+ uptake and extrusion 5.15Module 5: Figure ER/mitochondrial shuttle 5.25Module 5: Figure mitochondrial Ca2+ signalling 5.23Module 5: Figure mitochondrial motility 5.28Module 5: Figure phospholamban mode of action 5.21Module 5: Figure PP2A holoenzyme 5.9Module 6: Figure caveolae molecular organization 6.15Module 6: Figure circadian clock molecular mechanism 6.42Module 6: Figure integrin signalling 6.21Module 6: Figure IRS domain structure 6.9Module 6: Figure modular lipid-binding domains 6.5Module 6: Figure signalling hierarchies 6.2Module 6: Figure vinculin function 6.22Module 7: Figure α-cell signalling 7.126Module 7: Figure β-cell signalling 7.125Module 7: Figure brown fat cell 7.129Module 7: Figure collecting duct function 7.103Module 7: Figure control of food intake 7.6Module 7: Figure control of pancreatic secretion 7.113Module 7: Figure endothelial cell contraction 7.22Module 7: Figure glomerulosa cell signalling 7.120Module 7: Figure glycogenolysis and gluconeogenesis 7.115Module 7: Figure HCl secretion 7.91Module 7: Figure hepatic stellate cell 7.118Module 7: Figure intestinal Ca2+ reabsorption 7.77Module 7: Figure intestinal secretion 7.95Module 7: Figure kidney Ca2+ reabsorption 7.104Module 7: Figure L cell 7.94Module 7: Figure lipolysis and lipogenesis 7.128Module 7: Figure liver cell signalling 7.116Module 7: Figure mesangial cell 7.106Module 7: Figure metabolic energy network 7.5Module 7: Figure osteoclast podosome 7.81Module 7: Figure skeletal muscle E-C coupling 7.25Module 7: Figure smooth muscle cell cGMP signalling 7.43Module 7: Figure smooth muscle cell E-C coupling 7.42Module 8: Figure bone cell differentiation 8.38Module 8: Figure brown fat cell differentiation 8.48Module 8: Figure dorsoventral specification 8.18Module 8: Figure epidermal stem cell 8.32Module 8: Figure HSC regulation 8.35Module 8: Figure mammalian fertilization 8.5Module 8: Figure osteoclast differentiation 8.41Module 8: Figure Satellite cell activation 8.29Module 8: Figure signalsome expression 8.26
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge 2 �117
Figures Page no.
Module 8: Figure stem cell function 8.27Module 8: Figure white fat cell differentiation 8.47Module 9: Figure angiogenesis signalling 9.37Module 9: Figure cell cycle signalling mechanisms 9.7Module 9: Figure cytokinesis 9.15Module 9: Figure G1 checkpoint signalling 9.18Module 9: Figure growth factor signalling 9.22Module 9: Figure immunological synapse structure 9.28Module 9: Figure T cell signalling map 9.31Module 9: Figure target of rapamycin signalling 9.24Module 9: Figure TCR signalling 9.32Module 9: Figure VEGF-induced proliferation 9.38Module 10: Figure Ca2+-induced synaptic plasticity 10.36Module 10: Figure carotid body chemoreception 10.80Module 10: Figure gonadotroph regulation 10.55Module 10: Figure lactotroph regulation 10.57Module 10: Figure medium spiny neuron signalling 10.39Module 10: Figure neuronal gene transcription 10.40Module 10: Figure nociception 10.77Module 10: Figure phototransduction overview 10.67Module 10: Figure phototransduction 10.67Module 10: Figure postsynaptic density 10.11Module 11: Figure apoptosis 11.20Module 11: Figure autophagy 11.19Module 11: Figure FcεRI mast cell signalling 11.9Module 11: Figure formation and action of PAMPs 11.11Module 11: Figure inflammation 11.2Module 11: Figure macrophage signalling 11.12Module 11: Figure mast cell inhibitory signalling 11.10Module 11: Figure mast cell signalling 11.7Module 11: Figure neutrophil chemotactic signalling 11.15Module 11: Figure neutrophil chemotaxis 11.14Module 11: Figure platelet activation 11.6Module 12: Figure amyloid cascade hypothesis 12.6Module 12: Figure amyloid plaques and tangles 12.10Module 12: Figure astrocyte-induced neuronal death 12.11Module 12: Figure colon cancer 12.45Module 12: Figure hypertrophy signalling mechanisms 12.22Module 12: Figure polycystins and polycystic kidney disease 12.57
Tables Page no.
Module 1: Table G protein-coupled receptors 1.32Module 2: Table adenylyl cyclases 2.4Module 2: Table Ca2+ signalling toolkit 2.19Module 2: Table Hedgehog signalling toolkit 2.100Module 2: Table heterotrimeric G proteins 2.9Module 2: Table MAPK signalling toolkit 2.73Module 2: Table monomeric G protein toolkit 2.14Module 2: Table NF-κB signalling toolkit 2.77Module 2: Table Notch signalling components 2.102Module 2: Table redox signalling components 2.62Module 2: Table Smad signalling toolkit 2.92Module 3: Table ABC transporters 3.51Module 3: Table properties of Ca2+-sensitive K+ channels 3.42Module 3: Table receptor-operated channel toolkit 3.13Module 3: Table VOC classification 3.2
C©2009 Portland Press Limited www.cellsignallingbiology.org

Cell Signalling Biology Michael J. Berridge 2 �118
Tables Page no.
Module 3: Table voltage-dependent K+ channels 3.37Module 5: Table Ca2+ pumping toolkit 5.17Module 5: Table PDE family properties 5.11Module 5: Table PP1 regulatory, targeting and inhibitory subunits and proteins 5.6
C©2009 Portland Press Limited www.cellsignallingbiology.org